лаборатория докембрийских организмов ПИН РАН
лаборатория докембрийских организмов ПИН РАН
Гемологический центр, геологический ф-т МГУ
ЗАГАДКА ЭДИАКАРСКИХ ОРГАНИЗМОВ
В 1947 г. вышла первая статья R.C.Sprigg о находках в кварцитах Паунд в районе хребта Флиндерс близ Эдиакары, штат Южная Австралия, необычных ископаемых остатков организмов, позднее получивших название «Эдиакарская фауна». Это полувековой давности открытие имело особое значение для наук о Земле благодаря тому, что находки происходили из заведомо докембрийских отложений. В те годы докембрий считался палеонтологически немым, так как был охарактеризован лишь плохо изученными и мало понятными тогда строматолитами и сомнительными находками. Благодаря открытию R.C.Sprigg было обращено серьезное внимание и понята большая важность аналогичных ископаемых, известных еще до Второй мировой войны, в том числе и из юго-западной части Русской платформы. Окаменелости из Эдиакары стимулировали широкие поиски подобных ископаемых по всему миру, а эдиакарская фауна позволила Б.С.Соколову в 1985 г. обосновать выделение новой геологической системы в докембрии – вендской системы.
Главной особенностью эдиакарских окаменелостей является то, что это не скелеты – раковины, домики, панцири, кости, хитиновые, целлюлозные или пектиновые покровы или отдельные внутренние или внешние структуры, либо следы жизнедеятельности в виде измененных текстур осадка, то есть все то, что было известно из фанерозойской ископаемой летописи. Эдиакарские окаменелости – это отпечатки и слепки на поверхностях напластования и внутри слоев осадочных пород мягкотелых существ, не имевших не только твердых скелетов, но и каких-либо способных фоссилизироваться органических покровов или внутренних структур. Но что это были за существа? Как могли сохраниться остатки мягкотелых организмов в плотных зернистых породах, часто в кварцитах? Как могли образоваться высокорельефные отпечатки и даже трехмерные слепки, и почему они не были деформированы при литификации осадка? Какой образ жизни вели эти существа и в каких условиях они обитали? Почему эдиакарские организмы существовали в геологической истории короткое время и никак не связаны преемственностью с последующей раннекембрийской фауной? До самого последнего времени разгадать эти загадки эдиакарской фауны не удавалось, либо ответы были неубедительными. Начавшиеся в последние годы детальные исследования эдиакарских и других ископаемых остатков из разрезов верхнего венда Архангельской обл. (север России) проливают свет на этих загадочных организмов и позволяют дать обоснованные ответы.
ЖИВОТНЫЕ ИЛИ РАСТЕНИЯ
Непосредственно по самим остаткам эдиакарских организмов невозможно судить о том, каково было строение клеток, из-за чего нельзя и корректно ответить на вопрос: были ли эти организмы прокариотами или эвкариотами? Однако размеры остатков (от долей см до нескольких десятков см) позволяют достаточно уверенно предполагать, что они принадлежали эвкариотам. Но были ли это водоросли или животные? Единственным ключом к решению этой проблемы может быть только анализ сохранности. Одновременно с эдиакарскими организмами в тех же самых или экологически близких условиях в позднем венде (540-570 млн. лет тому назад) в бассейнах, раскинувшихся на территории Русской платформы, обитали разнообразные нитчатые серные бактерии, актиномицеты, цианобактерии, коккоидные и нитчатые эвкариотные водоросли, хитридиевые водные грибы и макроскопические лентовидные водоросли, сохранившиеся в тех же самых (даже на тех же поверхностях напластования, где найдены отпечатки эдиакарских организмов) или литологически близких породах в виде органикостенных микрофоссилий или фитолейм. Уникальная неизмененность пород и отсутствие выветривания в обнажениях Архангельской обл. и скважинах Московской и Мезенской синеклиз снимает важнейший аргумент, широко использовавшийся ранее для объяснения почему не сохранились органические структуры у эдиакарских организмов – органические вещества на остатках отсутствуют из-за того, что они либо были разрушены в результате диагенеза и постдиагенетических процессов, либо в результате выветривания. Столь широкий спектр сохранности остатков разнообразных бактерий, водорослей и грибов в верхнем венде Русской платформы позволяет уверенно говорить о том, что клетки эдиакарских организмов должны были иметь иное строение. Эти же данные опровергают представления G.J.Retallack об эдиакарских организмах как лишайниках и грибах с плотными тканями, так как подобные ткани, как мы видим из приведенных выше примеров, должны были бы сохраняться в ископаемом состоянии. Клеточные структуры животных организмов крайне редко сохраняются в ископаемой летописи кроме как у замороженных трупов или остатков в янтаре. Поэтому можно предполагать, что эдиакарские организмы были именно животными, а не растениями. При этом масса тела этих организмов была невелика, так как их остатки очень слабо пиритизированы, тогда как в тех же условиях широко распространены остатки нитчатых водорослей рода Striatella (их нитчатый таллом был шириной около 0,1 мм и длиной более 2 или даже 4 см), пиритизированные в результате деятельности серных бактерий – деструкторов органики.
КАК ОБРАЗОВЫВАЛИСЬ ОТПЕЧАТКИ И СЛЕПКИ
За полувековую историю изучения эдиакарской фауны было предложено много объяснений способа образования отпечатков и слепков мягкотелых организмов. Были проведены эксперименты по моделированию процессов захоронения мягкотелых организмов – главным образом медуз, так как первоначально многие из эдиакарских окаменелостей трактовались именно как остатки планктонных медуз. В экспериментах отпечатки были получены с применением разных материалов: A.G.Nathorst в 1881 г. использовал гипс, W.Schafer в 1941 г., R.D.Norris в 1989 г. и D.L.Bruton в 1991 г. применяли пески с разной сортировкой зерен, гранулометрией и содержанием глинистого матрикса, D.L.Bruton использовал также латекс. Однако искусственность получающихся результатов вступала в очевидное противоречие с фактическим материалом. Это вынуждало исследователей предполагать, что в действительности имел место иной процесс образования эдиакарских окаменелостей. H.D.Pflug полагал, что могло происходить посмертное окремнение остатков организма. R.J.F.Jenkins считал, что слепки образовывались в результате заполнения осадком полости, оставшейся от разложившегося внутри осадка тела, а форма полости при этом сохранялась за счет образования ореола минеральных веществ, пропитывавших окружающий осадок. A.Seilacher предположил, что тела одних организмов могли располагаться внутри осадка и рости одновременно с седиментацией, а для объяснения образования отпечатков на поверхностях напластования он предложил процесс образования «посмертных масок»: нижняя часть засыпанного осадком тела разлагалась в результате жизнедеятельности бактерий, в эту полость снизу проникал осадок, тогда как более прочные структуры, располагавшиеся в спинной части тела, некоторое время сдерживали вышележащий осадок. Но лишь проведенное Д.В.Гражданкиным изучение сохранности остатков нового рода и вида Ventogyrus chistyakovi Ivantsov et Grazhdankin, 1997 и обстановок осадконакопления верхневендских пород Архангельской обл. прояснило как были образованы отпечатки и слепки эдиакарских организмов группы Petalonamae.
Остатки V. chistyakovi были найдены в среднем течении р. Онеги на юго-восточном окончании хребта Ветреный Пояс на западе Архангельской обл. Обнажения сложены глинисто-терригенными породами, среди которых доминирует глинисто-алевритистая толща, демонстрирующая характерное для венда тонкое ритмичное строение. В толще переслаивания алевролитов и глин присутствуют турбидитоподобные прослои, слепки промоин и эрозионных котлов, выполненные мелкозернистыми песчаниками, содержащими по напластованию плоскую глиняную гальку. В толще переслаивания алевролитов и глин широко распространена бугорчатая слоистость. Остатки V. chistyakovi встречены только в слепках эрозионных котлов, но не во всех. Слепки котлов – это текстуры, образовавшиеся в результате многоэтапного последовательного заполнения округлых эрозионных углублений (диаметром 0,6-0,8 м и глубиной 0,4 м) на дне. Борта слепков гладкие, с одной стороны относительно крутые, с другой – положе. Слепки промоин и котлов, выполненные мелкозернистыми песчаниками и алевролитами, являются характерными текстурами среди группы штормовых отложений, или темпеститов. Считается, что промоины и котлы возникают во время штормов и образуются сильными придонными течениями, они указывают на высокую гидродинамическую активность обстановок осадконакопления и на комбинированный эффект горизонтальных и вертикальных завихрений среди однонаправленных придонных потоков. Упомянутая выше бугорчатая слоистость в толще переслаивания также относится к текстурам группы темпеститов, она служит хорошим индикатором глубины проникновения штормовых волн и взмучивания осадка.
 |
Текстуры темпеститов (бугорчатая слоистость, слепки промоин и эрозионных котлов) в фациях отложений турбидных потоков характерны для обстановок шельфов с волновым режимом осадконакопления. Толща переслаивания алевро-глинистых пород в таком случае отвечает условиям дистальных темпеститов: взмученный волнами тонкий и мелкозернистый материал переносился турбидными потоками в обстановки ниже глубины взмучивания осадка. Замеры направлений простирания слепков промоин, падения косой слоистости и ориентировки гребней знаков ряби показывают, что перенос осадков осуществлялся в юго-восточном направлении посредством штормовых потоков.
Шторма были, вероятно, главным фактором переноса и накопления песчаных, алевритовых и глинистых отложений на этом участке палеобассейна. Штормовые волны взмучивали большие объемы песка, а генерированные штормами потоки переносили суспензию на большие расстояния и в более глубоководные обстановки, ниже глубины взмучивания осадка волнами. При затихании штормов происходило осаждение осадка в виде покровов на значительной площади. На дне на глубине взмучивания осадка штормовыми волнами формировались бугры, а в области локализации штормовых потоков образовывались следы крупных размывов – промоины и эрозионные котлы.
Захоронение организмов происходило в результате частичного заполнения котлов слоем песка, принесенного во взвешенном состоянии потоком в период возрастания штормовой активности. Быстрое осаждение материала из взвеси без дальнейшего перемещения его по дну способствовало захоронению организмов in situ, часто в прижизненном положении. Об этом свидетельствуют, помимо данных седиментологии, следующие признаки: на одной поверхности напластования встречаются формы разных размеров, то есть не происходила сортировка остатков по размерам; все остатки залегают выпуклой поверхностью вниз, нет перевернутых остатков или следов их волочения по дну; длинные оси остатков ориентированы всегда в разные стороны. Остатки V. chistyakovi захоранивались группами, такие захоронения встречены в нескольких следующих друг за другом прослоях. Это позволяет предположить, что после затихания штормов новообразованный субстрат на дне котла заселялся новой популяцией V. chistyakovi. Все это дает основания считать, что популяции V. chistyakovi обитали на дне вдоль долговременных придонных токов, которые могли поставлять на глубины и в котлы кислород и питательные вещества. Именно событийное осадконакопление – быстрое осаждение алевро-песчаного материала из взвеси без дальнейшего перемещения его по дну – было главным условием образования отпечатков и слепков V. chistyakovi, при этом отпечатки и слепки отражают строение верхней поверхности тела.
КАК ВЫГЛЯДЕЛ VENTOGYRUS CHISTYAKOVI
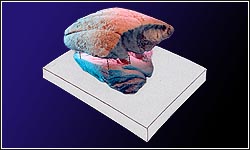
Ventogyrus chistyakovi имел тело, слегка напоминающее лодку. Длина тела от кормовой до носовой части была почти в два раза больше его ширины. Носовая и кормовая части тела были несколько приподняты, тогда как средняя часть тела, примерно равная его ширине, была горизонтальной.
 |
На верхней стороне тела располагалась система вертикальных гребней или перегородок, делившая ее на камеры. По середине проходила зигзаговидная перегородка, названная срединной. Один конец срединной перегородки соединялся с носовой частью тела, противоположный конец соединялся с кормовыми перегородками, отделявшими большую треугольную камеру, которая и формировала корму. Внутри кормовой камеры перегородки отсутствовали.
Срединная перегородка разделяла верхнюю поверхность тела на две половинки. От срединной перегородки и от кормовых перегородок почти перпендикулярно к ним, слегка отклоняясь в сторону носа, в обе стороны отходили перегородки разной длины. Среди них могут быть выделены разделительные перегородки, которые достигали боковых краев тела (бортов) и делили половинки на камеры, и дополнительные перегородки, не достигавшие бортов. Различаются дополнительные перегородки нескольких (до 5) порядков. Наиболее длинные дополнительные перегородки первого порядка имели длину не более 1/3 длины разделительных и располагались приблизительно посередине между ними. Дополнительные перегородки второго и следующих порядков, были еще короче и располагались также приблизительно посередине между разделительными и дополнительными перегородками первого и, соответственно, следующих порядков.
 |
Срединная перегородка была наиболее высокой в кормовой части в месте стыковки с кормовыми перегородками и достигала размеров, равных ширине тела или даже немного больше. Ее высота постепенно сокращалась в сторону носа. Кормовые перегородки были равны по высоте срединной в месте их стыковки, а затем из высота плавно сокращалась до нуля к краям тела. Высота разделительных перегородок в месте их отхождения от срединной была равна высоте последней, а к бортам плавно снижалась до нуля. Дополнительные перегородки были не только короче, но и, соответственно, ниже разделительных, их высота также сокращалась до нуля от срединной перегородки к их концам. Таким образом разделительные и дополнительные перегородки имели треугольную форму. Все перегородки были осложнены складочками.
В расположении разделительных и дополнительных перегородок наблюдается симметрия зеркально-скользящего отражения: перегородки двух половинок смещены на половину периода относительно плоскости, проходящей вдоль срединной перегородки. Именно по бороздам, отражающим на слепке перегородки, слепок может быть разобран на слепки камер, как на составные части. На обращенных к перегородкам сторонам слепков наблюдаются отпечатки каких-то структур тела, а на тонком прослое песчаника (иногда толщиной в одно-два зерна), подстилающим слепок, иногда наблюдаются слепки ветвящихся каналов и отпечаток морщинистой поверхности нижней стороны тела. Первоначально казалось, что тело имело также высокие (равные ширине тела) вертикально стоящие борта, однако последующее изучение не подтвердило это предположение.
 |
Столь необычное строение V. chistyakovi: полное отсутствие на отпечатках и слепках каких-либо следов рта, ротовых придатков и кишечника, форма тела, полностью отличная от формы тела известных беспозвоночных, условия обитания на дне в зонах сильных течений с периодической событийной седиментацией, – все это заставило обратить внимание на других, типичных, представителей группы Petalonamae, изученных ранее не столь детально и иначе реконструированных. С этой целью Д.В.Гражданкин переизучил вид Charnia masoni, Д.В.Гражданкин и А.Ю.Иванцов – Pteridinium nenoxa из богатейшей коллекции эдиакарских ископаемых, хранящейся в Лаборатории докембрийских организмов Палеонтологического института Российской Академии наук. Выяснилось, что и эти существа неподвижно лежали на поверхности дна и имели близкий план строения, а верхняя поверхность их тел была сложно расчленена гребнями, или перегородками. На отпечатках и слепках, принадлежащих этим видам, также не обнаружено никаких органов для сбора пищи и пищеварительной системы.
 |
КАКОЙ ОБРАЗ ЖИЗНИ ВЕЛИ ЭДИАКАРСКИЕ ОРГАНИЗМЫ
Ряд важных особенностей строения загадочных вендских мягкотелых организмов явно противоречит традиционным интерпретациям их как представителей кишечнополостных, разных групп червей, артропод, моллюсков и др. типов беспозвоночных. Такими особенностями являются: 1) отсутствие у большинства форм признаков рта, ротовых придатков, структур для сбора пищи; 2) отсутствие пищеварительной системы, тогда как у засыпанных осадком форм кишечник за счет его заполнения недопереваренной пищей (оболочки планктонных и клеточные структуры и оболочки бентосных водорослей и цианобактерий, а также мелкодисперсный органический детрит, то есть такая органика, которая хорошо фосссилизируется, сапропелизируется, или пиритизируется) должен четко выделяться на отпечатке или слепке; 3) отсутствие каких-либо устойчивых и способных мумифицироваться (то есть, по крайней мере, кутикулоподобных) покровных структур, не говоря уж о скелетах; 4) сравнение размеров и характера распределения в вендских отложениях отпечатков и следов передвижения других организмов свидетельствует о неподвижном бентосном образе жизни большинства эдиакарских организмов.
Приуроченность остатков к турбидитоподобным осадкам, темпеститам и отложениям, характеризующим обстановки с активной гидродинамикой среды, может говорить о мутности вод в зонах их обитания, что противоречит гипотезе M.A.S.McMenamin о существовании в теле эдиакарских организмов эндосимбиотических фототрофов. Современные животные, имеющие фотосимбионтов, а также пассивные фильтраторы обитают в иных обстановках, избегая мутных вод и засыпания осадком. Нетипичность пиритизации остатков или их определенных частей свидетельствует, что серные бактерии также не были эндосимбионтами этих организмов, так как по сере, накапливающейся внутриклеточно или экстрагируемой в среду из клетки, в раннем диагенезе быстро образуется пирит. Да и сами обстановки, в которых обитали эдиакарские организмы принципиально отличаются от гидротерм или метановых сипов, в которых сегодня обитают организмы, имеющие хемосинтезирующих бактерий в качестве эндосимбионтов. Остатки эдиакарских организмов также не приурочены к илам, обогащенным органическим веществом, что отличает их от современных Xenophiophoria, с которыми их сравнивал А.Ю.Журавлев.
 |
Проведенный М.Б.Бурзиным анализ характера распределения остатков фитопланктона, фитобентоса, животных и деструкторов по отдельным временным срезам в поздневендском бассейне Русской плиты показал, что отсутствие связи между распространением основных представителей фитобентоса и эдиакарских организмов, а это может означать, что эти животные не входили в одни и те же экосистемы с бентосными водорослями, то есть эдиакарские организмы не были звеньями пищевых цепей пастбищного типа. Вместе с тем, отмечается некоторая отрицательная корреляция в распространении этих животных и некоторых деструкторов – нитчатых матобразующих серных бактерий, аналогичных современным Beggiatoaceae.
Все эти особенности находят свое объяснение в рамках предложенной A.Seilacher интерпретации вендских мягкотелых животных эдиакарского типа как осмотрофных организмов, питание, выделение и дыхание которых происходило за счет диффузии через поверхность тела.
 |
Основным продуктом питания для этих животных могли служить растворенные в воде органические вещества – продукты жизнедеятельности микроскопических водорослей и прокариот, широко развитых в поздневендских бассейнах. Относительно большие размеры и необычная морфология тела этих животных могут быть связаны с необходимостью, во-первых, подняться над поверхностью дна и, во-вторых, образовать большую поверхность, омываемую водой. Обитание их в зонах действия придонных течений объясняется тем, что даже относительно небольшие концентрации биогенов в потоке воды могут давать очень высокие суммарные значения на барьере, через которые поток проходит, кроме того течение уносит также от организма продукты выделения, позволяя тем самым существовать локально очень плотным популяциям, что наблюдается в ископаемой летописи. Такой тип питания возможен только в отсутствии плотных непроницаемых покровов и не требует рта и пищеварительной системы.
 |
В современном мире обнаруживается аналогия образа жизни и способа питания вендских мягкотелых животных с эндопаразитами, обитающими в пищеварительной или кровеносной системах. Отличие заключается в том, что вендские животные обитали не внутри тел конкретных организмов подчас в агрессивной внутренней среде, а были своеобразными псевдопаразитами (термин предложен М.Б.Бурзиным в 1996 г.) экосистем, образованных прокариотами и низшими эвкариотами, в лоне которых они могли существовать до тех пор, пока эффективные фильтраторы в самом конце венда или в начале кембрия не изменили характер распределения биогенов в бассейнах и не сделали невозможным столь экзотический образ жизни примитивных животных.
Известное разнообразие форм эдиакарских организмов, по аналогии с некоторыми современными эндопаразитами, может определяться обитанием в обстановках с разной гидродинамикой: в условиях широких плоских потоков будут преобладать более широкие и плоские формы, в струйных потоках – с узкой вытянутой формой тела, а в условиях слабых течений, в толще воды или при пониженной концентрации биогенов, связанной с особенностями среды обитания, – стебельчатые и кустистые формы.
© 2001 Палеонтологический институт РАН; Все права
сохранены