Н.Н.Каландадзе, А.С.Раутиан (Палеонтологический институт РАН)
СОПРЯЖЕННАЯ ЭВОЛЮЦИЯ НАЗЕМНЫХ И ВОДНЫХ ТЕТРАПОД КОНЦА ПАЛЕОЗОЯ - НАЧАЛА МЕЗОЗОЯ
Экосистемные перестройки и эволюция биосферы. Москва, "Недра", 1994. С. 174-180.
Самым слабым местом в наших представлениях о юрском экологическом кризисе в сообществе наземных (не морских) тетрапод [6, 10] является отсутствие систематического анализа его исторических предпосылок. Часть из них рассматривается в этом кратком сообщении. В дальнейшем мы предполагаем проанализировать и предыдущий, меньший по масштабам, экологический кризис в сообществе наземных позвоночных самого конца перми. Часть событий временной окрестности этого кризиса кратко описана в диссертации А.Г. Сенникова (1991), часть - в настоящем сообщении.
Материал. Основу нашего исследования составляет ценотическая интерпретация динамики таксономического и экологического разнообразия тетрапод конца палеозоя и мезозоя (рисунок). Первичные данные о таксономическом составе фаун и стратиграфическом распространении составляющих их таксонов заимствованы из многочисленных литературных источников [18, 20]. В целом вновь построенная динамика разнообразия качественно согласуется с ранее выполненными подсчетами [6, 17-19].
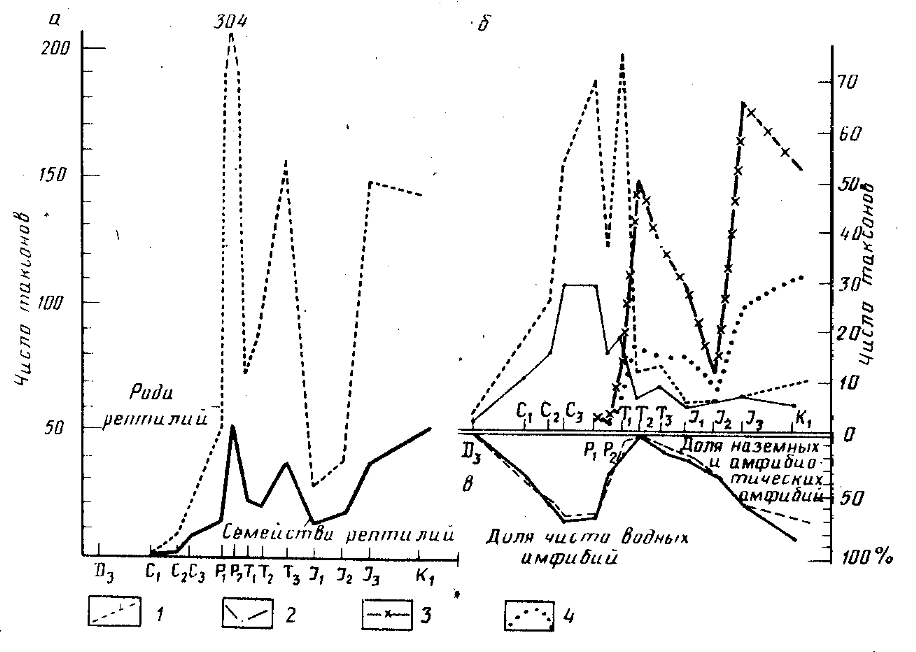
Динамика таксономического разнообразия наземных (а) и водных (б) рептилий и амфибий; доля (%) наземных и амфибиотичсских форм в фаунах амфибий разного возраста (в): / - рода амфибий, 2 - семейства амфибий, 3 - рода водных рептилий, 4 -семейства водных рептилий
Методика экологической интерпретации динамики таксономического разнообразия. Вероятность обнаружения и степень таксономического разнообразия той или иной группы организмов в палеонтологической летописи в данный момент геологического времени прямо пропорциональны всем основным критериям биологического прогресса, предложенным А.Н. Северцовым [12]: 1) фактическому таксономическому разнообразию группы в это время; 2) численности (биомассе); 3) широте адаптивной зоны и географического распространения [1, 22]. Таким образом, таксономическое разнообразие группы может быть использовано как некий интегральный показатель ее экологического процветания или угнетенности, а динамика разнообразия - как показатель изменения ее экологического состояния в течение геологического времени [11, 22].
Однако сравнение двух или более групп по этому показателю возможно лишь в том случае, если прочие условия, определяющие вероятность находки ископаемых - абсолютные размеры, наличие скелета и степень его склеротизации, условия захоронения, тафономия местонахождений [1, 22], способы ведения раскопок и т.п., - для этих групп достаточно сходны. Эти условия мы старались обеспечить сравнением таксонов относительно высокого ранга в глобальном масштабе и лишь с точностью до отдела стратиграфической шкалы. Все это позволило удовлетворяться достаточно ориентировочным экологическим сходством сравниваемых таксонов в пределах крупных адаптивных зон, понимая их в духе Дж.Г. Симпсона [14].
В тех случаях, когда рассматривались группы, занимающие различные адаптивные зоны, сравнение между ними носило чисто качественный характер на уровне сопоставления фаз максимального и минимального таксономического разнообразия, которые интерпретировались как фазы, соответственно, биологического прогресса и регресса.
Подобные соображения, но обычно в имплицитной (неявной) форме, лежат в основе подавляющего большинства исследований динамики таксономического разнообразия в течение геологического времени.
Основные результаты. Свободная и относительно автономная адаптивная радиация амфибий, превосходящих по разнообразию рептилий, продолжалась до ранней перми, когда семейственное и отрядное разнообразие их существенно превосходило современое. Доминантный блок крупных растительноядных и хищных рептилий в сообществе наземных тетрапод [13] только складывался. Первая, не имевшая продолжения попытка создания такого блока (пеликозавровая фауна) относится к позднему карбону-ранней перми Еврамерийской зоогеографической области [4-6]. Рептилии, известные с раннего карбона, в начале своей экологической экспансии находились в зависимости от амфибий, осваивая более доступные, не занятые или слабо освоенные ими биотопы. Об этом свидетельствует почти полное отсутствие специализированных водных рептилий в перми (таблица).
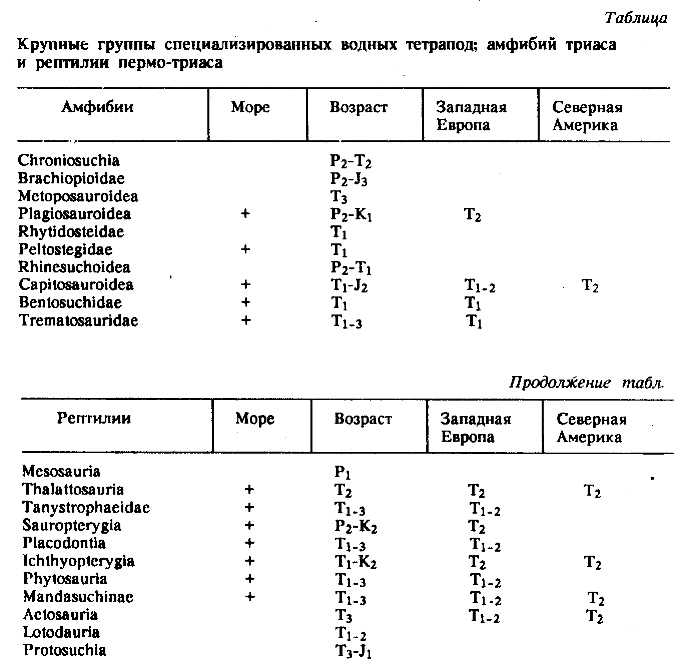
Примечание. В графе "море" отмечено (+) наличие находок в морских и прибрежно-морских отложениях; в графе "Возраст" дано полное стратиграфическое распространение групп. Наличие находок в изолированных морем в течение раннего - среднего триаса фаунах Западной Европы и Северной Америки отмечено в соответствующих графах.
Рост специализации, конкурентоспособности и разнообразия рептилий, насыщение первичной адаптивной зоны и выход за ее пределы, формирование доминантного блока крупных терапсид Гондванской зоогеографической области, филоценогенетические преемники которого просуществовали до конца мела, постепенно усиливали напряженность конкуренции в сообществе тетрапод. В этих условиях замедляется рост разнообразия амфибий, обладающих рядом морфофизиологических и экологических ограничений [2, З], а затем и рептилий. Кульминации этот процесс достиг в поздней перми, когда наблюдается первый в истории наземных позвоночных пик разнообразия рептилий и первый минимум - амфибий. В это время рептилии, очевидно, заняли все доступные им местообитания суши.
Дальнейшие события, как и предыдущие, развиваются в соответствии с предложенной нами моделью сопряженной эволюции сообщества и его биоты [9, 10]. Резко активизируется процесс вымирания и вытеснения менее конкурентоспособных групп за пределы сухопутного сообщества, в связи с чем сокращается общее разнообразие наземных тетрапод и открываются экологические лакуны. Последними, в частности, воспользовались перейазавры, занявшие в ряде биоценозов конца перми место основных консументов 1-го порядка. Тем же путем текодонты приобрели роль ведущих крупных хищников доминантного блока сообщества наземных тетрапод триаса. Предки тех и других были членами субдоминантного блока мелких инвертеброфагов [13]. Однако подробный анализ этих и других подобных случаев не входит в задачи настоящей статьи. Обратимся к не менее интересной проблеме конкурентного вытеснения.
Начиная с поздней перми, весь экогенез амфибий протекал "в тени" рептилий, а позже и млекопитающих. Из палеонтологической летописи поздней перми выпадают последние наземные (Dissorophidae) и амфибиотические (Lepospondyli, Eryopoidea) земноводные, резко обедняя набор жизненных форм. Исключение составляет лишь Micropholis из раннего триаса Южной Африки. Сохраняются, а в раннем триасе процветают, лишь высокоспециализированные крупные водные лабиринтодонты (понимаемые в широком смысле, включая Choroiosuchia - единственную уцелевшую в триасе группу антракозавров), не способные или практически неспособные выходить на сушу. Крупные группы лабиринтодонтов триаса либо начинают свою историю в поздней перми (Choroniosuchia, Brachiopoidea, Plagisaurbidea), либо, как Capitosauroidea и Trematosauroidea, имеют позднепермских предшественников (Rhinesuchoidea). Однако и эти группы постепенно сходят со сцены по мере освоения рептилиями водных пространств.
Миниатюризация большинства современных амфибий, изобретение плавания при помощи задних конечностей, а затем на этой основе скачущей локомоции (Anura); заселение быстрых ручьев с прохладной водой, а затем и приводно-почвенных биотопов (Urodela); глубокая специализация к жизни во влажной почве (Apoda) - все это создало жизненные формы, не характерные для рептилий [15]. Однако именно отмеченные особенности образа жизни понижают вероятность их сохранения в ископаемом состоянии [1, 22], чем и объясняется их весьма фрагментарная палеонтологическая летопись. Тем не менее, единичные находки триасовых предшественников современных отрядов амфибий известны. Это Triadobatrachus из раннего триаса Мадагаскара - единственный представитель отряда Ргоаnura, уже явно демонстрирующего стратегию адаптации, характерную для Anura. Это Epicris из позднего триаса Северной Америки, еще имеющий конечности, но отнесенный к отряду Apoda. И, наконец, Triassurus из позднего триаса Средней Азии, условно отнесенный к Urodela. Юрские и раннемеловые находки уже несомненно относятся к современным отрядам.
Возникновение, начиная с триаса, групп амфибий, практически не конкурирующих с рептилиями, подтверждает и вторичный рост доли наземных и амфибиотических форм после среднего триаса (см. рисунок).
Обострение конкуренции на суше стимулировало освоение водной стихии рептилиями, До поздней перми известна лишь одна группа специализированных водных рептилий (Mesosauria), представленная двумя родами в ранней перми Гондваны. Уже в поздней перми появляются древнейшие Sauropterygia, а в триасе - большая часть крупных групп водных рептилий (таблица). Первого в своей истории пика разнообразия они достигают уже в среднем триасе.
Большая часть, если не все, водных рептилий происходит от конкурентно вытеснявшихся на суше форм. Наиболее специализированные водные рептилии Ichthyopterygia, Sauropterygia и Placodontia имели предков среди Euryapsida - крупнейшего полностью вымершего таксона Sauropsida, a Tanystrophaeidae и Thalattosauria - среди примитивных Eosuchia. Менее специализированные Mandasuchinae в течение триаса неуклонно вытеснялись более крупными и специализированными Rauisuchinae (Rauisuchidae). Все специализированные водные текодонты относятся к Crodylomorpha - первично четвероногим архозаврам, вымершим или вытесненным в водную стихию (крокодилы) к концу позднего триаса.
Воспользовавшись выраженной водной адаптацией, отсутствовавшей у наземных рептилий, амфибии дали мощный всплеск разнообразия специализированных водных лабиринтодонтов в раннем триасе, успев это сделать до расцвета водных рептилий. В дальнейшем их разнообразие резко сокращается, особенно в юре, когда их единичные находки приурочены к реликтовым биотопам [16, 21, 23]. Наиболее поздняя находка лаби-ринтодонта (Plagiosauridae) относится к концу раннего мела [21] изолированной в это время Австралии [7, 8].
Глубина водной специализации лабиринтодонтов и текодонтов позволила некоторым из них, преодолев сравнительно небольшие морские проливы, заселить изолированные в раннем - среднем триасе [6] Западную Европу и Северную Америку (см. табл.). Причем в Западную Европу, расположенную ближе к гондванским странам, проникло большее число таксонов, чем в более удаленную Северную Америку. Особенно неожиданной оказалась способность амфибий преодолевать морские преграды, учитывая отсутствие случаев такого проникновения в фаунах перми [4-6]. Это обстоятельство подтверждает представление о значительном углублении водной специализации у триасовых лабиринтодонтов под давлением водных рептилий.
Таким образом, рост таксономического и экологического разнообразия рептилий и насыщение ими доступных местооби-таний суши, усилившие напряженность конкуренции в сообществе тетрапод, определили специфику всего дальнейшего экогенеза амфибий и освоение водной стихии рептилиями. Иными словами, позднепермский пик разнообразия рептилий явился поворотным пунктом, определившим вызревание в течение триаса предпосылок радикальной смены фауны амфибий во временной окрестности юрского экологического кризиса в сообществе наземных тетрапод. Следовательно, каковы бы ни были непосредственные внешние пусковые механизмы юрского кризиса, его последствия, по-крайней мере для амфибий, только этими причинами объяснить невозможно.
ЛИТЕРАТУРА
1. Ефремов И.А. Тафономия и геологическая летопись. М.: изд. АН СССР. 1950.
2. Иорданский Н.Н. Неравномерность темпов макроэволюции и ключевые ароморфозы // Природа. 1977. №6. С. 36-46.
3. Иорданский Н.Н. Эволюция комплексных адаптаций. Челюстной аппарат амфибий и рептилий. М.: Наука, 1990.
4. Каландадзе Н.Н., Раутиан А.С. К исторической зоогеографии наземных тетрапод конца палеозоя - начала мезозоя // Палеонтология. Стратиграфия. М.: Наука, 1980. С. 93-102.
5. Каландадзе Н.Н., Раутиан А.С. Межконтинентальные связи наземных тетрапод и решение проблемы шотландской фауны Элгин // Жизнь на древних континентах, ее становление и развитие. Л.: Наука, 1981. С. 124-133.
6. Каландадзе Н.Н., Раутиан А.С.- Место Центральной Азии в зоогеографической истории мезозоя // Ископаемые рептилии Монголии. - М.: Наука, 1983. - С. 6-44.
7. Каландадзе Н.Н., Раутиан А.С. Историческая зоогеография млекопитающих // Оперативно-информационные мат. к 1 Всесоюзн. совещ. по палео-териологии. - М.: Наука, 1989. - С. 28-30.
8. Каландадзе Н.Н., Раутиан А.С. Система млекопитающих и историческая зоогеография // филогения млекопитающих. - М.: Изд. МГУ, 1992.
9. Каландадзе Н.Н., Раутиан А.С. Эвристическая модель эволюции сообщества и его таксономического и экологического разнообразия // Труды ЗИН РАН. - 1993. (В печати).
10. Каландадзе Н.Н., Раутиан А.С. Юрский экологический кризис сообщества наземных тетрапод и эвристическая модель сопряженной эволюции сообщества и биоты // Проблемы доантропогенной эволюции биосферы. - М.: Наука, 1993. (В печати).
11. Парамонов А.А. Пути и закономерности эволюционного процесса // // Современные проблемы эволюционной теории. - Л.: Наука, 1967. -С, 342-441.
12. Северцов А.Н. Морфологические закономерности эволюции. - М.-Л.: Изд. АН СССР, 1939.
13. Сенников А.Г. Ранние текодонты Восточной Европы. - М.: Изд НИН РАН, 1991.
14. Симпсон Дж.Г. Темпы и формы эволюции. - М.: Изд. ин. лит., 1948.
15. Татаринов Л.П. О роли условий жизни в филогенезе земноводных. -М.: МГУ, 1953.
16. Шишкин М.А. Лабиринтодонт из юры МНР // Налеонтол. журнал. -1991. - № 1. - С. 81-95.
17. Benton M.f. Mass extinction among families of non-marin Tetra-pods // Nature. - 1985. - V. 306. - N 6031. - Р. 811-814.
18. Benton MJ. Mass extinction among families of non-marin Tetro-pods: the data // Mem. Soc. Geol. France. - 1987. - N 150. - P. 21-32.
19. Benton M.I. Patterns of evolution and extinction in Vertebrates // Evolution and fossil record. London.-Belhaven Press. - 1989. P. 218-241.
20. Carrol R.L Vertebrate paleontology and evolution. - N.-Y. W.H. Freman and Co, 1988.
21. Milner A. Late extinction of amphibia // Nature. - 1989. -V. 338. - P. 117.
22. Muller А.Н. Lehrbuch der Palaozoologie. - Bd. 1, 3. Jena. VEB G. Fischer. 1966-1976.
23. Nessov L.A. Late mesozoic amphibians and lizards of Somiet Middle Asia // Acta Zool. - Cracov, 1988. - V. 34. - N 14. - P. 475-486.