ЭНЕРГИЯ И ЖИЗНЬ
Воспроизведено по изданию:
Печуркин Н.С. Энергия и жизнь. — Новосибирск: Наука. Сиб.отд-ние, 1988.— 190 с. — (Серия «от молекулы до организма»).
Что движет эволюцибю жизн на нашей планете? В каком направлении развивается жизнь? Отчего «процветают» примитивные паразиты? Может ли разум человека влиять на судьбы Вселенной? На эти (и близкие им) вопросы делает попытку ответить автор, развивая энергетический подход к изучению живой природы.
Книга будет интересна для биологов, физиков, химиков, биофизиков, а также всех интересующихся общими вопросами развития.
Глава 1
О прогрессивном развитии в биологии
Мы, разумные существа, не должны забывать, что наша цивилизация — лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии солнечного излучения.
Ю. Одум
Вопрос о движущих силах развития жизни, о направлении эволюции, ее прогрессе, по образному выражению Н. В. Тимофеева-Ресовского, «это самое больное место всех биологов». В одной из последних работ Николай Владимирович писал, что не существует мало-мальски приемлемого, логичного понятия прогрессивной эволюции и что «сегодня никто не может дать серьезный ответ на вопрос, ведет ли отбор автоматически к прогрессивной эволюции» [Тимофеев-Ресовский, 1980. с. 63].
Проиллюстрируем неоднозначность и неопределенность трактовки прогрессивного развития жизни на нескольких примерах. Известные теоретики П. Эрлих и Р. Холм в книге «Процесс эволюции» [М., 1976] отмечают, что основной вопрос остается без ответа: почему в ходе эволюции ДНК создала для своего собственного воспроизведения трубкозубов и людей, тогда как бактерии и другие простые организмы, казалось бы, могут не хуже служить для этой цели? Наиболее образно высказался современный биолог-эволюционист Р. Левонтин [1978]. Он сравнил существующую теорию эволюции с машиной, активно перерабатывающей огромное количество сырья (особенно с помощью новых методов молекулярной биологии и генетики), которое, к сожалению, почему-то не превращается в готовый продукт.
Можно привести еще много таких высказываний. Их основу составляет неудовлетворенность современным состоянием эволюционной теории, особенно при объяснении прогрессивного развития. Вопрос действительно «больной», и однозначного мнения нет. Некоторые представители ортодоксальной точки зрения, полагая. что здание эволюционной теории полностью построено, склонны аттестовать любые виды критики как рецидив ламаркизма, другие соглашаются, что есть недоработки, поскольку в последние годы число попыток модифицировать представления о развитии жизни заметно возросло.
В целом эволюционная теория с успехом выполняет пока только функцию объяснения существующего и «практически неспособна выполнять задачи предсказания» [Борзенков, 1982, с. 16]. Описательность биологии бросается в глаза; однако умение теории отвечать на вопрос «как?», не затрагивая ответа на вопрос «почему?», уменьшает ее шансы называться теорией.
С исторической точки зрения понятно, почему представления о прогрессивной эволюции не были необходимым звеном развития дарвинизма на начальных этапах. Сам Ч. Дарвин, разрабатывая учение о происхождении видов путем естественного отбора, одним из главных достижений считал изгнание телеологии из теории об эволюционном развитии жизни. В то время это было действительно важно для материалистического понимания и объяснения эволюции, И поэтому Ч. Дарвин, всегда корректно и внимательно относившийся к взглядам оппонентов, резко выступал против укоренившихся натурфилософских представлений о внутреннем стремлении живых организмов к усложнению организации, к совершенству (по его словам, «этого дурацкого ламарковского стремления к совершенству»). Отсюда и появляется избыточное акцентирование внимания на стохастичности в противовес стремлению к предначертанным идеальным формам структуры или движения, представления о которых развивал великий Аристотель.
По Ч. Дарвину, процесс приспособления способен случайно приводить к изменениям, которые можно рассматривать как прогресс, но нет внутреннего механизма, обеспечивающего неуклонное совершенство. Сам Ч. Дарвин в своих письмах напоминал; «Никогда не применяйте слова „выше” или „ниже” (цит. по. [Майр, 1981, с. 20]).
Однако, оценивая факты, иллюстрирующие «уравновешение и экономию роста», Ч. Дарвин [1912, с. 117] отмечал, что они «могут быть подведены под общин принцип, а именно, что естественный отбор непрерывно пытается экономизировать всякую часть организации...».
Таким образом, начиная с основателя, дарвинизм не дает определения прогресса, особенно на уровне организма, но он дает в руки эволюциониста ясное понимание того, что существует очень сильное оружие, действующее на популяционном уровне,— естественный отбор (как отмечает Э. Майр, именно популяционное мышление было наиболее революционной идеей теории Ч. Дарвина).
Крупнейший синтез биологии XX в., исторически трудно складывавшийся — синтез дарвинизма и генетики — современная синтетическая теория эволюции (в основном микроэволюции), также не дает строгого определения «прогресса» в направлении эволюции. «Современная наука пока не в состоянии дать общего определения понятию «прогресс» в биологии. Возможно, такого рода определение не будет найдено и в будущем...» [Яблоков, Юсуфов, 1976, с. 268]. Однако, отмечая «постоянно растущую в процессе эволюции сложность биосферы, связанное с ней усиление давления жизни и разнообразие групп организмов», авторы цитированного учебника говорят о неизбежности «не только появления все более сложных существ и органов, но и их энергетического совершенствования» [Там же, с. 266].
Естествознание XIX в. по праву гордилось двумя крупнейшими достижениями: разработкой материалистической концепции эволюции в науках о живой природе и разработкой концепции энергии в развитии физики. Несомненно, что поиск внутренней связи между этими концепциями был предметом многих исследований. Так, К. А. Тимирязев еще в 1912 г. подчеркивал, что вопрос о космической роли растений является какой-то пограничной областью между двумя великими обобщениями прошлого века, между учением о рассеянии энергии и учением о борьбе за существование. Однако попытки найти простые формальные связи и вывести на их основе энергетические принципы развития жизни оказались практически безрезультатными. Более того, непосредственное приложение термодинамических законов к анализу явлений жизни привело к прямому противоречию: эволюция (развитие) живых систем происходит в направлении, противоположном указываемому вторым началом термодинамики (вместо деградации системы и роста энергии — повышение организации системы). Следовательно, согласно представлениям классической термодинамики, жизни как устойчивого явления не должно существовать. Сам факт наличия и развития жизни убедительно демонстрирует некорректность выводов подобного рода. «Жизнь не укладывается и посылки, в которых энтропия установлена», — отмечал В. И. Вернадский [l960, с. 85].
Потребовалось развить новую область, термодинамики — неравновесную термодинамику, на основе которой оказалось возможным ввести термодинамические критерии эволюции открытых систем. В применении к живым системам, открытость которых является одним из важнейших свойств, эти критерии определяют устойчивость стационарного состояния (а не равновесия — аналога смерти!), в котором скорость производства энтропии и, следовательно, рассеяния энергии минимальна.
И опять физический критерий эволюции не соответствует развитию реальных живых систем, которые в эволюции явно увеличили и рассеяние, и использование потоков энергии, пропускаемых через себя.
Физики и механики назвали энергию «царицей мира», а энтропию — ее «тенью». Понятие энтропии имеет двойственную природу. (Третью сторону — информационную — мы пока не затрагиваем.) С одной стороны, энтропия характеризует рассеиваемое системой «бесполезное» тепло, а с другой— является мерой упорядоченности (с ростом энтропии увеличивается беспорядок — в этом проявляется «теневой» смысл энтропии). Так вот в биологии, где упорядоченность структур почему-то возрастает, больше внимания уделялось энтропии, чем энергии. «Царица мира»— энергия оказалась в тени своей собственной «тени» — энтропии. Много говорилось об отрицательной упорядочивающей энтропии, присущей живым организмам. Даже солнечный свет предпочитали рассматривать как «мощный источник отрицательной энтропии», а не как поток энергии [Шредингер. 1972, с. 70]. А между тем для существования любого стационарного состояния открытой системы необходим поток свободной энергии извне, а не поток отрицательной энтропии в систему, или негэнтропии, как это следовало из вывода Э.Шредингера, наиболее часто упоминаемого в литературе. Самым простым подтверждением этому является возможность гетеротрофного роста клеток (т. е. синтеза сложных биополимеров и структур) на простых неорганических солях и углеводах (и даже углеводородах). Еще более убедителен хемо- и фотоавтотрофный рост, где используются лишь простые неорганические соединения и поэтому о питании отрицательной энтропией (или на языке термодинамики — высокоупорядоченными структурами) не может быть и речи. Однако абсолютно необходимым условием развития во всех упомянутых случаях является поток свободной энергии в различных формах (при окислении органических соединении; выделяемой в экзергонических реакциях окисления неорганических веществ типа реакции образования «гремучего газа»; энергии квантов света).
В целом термодинамический анализ возможности устойчивого существования стационарных состоянии диссипативных структур (т. е. обладающих определенным уровнем организации), согласно И. Пригожину, не указывает пути эволюции этих состояний.
Итак, и к настоящему моменту физика и биология не дают единой картины развития, перехода от сложных физических к простым (но еще более сложным на самом деле) биологическим структурам. Ситуация настолько драматична, что вместо ожидаемого синтеза имеет место прямое размежевание. Физики в данном случае «отгородились» принципом дополнительности, который ввел знаменитый Н. Бор. Согласно этому принципу, некоторые понятия несовместимы и должны восприниматься как дополняющие друг друга. «Идея дополнительности,— пишет известный физик-теоретик А. Б. Мигдал [1983, с. 39],— позволяет понять и примирить такие противоположности, как физическая закономерность и целенаправленное развитие живых объектов». Дополнительность, а с нею и несводимость физико-химической причинности и биологической целенаправленности декларировал и сам Н. Бор.
В свою очередь, и биологи «не остаются в долгу», заявляя об уникальности и неповторимости биологической эволюции. По словам крупного эволюциониста Э. Майра [1981, с. 26], «биологическая эволюция — это результат особых процессов, вторгающихся в особые системы», а органическая эволюция «отличается от эволюции Вселенной и от других процессов, с которыми имеют дело физики». Об эволюционном прогрессе Э. Майр пишет: «Никакой программы, которая регулировала или направляла этот прогресс, не было; он был результатом решений, принимаемых отбором, „на каждый данный момент”» [Там же, с. 23].
Такие размежевание наук о природе приводит к формулировке гипотез, иногда почти совпадающих с вымыслами художественной фантастики. Например, выдвигается так называемый антропоцентристский, или антропный, принцип. Суть его сводится к тому, что Вселенная устроена таким образом, чтобы в ней мог существовать человек. На примерах анализа фундаментальных физических констант показывается, что только в узком диапазоне их значений возможно существование сложных структур вплоть до живых систем. И поэтому данные константы (или их комбинации) именно таковы, чтобы наши живые структуры могли существовать. В последнее время делается вывод об особом положении Солнца в так называемом «галактическом поясе жизни», в относительно спокойной зоне синхронного вращения спиральных рукавов Галактики и межзвездного газа. Такой вывод близок к теологическим толкованиям, ставившим в основу мироздания нашу планету и ее Творца, создавшего человека по своему образу и подобию.
Но все успехи естествознания были связаны именно с отходом от антропоцентризма. Достаточно вспомнить Коперника, Галилея, Ньютона, Лапласа, Эйнштейна.
В биологии развития очевидным следствием антропоцентризма и натурфилософии является моноцентризм. Первым его проявлением (имеется в виду естественнонаучная трактовка эволюции) был организмоцентризм. Концепция организмоцентризма привела к ламаркистскому чисто телеологическому объяснению. Затем сложилось представление о виде, и он стал претендентом на центральное положение в биологии развития. Фундаментальное учение Ч. Дарвина о естественном отборе — результат видоцентризма. Ограниченностъ видоцентризма в том, что естественный отбор выступает чем-то самодовлеющим, «вещью в себе», с его непредсказуемым стохастическим механизмом действия (см. выше цитату из Э. Майра). Недаром яркий и оригинальный критик дарвинизма А. А. Любищев {М., Проблемы формы, систематики и эволюции организмов, 1982, с. 161, 196] обвинял оппонентов в том, что у них «отбор исполняет обязанности всемогущего господа бога», особенно в объяснении прогрессивной эволюции. Однако сам, находясь в плену организмо- и морфоцентризма. прибегал к схоластическим представлениям о единой гармонии природы и о наличии творческого начала, подобного сознанию, когда пытался объяснить сходства и различия форм среди неживой и живой природы.
Недостаточность одного из относительно новых «центризмов» (можно назвать его «ДНК- или геноцентризмом») в последнее время становится также наглядной, особенно при объяснении движущих сил развития жизни (вспомним цитату из П. Эрлиха и Р. Холма). Ответ молекулярных биологов и генетиков — «гены хотят жить и размножаться в замкнутой системе ДНК» — также нельзя признать научным, хотя он используется в завуалированной форме. При этом подходе центральным кирпичиком мироздания служат клетка и со компоненты, включая полимерные молекулы, но в целом это — все тот же организмоцентризм.
Бурное развитие экологии в последнее время приводит к развенчанию моно- и к пониманию идей полицентризма. Ценность экологических исследований для экспериментального изучения действия естественного отбора стала понятной после С. С. Четверикова, положившего начало генетико-популяционному подходу к изучению естественного отбора. Однако сложности экспериментов с популяциями и экосистемами сильно затормозили развитие экологического направления в эволюционных исследованиях. Необходимость «учета в эволюционных построениях глобального биосферного биоценотического фактора» [Татаринов, 1985, с. 16] все более осознается в настоящее время.
Количественные экологические исследования, особенно синэкологического направления, позволяют поставить экосистему, а точнее, круговорот веществ в ней в центр картины развития живой природы. И здесь, может быть даже несколько неожиданно, по совершенно очевидно, проявляется источник движения и развития жизни. Не таинственное стремление к самосовершенствованию, не самоорганизация биологических структур, «не могущих жить без метаболизма», а постоянная накачка потоком свободной энергии и вынужденность вращения вещества под воздействием этого возмущения.
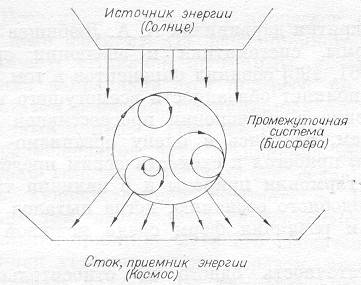
Рис 1. Схема трехзвенной системы с притоком энергии
Покажем схематическую систему, накачиваемую потоком энергии (рис. 1). Обязательным элементом является и третье звено — приемник энергии, или сток, в который энергия в рассеянном виде переходит от. промежуточной системы. Для нашей биосферы основным источником энергии служит Солнце, а приемником — Космос. В любых промежуточных системах кого типа (с протоком энергии) по законом физики возникают циклы в виде динамических структур. В большинстве случаев нам кажется, что они возникают сами по себе, и мы даже называем это явление «самоорганизацией структур», что придает некоторый налет загадочности.
Но в каждом случае обязательно есть «творец», и он имеет материальную природу. На Земле — это поток энергии от Солнца, который вызывает и организует круговороты в ограниченной системе (от простых физических: воды и воздуха, до сложного, биотического ). Однако функционально биотический круговорот. совсем несложен: это цикл реакций восстановления и окисления, где этап восстановления и подкачивается энергией квантов света. Развивался он постепенно из физико-химического. Обрастание сложными структурами — это вторичное явление. Будут выживать те, которые лучше вписываются в круговорот, способствуют его ускорению и умощнению (например, скорости химических реакций в живой природе возросли в миллионы и миллиарды раз). При этом круговорот использует все больше энергии и даже захватываются ее дополнительные потоки. Принципы развития очень просты и, главное, могут быть выражены количественно.
Роль «царицы мира», энергии, при таком подходе начинает проявляться по-настоящему, а ее «тень», энтропия, своим ростом только демонстрирует возрастание потоков свободной энергии, использованной экосистемой. Поэтому упомянутые представления Э. Шредингера о негэнтропии скорее можно назвать «поэтическими» (по выражению профессора Моровица, автора книги «Потоки энергии в биологии»), нежели физическими: для неравновесных систем энтропию очень часто трудно определить, тогда как энергия и ее потоки гораздо легче поддаются количественным измерениям.
Естественный отбор в этом случае перестает выступать в роли «господа бога», а к очевидностью выполняет и творческую (поступательное развитие круговоротов, прогресс), и сдерживающую, стабилизирующую роль (устранение избытков траты вещества, вплоть до явного регресса формы организмов). Телеологичность становится не только более явной, но и явно материалистической. А действие естественного отбора проверяется и контролируется не только на уровне видообразования (горизонтальном), но и на уровне совершенствования целых экосистем (вертикальном).
По методологии все современные концепции развития жизни можно отнести к трем основным типам: субстратные, энергетические и информационные.
Разработка общей теории биологического развития, прежде всего прогрессивного, должна естественным образом опираться на все три концепции, органически связывая их друг с другом во взаимодополнении и обогащении.
Исторически сложилось так, что первой стала развиваться субстратная концепция, начавшись с морфологии организмов. Дальнейшее развитие биохимии и физиологии углубило понимание биохимического единства живой природы, а бурный взрыв исследований по молекулярной биологии и генетике в последние десятилетия доказал единую генетико-молекулярную основу всех процессов жизнедеятельности. Триумфальные шествие этой концепции автоматически привело к абсолютизации некоторых ее положений, что сводится к одностороннему толкованию причин возникновения и развития жизни (вспомним: «Гены хотят жить и размножаться» ).
Информационная концепция, появившись самой последней, начала бурно развиваться с совершенствованием кибернетики и теории информации. Отметим, что даже в первом основополагающем труде по кибернетике Н. Винера речь шла об управлении и связи как в машине, так и в живом организме. Кибернетический подход позволил многое понять в развитии механизмов управления в живой природе, дав представление и о темпах эволюции. Однако и он, по попятным причинам, односторонен. Не ясно, как конкретно применять основанный на использовании этого подхода вывод о «возрастании ценности информации в эволюции».
Энергетическому подходу, как мы уже говорили, повезло меньше двух первых из-за различий в методологии физики и биологии. Однако именно он указывает направление развития сложных открытых систем, подвергающихся постоянной накачке энергией извне: это совершенствование циклов вещества; их умощнение и ускорение; возрастание переработки энергии каждой единицей структуры. Но и здесь абсолютизация недопустима, ибо один энергетический подход не способен дать представление о структурных особенностях эволюционирующих структур.
Например, рассуждая о возникновении жизни, точнее говорить не о «биохимическом предопределении» (а именно так называется книга Д. Кеньона и Г. Стейнмана о проблемах происхождения жизни, [М., 1972]), а об «энергетическом предопределении» зарождения живых структур. При этом важнейшая роль остается за субстратным подходом, т. е. за биохимическим «исполнением» жизни.
1. Субстратный плюс Информационный (без Энергетического) : С+И—Э. Явная телеология при всех долевых вариантах их отношений и в итоге — нет направления развития: «кто» эволюционирует — ясно, «как» и какими темпами — ясно, неясно — «куда».
2. Субстратный плюс Энергетический (без Информационного) : С+Э—И. Известно, «кто» и «куда» эволюционирует, неясно— «как» (с какой скоростью).
3. Энергетический плюс Информационный (без Субстратного): Э+И—С. Известно, «куда» и «как», только неясно — «кто» эволюционирует (биологические системы или, к примеру, промышленные технологии).
Для плодотворного, равноправного синтеза время только наступает, и пока менее всего готов к нему, слабее всех разработан энергетический подход — это следует подчеркнуть еще раз! Различные, но далеко не все аспекты этого подхода, по возможности в сочетании с субстратным и информационным, освещаются в данной книге.
Глава 2
Потоки энергии и «самоорганизация» структур
Все обменивается на огонь и огонь на все, как на золото — товары, на товары — золото.
Гераклит
Несмотря на внешнее разнообразие, для большинство процессов переноса (тепла, вещества, электричества) имеются общие закономерности, которые связывают скорость того или иного процесса и силы, вызывающие этот процесс.
Например, перенос вещества (диффузионный поток вещества) в направлении Х по закону Фика имеет вид
![]()
где dC/dX — градиент концентрации диффундирующого вещества С. В уравнении слева — поток, или перенос;справа — движущая сила процесса переноса. Коэффициент пропорциональности — это коэффициент диффузии D, характеризующий природу явления переноса и определенные условия его протекания. Такие же уравнения описывают перенос тепла, электричества. механическое движение.
Знак «минус» означает, что данный перенос и все типы переносов происходят в направлении отрицательных градиентов, т. е. должны приводить к уменьшению всех потенциалов, всех сил в тех простых системах, о которых мы здесь упоминаем. Такова реакция любой простой системы на возмущение: она стремится его уменьшить. А внешние возмущения, подвод энергии вынуждают систему опять организовывать «нейтрализующие» потоки. Без подвода энергии не будет движения (широко известна «тепловая смерть» изолированных закрытых систем, которые приходят в равновесие). Таким образом, в открытой системе постоянному действию сил соответствуют (и противостоят) постоянно работающие потоки. В этом и заключается суть известного в термодинамике принципа Ле-Шателье. Одна из наиболее распространенных формулировок этого принципа такова: всякая замкнутая система, выведенная из равновесия каким-либо возмущающим воздействием, стремится ослабить результат этого воздействия. Это — типичная формулировка, в которой затушевывается роль внешнего воздействия как источника движущих сил, а все внимание переносится на воздействующую систему. Получается, что система сама организует потоки, создает структуры, совершенствуется и т. д. В наше время стало модным говорить о «самоорганизации» сложных структур. Об этом чуть позже, а пока подумаем о том, как должна реагировать ограниченная по объему система, к которой постоянно подводится возмущающая систему энергия. Как правило, подвод энергии не может осуществляться равномерно (потому-то и есть градиенты). Единственная возможность непрерывного ответа, бесконечного во времени реагирования на внешнее возмущение, на постоянную накачку — это организация циклов вещества в замкнутой системе. Еще в 1949 г. академик В. Р. Вильяме писал, что единственный способ придать ограниченному количеству свойства бесконечного — это заставить его вращаться по замкнутой кривой.
Самой простой иллюстрацией сказанного является колесо — колесо старинной ветряной мельницы, вертушка стиральной машины или турбина новейшей ГЭС. Постоянный подвод (и расход) энергии в виде ветра, электричества или падающей воды заставляет эти колеса вращаться несчетное количество раз. Менее очевидна, по имеет ту же природу конвекция (вращение) воздуха в комнате под действием тепла от горячий печки или от батареи центрального отопления. Да и сами циклические потоки горячей воды по отопительной системе — это тоже результат накачки тепла от энергии парового котла.
Но не будем увлекаться примерами вращений, изобретенных изощренным умом человека. О естественных движителях, главными из которых являются поток солнечной энергии и внутреннее тепло Земли, мы поговорим особо — они этого заслуживают. О вызываемых ими глобальных циклах вещества — тоже отдельный разговор. А теперь, чтобы завершить обсуждаемую здесь тему о вынуждающих потоках и вызываемых ими циклах, поговорим об одном удивительном явлении. Это — возникновение сложных структур под влиянием потоков энергии в разного рода открытых системах, где циклическое перемещение вещества вдруг обретает устойчивые формы, когда сложная структура возникает, казалось бы, на пустом месте. Это явление носит название «самоорганизация сложных систем».
Впервые организацию сложной структуры в простой системе наблюдал французский физик Бенар в 1900 г. Это были шестиугольные ячейки, названные теперь в его честь бенарами. Получить их несложно. Надо налить на сковороду немного вязкой жидкости (например, растительного или минерального масла) и нагревать ее на медленном огне. Пока тепловые потоки малы, т. е. нагрев слаб, жидкость неподвижна, поток тепла усиливает молекулярную диффузию. Внизу образуется более горячий слой с меньшей плотностью, который стремится вверх, а на его место опускаются более холодные слои. По мере роста потока тепла должна возникнуть та самая конвекция, благодаря которой осуществляется выравнивание температуры в комнате. Однако при медленном возрастании перепада температур между верхним и нижним слоем, как обнаружил Бенар, неожиданно вся поверхность масла преображается — она разбивается на отдельные ячейки, большинство из которых имеет форму шестигранников, т. е. призматических шестиугольных цилиндриков от поверхности до дна сосуда. Образуется цикл по веществу: жидкость поднимается вверх в центре ячейки и опускается вниз у граней. Такая структура характеризуется более эффективным теплопереносом или более интенсивной диссипацией энергии. При этом происходит конкуренция и «выживают» именно шестигранники — наиболее эффективные рассеиватели энергии, способные полностью заполнять пространство [Кадомцев, Рыдник, 1981].
При дальнейшем увеличении теплопотока ячейки мельчают, их становится больше, а затем они исчезают. жидкость переходит в стохастическое турбулентное движение, так как теперь уже турбулентность более эффективно обеспечивает перенос тепла.
Таким образом, имеет место но «самоорганизация» структуры, а вынужденная организация циклических потоков вещества под влиянием внешней возмущающей силы ( потока тепла в данном случае).
Аналогичную физическую природу имеют вихри, открытые Дж. Тейлором в 1923 г. Они возникают в жидкости, налитой между двумя концентрическими цилиндрами, при вращении внутреннего цилиндра с определенной частотой.
В крупных масштабах интересное явление «самоорганизации» сложных, симметричных структур можно наблюдать в морях и океанах. Оно связано с образованием так называемых вихрей Лангмюра. Суть этого процесса заключается и следующем. При слабом ветре образуются довольно устойчивые вихри, в которых вода циркулирует вверх и вниз, образуя ряд зон подъема и опускания, которые чередуются в пространстве (рис. 2). Это может приводить к пятнистости планктона, скапливающегося в зонах подъема или опускания в зависимости от его плавучести, отрицательной или положительной. Иногда, в ночное время, эта сложная картина хорошо прослеживается по чередованию полос или более сложных структур свечения морской коды, так как многие виды планктеров способны к биолюминесценции. С увеличением ветра организованные в вихри структуры исчезают, начинается турбулизация, а с нею — образование новых структур, гораздо более подвижных. Мы к ним привыкли, это — самые обыкновенные волны. Однако не мешает подумать, что все они — тоже вариант «самоорганизации», вариант диссипативных структур, так как возникают при постоянно дующем ветре, не имеющем строгой волновой природы, хотя и несущем вихри. С их образованием диссипация энергии ветра несомненно увеличивается.

Рис. 2. Вихри Лангмюра, возникающие в достаточно спокойной воде при слабом ветре.
Зоны возможного скопления организмов с положительной (а) и отрицательной (б) плавучестью.
Конвективные ячейки можно наблюдать и при циркуляции воздушных масс, вызванной неравномерным нагреванием суши и атмосферы. Описаны и зарисованы почти правильные шестигранники из облаков со сторонами в несколько километров. Движущая сила — более высокая температура поверхности Земли. Организация циклов движения воздуха такая же, как в ячейках Бенара. Энергетическая выгодность шестигранников также не вызывает сомнения.
Наконец, несколько неожиданная картина «структуризации» Солнца была обнаружена при фотографировании участков солнечной поверхности через телескопы, поднятые в стратосферу. Поверхность оказалась состоящей из гранул (зерен). Как будто рассыпан слой зерна. Эти гранулы являются результатом конвекции во внешнем слое Солнца. В центре гранул солнечная плазма поднимается; по краям, отдав энергию на излучение и охладившись, она опускается. Разность температур приводит к контрасту яркости, что мы и воспринимаем как ячейки. Размеры гранул 200—1300 км, «живут» они в среднем 10 мин, затем границы старой гранулы размываются и вместо отмершей возникает новая структура. Легко можно видеть глубокую аналогию таких гранул с ячейками Бенара. Напомним, что во всех примерах динамические структуры образовывались в промежуточной системе, через которую шел поток энергии от источника к стоку (см. гл. 1).
Теперь можно сделать несколько выводов общего плана. Прежде всего ответим на вопрос, бывает ли «самоорганизация» сложных структур в таких системах? Корректнее отвечать — не бывает! Не бывает «само» организации. Бывает вынужденная организация циклических потоков вещества в виде динамических структур под влиянием внешней возмущающей силы. И такая структура (от маленькой ячейки на сковороде, облачных шестигранников до огромных солнечных гранул) существует до тех пор, пока она более эффективно выполняет функцию переноса, чем если бы этот перенос осуществлялся без нее. Итак, выводы в сжатой форме.
1. Внешний движитель — источник энергии вызывает циклические переносы вещества, организуя динамические структуры в промежуточной системе. Сам момент появления такой структуры случаен, он связан с флуктуациями в системе.
2. Эти структуры более интенсивно выполняют функцию переноса, выживают в конкурентной борьбе наиболее эффективные (их можно назвать «приспособленные» ).
3. Если при изменении потока энергии структура начинает хуже выполнять функцию переноса, то она либо заменяется на новую, либо исчезает совсем.
Самый общий вывод: в системе с протоком свободной энергии структура вторична, функция первична.
Этот вывод можно было бы даже окрасить в теологические тона (кому нужна функция: уж не от бога ли все это?), но вопрос этот сам собой снимается, так как есть материальный носитель, «возмутитель спокойствия» и организатор движения: на нашей планете это прежде всего поток солнечной энергии. Некоторую роль играет и поток тепловой энергии от центра Земли, особенно за геологическое время. Рассмотрим подробнее поток свободной энергии от Солнца.
Глава 3
Солнце — главный источник энергии для поверхности Земли
О солнце, ты живот и красота природы,
Источник вечности и образ божества!
Тобой живет земля, жив воздух, живы воды,
Душа времен и вещества!
А. П. Сумароков
Из большого числа возможных источников энергии, имеющихся у нашей планеты, первое место, несомненно, следует отдать солнечному потоку, который поддерживает необходимые температурные условия Земли (чтобы мы не испарились, перегревшись, или не замерзли, переохладившись).
Культ Солнца был развит у большинства народов, населяющих Землю, и недаром поток солнечной энергии составляет основу всех потоков энергии на нашей планете (рис. 3).
К внешней границе тропосферы подводится поток солнечной радиации примерно 1000 ккал/(см2·год) (или около 2 ккал/(см2·мин)). Из-за шарообразности Земли на единицу поверхности внешней границы тропосферы в среднем поступает четвертая часть — примерно 250 ккал/(см2· год). Треть этого потока отражается, и, следовательно, Земля поглощает 167 ккал/(см2· год). Из них 59 ккал/(см2· год) поглощает атмосфера, и на долю поглощения земной поверхностью приходится 108 ккал/(см2·год). Эта энергия «перерабатывается» различными способами. В виде длинноволнового инфракрасного излучения с поверхности Земли уходит 36 ккал/(см2· год).

Рис. 3. Укрупненная схема энергетического баланса Земли
(составляющие энергетического баланса, ккал/(см2 ·год)) [Будыко, 1984].
Благодаря парниковому эффекту поверхность Земли получает около 72 ккал/(см2·год) радиационной энергии, часть которой — 60 ккал/ (см2·год) — идет на испарение воды (нижний кружок на рис. 3), а часть — 12 ккал/ (см2·год) — возвращается в атмосферу через турбулентные потоки воздуха.
Основной передатчик тепла между космосом и Землей — атмосфера — получает от Земли «свои» 60 ккал / (см2·год) за счет конденсации водяных паров (верхний кружок на рис. 3), упомянутые 12 ккал/(см2·год)— за счет турбулизации и непосредственно от радиации Солнца — 59 ккал/(см2 · год). Итог: приход равен 131 ккал/ (см2·год). И соответственно расход тепла через эффективное излучение — той же величине —131 ед. Вместе с результирующими 36 ккал/(см2·год) длинноволнового излучения от земной подстилки мы и получим расход в целом — 167 ккал/ (см2·год), в точности равный приходу энергии с потоком солнечной радиации.
Таким образом, на нашей планете работает «система жизнеобеспечения» с определенным интервалом температур. Среднегодовая температура составляет 14,25°С, при этом в Северном полушарии средняя температура 15,2°С, а в Южном — только 13,3°С из-за высокой отражательной способности ледового панциря Антарктиды.
Из 72 ккал, поглощаемых каждым квадратным сантиметром земной поверхности в год, океан «забирает» почти вдвое больше, чем суша, — 90 и 50 ккал соответственно. Это объясняется большой теплоемкостью воды и ее подвижностью. Океаносфера является мощным аккумулятором солнечного тепла, она накапливает в 21 раз больше того количества тепла, которое за год поступает от Солнца ко всей поверхности Земли (7,6 · 1023 ккал по сравнению с потоком в 3,65 · 1020 ккал/год). Поэтому ее взаимодействием с атмосферой определяется погода на земном шаре. Тепло, поглощаемое в тропиках, переносится течениями в высокие широты, смягчая климат умеренных и полярных областей. Один Гольфстрим несет в 22 раза больше тепла, чем все реки суши.
В целом гидросфера работает под влиянием накачки солнечной энергии как гигантская тепловая машина. Можно даже оценить ее коэффициент полезного действия. «Чистая» энергия движения, перемещения воздушных и водных масс, т. е. та часть, которая может совершать нужную нам работу, оказывается совсем небольшой: для атмосферы (со средней скоростью ветра, несколько превышающей 10 м/с у поверхности Земли)— всего 1,6% от поглощаемого солнечного тепла, а для океаносферы (со средней скоростью течения во всей толще вод, равной 3,2 см/с) — еще на пару порядков ниже. Конечно, одна из наиболее серьезных энергетических затрат — это затрата энергии на физический круговорот воды, прежде всего на испарение. Ее тоже можно оценить по уже приведенным данным. Около 55% — таков расход энергии, дошедшей до земной поверхности, на испарение.
В атмо- и гидросфере сложное переплетение циклов, различающихся по пространству и времени существования, определяет и мгновенное состояние — погоду в любой точке Земли и климат в каждой зоне. Климат есть результат усреднения прошлых погод каждого дня в каждой точке. Не затрагивая очень спорного, но злободневного вопроса о прогнозировании погоды, подчеркнем только, что само понятие о климате было введено еще учеными Древней Греции. Слово это греческого происхождения (klima) и означает «наклон». То есть еще в то время греки хорошо понимали, что климат местности зависит от среднего наклона солнечных лучей к поверхности Земли. Молодцы, древние греки! Представление о первоисточнике движения у них было самое верное.
Глава 4
Круговороты вещества в неживой природе
... Опять в морской простор
Спешит вернуться то, что небо сушит.
А реки снова устремляют с гор ...
А. Данте
4.1. Циклы воды и воздуха
Перейдем к описанию наиболее существенных круговоротов вещества на нашей планете, которые, как мы знаем, «раскручиваются» не сами по себе, а потому, что их непрерывно «заставляют» это делать потоки энергии, главным образом солнечной. Проанализируем работу одного из наиболее интенсивных циклов — круговорота воды и воздуха на земной поверхности.
Из всех круговоротов циклическое перемещение огромных количеств воды было наиболее интересным и важным для человечества. Общеизвестно, что древний человек селился главным образом по берегам рек и озер; сильно страдал от избытка и недостатка осадков, приносимых ветрами. Дождю, ветру, воде поклонялись, о них слагали легенды, их обожествляли (достаточно вспомнить легенды о страшном ливне и Всемирном потопе, которые в разных вариантах сохранились у многих народов).
Вполне естественно, что человек хотел знать, откуда берутся реки и ручьи, почему они не иссякают (или вдруг иссякают), почему выпадает дождь или снег, почему вода в море соленая, а в реке пресная и т. д.
Давайте вкратце рассмотрим на примере развития представлений о круговороте воды, как шло накопление знаний, каковы были наиболее типичные заблуждения и ошибки, как пробивала себе дорогу Истина. Особое внимание уделим вопросу, наиболее важному для энергетического подхода и для методологии в целом,— вопросу о движущих силах круговорота.
Сначала вспомним Фалеса Милетского (VI в. до н. э.), который считал воду основой всего многообразия природы. Он полагал, что ветер вгоняет воду в недра Земли, горные породы выдавливают ее вверх, откуда и текут реки и ручьи.
В основу учения Аристотеля (IV в. до н. э.) легли представления о круговороте несколько ранее жившего Демокрита, родоначальника атомистического учения. Демокрит, объясняя разливы Нила, утверждал, что проливные дожди в бассейне великой реки связаны с испарением снега и воды на севере и их переносом пассатами. Аристотель справедливо считал, что источником всех вод является влага атмосферы, но дождевых осадков недостаточно для образования и поддержания речных потоков. Поэтому он придумал конденсационную теорию образования воды из воздуха в многочисленных холодных земных пустотах. (Таким образом, опирался на трансформацию основных элементов: воды и воздуха).
Плиний Старший (1 в. н. э.), автор «Естественной истории»— сочинения, популярного вплоть до XVI в., писал, что вся вода стремится к центру Земли и собирается в низких местах, а поднимается вверх до истоков ручьев и рек и изливается в виде родников с помощью духа, который побуждает воду «раздуваться».
Варварство и феодализм вплоть до эпохи Возрождения отличались упадком и застоем в развитии познания природы, в том числе и нашей проблемы.
Например, в период средневековья единственное представление, разрешенное христианской церковью, основывалось на высказывании из «Книги Экклесиаст»:
Восходит солнце, и заходит солнце, и на место свое поспешает,
Чтобы там опять взойти;
Бежит на юг и кружит на север, кружит, кружит, на бегу своем ветер,
И на круги свои возвращается ветер;
Бегут все реки в море,— а море не переполнится,
К месту, куда реки бегут,—
Туда они продолжают бежать *.
* Цит. по: Поэзия и проза Древнего Востока: Библиотека всемирной литературы.— М.: Худож. лит., 1973.— С. 638.
Высказывание довольно точное и емкое, все участники событий перечислены, но вопрос о движущих силах круговорота, например, как и почему вода поднимается вверх, к истокам рек, увы, практически не ставится, да и не мог ставиться при теологическом толковании мира. (Так создано богом, так надо — вот стандартный догматический ответ на все вопросы).
Эпоха Возрождения поистине возродила интерес к познанию природы, дала миру гигантов мысли и чувства. Например, Леонардо да Винчи (1452—1519), задумываясь об источнике питания рек, близко подошел к пониманию влияния теплоты Солнца, но атмосферные осадки не считал основными в круговороте. Как и древние греки, он полагал, что морская вода способна подниматься вверх. Он уподоблял всю Землю живому организму (вот они, естественные истоки организмоцентризма, который так дорого стоил биологии, но об этом—позже!), где движение воды соответствует движению крови. Вода Земли, как кровь организма, согласно великому Леонардо, оживотворяет Землю и ее создания.
Но загадка источника движущих сил (подъема воды к истокам рек) так и осталась недоступной даже для лучших умов натурфилософии всех времен. Хотя для нас с вами разгадка кажется очевидной. И не в том дело, что мыслители прошлого были слабы; упомянутые здесь имена, сохраненные историей, сами говорят за себя. Слабость заключалась в натурфилософском подходе к изучению природы. Выделим две главные причины. Первая — не вставши на путь строгого количественного эксперимента, нельзя было выбрать единственно правильную из большого числа гипотез, призванных объяснять то или иное природное явление. Поэтому так важна была и вторая причина: схоластичность и авторитет учителя.
И только XVII в. с развитием капитализма, с его высокими требованиями к техническим средствам привел к быстрому прогрессу опытной науки — естествознания в современном понимании.
И сразу все стало на свои места! Почти триста лет назад знаменитый и в наши дни Э. Галлей (описанная им комета недавно вновь приближалась к нам) измерил в небольшом сосуде ежедневное испарение воды солнечными лучами. Убыль воды составила примерно 0,1 дюйма (1 дюйм = 2,54 см). Он пересчитал это на поверхность Средиземного моря, взял приток воды в него примерно в 90 стоков Темзы и получил удивительные данные, которые показали, что испарение не только компенсирует сток, но и превышает его в 3 раза! А ведь именно нехватка испарения и считалась камнем преткновения для натурфилософов, начиная с Аристотеля.
Все дальнейшее стало уже делом техники, хотя и не простым. Но для оценки методологии, для нашего рассмотрения это уже не имеет принципиального значения. В XVIII и XIX вв. потребовалось много усилий, чтобы баланс круговорота был уточнен в деталях. К настоящему времени поверхность Земли покрыта сетью гидрометеорологических станций и балансы круговорота просчитываются на больших вычислительных машинах для длинных и коротких временных интервалов. Для нас важно, что организация этого потока вызывается потоком энергии солнечного излучения. Факт сам по себе банальный, но поучительный, если не забывать историю его установления — более чем двухтысячелетние заблуждения светлых умов натурфилософии.

Рис. 4. Гидрологический цикл на нашей планете.
1—океан; II—сточные области суши; III — замкнутые области суши. Стрелками обозначен перенос влаги. Цифры у стрелок соответствуют объемам воды в десятках тысяч кубических километров.
Приведем данные по глобальному круговороту воды на Земле (рис. 4). В целом для земной поверхности величина осадков, равная испарению за год, составляет 113 см [Будыко, 1977]. Общий объем испаряемой воды в год чуть превышает объем Черного моря.
Интересно оценить интенсивность участия различных вод в круговороте: глубинные воды имеют цикл обновления, они возобновляются за 5000 лет, воды Мирового океана — за 3000, воды непроточных озер в среднем — за 200 лет, воды проточных озер — за десятки лет, почвенная влага — за год, речные воды — за 12—15 дней, атмосферная влага—за 8—10 дней.
Таким образом, особенно напряженны процессы влагообмена, связанные с атмосферой и речными стоками.
Поражает воображение сам факт длительной, в течение миллионов лет, «жизни» любой крупной реки. Если перегородить русло и изъять источники, то вся вода реки может вытечь за несколько дней и оголится русло до самого океана! Устойчивость этой подвижной системы обеспечивается тем, что она является неотъемлемой частью «вечного» круговорота, который раскручивается самым мощным движителем на нашей планете — Солнцем.
Оценивая глобальный круговорот воды, конечно же, не следует забывать о мощнейших океанических и морских течениях, которые могут практически постоянно переносить громадные объемы воды. Всем известен Гольфстрим, который в десятки раз мощнее всех рек суши, вместе взятых, и который своим теплом греет Европу и даже влияет на климат Сибири. Менее известно антарктическое циркуляционное кольцо, которое является главным переносчиком вод из океана в океан (кроме «маленького» Северного Ледовитого).
Даже реки никогда не представляют собой направленные ламинарные потоки, текущие строго в одном направлении. Вот, например, как образно описывает известный писатель В. П. Астафьев в романе «Царь-рыба» вид на Енисей с борта самолета: «Поле реки, точно от взрывов мин, опятнанное воронками — кружилась вода на подводных каргах, было широко и в общем-то покойно, лишь эти воронки да царапины от когтей каменных шиверов и в крутых поворотах сморщенная, как бы бороной задетая гладь только и показывали, что внизу под нами все же не поле, а река, наполненная водой и неостановимым движением...»
Всем нам хорошо знакомы завихрения воды в любом бегущем ручейке, а такие большие заводи, омуты, в которых вода закручивается и движется навстречу основному потоку, памятны с детства.
Круговорот воздуха тесно связан с круговоротом воды, так как атмосфера является основным источником водяных паров. В отличие от воды, которая испытывает фазовые превращения (лед — вода — водяной пар) и гораздо более энергоемка, воздух только механически перемещается согласно градиенту температур и давлений. Зато скорость его перемещения может быть очень высокой по сравнению с водными потоками. Достаточно вспомнить о тропических ураганах или арктической пурге. И хотя иногда кажется, что нет никакой системы в перемещении воздушных масс, на самом деле глобальный круговорот воздуха достаточно хорошо организован и поддерживается нагревом поверхности Земли, равномерно распределенным в пространстве потоком солнечных лучей. Наибольшая разность температур отмечается между тропическими и полярными районами.
Воздух тропиков, нагреваясь, расширяется и поднимается к верхним слоям атмосферы. Воздух полярных районов охлаждается и, сжимаясь, опускается вниз и растекается по поверхности Земли, устремляясь в тропические районы. Вращение Земли приводит к отклонению циркуляции воздуха от меридионального направления. Особенности земной поверхности вызывают дополнительные завихрения и потоки, образование которых трудно предсказать (бури, ураганы, циклоны, антициклоны, смерчи и т. д.). Однако имеются и более постоянные перемещения воздушных масс. В приэкваториальной зоне большую роль играют постоянно дующие ветры — пассаты (северо-восточные — к северу и юго-восточные—к югу от экватора). На границах материк — океан перенос воздуха и влаги осуществляется периодическими ветрами — муссонами, которые компенсируют разность температур из-за неодинакового нагрева воздуха над сушей и водой. Летние муссоны дуют с моря, более холодного, зимние муссоны несут более холодный воздух суши. И наконец, существуют менее мощные ветры — бризы, которые меняют направление в течение суток. Этот ветер способен проникать на 40 км в глубь суши и захватывать столб воздуха высотой до 0,5 км, выше — ветер дует в обратном направлении. На этом примере можно хорошо видеть работу одного из частных круговоротов, вызванного к жизни потоком солнечной энергии.
Рассматривая круговороты воды и воздуха, мы должны учитывать их разномасштабность: от огромных глобальных до мельчайших локальных. И все они вызываются потоками энергии, основу которых составляет поток энергии от Солнца. Число локальных круговоротов неисчислимо.
4.2. Геологический цикл
На земной поверхности под влиянием потока солнечной энергии происходит выветривание горных пород, их разрушение. В результате такой эрозии в течение года ветрами, реками, ледниками уносится до 1010 т твердого и растворимого вещества.
Помимо физического переноса веществ происходит с неодинаковой скоростью выщелачивание химических элементов из основных пород. Например, кальций, калий, натрий, хлор, сера сравнительно легко уходят из кристаллических решеток первичных минералов; кремний, алюминий, титан, марганец, наоборот, крайне плохо растворимы. В образующихся рыхлых породах коры выветривания широко развиты процессы окисления (железо, сера, марганец в изверженных породах, как правило, двухвалентны, а в коре выветривания дают соединения Fe2O3; Na2SO4; MnO2(Fe+3; S+6; Mn+4)) [Перельман, 1977]. Часть выносимого осадка остается на шельфе, а часть доходит и до океанических впадин. Со временем осадок захороняется, погружаясь на все большую глубину, в том числе и под собственным давлением. При этом осадочные породы испытывают метаморфизм и подвергаются переплавке за счет тепла Земли. После чего в зонах подъема, частично перемешиваясь с глубинным веществом, они поднимаются вверх в горных цепях в виде магматических пород. И опять подвергаются выветриванию, попадая в следующие химические циклы.
Геохимические процессы, грубая канва которых приведена выше, совершаются на протяжении многих тысяч и миллионов лет. Понятно, что они испытывают очень сильное влияние биотического круговорота, интенсивность процессов в котором несравненно выше. Растительность, почва и особенно микроорганизмы вносят заметный вклад в геохимические явления, по еути превращая их в биогеохимические. В.И.Вернадский, [1965] в одной из последних работ писал, что процессы выветривания, в частности резко выраженные в биосфере, всегда биогенны и биокосны; микроскопическая жизнь в них играет ведущую роль.
Глава 5
Живая природа. Нехватка вещества и его циклы
... Вся поверхность шара состояла из капель, плотно сжатых между собой. И капли эти все двигались, перемещались и то сливались из нескольких в одну, то из одной разделялись на многие. Каждая капля стремилась разлиться, захватить наибольшее пространство, но другие, стремясь к тому же, сжимали ее, иногда уничтожали, иногда сливались с нею.
— Вот жизнь,— сказал старичок-учитель.
Л. Толстой
5.1. Главная ячейка жизни — клетка
Определение жизни с позиций функционального подхода (метаболизм, размножение, расселение в пространстве) можно дать в следующей форме [Печуркин, 1982]: это открытая система, развивающаяся на основе матричного автокатализа под влиянием внешнего потока энергии, ограниченная по веществу и использующая циклы реакций.
Что такое матричный автокатализ, известно из школьного курса биологии, и мы не будем его описывать.
Энергетические траты живой клетки осуществляются через запасание энергии в молекуле аденозин-трифосфата (АТФ) и передачу ее соответствующим «работающим» молекулам (подробнее позже).
Полученную энергию клетка расходует на поддержание активности и многочисленные синтезы. Несмотря на сложность и большое разнообразие органических молекул и клеточных структур, все они строятся из малого набора простых соединений-предшественников, поступающих из внешней среды. Начало синтезов идет от двуоксида углерода, воды и минеральных солей. На первом этапе они превращаются в промежуточные продукты, из которых на втором этапе создаются основные строительные блоки и среди них аминокислоты и мононуклеотиды. На третьем этапе происходит сборка четырех типов макромолекул из строительных блоков. На следующих этапах образуются функциональные надмолекулярные комплексы, которые на высшем уровне организации объединяются в целостный организм — клетку как основную (и единственную) ячейку жизни.
Давая краткое описание работы клетки, подчеркнем важнейшую особенность жизни. «Поразительным открытием молекулярной биологии за последние три десятилетия» назвала обнаруженную всеобщность фундаментальных химических процессов в живой клетке известная исследовательница эволюции жизни, профессор Бостонского университета Л. Маргелис, [1983]. Действительно, функциональное единство самых существенных биологических феноменов не может не поражать. Так, генетический код, определяющий соотношение между последовательностями нуклеотидов и аминокислот в белке, универсален. По существу, он одинаков у всех изученных организмов — от древнейших бактерий до человека.
Связывание информационных РНК, комплементарных генной ДНК, с рибосомами при синтезе белков, по-видимому, тоже универсально. Наконец, энергетическая валюта — АТФ — также едина для представителей всех царств живого мира.
Остановимся еще на одной немаловажной особенности клеточной организации. Это — энергетическая экономичность генетического кода. Одним из ее проявлений может служить корреляция между распространенностью аминокислоты в белках и энергетической стоимостью ее синтеза. Из статистического анализа более 600 белков вирусов, микроорганизмов, растений и животных удалось достоверно установить, что чем выше затраты АТФ на биосинтез данной аминокислоты, тем реже входит она в состав белков. С этой точки зрения становится понятным явление незаменимости аминокислот, т. е. неспособности некоторых из них синтезироваться в организмах высших животных и человека. (Поэтому нам и требуется животный белок, содержащий эти аминокислоты, в свою очередь полученные от растений.) Оказывается, что энергетическая эффективность биосинтеза белка у организмов, не синтезирующих, а потребляющих извне эти аминокислоты, на целых 20 % выше, чем у организмов, которые синтезируют все необходимые аминокислоты. Кроме того, для синтеза незаменимых аминокислот требуется гораздо большее число ферментов, чем для синтеза заменимых, что также связано с дополнительными тратами вещества и энергии.

а: 1 — жгутик, 2 — клеточная мембрана, 3 — нуилеоид (генофор), 4— малые рибосомы, 5— нецеллюлозная стенка; б: 1 — пластида, 2 — ядерная мембрана, 3 — большие рибосомы, 4 — кинетохор, 5 — хромосома, 6 — эндоплазматический ретикулум, 7 — ундулиподия 9 + 2 (микротрубочки), 8—кинетосома 9 + 0, 9— клеточная мембрана, 10 — клеточная стенка, 11 — митохондрии.
Выделяют две главные таксономические единицы — надцарства: прокариоты и эукариоты.
Название «прокариоты» происходит от латинского слова pro (вперед, вместо) и греческого káryon (ядро) (рис. 5). Клетки прокариот не содержат ядра с мембраной, их кольцевая ДНК располагается в клетке свободно. Слабо выражено деление пространства клетки на отдельные части. Ограничено количество клеточных компонентов — органелл. Отсутствуют пластиды и митохондрии, отвечающие за энергетические превращения в более высокоорганизованных клетках. Клеточная стенка состоит из гетерополимерного вещества — муреина, которое не встречается у других групп организмов. Аппараты движения (жгутики) либо отсутствуют, либо относительно просто устроены. Наконец, размеры прокариотных клеток очень малы, в среднем единицы микрометров (мкм), что находится на грани разрешающей способности светового микроскопа.
Простота структуры у прокариот компенсируется высокой лабильностью и многообразием метаболических процессов. Способ питания может быть как автотрофным, так и гетеротрофным. Прокариоты питаются путем всасывания, или абсорбции, питательных веществ через клеточную стенку. Обычный тип размножения бесполый, простое деление пополам, однако обмен генетическим материалом иногда происходит при слиянии клеток за счет парасексуальных процессов.
Если прокариоты справедливо считаются первичными формами, возникшими в начале эволюционного пути, то развившиеся из них эукариоты представляют собой следующую ступень эволюции (см. рис. 5).
Клетки эукариот имеют выраженное ядро, окруженное мембраной. Генетический материал (ДНК) связан с белком в отдельных образованиях — хромосомах. Имеется целый набор органелл клетки: вакуоли, гранулы, нитевидные и палочковидные структуры. Энергетические процессы локализованы в митохондриях. Хорошо развита эндоплазматическая мембранная система, несущая множество пузырьков и цистерн. Мелкие нуклеопротеидные частицы — рибосомы, в которых производится синтез белков, либо связаны с мембранами эндоплазматической сети, либо взвешены в цитоплазме. Обычный тип размножения — через половой процесс с чередованием слияния ядра в зиготе и редукционного деления с образованием половых клеток — гамет. Возможны и неполовые способы размножения: простым делением, как у прокариот, почкованием, образованием спор и т. д., что наиболее часто встречается у микроорганизмов. Жгутики, или реснички, если они есть, имеют более сложное, чем у прокариот, строение. Питание эукариот может быть автотрофным и гетеротрофным: абсорбционным, как у прокариот, или голозойным, при котором пища заглатывается и перерабатывается внутри организма.
Размеры эукариотных клеток намного превышают размеры прокариотных, к примеру, митохондрии сопоставимы по величине с отдельными прокариотными клетками.
Подводя итог краткому описанию структуры и функции основной единицы живого — клетки, выделим «три кита» биологической организации: биохимическое единство, экономия материала и энергетическая эффективность.
5.2. Биологический взрыв и нехватка вещества
Одной из важных черт жизни является способность к рождению себе подобных, которые также могут размножаться, давая новые единицы, способные к размножению, и т. д. Это и есть известное явление автокатализа в физике, химии, при механических переносах: это — лавины, сели, реакции возгорания и взрыва (от простого пожара до атомного и термоядерного взрывов). Математически такому процессу соответствует уравнение экспоненты. Если размножение идет простым удвоением, типа бактериального деления пополам один родитель дает два потомка и т. д.), то формула для числа потомков Х имеет вид
где g — длительность одного поколения, т. е. время от рождения до следующего рождения; n — число поколений; Х0 — начальное число размножающихся единиц (клеток, организмов).
Первое представление об экспоненте и ее стремительном росте во времени связывают со старинной восточной легендой о появлении шахмат. Правитель решил отблагодарить мудреца-изобретателя и предложил ему самому назначить награду, гарантируя исполнение. Мудрец попросил, казалось бы, немного: дать ему одно зернышко — на первый квадратик шахматной доски, два — на второй, четыре — на третий и т. д., удваивая каждый раз цифру (вплоть до 263, так как 20 = 1 на первой клетке). Правитель был вначале поражен скромностью просьбы, а потом... оказалось, что она невыполнима. Чтобы заполнить все клетки доски, потребовалось бы по весу около 100 млрд т зерна, т. е. многократный мировой урожай!
В живой природе такая способность к быстрому возрастанию, автокаталптическому размножению была не только отмечена около тысячи лет назад, но уже и сформулирована в начало XIII в. (1202 г.) в виде математической модели итальянским математиком Леонардо Пизанским (он был родом из той самой Пизы, где находится знаменитая падающая, но до сих пор не упавшая башня). Этот Леонардо более известен под именем Фибопаччи. Рассуждая о числе потомков, появляющихся в последовательных поколениях, от единственной пары кроликов, Леонардо получил растущую последовательность чисел: 1; 1; 2; 3; 5; 8; 13; 21 и т.д., где каждое последующее число — сумма двух предыдущих (это и есть знаменитый ряд чисел Фибоначчи). Таким образом, численность популяции очень резко возрастает с числом поколений; так, если к десятому поколению общее число потомков приблизится к 100 особям, то уже к шестнадцатому будет свыше 1500 особей.
Великий классификатор и систематик живой природы К. Линней в середине XVIII в. вычислил, что «если бы однолетнее растение производило только пару семян („Нет ни одного растения, которое было бы так неплодовито”, — отмечает Ч. Дарвин, который приводит эти вычисления Линнея), его потомки на следующий год снова по паре семян и т.д., то в 20 лет было бы миллион растений» (цит. по: [Дарвин, 1912, с. 56]).
Сам Ч. Дарвин находился под глубоким впечатлением от высокой скорости размножения живых организмов в геометрической прогрессии. «...Все органические вещества естественно возрастают в такой прогрессии, что, если бы они не погибали, земля вскоре была бы покрыта потомством одной единственной пары... Даже медленно размножающийся человек удваивает численность в 25 лет, и по этой пропорции менее чем в тысячу лет буквально не осталось бы для его потомства места, где можно было бы поставить ногу» [Там же, с. 56].
Наиболее впечатляющие цифры можно привести из кинетики роста микроорганизмов. Если взять среднюю массу бактерии равной 6 · 10—13 г и сравнить ее с массой Земли, равной 6 · 1027 г, то получим величину, в 1040 раз меньшую. Однако прирост биомассы бактерий в такое число раз при размножении простым делением можно получить примерно за 130 последовательных поколений (1040 = 2130). Если длительность поколения принять за 20 мин (это — средние данные для кишечной палочки на богатой среде), то получим необходимое время — несколько менее 2 сут. Таким образом, при хороших условиях размножения потомки одной бактериальной клетки способны в течение всего лишь 2 сут создать биомассу по величине, равную массе всей планеты, а по объему превышающую ее в пять раз с лишним. Поистине огромен потенциал живой приводы к размножению!
Мы уже говорили, что биохимическую основу, обеспечивающую высокие скорости роста и развития клеток, организмов, популяций, составляют хорошо сбалансированные системы реакций воспроизводства (автокатализа) макромолекул, прежде всего нуклеиновых кислот и ферментов. Например, эффективность иона железа как катализатора реакции разложения перекиси водорода возрастает на 9—10 порядков (т. е. в миллиарды раз), если он в составе молекулы порфирина входит в фермент — каталазу. Примерно такие же, т. е. в сотни миллионов и миллиарды раз большие по сравнению со скоростями реакций в неживой природе, скорости процессов, протекающих на ферментативной основе в живых системах.
В живых организмах «... скорость химической реакции почти всегда достигает предельного значения, которое определяется законами физики. Во всех случаях обнаруживают оптимальное соотношение тенденции „как можно быстрее” и „настолько точно, как это нужно”»,— пишут в книге «Игра жизни» известный исследователь физико-химических основ эволюции лауреат Нобелевской премии Манфред Эйген и его соавтор Роберт Винклер [М. 1979, с. 96].
Однако возможности неограниченного размножения не могут реализоваться: и для популяционного уровня, и выше «взрывы» численностей имеют место гораздо реже, чем поддержание стационарных уровней, и бывают кратковременными. Живая природа упирается в ограничение косного окружения. «Напор жизни» (по выражению В. И. Вернадского), нехватка вещества загоняют ее в условия сильного лимитирования. (Не зря существует выражение: «Голод правит миром».)
Живая система использует все, что может взять у среды. Главная черта, характеризующая «хитрость» живой природы, ее «умение» справляться с лимитированием по веществу при постоянной накачке потоком солнечной энергии,— это повсеместное развитие циклов вещества. Рассмотрим некоторые примеры циклов вещества в живой природе — от молекулярных структур клетки до биосферы в целом — на разных уровнях ее организации.
5.3. Живые циклы: от электронного до биосферного
«„Wheels within wheels within wheels”— циклы, включающие циклы, которые, в свою очередь, включают циклы,— так определяется биологический процесс в целом»,— пишут в книге «Наука о живом» известные биологи П. и Дж. Медавары [М., 1983]. Мы начнем описание этих циклов с самого основного — энергодающего.
Представим простую схему протонного (электронного) цикла (рис. 6). С помощью белков клетка способна использовать энергию света, перенося водородные ионы и, соответственно, электроны через мембрану. Таким способом создается разность потенциалов, электрохимический градиент. А он и будет движущей силой процесса, в данном случае химического. Его величина, порядка 0,25 В, вполне достаточна, чтобы компенсировать потери энергии при синтезе АТФ из АДФ. На каждую синтезированную молекулу АТФ, этой энергетической валюты, «расходуется» два Н+, т. е. два протона обратно возвращаются через мембрану. Так работает цикл. Энергетические ресурсы клетки могут быть разнообразными для авто- и гетеротрофов, в последнее время много работают с бактериородопсином как источником электрохимического потенциала. Этот светочувствительный белок наряду с широко известным хлорофиллом также можно назвать генератором электрического тока. У эукариотных гетеротрофных клеток энергодающим источником является глюкоза, а образование АТФ связано с мембранами митохондрий. Последние образно называют «электростанциями клетки». Теперь, в связи с пониманием протонного цикла, оказалось, что это вовсе не метафора. А в целом, по энергетике, автотрофную клетку можно назвать «фотоэлектрическим элементом», а гетеротрофную—«электрохимическим элементом» на основе циклов.

Рис. 6. Схема энергодающего протонного цикла.
Одной из главных особенностей живого является наличие специфических белковых катализаторов — ферментов. Работа этих катализаторов также циклична. Существовал специальный термин—«число оборота» фермента, т. е. сколько молекул субстрата «перерабатывает» одна молекула фермента в единицу времени. (Теперь эту характеристику называют молекулярной активностью). И этот показатель может быть очень большим, достигая, например, тысячи или даже миллиона в минуту. Миллион операций в минуту! — Такова «скорострельность» фермента, так работает эта сложная машина, циклически меняющая свою пространственную конформацию с огромной скоростью.
Основа деятельности зеленых растений — фотосинтез, и осуществляется он наверху, в листьях, содержащих хлорофилл. Газообмен с окружающей средой при фотосинтезе и дыхании растения осуществляется через межклеточные пространства — устьица, величина которых может регулироваться. А необходимая для фотосинтеза вода подается по специальной транспортной системе, которая называется ксилемой. Иногда эту систему тонких трубочек называют водопроводной. В воде растворены минеральные соли, необходимые для многочисленных биосинтезов всей органики растения. Все это поступает из корней под влиянием корневого давления и транспирации воды в листьях. Но корни тоже «хотят жить», а они гетеротрофны, и вот по другой системе трубочек — флоэме к ним устремляется источник энергии — образовавшийся в листьях сахар. Движущая физико-химическая природа этого потока, иногда очень быстрого (до 10 м/ч), не совсем ясна. Ксилема и флоэма и образуют циркуляционную систему растения, по которой проходит огромное количество вещества. Например, на фотосинтез используется 1–2% поступающей от корней воды, а остальное количество, в 50—100 раз больше, уходит при транспирации. Прямо или косвенно, через атмосферные процессы (осадки, конденсация, потоки) эта вода опять возвращается к корням. Так замыкается ее цикл.
Более совершенная и более замкнутая циркуляционная система имеется у животных, особенно у высших, включая человека. Здесь работают настоящие насосы, перекачивающие энергетическое топливо, кислород, питательные элементы и выводящие отходы метаболизма. Главным носителем является кровь. В легких она обогащается кислородом, который запасает в эритроцитах (дискообразных форменных элементах) и выделяет CO2 как результат окисления глюкозы в энергодающих процессах. В кишечнике она получает питательные вещества, образующиеся в результате метаболического усвоения, «переваривания» пищи. Самый главный мотор организма животного — это его сердце. Выталкивая через артерии обогащенную кислородом и питанием кровь, оно через систему ветвлений и капилляров доводит ее до каждой работающей клетки, где путем диффузии происходит обмен принесенных веществ на метаболические отходы. Венозная кровь, замыкающая цикл, прокачивается через выделительные системы, где освобождается от метаболитов и вновь приходит по малому кругу к легким. Удивительна работа сердца как насоса. За минуту у человека, находящегося в состоянии покоя, оно перекачивает около 5 л крови, а за час это составит уже по весу примерно 4–5 весов взрослого человека. За 70 лет жизни сердце человека в среднем перекачивает свыше 150 млн л крови, что более чем в 2 млн раз превышает вес человека. Такова работа этой циркуляционной системы, обеспечивающей жизнь организма.
Рассмотрим работу планетарного биотического круговорота. Очень конкретно и точно выразил свою точку зрения на «энергетический цикл жизни» один из выдающихся биохимиков нашего времени А. Сент-Дьёрдьи [1964, с. 30]: «Электроны сначала поднимаются на более высокий энергетический уровень фотонами (квантами света), а затем в живых системах падают на свой основной уровень, отдавая при этом свою избыточную энергию, которая приводит в действие машину жизни».
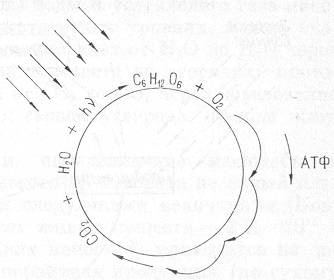
Рис. 7. Схема основного энергодающего цикла в биотическом круговороте. Параллельные стрелки показывают поток энергии Солнца.
Поток возбужденных, богатых энергией электронов, или электронный каскад, можно уподобить ряду водопадов: каждый водопад приводит в движение циклы вещества, вращает «турбины» ферментативных реакций, в ходе которых энергия электронов связывается в биологически полезной форме — в виде энергии макроэргических соединений, например всем известного аденозинтрифосфата, или АТФ, которую часто называют «энергетической валютой жизни».
И в данном случае, как и при описании других типов круговоротов, очевидна необходимость циклов вещества для длительного использования «вечного» потока энергии от Солнца.
Основой для расчета циклов главных элементов, прежде всего углерода, кислорода и водорода, составляющих 9/10 массы всех живых тел, может служить уравнение реакции фотосинтеза (или дыхания), представленное на рис.7. Для прямого протекания этой главной для жизни реакции необходима энергия солнечного света (Q=120 ккал/моль), а обратная реакция — дыхание, связанная с потреблением глюкозы, осуществляется за счет использования энергии, запасенной в углеводах.
Для замыкания круговорота достаточно иметь всего два звена: фотосинтезирующее, автотрофное, которое производит органические соединения (растения суши и водоросли), и звено потребителей этой энергии, гетеротрофное (бактерии). Работа бактерий сопровождается освобождением элементов неорганического питания для последующего использования автотрофным звеном и т. д. (рис. 8).

Рис. 8. Схема биотического круговорота и потоков энергии через основные звенья упрощенной экосистемы.
Сплошные линии — потоки вещества; штриховые — передача энергии; стрелки, отходящие от круга, указывают потери энергии в каждом звене, т. е. отток энергии в космос.
Итак, растения-продуценты, фиксирующие и аккумулирующие солнечную энергию в своей биомассе, могут как потребляться травоядными животными, так и, отмирая, перерабатываться бактериями и грибами в запас неорганических биогенных элементов в почве и воде. При этом, казалось бы, что травоядные (хищники 1-го рода) создают новую биомассу. Но надо помнить, что для создания ее они расходуют примерно в 10 раз больше живого вещества с предыдущего уровня продуцентов. Соответственно и теряется энергия. Следующий трофический уровень — плотоядные (хищники 2-го рода), потребляя травоядных, также рассеивают энергию, но они уже могут использовать до 30% от потребленной энергии. Трофических уровней потребителей может быть несколько, обычно не более 4–6, из-за потерь энергии на каждом из них. Кстати, о человеке, с этой точки зрения, можно говорить как о хищнике 1-го, 2-го и последующих родов.
В конечном счете все органические молекулы расщепляются до неорганических соединений, пополняя запас биогенных элементов, но они опять расходуются на синтез фитомассы, самой большой массы органики на нашей планете.
Поскольку молекулы воды и углекислого газа находятся на низких энергетических уровнях, можно сказать, что цикл превращений идет от H2O до H2O через скачок с помощью квантов света до «горячих» протонов и электронов, или от CO2 до CO2 через образование энергетически богатых связей углерода, прежде всего глюкозы.
Общие показатели, определяющие масштабы и энергоемкость биотического круговорота на нашей планете, характеризуются следующими величинами [Ковда, 1975]: биомасса всех живых существ — 2,42·1012 т (по сухому весу), из них менее 1% приходится на долю мирового океана; первичная продукция (по сухому веществу) — 2,32·1011 т/год, из них 1,72·1011 т/год — продукция континентов; 0,6·1011 т/год — продукция Мирового океана. Из всей приходящей на поверхность Земли солнечной энергии на фотосинтез расходуется менее 0,1% (на суше несколько выше 0,1%, на поверхности Мирового океана примерно 0,04% из-за низкопродуктивных центральных частей, соответствующих пустыням суши).
Согласно А. А. Ничипоровичу [1967], годовая продукция фотосинтеза на Земле оценивается в 46·109 т органического углерода. По уравнению реакции фотосинтеза для производства этого количества углерода требуется, чтобы 170·109 т углекислоты связывались с 68·109 т воды, в результате чего усваиваются 44·1016 ккал ФАР и образуются 123·109 т кислорода и 115·109 т сухого органического вещества.
Не будем останавливаться на видовом разнообразии организмов (насчитывают около 2 млн видов растений и животных, не говоря уже о бактериях, среди которых известна едва ли не десятая часть видов). Отметим, что весь этот калейдоскоп, составляющий живую оболочку планеты — биосферу, занимает всю тропосферу и нижнюю часть стратосферы (до озонового экрана, примерно 30–40 км), а снизу ограничен отложениями на дне океанов и глубиной проникновения подземных вод вместе с микроорганизмами (до глубины порядка 10 км).
Скорость оборота всего живого вещества достигает примерно 10% в год, этот же показатель характерен и для продуцентов, точнее, зеленых растений суши, составляющих более 99% общей биомассы. Для деструкторов, едва ли достигающих 1 % от общей биомассы планеты и вынужденных перерабатывать всю массу органического вещества (в 10 раз превышающую их собственный вес), скорости оборота соответственно во много раз выше.

Рис. 9. Зависимость максимальной продуктивности от радиационного баланса для территории СССР.
Максимальная продуктивность растительного покрова определяется радиационным балансом (рис. 9). Естественно, что для более точного описания характеристик круговорота, в особенности его продуктивности и интенсивности, необходимо учитывать и конкретные условия развития экосистем данной зоны, и условия обеспеченности водой, и результирующие температуры, но для нас с вами в данном случае важно подчеркнуть, что условие обеспечения потоком энергии является первичным и наиважнейшим.
Давая краткий обзор характеристик глобального биотического круговорота, еще раз выделим общепланетарное значение живого вещества, которому придавал огромное значение В. И. Вернадский. Проиллюстрировать это можно с помощью несложных расчетов. По сухому веществу наличная биомасса Земли, имея вес 2,42·1012 т, составляет лишь 0,00001 % от веса земной коры (2·1019). Однако принимая 10 лет за период ее обновления и полагая, что продукция по объему мало изменилась за последний миллиард лет, можно получить суммарную величину массы вещества, использованного жизнью: 2,0·1012 × 10-1 × 1·109 т = 2·1020 т. А это уже в 10 раз превышает вес коры Земли. Мы можем утверждать, что атомы, составляющие наши тела, побывали и в древних бактериях, и в динозаврах, и в мамонтах. Положение о ведущей роли живого вещества в биосфере предложено назвать законом Вернадского. В одной из наиболее полных формулировок он гласит: «Миграция химических элементов на земной поверхности и в биосфере в целом осуществляется или при непосредственном участии живого вещества (биогенная миграция), или же она протекает в среде, геохимические особенности которой (кислород, углекислый газ, сероводород и т. д.) преимущественно обусловлены живым веществом, как тем, которое в настоящее время населяет данную биокосную систему, так и тем, которое действовало на Земле в течение всей геологической истории» [Перельман, 1977, с. 128].
Приведем некоторые показатели, характеризующие «экономию биосферы», или степень замыкания круговорота по одному из важнейших элементов — углероду за последний миллиард лет. Годичная продуктивность биосферы по углероду достигает 9·1010 т. За миллиард лет — 9·1019 т. Запасы мертвого органического вещества биогенного происхождения, выпавшего из круговорота в толщах осадочных пород, т. е. ушедших из цикла древних биосфер, составляют по различным оценкам от 4·1015 до 15·1015 т (в пересчете по углероду), за среднее можно принять цифру 9·1015 т. Отсюда можно оценить степень несовершенства круговорота как отношение потерянного углерода ко всему задействованному:

или в процентах: К = 0,01% (соответственно степень замкнутости определяется числом с четырьмя девятками — 99,99%). А это значит, что в среднем каждый атом углерода участвовал в цикле примерно десять тысяч (!) раз, прежде чем был потерян для жизни в захоронениях литосферы. Так удивительно экономна наша биосфера как единая функционирующая единица, так совершенен наш глобальный биотический круговорот. А ведь углерод не является основным лимитирующим биогенным элементом, поэтому цифры по азоту и фосфору должны быть еще более впечатляющими.
Более того, и те органические остатки, которые захоронены в седиментах литосферы, если говорить о геологическом времени, т. е. учитывать геохимические циклы, находятся там не вечно.
По образному выражению А. В. Лапо, автора интересной книги «Следы былых биосфер» [М., 1979, с. 111], консервация биогенных веществ в экосистемах — «явление сугубо временное, нечто вроде хранения багажа в автоматической камере на вокзале. Нормально жизнь забирает обратно свой багаж (то бить небиогенное вещество). Нужны какие-то исключительные обстоятельства, чтобы ячейка камеры осталась невскрытой, а багаж — замурованным в ней». Один из наиболее ярких примеров использования захоронений такого рода — это добыча полезных ископаемых человеком гораздо более высокими темпами, чем средние скорости их образования. Недаром так высока угроза быстрого истощения ископаемых ресурсов, а с ней и возможностей нашего «паразитирования» на несовершенствах и катастрофах былых биосфер.
Заканчивая рассмотрение особенностей жизни, мы можем еще раз подчеркнуть, что возмущающий поток энергии раскручивает циклы на всех уровнях организации живого вещества.
Тем самым жизнь не останавливается, не загоняется в тупик из-за нехватки вещества, а совершенствуется, ускоряя и умощняя свои циклы. «Хочешь жить — умей вертеться» — гласит лукавая пословица. Для оценки развития круговоротов (а не в приложении к одной популяции) она теряет большую часть метафоричности и довольно точно отражает одну из главных черт биотических циклов. Действительно, в физическом круговороте произошел отбор самого энергоемкого и подвижного носителя — молекулы воды. В биотическом круговороте и в структурах живого вещества она тоже играет одну из важнейших ролей.
Это удивительное вещество пронизывает всю биосферу. В атмосфере — это испарение и громадные переносы облаков по всей планете; в гидросфере — это аккумуляция тепла океаническими водами и глобальные течения; в литосфере — это теплоноситель отопительной системы, позволяющий выводить внутренние потоки тепла. И наконец, вода — основа всего живого. Все живое вещество состоит более чем на 2/3 из воды. Например, человек за свою жизнь в среднем прокачивает воды около 75 т (а это в тысячу раз больше его веса). А главное — вода участвует в энергодающих метаболических процессах, без которых жизнь невозможна.
Тесную взаимосвязь воды и жизни много раз отмечал В. И. Вернадский, говоря, что вода и жизнь генетически связаны, а известный немецкий физиолог прошлого века Эмиль Дюбуа-Реймон называл жизнь «одушевленной водой». Можно сделать некоторые выводы.
1. Поток энергии является источником движения в любой системе.
2. Под влиянием постоянной накачки энергией в любой ограниченной системе возникают циклические перемещения вещества вплоть до сложных динамических структур. Неравновесность является характерной чертой для систем с накачкой (а не с особым свойством жизни).
3. В системах с циклами имеет место отбор наиболее подвижного и энергоемкого носителя, примером которого на нашей планете может служить вода.
4. Ведущую роль в трансформации веществ на Земле играет биотический круговорот, составляющий основу жизни.
Глава 6
Два энергетических принципа биологического развития
В особой земной оболочке, которой является биосфера, характеризуемая необратимыми процессами, жизнь будет увеличивать, а не уменьшать с ходом времени свободную энергию этой оболочки.
В. И. Вернадский
6.1. Трудности измерения эволюции
Древнейшие натурфилософы-материалисты, особенно в Греции и Индии, не могли не обращать внимания на энергетические потоки в виде тепла, огня, света, считали их материальными носителями движения. Представители экспериментальной науки, начиная с XVII в., занимаясь вопросами энергетики в технике и физике, неоднократно сталкивались с вопросами энергетики и движущих сил жизни. К примеру, одной из первых единиц мощности была в буквальном смысле «живая» единица — лошадиная сила. Говоря об истории идей, относящихся к энергетическим аспектам жизни, В. И. Вернадский отмечал, что целый ряд мыслителей прошлого, философов и естествоиспытателей, независимо приходили почти к одинаковым представлениям.
Сам В. И. Вернадский придавал большое значение энергетической функции вещества. Второй биогеохимический принцип Вернадского непосредственно связывает вопросы направленной эволюции с энергетикой жизни. По В. И. Вернадскому [1960], преимущества в ходе эволюции получают те организмы, которые приобрели способность усваивать новые формы энергии или «научились» полнее использовать химическую энергию, запасенную в других организмах.
Наибольшее развитие в приложении к анализу живых систем энергетический подход получил в работах исследователей экологического направления. Одной из первых попыток можно считать гипотезу А. Лотки о том, что эволюция экосистемы направлена на увеличение суммарного потока энергии через нее, а в стационарном состоянии этот поток достигает максимума. Эта закономерность проявляется в широко известной (и в ряде случаев поражающей воображение) способности живого к распространению, к развитию, во «всюдности» жизни согласно первому биогеохимическому принципу Вернадского.
Не останавливаясь на конкретных вариантах трактовки, можно сделать вывод о том, что к настоящему времени основные трудности развития и применения этих принципов заключаются не в недостаточности идей и теоретических обобщений. Главная трудность состоит в сложностях строгой экспериментальной проверки этих принципов и в недостаточной точности, а иногда и невозможности измерений на природных биологических системах. Таким образом, не выполняется одно из основных методологических требований естественных наук: экспериментальная проверяемость результатов, при которой теория должна опираться на эксперимент. Это условие очень важно для обобщений в биологии, где большое разнообразие явлений может приводить к неменьшему разнообразию их истолкований; поэтому здесь тем более справедливо замечание Ю. Одума [1975, с. 149]: «Утвердиться в науке и оказаться полезным на практике может лишь тот принцип, который поддается не только качественному, но и количественному анализу».
Таким образом, необходимость строгих дискриминационных экспериментов очевидна, а их отсутствие является «узким местом», сдерживающим накопление количественного знания. Нужно иметь хорошо воспроизводимые стационарные состояния систем н переходы между ними с измерением потоков энергии и вещества.
Однако трудности количественной экспериментальной (тем более натурной) работы с экосистемами или их отдельными звеньями невообразимо велики. Трудно себе представить стационарные состояния таких систем со строгим измерением и контролем параметров среды и живого вещества, еще более трудно провести надежные измерения эволюционных переходов в них. Поистине не от «хорошей жизни» измерения трансформации энергии проводились на отдельных изолированных организмах, хотя узость и ограниченность такого подхода хорошо осознавал и резко критиковал В. И. Вернадский [1978, с. 141] еще в начале нашего века: «Благодаря тому, что в огромной массе проблем биологии зависимость организма от внешней среды исчезает из поля нашего зрения, вместо живого организма изучается в ней искусственно отделенное от внешней среды тело, не отвечающее реальному объекту Природы».
Таким образом, задача кажется неразрешимой и трудно представить, как выйти из этого «порочного круга»: в экологии можно правильно поставить задачу, но нельзя провести строгих измерений (например, только для одного эволюционного эксперимента могут понадобиться десятки поколений исследователей), а в возможных (но далеко не простых!) экспериментах по изучению роста и развития отдельных организмов теряется основа для корректной постановки задачи.
И все же дело не столь безнадежно, если обратиться к объему, удивительным образом сочетающему в себе достоинства обоих упомянутых подходов, а именно к популяциям микроорганизмов. Вновь процитируем В. И. Вернадского [1978, с. 141]: «Во всех вопросах микробиологии мы для получения нужных результатов, для изучения изменения среды под влиянием организмов идем одним путем. Мы изучаем не влияние отдельного организма, а проявление массового воздействия их совокупности». Представления В. И. Вернадского о живом веществе как о совокупности живых организмов, производящих на земной поверхности химическую работу и являющихся носителями свободной энергии, идейно тесно смыкаются с представлениями об энергетических потоках, развиваемых в данной работе.
Итак, что же может предложить современная микробиология (вернее, ее кинетический и популяционный разделы) для решения поставленных задач?
Со второй половины нашего столетия начали активно развиваться методы непрерывного культивирования микроорганизмов, что привело к разработке уникальных экспериментальных моделей популяционной динамики в открытых системах. Наконец-то появилась экспериментальная возможность на живых надорганизменных системах получать стационарные состояния, измерять переходы между ними и даже изучать эволюцию этих состоянии. Быстрое развитие техники непрерывного культивирования усовершенствовало разнообразные методы экспериментального исследования: при низких и высоких скоростях роста популяций; с различной степенью и характером ограничения роста (лимитирование по различным веществам, ингибирование метаболитами и внешними соединениями типа загрязняющих веществ); рост без внешнего лимитирования; в пространственно неоднородных и гомогенных средах; в монокультурах и различного рода ассоциациях одного или нескольких трофических уровней вплоть до малых экосистем с разными уровнями замыкания по веществу, т. е. круговорота; в стабилизированных, переходных или меняющихся по какому-либо заданному закону условиях среды. При этом целый ряд количественных показателей не только находится под контролем, но также может автоматически измеряться и регистрироваться [Печуркин, 1978].
Для иллюстрации действия энергетических принципов, обсуждаемых в этой главе, мы будем использовать некоторые данные, полученные в экспериментах с микробными популяциями и ассоциациями в открытых системах [Печуркин, 1982].
В этой главе мы обсудим два энергетических принципа: экстенсивного и интенсивного развития жизни. Первый связан с захватом энергии биологическими системами, а второй — с эффективностью ее использования.
6.2. Энергетический принцип экстенсивного развития (ЭПЭР)
Обозначим через Нпад поток падающей энергии, т. е. поток энергии, который потенциально может быть использован некоторой биологической системой. Примерами Нпад являются поток солнечной энергии для автотрофов, количество органической пищи (в калориях), доступной для использования гетеротрофными звеньями в единицу времени. Не весь падающий поток энергии может быть захвачен и использован системой:
| Нисп = Нпад — Ннеисп . | (1) |
Сам поток энергии, захваченной системой, складывается из двух частей: доли, связанной с запасением энергии в биомассе — В, и потерь на организацию, поддержание и активность, т. е. расход энергии системой, — R:
| Нисп = µ В — R . | (2) |
где µ — показатель прироста биомассы (или обновления в стационарном состоянии открытой системы).
Объединив (1) и (2), получим общее соотношение для потоков энергии в систему и ее трансформации в этой системе:
| Нпад — неисп = Нисп = µ В + R | (3) |
Дадим формулировку энергетического принципа: в процессах развития надорганизменных систем (эволюции, экологических сукцессиях и перестройках) использованный биологической системой поток энергии Нисп возрастает, достигая локальных максимальных значений в стационарных состояниях.
Подчеркнем еще раз, что в формулировке данного принципа речь идет о стационарных состояниях, которые достигаются в процессе развития открытых биологических систем популяционного и более высоких уровней структурной организации, а также их эволюции.
Представим схематически наиболее предпочтительный тип изменения потоков энергии во времени, использованных системой (рис. 10,а). Здесь показаны и рост Нпад, т. е. рост захваченной энергии, и уменьшение Ннеисп, т. е. снижение потерь. Штриховкой обозначено, что при этом происходит увеличение рассеяния энергии внутри системы Д с уменьшением трат на образование биомассы (это относится уже ко второму энергетическому принципу и будет обсуждаться позже). Естественно, что монотонное линейное увеличение Нпад или снижение Ннеисп не может иметь места в реальных системах. Гораздо типичнее скачкообразные переходы, связанные с качественными изменения в самой системе (рис. 10,б—г). Условия роста (или по крайней мере не убывания) потока использованной энергии Нисп остаются для каждого момента времени.

6.3. Экспериментальные эволюционные машины (ЭЭМ)
В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.
Суть непрерывного процесса заключается в постоянной подаче питания в зону развития популяции и одновременном отборе избыточной части из рабочего пространства. Природная популяция, таким образом, осуществляя непрерывный обмен веществом с внешней средой и получая энергию извне, развивается в открытой системе, что является главной чертой ее динамики. В лабораторных условиях такой обмен осуществляется с помощью непрерывного потока питающей среды в рабочий объем ферментера и соответствующего оттока культуральной жидкости.
В классических периодических процессах нелимитированный рост популяции отмечается лишь во время экспоненциальной фазы, которая сменяется фазой замедления, т. е. торможения роста. При непрерывном культивировании можно застабилизировать рост популяции в любой точке на восходящей ветви S-образного роста популяции, в том числе и в экспоненциальной фазе. Для этого, как уже говорилось, необходимо непрерывно подавать свежую питательную среду для популяции и удалять избыточную часть популяции из рабочего объема. Для поддержания плотности популяции в заданной точке фазы нелимитированного роста применяются различные способы управления скоростью протока. Основное их свойство — наличие обратной связи между приростом концентрации биомассы и удалением части популяции из ферментера. Эти величины должны быть равными и это равенство, а с ним и концентрация биомассы — поддерживаются с помощью автоматических измеряющих и следящих устройств. Среди таких способов первым был турбидостат, в котором концентрация клеток поддерживается на определенном уровне за счет регулирования оптической плотности культуры. Применение его ограничено работой с оптически однородными средами.
Для процессов культивирования, в которых имеется прямая связь между приростом биомассы и изменением рН-культуры (например, при потреблении физиологически кислого источника азота), разработан и используется рН-статный способ управления скоростью протока. При этом скорость протока с помощью автоматических устройств уравнивается со скоростью изменения рН растущей популяцией, а следовательно, и со скоростью роста, что обеспечивает поддержание концентрации биомассы на заданном уровне.

Рис. 11. Блок-схема процессов непрерывного роста микробных популяций.
1 — культиватор; 2 — датчик-измеритель; 3 — самописец с регулятором; 4 — система управления насосами-дозаторами; 5 — система дозирования; 6 — сосуд с питательной средой; 7 — сборник урожая [Печуркин, 1982]. (Штриховой линией отмечена блок-схема хемостата.)
Наиболее просто и надежно система поддержания концентрации биомассы (клеток) и управления скоростью протока работает в импульсном режиме (рис. 11). Основу схемы составляет датчик-измеритель. Для турбидостата это — датчик оптической плотности, для рН-стата — электродная пара рН. Измеренное значение параметра в виде электрического сигнала подается на схему управления протоком (блоки 3 и 4). Если величина параметра достигает заданной, то в системе управления вырабатывается управляющий сигнал, который передается в систему дозирования. Происходит долив порции свежей питательной среды и одновременно такой же по объему слив части суспензии из ферментера. Разбавление культуры свежей питательной средой приводит к снижению величины управляющего параметра, и проток выключается. Продолжающийся рост популяции снова вызывает возрастание величины управляющего параметра до уровня срабатывания следящей системы и т. д. На самописце 3 вырисовывается «пила», частота и размер зубцов которой определяются особенностями схемы управления и скоростью роста популяции.
В отличие от описанного выше турбидостатного культивирования в нелимитированных условиях, когда для поддержания устойчивого состояния требуются регуляторы, хемостат характеризуется обязательным внешним ограничением роста. Такое ограничение является устойчивым регулятором стационарности параметров среды и популяции. Как правило, это — лимитирование недостатком одного из компонентов питания или ингибирование роста продуктами метаболизма. Основа хемостата — подача питательной среды с постоянной скоростью протока. Популяция, утилизируя субстрат, «загоняет» себя в условия лимитирования по одному из компонентов среды, потребляя его до низких остаточных значений. Хемостат получил широкое распространение, так как простота аппаратурного оформления сочетается в нем с широкими экспериментальными возможностями.
Для нас здесь необходимо подчеркнуть два важных свойства турбидо- и хемостата. Первый соответствует росту популяций в нелимитированных условиях, что в природе может встречаться на ранних фазах экологической сукцессии, например при заселении новой экологической ниши. Второй — с глубоким лимитированием роста — является аналогом большинства природных ситуаций, где повсеместно встречается ограничение роста.
С точки зрения функционирования открытых систем хемостат и турбидостат — это термодинамические системы, способные находиться в устойчивых стационарных состояниях. Причем хемостат соответствует случаю постоянных потоков, а турбидостат — случаю постоянной организации (или постоянных реакционных сил). Других условий стационарности в открытых системах просто не бывает. Таким образом, в руках экспериментаторов имеются открытые системы двух основных типов развития (и для экологии, и для термодинамики). Если в таких системах будут происходить эволюционные изменения, переход от одного стационарного состояния к другому в результате изменения качественных свойств систем (например, в результате процессов мутирования и отбора), то главные характеристики этих сукцессионных перестроек, или шагов эволюции, можно измерить, не теряя общности подхода с точки зрения как биологии, так и физики. Естественно, что основу такого единства составляет энергетический подход.
Накоплено достаточно много данных по работе «эволюционных машин» обоих типов. Кинетика накопления активных мутантов, вытесняющих исходную форму из ферментера в результате процессов автоселекции, выглядит очень разнообразной (см. [Печуркин, 1978, 1982]). Здесь мы только перечислим наиболее интересные варианты.
В турбидостате это — мутанты с повышенной максимальной удельной скоростью роста; более «резистентные» мутанты, т. е. способные расти с повышенной скоростью в условиях ингибирования роста. Для хемостата характерны мутанты с повышенным сродством к лимитирующему субстрату; более экономичные формы; более жизнеспособные мутанты, т. е. с пониженной скоростью отмирания, и т. д.
В турбидостате микроэволюционный переход сопровождается увеличением скорости протока, а следовательно, и Нпад (в соответствии с рис. 10, б); в хемостате процесс автоселекции приводит к более полному использованию лимитирующего субстрата, т. е. к снижению Ннеисп (в соответствии с рис. 10, в). И для обоих случаев характерно увеличение потока использованной энергии, т. е. Нисп. Таким образом, энергетические принципы однозначно указывают направление эволюционных перестроек, несмотря на кажущееся внешнее разнообразие эволюционных переходов.
Оказалось, что в хемостате и турбидостате можно хорошо экспериментировать с более сложными взаимодействиями, например изучать сосуществование видов, а также трофические отношения типа хищник — жертва, паразит — хозяин. При длительном развитии таких систем в проточных условиях наблюдалась хорошо выраженная тенденция к стабилизации их развития и коэволюции, причем Нисп возрастал.
Эволюционные эксперименты по типу турбидостата были проведены в лаборатории доктора С. Спигелмана (США) не с живыми клетками, а с макромолекуляр-ными системами, способными к воспроизведению. Действию отбора в ряду последовательных разведений (переносов в пробирки с приготовленной для размножения средой) подвергался носитель наследственности — молекула РНК бактериофага Qβ или ее укороченные варианты. Репликация РНК осуществлялась с помощью фермента РНК-полимеразы, выделенной из зараженной фагом культуры кишечной палочки.
Остановимся подробнее на одном из экспериментов по накоплению и идентификации мутантов, резистентных к этидийбромиду. В качестве исходной формы в опыт бралась малая реплицирующаяся молекула РНК (MDV), состоящая из 218 нуклеотидов, расположенных в известной последовательности. Реакционная смесь в каждой из 25 пробирок содержала 15 мкмоль ингибитора — этидийбромида, который способен обратимо снижать скорость удлинения цепи молекулы. В каждой пробирке, начиная с первой, реакция инкубировалась в течение 10 мин, а для переноса в последующую пробирку использовалась малая доля (10-5) материала предыдущей. В данных условиях эксперимента происходило экспоненциальное размножение РНК в каждой пробирке, так что действие отбора было направлено на увеличение скорости репликации, которая и возросла почти в 2 раза.
Изучение последовательности нуклеотидов в мутантной РНК показало, что три нуклеотида отличаются от нуклеотидов «дикого» (исходного) типа. Каждая из трех единичных мутаций происходила в разное время. Химическая основа резистентности, по-видимому, заключается в элиминации сайтов, связывающих этидий, благодаря специфическим изменениям в последовательности нуклеотидов. Это выражалось в том, что мутантная РНК связывала меньше этидия, чем молекулы «дикого» типа.
Эти эксперименты стимулировали теоретические исследования уже упоминавшегося нами М. Эйгена [1973] по эволюции самовоспроизводящихся структур типа гиперциклов замкнутых цепочек из ферментов и нуклеиновых кислот. Он рассматривал конкуренцию таких единиц в открытых системах при двух типах селекционных ограничений: постоянные потоки (аналог хемостата) и постоянная организация (аналог турбидостата). Близость его результатов к расчетам по кинетике эволюционных переходов в микробных популяциях оказалась удивительной. Если же дополнить его расчеты, введя поток богатых энергией мономеров, которые служат источником энергии и вещества для синтеза исследуемых полимеров, то действие энергетического принципа экстенсивного развития становится совершенно очевидным для обоих случаев селекционных ограничений [Печуркин, 1982]. Таким образом, и на неживых системах с автокатализом и внешним ограничением «работа» энергетических принципов может быть достаточно легко продемонстрирована.
К сожалению, получить данные со строгими измерениями кинетики и эволюции сложных экологических систем гораздо труднее. Трудно замерять и динамику потоков энергии. Однако некоторые результаты экологических сукцессий и эволюционных перестроек достаточно хорошо иллюстрируют действие энергетических принципов.
Мы уже говорили, что энергетические принципы и биогеохимические принципы Вернадского очень хорошо соответствуют друг другу. «Всюдность» жизни определяется «всюдностью» потока энергии, и можно только удивляться подгонке локальных круговоротов вещества к захвату солнечной энергии. В этом разделе мы рассмотрим ряд примеров, иллюстрирующих действие энергетических принципов в трех основных звеньях круговорота: автотрофах-продуцентах, гетеротрофах-консументах и разлагателях-редуцентах.
Общеизвестно, что при изменении (даже очень резком) условий среды может произойти практически полная смена видов, особенно доминирующих в экосистеме, но функция круговорота страдает менее всего. Например, при сукцессионных перестройках в экосистемах работает правило, сформулированное А. Лоткой еще в 1925 г.: максимизируется энергия в биологических системах. Сукцессия сопровождается удлинением жизненных циклов, ростом степени замкнутости круговоротов вещества и в конечном счете переходом к стабилизированному состоянию, где весь прирост биомассы в автотрофном звене компенсируется ее расходом в последующих звеньях. Такой «гомеостаз» системы может быть ярко выражен. Около 40 лет назад было отмечено, что содержание хлорофилла на 1м2 в разных сообществах бывает примерно одинаковым [Гесснер, 1949; цит. по: Одум, 1975]. То есть в целых сообществах содержание зеленого пигмента гораздо более равномерно распределено, чем в отдельных растениях или их частях. В сложившихся сообществах самые разнообразные растения — старые, молодые, освещенные, затененные, многолетние, однолетние — представляют собой единое целое и по-своему приспособлены к захвату поступающей солнечной энергии. Растения, адаптированные к слабому свету, содержат меньше хлорофилла на единицу действующей поверхности, чем растения верхнего яруса. Степень поглощения светового потока хорошо определяется невооруженным глазом в глухом таежном лесу, в старой дубраве и в многоярусных джунглях: даже в яркий день там царит полумрак.
Содержание хлорофилла иногда предлагается считать за одну из основных продукционных характеристик природных экосистем (точнее ассимиляционное число, т. е. продуктивность на 1 г хлорофилла, выраженная в граммах кислорода за час). И для водных, и для наземных экосистем этот показатель является одним из наиболее устойчивых.
Может быть, еще ярче проявляется эта важнейшая особенность биосистем — функциональное сходство при различиях видового состава — при изучении суммарной биологической активности почв. Несмотря на явные различия в составе организмов, результирующая активность определенного горизонта, слоя или даже типа почвы остается примерно одинаковой. То же можно отнести и к дыханию, т. е. выделению углекислоты. И все это имеет место, несмотря на сложную пространственную микроочаговость почвенных ценозов. Такой микроочаг (0,05–5 мм в диаметре) иногда включает и автотрофный компонент; тогда он служит реальной природной структурой, соответствующей представлению о простейшей микробной экосистеме. Таким образом, в природной обстановке уже в пределах 3–5 мм складываются сложные сообщества, осуществляющие круговорот! И такие сообщества относительно устойчивы по функционированию.
Крупные экосистемы воды и суши с точки зрения структурной организации и наличия имеющихся видов трудно объединить по сходству. Возможно, что ни одного вида, общего для таких экосистем, и не удастся отыскать. Но с точки зрения функционирования, сукцессионной динамики и эволюции они во многом схожи. По крайней мере эволюция в пределах разных групп экосистем может количественно характеризоваться ростом эффективности использования солнечной энергии.
Из-за низкой плотности и рассеянности потока квантов солнечного излучения даже в сбалансированных ценозах используется на фотосинтез около 1% падающего потока, а в целом по биосфере — ниже 0,5%. Это кажется очень малым, но мы теперь уже знаем, как непросто живым экосистемам в условиях нехватки вещества производить его всевозможные циклы. Мы уже описывали одно из удивительнейших чудес природы — растение, которое способно на огромную высоту навстречу солнечному лучу поднимать растворы необходимых солей, чтобы добыть энергию себе и последующим звеньям, поставляющим эти соли растению в круговороте.
Зато с каждым последующим звеном коэффициент использования энергии повышается, травоядные животные используют 10—15% от их кормовой базы, а хищники — до 30%. Поскольку консументы берут около 10% энергии растений, то, как отмечает С. С. Шварц [1980], часто делается вывод о том, что не энергетические ресурсы лимитируют развитие. Однако более тонкие наблюдения говорят, что такие выводы надо использовать с осторожностью. Оказывается, энергетический баланс животных очень напряжен. Поддержание энергетического баланса может рассматриваться как основа адаптаций.
Однако нельзя отводить растениям слишком пассивную роль. Живая природа более изобретательна, чем мы склонны считать. Одним из ярких примеров этому может служить увеличение захвата энергии растениями в эволюции. Если первичный бактериальный фотосинтез был связан с энергетически низкой длинноволновой частью спектра солнечного излучения, то современные зеленые растения используют наибольший возможный поток солнечной энергии. Считающийся более древним бактериохлорофилл имеет максимум поглощения между 800 и 900 нм, где поток солнечной энергии существенно меньше.
Говоря о действии ЭПЭР в биосфере, обратимся к выводам теоретиков. В обстоятельном труде «Самоорганизация в неравновесных системах» Г. Николис и И. Пригожин [М., 1979] затрагивают аспекты эволюции экосистем. Рассматривая вопросы устойчивости системы против структурной флуктуации с новой функцией (что-то типа активного мутанта в популяции), авторы приходят к выводу, что «в качестве движущей силы эволюции следует рассматривать энергетическую диссипацию» и что «процессы эволюции приводят к усилению эксплуатации окружающей среды» (с. 456). Обсуждавшийся нами энергетический принцип достаточно полно и точно соответствует этим выводам.
6.4. Энергетический принцип интенсивного развития (ЭПИР)
Говоря об ЭПЭР, мы подчеркивали возрастание способности живой системы захватывать энергию, способности к экспансии, распространению в новые места. При этом качественных изменений энергетики организмов можно и не требовать, им достаточно за счет автокатализа как можно быстрее «наплодить» себе подобных.
Однако при длительном развитии и особенно эволюции живых систем все более существенную роль должны играть процессы, направленные на улучшение качества использования энергии. И это очевидно, так как живая система в результате автокатализа быстро попадает в условия жесточайшей нехватки вещества (об организации и развитии циклов мы неоднократно упоминали на страницах этой книги). А при быстром размножении и лимитировании по веществу потребуется и быстрое отмирание, что сопровождается потерями энергии и информации и ставит популяцию в невыгодные условия. В этом случае гораздо выгоднее структуры с более длинным циклом развития. И не зря у многоклеточных организмов доля размножающихся клеток падает от 100% на ранних стадиях до 1% и ниже во взрослом состоянии.
Увеличение длительности существования считается одним из наиболее характерных проявлений эволюционного прогресса. «Подъем энергии жизнедеятельности» и, в частности, «повышение дыхательной функции», по А. Н. Северцову, является одним из главных эволюционных изменений. При этом очень важно, чтобы траты энергии на образование самой структуры и ее содержание без выполнения других функций, типа основного обмена у животных, минимизировались (или по крайней мере возрастали медленнее общих трат).
Исходя из принципа оптимальной структуры [Розен, 1969], требуется минимизация «метаболической цены», которая измеряется энергией, расходуемой организмом на образование и поддержание структуры.
Введем показатель уровня энергетического развития, характеризующий интенсивность использования энергии на единицу возобновляемой биологической структуры:
 .
.Для простых случаев (без учета возрастных структур и т. д.) между скоростью обновления биомассы µ и длительностью поколения g существует связь в виде
 .
.Здесь R — число одновременно появляющихся потомков; R = 2 при делении клеток, почковании, при последовательном появлении по одному потомку и сохранении активности родителя, т. е. в этом случае время удвоения биомассы равно длительности поколения. Для подобных случаев можно записать
 .
.В любом из вариантов энергетический принцип интенсивного развития гласит: любая живая система надорганизменного уровня развивается (эволюционирует) таким образом, что поток использованной энергии на единицу биологической структуры (за время существования этой структуры) возрастает.
Подчеркнем, что введенный показатель, отражая роль и функции структуры биосистем, остается безразмерным, и это немаловажно для сравнительного анализа (биомасса выражается через ее энергосодержание).
Остановимся подробнее на некоторых наиболее существенных выводах.
А. Рост интенсивности энергообмена (согласно ЭПИР). В биологии развития энергетический подход считается одним из наиболее важных, хотя основное внимание здесь уделялось измерениям энергозатрат отдельных особей в малоподвижном состоянии. Многочисленными исследованиями показано, что скорость теплорассеяния
(часто измеряемая косвенно по скорости дыхания  ) зависит от массы тела W:
) зависит от массы тела W:
 ,
,где а и k — константы.
«Справедливость этого уравнения установлена для большинства групп животных, от простейших до млекопитающих. Можно сказать, что эта зависимость является эмпирическим законом, справедливым для всех животных», [3отин, Зотина, 1976, с. 49]. Величина k, меньшая единицы, хорошо соответствует известному закону поверхностей Рубнера, так как метаболизм соответствует массе, числу функционирующих клеток, а теплоотдача идет с поверхности. Это означает, что с ростом размеров организма падает величина удельной теплопродукции, так как уменьшается отношение поверхность: объем. Для нашего анализа пока больший интерес представляют данные о том, что в процессе эволюции, а не индивидуального развития коэффициент a, характеризующий в данном уравнении интенсивность энергообмена, существенно возрастает. Согласно данным, суммированным Хеммингсеном для трех, далеко отстоящих по организации групп организмов, константы уравнения равны в среднем: для одноклеточных a1 = 0,084 кал/ч; для пойкилотермных a2 = 0,69; для гомойотермных животных a3 = 19,68 кал/ч. Следовательно, интенсивность обмена возрастает от простейших к гомойотермным более чем на два порядка, более чем в 200 раз. Поддержание постоянной температуры тела для гомойотермных животных обходится примерно в 30 раз дороже по тратам на обмен, по сравнению с пойкилотермными того же размера. Казалось бы, огромная расточительность! Однако преимущества такого ароморфоза позволили гомойотермным животным занять места, недоступные для пойкилотермов. Вспомним белых медведей и песцов в Арктике или пингвинов в Антарктике; очень впечатляют с этой точки зрения сезонные миграции птиц на многие тысячи километров. Пример с птицами особенно наглядно показывает резко возросшие энергетические возможности гомойотермных животных.
К сожалению, данных по полному энергетическому обмену организмов, тем более для популяционпого или экосистемвого уровня, явно недостаточно. Приведенные выше результаты относятся главным образом к основному энергетическому обмену, который составляет лишь часть полного. Хотя наиболее вероятно их параллельное возрастание, так как за каждое новое «изобретение» организму необходимо расплачиваться прежде всего дополнительным расходом энергии. Но в целом возрастание активного обмена и есть итоговая мера прогресса.
Особенно наглядны расхождения обменов для человеческой популяции, вовлекающей в свою среду дополнительные энергетические источники. Если по уровню основного обмена человек занимает срединное положение в группе млекопитающих (согласно известной диаграмме «от мыши до слона»), то дополнительное использование и производство энергоресурсов у него в среднем в 20 раз выше. А в развитых странах и в 50–100 раз! Следовательно, согласно ЭПИР, человек в 20 раз более активен, чем млекопитающие и птицы, обладающие самыми высокими показателями энергорассеяния.
Б. Рост размеров особи и длительности поколения. В теории морфофизиологической эволюции рост продолжительности жизни и уменьшение числа потомков являются одним из показателей увеличения приспособленности животных. Для всего развития жизни в условиях нехватки вещества и постоянной накачки энергией это почти очевидно: резко снижается зависимость от лимитирования по веществу и сохраняется возможность использовать энергетические потоки.
С виду несколько противоречащим ЭПИР кажется увеличение средних размеров организмов в эволюции. Однако не следует забывать, что с увеличением размера организмов, согласно уравнению теплообмена, падает основной обмен, в то время как активный обмен возрастает. (Например, более крупные животные способны перемещаться на большие расстояния и из большего числа выбирать места расселения.)
Подчеркнем высокую напряженность обмена у мелких животных, т. е. большую долю поддерживающего метаболизма по сравнению с активным. Подсчитано, например, что маленькие грызуны расходуют до 95% своей энергии на поддержание постоянной температуры тела и основной метаболизм — тут переваривание и усвоение пищи, работа внутренних органов, мышц, нервной системы — и лишь единицы процентов на активную жизнь: миграции, поиски полового партнера, освоение новых местообитаний [Шварц, 1980]. Относительная доля поддерживающих трат у мелких млекопитающих в десятки раз больше, чем у крупных.
Несмотря на огромные относительные траты энергии, мелкие млекопитающие не имеют такой надежной терморегуляции, как крупные. (И это очень важно в эволюции для увеличения размеров.) «Мелочь» гораздо быстрее гибнет от охлаждения и перегревания, в то время как крупная живность легко переносит подобные условия (примеры распространения крупных животных к полюсам холода широко известны). Повышенная долевая активность энергетического обмена позволяет крупным гомойотермным животным освоить огромные территории, недоступные для других организмов. Это и есть биологический прогресс, по А. Н. Северцову.
Если говорить о соотношении активного и поддерживающего обменов у человека как биологического вида, то оно примерно такое же, как у других млекопитающих его размера (примерно в 2–3 раза). Однако интересно оценить мгновенные максимальные возможности человека по активному расходу энергии. Возьмем, например, рекордсмена-штангиста, который в рывке примерно за полсекунды поднимает на высоту 2 м около 200 кг. Мощность, развиваемая им, составляет около 8 кВт (или около 2 ккал/с). Основной обмен (около 1 ккал/мин) превышает это более чем в 100 раз. Однако в процессе общественного развития человека мускульная энергия заменялась энергией механизмов и машин (об этом мы будем говорить далее).
И все же у млекопитающих и птиц, даже очень мелких, активность существенно выше, чем у пойкилотермных животных. Это и является несомненным завоеванием макроэволюции, если перевести на язык работы «живого вещества»: вещество, заключенное в млекопитающих, энергетически более нагружено и активно. В целом это соответствует энергетическому биогеохимическому принципу Вернадского.
К сожалению, из-за больших трудностей надежных измерений потоков энергии через популяции животных в природных условиях, с учетом соотношения активного и пассивного обменов, практически не приводилось. В зависимости от условий существования указанное соотношение может сильно изменяться, потому-то и необходимы надежные измерения, а не просто оценочные данные, как это часто делается в экологии [Уиттекер, 1980].
И все же очевидным, несмотря на недостаток данных, выглядит увеличение показателя интенсивности развития для млекопитающих: только 1–2% от потребленной пищи используют на рост эти высокоорганизованные гомойотермные животные, принадлежащие к той же филогенетической ветви, что и «венец природы»— человек.
В. Действие естественного отбора на относительное уменьшение генома в клетке в соответствии с ЭПИР. Это требование не только не очевидно, но оно на первый взгляд абсурдно в наше время, во время расцвета молекулярной генетики, когда кажется более корректным заявлять, что жизнь — способ существования нуклеиновых кислот и чем выше их содержание, тем лучше для организма. Кроме того, известно, что геном эукариотной клетки содержит в 100–1000 раз больше генов, чем ее предшественница по эволюции, гораздо проще устроенная прокариотная клетка. И действительно, более правомерным кажется, что геном человека должен быть гораздо больше по размеру, чем геном какой-либо лягушки или рыбы, не имеющей даже строгих терморегуляционпых надстроек. Однако это не совсем так. Попробуем разобраться в несоответствиях.
1. Увеличение абсолютного размера генома организмов в макроэволюции и усложнение его организации — далеко не однозначные процессы. Новые, молодые процветающие виды далеко не всегда имеют увеличенный геном; гораздо более частым бывает существование «на задворках» малочисленного полуископаемого вида, обладающего большим геномом (вспомним гаттерию). Одним из основных правил эволюции считается то, что новые виды образуются не из высокоорганизованных и специализированных форм, а из относительно простых. По-видимому, это относится и к размерам наследственных структур.
Покажем изменение размеров геномов у животных, доминировавших в воде и затем оккупировавших сушу и воздух (рис. 12). По этим данным можно видеть, что костистые рыбы, будучи несколько «моложе» хрящевых, заметно уступают им по размеру генома и, как мы хорошо знаем, теперь доминируют практически во всех водоемах. По этим же данным можно судить, что наиболее «тяжело» дался выход на сушу, т. е. появление амфибий (пришлось увеличить геном во много раз). По-видимому, действительно не просто одновременно заселять два типа пространства — и воду, и землю, надо содержать много кодирующих наследственных структур. Более специализированные рептилии, обитающие на суше, резко уменьшили размеры генома (змеи, например, утратили даже конечности). «Захват» воздушного пространства сопровождался еще большим снижением ненужных структур (чтобы их не носить зря в воздухе), и человек, появившийся сравнительно недавно, как мы видим, не отличается увеличением генома.
В целом можно даже говорить о тенденции к снижению величины генома в макроэволюции животных, по крайней мере верхних значений его амплитуды (на рис. 12 это условно показано стрелками I и II).
Феномен больших разбросов в размерах геномов даже у близкородственных видов «остается совершенно загадочным», — пишет знаменитый генетик Дж. Уотсон в обстоятельном труде «Молекулярная биология гена» ,[М., 1978, с. 507]: «В настоящее время единственным ключом к решению проблемы мог бы служить тот факт, что у близкородственных растений, где содержание ДНК может различаться в 10 раз, количество ДНК коррелирует со сроком жизни. Меньшие величины характерны для короткоживущих однолетних растений...» Кроме того, для растений подмечено, что древовидные папоротники саговники и прочие древние виды, не выдерживающие конкуренции с современными формами, очень часто полиплоидны, в то время как эволюционно продвинутые сложноцветные имеют наименьшее количество ДНК на клетку [Медников, 1980]. Такое заключение генетиков непосредственно «льет воду на мельницу» ЭПИР, так как, согласно показателю интенсивности развития, современники, занимающие близкие ниши, должны иметь близкие энергетические показатели этого типа.

Рис. 12. Изменение размеров (амплитуда) геномов животных в эволюции [по Айала, 1984, модифицировано]. Крестиком отмечено положение человека.
Следует напомнить, что наше рассуждение относилось к абсолютным значениям размеров геномов. И тут нельзя забывать о том впечатляющем факте, что геномы эукариот в сотни, а то и в тысячи раз больше геномов прокариот (см. начало этого параграфа). Например, в клетке млекопитающего содержится приблизительно в 1000 раз больше ДНК, чем в клетке хорошо изученной генетиками кишечной палочки. Абсолютное увеличение налицо. Поговорим теперь об относительных размерах геномов про- и эукариотных клеток.
2. По-видимому, доля ДНК в любой клетке может считаться несомненной характеристикой ее структурного компонента. Можно также полагать, что и содержание РНК в клетке коррелирует с ее энергетической нагрузкой по структуре, так как работающие гены остаются генами, т. е. все же это — единицы структуры, хотя и управляющие через ферменты функционированием клетки. Тогда суммарное содержание нуклеиновых кислот в клетке можно принять за ее базальную структурную часть, а их долю по отношению к общей массе клетки можно полагать за показатель отягощения клетки структурной частью (явно структурная часть биомассы). Приведем данные по относительному содержанию нуклеиновых кислот в клетках разных типов (рис. 13). Обращает на себя внимание сильное «отягощение» прокариотных клеток структурными компонентами: до 18% веса кишечной палочки могут составлять нуклеиновые кислоты. Большую часть из них представляют РНК, и их доля естественным образом возрастает с увеличением скорости роста или уменьшением длительности поколения, т. е. с возрастанием белоксинтезирующей активности клетки. Но и доля ДНК в клетках высших организмов (отмечена штриховой линией на рис. 13) тоже ниже по сравнению с прокариотными клетками и относительно быстро растущими одноклеточными эукариотами (дрожжами и хлореллой).

Рис. 13. Относительное содержание нуклеиновых кислот (РНК+ДНК) в клетках разных типов.
Прокариоты: 1 — кишечная палочка, 2 — водородные бактерии, 3 — светящиеся бактерии; одноклеточные эукариоты: 4 — кормовые дрожжи, 5 — микроводоросль хлорелла; мышечные ткани многоклеточных эукариот: 6 — рыб, 7 — млекопитающих.
Относительное снижение доли нуклеиновых кислот в клетках при прогрессивной эволюции автоматически означает, что энергетический поток на единицу генетической структуры увеличивается. А с учетом удлинения времени жизни структур Ки.р. возрастает дополнительно. Вот почему прокариоты имеют более низкие показатели энергетической интенсивности по сравнению с эукариотами, особенно с млекопитающими, хотя абсолютная скорость их развития гораздо выше. Просто их высокие скорости трансформации энергии соответствуют гораздо большим величинам работающих биомасс; отсюда и каждая единица структуры менее эффективна энергетически (согласно ЭПИР).
Таким образом, можно сделать нетривиальное предположение, что с ходом прогрессивной эволюции имеет место относительное упрощение биологической структуры. Методологически это означает, что принцип простоты поддается экспериментальной проверке в измерениях развития биологических систем. (Это очень уж отдает крамолой: выводить принцип простоты из биологии, а не из физики, например, где он должен быть более очевидным.) Тем не менее в экспериментах с эволюционными машинами можно убедиться, как отбираются наиболее простые структуры, выполняющие определенные функции, в частности, по энергетике.
Особо оговоримся, что в данном случае мы затрагиваем только энергетические показатели развития, т. е. анализ поневоле односторонний. Например, оценивая в целом эволюцию генома, следует учитывать такие сажные изменения его характеристик, как увеличение надежности или повышение ценности кодируемой информации; правда, последнее трудно интерпретируется. Можно говорить, что эволюция генома от прокариот к эукариотам и далее до человека шла не на увеличение структуры генома, а на повышение качества его функционирования, что находится в хорошем соответствии с энергетическим подходом. Поскольку информация пропорциональна логарифму разнообразия то небольшое увеличение генома позволяет кодировать гораздо большее количество структур. Совершенствование систем регуляции, энергетически очень недорогое, может давать многое для победы в конкурентной борьбе. Становится понятным, почему по структурным генам человек и шимпанзе так удивительно схожи, более 99% структурных генов у них одинаковы. Основная разница заключается в совершенствовании регуляторных функций генома.
Глава 7
Первая ступень эволюции жизни на Земле: от химического к биотическому круговороту
Пожалуй, самое удивительное в эволюции жизни на Земле, это то, как быстро она происходила.
Р. Е. Дикерсон
7.1. Как в химических циклах могли образоваться протоклетки
Происхождение и развитие жизни на Земле через химическую эволюцию к преджизни, а затем — к современной жизни у эволюционистов-материалистов не вызывает сомнений. С широко известных работ А. И. Опарина и Д. Б. С. Холдейна, написанных в 20-х годах нашего века, началось изучение и научное обоснование этапов развития жизни. Субстратный подход занял главенствующее положение в изучении первых этапов химической и биологической эволюции. Однако огромное разнообразие возможных вариантов и даже путей образования структур в принципе не поддается описанию и приходится говорить либо о «закономерном появлении новой формы движения материи» (см. гл. 1), либо о «самоорганизации» (см. гл. 2).
Энергетический подход в сочетании с субстратным позволяет резко уменьшить рассматриваемое число возможных вариантов и путей развития, заменив традиционный вопрос: что было раньше — наследственная молекула или белок («яйцо или курица»), на другой: как выглядела первичная экосистема. Или точнее: как изменялось первичное вещество на поверхности нашей планеты с учетом непрерывной накачки энергией (главным образом от Солнца, но на определенных этапах формирования нашей планеты и изнутри ее). При этом первичные химические преобразования должны были носить циклический характер, так как потоки энергии прежде всего вызывают механические циклы перемещения газов и жидкостей. Мы знаем, что в ограниченной системе образуются циклические потоки, разнообразные по размерам и длительности (в гл. 4 мы их обсуждали). При этом вещество то попадает в зону действия потока энергии (УФ-лучи, разряды молний и прочее), то уносится из этой зоны в спокойные тихие места, где направление химической реакции может и должно смениться на противоположное. Особенно эффективной в организации наиболее быстрых, а следовательно, и наиболее активных циклов химических реакций и потоков вещества является смена дня и ночи. И уже здесь мы можем говорить от отборе, идущем в двух направлениях. Первое — уход тех «участников», которые выпадают из круговорота (в осадки, захоронения или улетают в космос). Так отбиралась, например, вода как активный участник физического круговорота. Второе направление, более интригующее, это отбор по ускорению химических реакций. Этот отбор связан с увеличением потоков вещества и его трансформацией через те реакции, в которых это вещество быстрее реагирует — в полном соответствии с известным принципом максимума скоростей реакций в открытых химических системах.
В данной главе мы и рассмотрим схематически этапы становления и развития жизни на нашей планете, опираясь на субстратный и подчеркивая энергетический аспект развития циклов от химических до биотического. При этом энергетическая оценка, определяя направление развития (т. е. ответ на вопрос: «куда» шло развитие), позволяет несколько снизить остроту вопроса: «кто» конкретно развивался.
Если оценивать возраст Земли в 4,6 млрд лет, что общепринято, и учесть, что о первом миллиарде лет почти ничего не известно (пет геологических свидетельств), то уже практически в первых сохранившихся отложениях осадочных пород обнаруживаются микроокаменелости, напоминающие бактерии. По крайней мере разрыв не превышает 0,3 млрд лет. Отсюда понятна и точка зрения тех исследователей, которые полагают, что жизнь была занесена на нашу планету извне. Однако прибегать к теории панспермии, да еще направленной, нет особой необходимости, так как большинство этапов абиогенного синтеза в настоящее время имеют естественно-научное объяснение, а целый ряд их может быть воспроизведен экспериментально.
Сама проблема происхождения жизни может быть разбита на пять отдельных вопросов: 1) образование планеты с атмосферой и гидросферой, где имеется «сырье» для возникновения жизни; 2) синтез биологических мономеров, исходных кирпичиков жизни, например аминокислот, Сахаров и органических оснований; 3) синтез биологических полимеров типа белковых и нуклеиновокислотных цепей; 4) вычленение отдельных частиц типа капель бульона по Холдейну, микросфер по Фоксу, коацерватов по Опарину, их превращение в пробионты с собственным химизмом; 5) возникновение репродуктивного аппарата, передающего дочерним клеткам химические и метаболические потенции родителей. Кратко это формулируется как проблемы исходного сырья, образования мономеров, полимеров, изоляции и репродукции.
Чтобы не остаться в рамках только субстратного подхода к изучению жизни, мы должны обратить внимание на «движущие силы» химической эволюции, а именно на источники энергии для химических синтезов в примитивной атмосфере Земли. И еще раз вспомним о вызываемых ими циклах реакций: синтеза — ре-синтеза, образования — разрушения. Каковы же были эти источники энергии? Основным источником, как и теперь, было Солнце, спектральный состав излучения которого не изменился. Однако свет проходил через атмосферу иного состава. Кислорода, который имеет биогенное происхождение, не было, а следовательно, не было и озонового экрана, который поглощает теперь почти все коротковолновое ультрафиолетовое излучение. Тогда большая часть этого высокоэнергетического излучения достигала земной поверхности. Следовательно, большие количества активной фотохимической энергии были доступны для взаимодействия с веществом. Может быть, с учетом этого высокая скорость химической эволюции и не покажется столь удивительной. Кроме того, возможно, что и электрические разряды были более мощными, так как для первобытной земной поверхности была характерна высокая вулканическая активность. Жар лавовых потоков, сильные ливни создавали самые разнообразные условия для протекания химических реакций. Например, выпадение вулканического пепла могло служить в локальных водоемах хорошим временным экраном от разящих УФ-лучей.
К настоящему времени проведено множество модельных экспериментов по химической эволюции. Было обнаружено, что при воздействии разных видов энергии на газообразный углерод, азот, воду, водород, входящие в состав примитивной атмосферы, вначале образуются реакционноспособные промежуточные продукты. А они затем дают множество биологических или близких к биологическим мономеров и некоторые полимеры. Как подчеркивает К. Фолсом, автор книги «Происхождение жизни» [М., 1982], для суждения о процессах на примитивной Земле необходимо рассматривать не отдельно взятый эксперимент, а всю их совокупность. Первые циклы могли иметь такую структуру:

Направление реакций определялось притоком энергии в зависимости от места и времени (например, времени суток). Можно считать, что проблема синтеза мономеров не заключает в себе каких-либо фундаментальных трудностей или трудностей философского характера.
При синтезе полимеров необходимы одновременно приток энергии и отщепление воды. Как и при синтезе мономеров, проблема небиологического синтеза полимеров не имеет принципиальных трудностей для понимания, хотя некоторые затруднения здесь имеются (к примеру, для конденсации лучше всего подходят безводные условия, в которых очень сложно представить ход эволюции живых систем).
Следующий этап химической эволюции — развитие фазово-обособленных систем. И здесь модельные эксперименты дают нам большое разнообразие возможных вариантов. Это — коацерватные капли Бунгенберг-де-Йонга и Опарина, пузырьки Голдейкра, микросферы Фокса и т. д. Отметим очень важное свойство фазовой обособленности или наличия границ в замкнутой системе. Полимеры, возникающие в растворах, не могут достичь высоких концентраций, в частности, из-за протекания обратных реакций. А полимеризация в ограниченном, выделенном объеме снижает в нем концентрацию мономеров и, соответственно, понижает осмотическое давление. Такое снижение приводит к перекачке мономеров из окружающей среды. И таким образом пробионты способны «высасывать» органику из первичного бульона, а значит, расти и почковаться или делиться. По образному выражению профессора Б. М. Медникова, [1980, с. 425], «не жизнь породила клетку, а клетка возникла раньше самой жизни».
Действительно, можно выделить ряд свойств пробионтов, чтобы они могли стать прародителями первичных живых клеток: способность к обмену с окружающей средой (проницаемая мембрана); способность к росту, увеличению объема; способность к делению и почкованию. Особого внимания заслуживает способность пробионтов к первичному метаболизму, т. е. к протеканию специфических синтетических и биохимических реакций. Это приводит к тому, что локальные условия в них сильно отличаются от условий внешней среды. Например, коацерваты Опарина, состоящие из полинуклеотида и белка, при добавлении полинуклеотид-фосфорилазы в присутствии АДФ способны синтезировать полинуклеотид-полиадениловую кислоту. При этом капли растут в размере и способны к механическому разделению.
В экспериментах Фокса в результате нагрева смеси аминокислот, с последующим охлаждением и переносом в воду, образовывались протеиноподобные микросферы. Они также характеризовались определенной каталитической активностью и были способны к почкованию или делению, как и большинство бактерий.
В модели Бернала полимеры сорбировались на глинистых минералах, и предполагалось, что далее они самоорганизуются в протоклетки с метаболизмом и отбором.
Эксперименты Опарина, Фокса и других — всего лишь демонстрация того, как работают физико-химические фазово-обособленпые системы. Но они показывают аналогии жизненных процессов в простых системах и позволяют проиллюстрировать идеи выживания и отбора на уровне химических систем. Из этих экспериментов следует, что образование коацерватных капель и микросфер — это типичное поведение полимеров в растворах. Шансы таких капель на выживание повышаются, если они способны к каталитической активности, в результате которой могут расти в размерах. Те из них, которые обладали повышенной скоростью «высасывания» мономеров из окружающей среды, развивались быстрее и побеждали в конкурентной борьбе.
Таким образом, можно себе представить, что на протяжении целых геологических эр действовал мощный химический отбор. Он приводил к ускорению химических процессов. Механизм этого действия практически очевиден.
Согласно принципу максимальных скоростей реакций в случае нескольких открытых химических систем с общей внешней средой основной поток вещества идет через систему, которая обеспечивает наибольшую скорость химических превращений. Такие пробионтные системы в «первичном бульоне» получали преимущество перед соседними и начинали вытеснять более медленные (менее приспособленные) формы. Под воздействием внешних механических сил, таких как ветер и волны, происходило дробление (деление) капель. Запасы готовых органических веществ, пригодных для прямого использования, естественно, были ограниченны, что приводило к конкуренции за субстрат и, таким способом, к возникновению «предбиологического естественного отбора». Применение термина «естественный отбор» к эволюции коацерватов-пробионтов представляется вполне допустимым, так как никаких специфических отличий между популяциями протобионтов и современных микроорганизмов с точки зрения действия отбора не имеется. В том и другом случае отбор приводит к увеличению приспособленности популяции, что выражается через изменение действующих скоростей роста. А характер и направление отбора определяются условиями среды.
В этом смысле применение методов непрерывного культивирования, разработанных для исследования микробных популяций, по-видимому, является весьма перспективным для изучения действия отбора в популяциях протобионтов и в конечном счете для моделирования данного этапа эволюции, заключающегося в возникновении и совершенствовании метаболизма.
Совершенствование метаболизма может изучаться в проточной системе по методу, основанному на модели Н. Горовица. Логика рассуждений данного автора сводилась к тому, что в некоторый момент в первичном бульоне усваиваемые вещества A оказались полностью израсходованными; тогда те протобионты, которые были способны производить A из других доступных соединений B, получили преимущество. Когда, в свою очередь, снизилось количество вторичных питательных веществ B, возникла необходимость в образовании A и B из C и т. д. Приобретение соответствующих катализаторов, ускоряющих эти реакции, от простых катализаторов до ферментов, определяло степень усложнения этого процесса и ускорения метаболизма.
В соответствии со схемой Горовица об удлинении цепей метаболизма легко представить себе замыкание этих цепей в циклы, первые круговороты вещества с участием клеток. Причем необязательно это могло осуществляться в одном типе фазово-обособленных систем, возможно распределение по звеньям цикла, с вычленением звеньев. Вначале это гетеротрофное звено с наиболее древним источником энергии — гликолизом; затем, по мере исчерпания органики, подключение автотрофных вариантов. Подробнее мы обсудим это в следующем параграфе, а пока коротко оценим два альтернативных варианта использования энергии при развитии протобионтов.
Конкурентную гипотезу о прямом использовании энергии протоклетками развивает американский исследователь К. Фолсом. Он обращает внимание на то, что в экспериментах при воздействии энергии на смесь первичных газов, т. е. уже на первом этапе, не только образуются малые органические молекулы, но и обнаруживается полимерный материал, содержащий большое количество углерода. Обычно он осаждается на стенках реакционного сосуда или на электродах, иногда образует маслянистую пленку на поверхности воды. Химически он трудно интерпретируется. При встряхивании или при перемешивании такая пленка может образовывать сферулы от 1 до 20 мкм в диаметре. Они имеют двойную гидрофобную мембрану. После самосборки они медленно опускаются на дно сосуда. Такие структуры имеют одну удивительную способность: после начала реакции в искровом разряде их число возрастает во времени экспоненциально. По замечанию К. Фолсома, это может свидетельствовать о том, что одна микроструктура служит центром для самосборки других, а именно такого рода автокатализ и является характеристикой биологических популяций.
Прямое использование энергии и большой выход реакции (практически весь углерод переходит в эти структуры) заставляют обратить на такие сферулы особое внимание. Рецепторами энергии в них могут служить порфирины, которые легко получаются в экспериментах по имитации химической эволюции пирролов. Протоклетки, имеющие гидрофобную границу раздела фаз, способны избирательно адсорбировать порфирины. В свою очередь, сорбированные порфирины могут служить рецепторами ультрафиолетового излучения, устанавливать протонные градиенты и превращать энергию излучения в потенциальную энергию химических связей. Следовательно, на самой ранней стадии возникновения жизни возможно существование гетеротрофных фотосинтезирующих организмов, использующих УФ-излучение для создания полимеров. Даже нерегулярные полимеры аминокислот, образующие комплексы с ионами металлов, обладают слабой каталитической активностью. Так открывается поле деятельности для естественного отбора.
Вторая из конкурентных гипотез имеет дело с прямым использованием энергии первичных газовых выбросов изнутри нашей планеты. В гл. 4 мы подчеркивали, что основу функционирования живых систем составляет цикл реакций окисления — восстановления. В первичной атмосфере окислительные условия создавались за счет фотохимических реакций, к примеру отщеплением водорода с его диффузией в космос. По расчетам, восстановленные соединения типа CH4 в такой атмосфере неустойчивы и быстро окисляются. Глубины Земли, наоборот, являются источником восстановительных газов, которые поступали изнутри особенно интенсивно на ранних этапах развития самой планеты.
Представляется возможным даже полностью независимое развитие и существование литотрофных организмов за счет энергии водорода и других восстановительных газов, имеющих как ювенильное, так и метаморфическое происхождение. Одним из главных условий поддержания и развития микроорганизмов (первичных организмов) является наличие длительного и достаточного потока энергии. По крайней мере, хемолитотрофные организмы способны окислять все основные компоненты вулканических газов: H2, CO, NH3, CH4, SO2 и т. д. Поэтому в местах длительного выхода глубинных газов может развиваться микробное сообщество, использующее не продукты разложения органического вещества, синтезированного каким-то другим, а первичные продукты газовых выделений [Заварзин, 1984], таким образом вместо фототрофии имеется возможность хемолитотрофии.
Совершенствование пробионтов под влиянием естественного отбора постепенно привело к появлению живых клеток. По метаболизму ни одно живое существо в принципе не делает больше того, что могли делать пробионты [Медников, 1980]. Поэтому возникновение систем репликации и передачи наследственного материала от родительских к дочерним клеткам следует считать одной из важнейших черт жизни. Однако именно здесь кроется самая большая тайна. Можно согласиться с Р. Дикерсоном [1981], что эволюция генетического аппарата — это тот этап эволюции, для которого лабораторных моделей не найдено, поэтому рассуждать о ней можно бесконечно, не смущаясь неудобными фактами. Действительно, генетический аппарат современных организмов настолько сложен и универсален, что почти невозможно его себе представить в примитивном виде. А это значит, что главные принципы эволюции — ее непрерывность и последовательность — пока еще четко не продемонстрированы.
Не вызывает сомнения, что генетический аппарат эволюционировал согласованно (т. е. «курица и яйцо» вместе) из наиболее простых форм. Важно отметить, теперь уже с позиций энергетического подхода, что простые первичные варианты, как неэффективно функционирующие, были вытеснены в дальнейшей конкурентной борьбе и исчезли впоследствии. О них теперь можно только гадать. Одной из самых загадочных является проблема возникновения рибосомального аппарата биосинтеза белков. Тут сразу требуется несколько десятков молекул специфических белков и не менее трех типов молекул РНК с различными молекулярными весами.
Постепенность развития биополимеров в протоклетках связана с увеличением их малых, по сравнению с современными биополимерами, размеров. Первичные «белки» могли быть совсем небольшими молекулами, могли состоять лишь из пяти — семи аминокислотных остатков. И первичные полинуклеотиды содержали не миллионы, а десяток-другой оснований. Такие полимеры и получаются во многих экспериментах, имитирующих начальные условия. Напомним, что каталитический активный центр фермента почти всегда гораздо меньше всей молекулы фермента, он имеет лишь небольшое число аминокислотных остатков. Остальную часть большой молекулы можно считать позднейшей надстройкой: она не связана с катализом отдельной реакции, а служит для целостного контроля в клетке.
При таком подходе можно постепенно двигаться дальше. Представим небольшую генераторную РНК, выполняющую и генетическую, и матричную роль. Более устойчивая ее форма — кольцо. В этой же клетке может быть несколько коротких тРНК. Генераторная РНК способна реплицироваться без ферментов, хотя и медленно. На циклическом генераторе могут непрерывно реплицироваться новые РНК, гораздо более длинные, двух типов: крупные кольцевые и линейные. Для ускорения реакции необходимы простые полипептиды, катализирующие синтез олигопуклеотидов. Синтез таких пяти-, семичленных пептидов, но уже со специфической последовательностью, возможен с помощью коротких первичных тРНК на генераторной РНК, выполняющей роль матричной РНК. Связывание аминокислоты с тРНК, возможно, обеспечивалось энергией пирофосфатпых связей. Внешняя среда служила источником всех необходимых малых молекул, т. е. они «высасывались» протоклеткой из среды по правилам химической кинетики. Пирофосфаты образовывались под влиянием потока энергии ультрафиолетового излучения Солнца. Так могла работать первичная живая клетка по К. Фолсому. (Но, может быть, и не совсем так.)
Скорости функционирования таких протоклеток были невысоки, но и специфичность катализа тоже была невысокой, а это резко снижает требования к уникальности биополимеров. Сходные функции способны выполнять разнообразнейшие структурные сочетания, миллиарды миллиардов вариантов! (Это, как правило, не учитывается в расчетах по вероятности возникновения жизни, так как принимается в расчет вероятность образования определенной конкретной структуры биополимера, якобы обладающей уникальной функцией.) В сочетании с малыми размерами первичных биомолекул шансы на быстрое образование первичных клеток, т. е. на возникновение жизни, резко повышаются.
Очень доказательными в этом смысле являются эксперименты по молекулярной эволюции, проведенные группой Спигелмана, о которых мы писали в предыдущей главе. Помимо изменения размеров фаговой РНК, реплицирующейся с помощью фермента репликазы, обнаружено, что фермент способен катализировать синтез рибонуклеотидных цепочек и без матрицы. Синтез шел медленнее, до тех пор пока образующаяся цепь не становилась матрицей сама. Полученная РНК оказалась совершенно непохожей (!) на фаговую РНК. Она была случайной последовательностью нуклеотидных остатков, но реагировала на факторы отбора подобно ее специфической фаговой форме. Следовательно, не строгая структура определяет функцию, а под функционирование подбирается структура. И, как мы обсуждали в предыдущей главе, можно предсказывать направление отбора соответствующих структур согласно энергетическим принципам.
Коротко резюмируем суть рассмотренного этапа развития жизни — химической эволюции, вплоть до образования первых живых клеток. Основу его составляет физико-химическое концентрированно абиогенно образованного органического вещества в пробионтах. Отбор, возникающий уже на этом предбиологическом этапе, действует не на отдельные молекулы, а на целостные фазово-обособленпые структуры. Выигрывали те из них, которые наиболее эффективно прокачивали через себя вещество под влиянием внешнего потока энергии (структура подгонялась под функцию). Прямым или косвенным источником этой энергии был поток солнечного излучения и, возможно, поток доступной энергии изнутри Земли, например с газовыми выделениями. Возникновение генетического кода резко ускорило ход эволюции и действие отбора, так как появился автокатализ в ограниченной среде (основа для действия отбора в открытых системах, неважно: живых или неживых).
В заключение особо оговорим энергетические преимущества перехода к живым системам, совершенствования и усложнения структур протоклеток. Фазовое обособление структур очевидно из физико-химических требований. Однако по энергетике, например при прямом взаимодействии с квантами света, молекулы, связанные в полимер и укрытые в клетке, могут даже частично проигрывать по сравнению со свободными молекулами того же типа, оставшимися в первичном бульоне. В частности, это может происходить из-за эффекта затенения их друг другом или оболочкой клетки. Но мощнейшим противовесом, компенсирующим все потери, служит возникающий метаболизм. Высасывание по законам химической кинетики органических молекул с запасенной в них абиогенно энергией из объемов, гораздо больших, чем размеры самих пробионтов, резко увеличивает энергетическую нагрузку на каждую включенную в состав протоклетки молекулу. Первичные варианты, способные к автокатализу, относительно быстро смогли использовать самые доступные органические молекулы из первичного бульона — это и есть развитие по ЭПЭР (захват энергии и пространства без изменения качества). А при нехватке доступного источника стала в отборе совершенствоваться качественная сторона — интенсификация процесса метаболизма старых соединений и возрастание умения утилизировать новые источники. Это — прямое проявление действия ЭПИР (совершенствование структуры для выполнения функций, связанных с перекачкой энергии). Развитие и совершенствование циклов недостающих веществ выглядит здесь очевидным.
Итак, всего вероятнее, что первый шаг в «оживлении» химического круговорота, первый разрыв химического цикла, а точнее, встраивание в него, были сделаны гетеротрофными анаэробными формами. По сравнению с остальными организмами их пути метаболизма гораздо короче, а энергия, используемая ими, заключена в транспортабельной форме в абиогенно образованных органических молекулах. Первичные сопрягающие агенты в форме полифосфатов также могли иметься в наличии в результате простых химических синтезов.
Основным поставщиком энергии, самым простым, самым универсальным, является гликолиз, или анаэробное брожение. При нем происходит разрушение глюкозы или родственных ей соединений, и высвобождаемая энергия запасается в форме «моченого полифосфата», т. е. в форме молекулы АТФ, которая и является универсальной энергетической «валютой». Следовательно, по схеме эволюции «от простого к сложному» («задом наперед», по Горовицу) гликолитический путь получения энергии возник после исчерпания полифосфатов, образовавшихся абиогенно путем химической конденсации. Видимо, скорости химического образования полифосфатов уже не могли удовлетворять энергетические потребности растущей популяции примитивных организмов. И это естественно. Автокатализ, характерный для живых систем, быстро показал неэффективность химического синтеза вещества, а особенно его энергетическую недостаточность.
При гликолизе, в результате 10 согласованных реакций, каждая молекула глюкозы расщепляется на две молекулы пирувата, а клетка получает две макроэргические фосфатные связи в виде молекул АТФ (из АДФ). Подчеркнем единство энергетического и субстратного подходов. Пируват служит субстратом для целого семейства нужных для клеток соединений, например этилового спирта, молочной кислоты и ряда других кислот типа муравьиной, уксусной, янтарной, масляной; пропилового и бутилового спирта, ацетона, газообразного водорода и т. д. Это определяется природой конечного акцептора электрона. И, что очень существенно, весь этот «букет» достигается включением лишь небольшого числа дополнительных реакций и с применением сходных каталитических механизмов. Все эти вещества служат строительным материалом для дальнейших синтетических реакций конструктивного обмена. Если к этому добавить, что гликолиз характеризуется необычайно высокими скоростями протекания реакции, а следовательно, и получения энергии, то становится ясным, почему он так широко распространен в современном живом мире.
Недостатком его является невысокая степень высвобождения энергии из исходного субстрата: продукты его остаются еще высокоэнергетическими. Поэтому электроны, поднятые прямо или косвенно энергией фотонов на высокий энергетический уровень, опускаются не на нижний основной уровень, а совсем немного, останавливаясь на промежуточных уровнях восстановленных соединений-акцепторов.
Подытожим результаты рассмотрения первого этапа становления биотического круговорота. Это — гетеротрофные, анаэробные одноклеточные организмы, возможно похожие на современные бактерии, такие как клостридии, живущие за счет брожения. Они существуют за счет распада богатых энергией органических соединений, образовавшихся абиогенно. Они играют роль «мусорщиков», уничтожая органику химического происхождения, возникшую под влиянием УФ-лучей, электрических разрядов, ударных волн и прочих источников энергии. Основная функция гетеротрофов — деструкция органических соединений. Они ее выполняют быстро за счет автокатализа и таким образом «выжимают» все, что может дать химический синтез, гораздо более медленный по сравнению с биологической деструкцией. Наступает первый кризис из-за несбалансированности круговорота. И кризис этот — энергетический, так как косвенного производства энергии через полифосфаты явно недостаточно. Кроме того, низкий энергетический выход процессов брожения требует переработки громадного количества субстрата для обеспечения энергией биосинтетических процессов в клетке. Например, по сравнению с позднее возникшим окислительным фосфорилированием гликолиз забирает лишь около 7% энергии, запасенной в молекуле глюкозы (но об этом чуть позже). Таким образом, первичной жизни не хватало доступной энергии.
И в то же время потоки энергии «бушевали» вокруг простых протоклеток, задевая их самих. Источником этих потоков были Солнце и ядро Земли, дававшее богатые энергией газовые эксгаляции.
7.2. Первый биотический круговорот
(цианобактериальное сообщество)
Первичные гетеротрофные клетки, естественно, были частично окрашенными и потому взаимодействовали с потоком солнечного излучения. Оно могло разрушать молекулы, особенно его УФ-часть, или терялось в виде тепла. Но в толще воды, более интенсивно поглощавшей коротковолновое излучение, могли иметь место и другие взаимодействия, в частности взаимодействия о длинноволновым излучением. Поглощение света сложными молекулами могло приводить к развитию фотохимических реакций, в которых за счет энергии света при обычных температурах преодолевались высокие энергетические барьеры. Это приводило и к ускорению скоростей реакций, и к практически необратимому синтезу еще более сложных соединений. Естественно считать, что флюоресцирующие сложные молекулы, входящие в состав живых клеток, вначале приводили к ускорению лишь некоторых процессов метаболизма, а не к прямому фотосинтезу органических веществ.
Среди фотосенсибилизаторов, т.е. оптически активных молекул, возбуждаемых квантами света, наиболее часто встречаются (как составные части ферментов) соединения, построенные путем сочетания неорганических ионов с органической основой, которую, как правило, составляет порфириновый цикл. Порфирины могут образовываться из пирролов и формальдегида, несколько хуже в восстановительной, чем в окислительной среде, а для синтеза наиболее восстановленных порфиринов требуются строго анаэробные условия. Включение металла в центр порфириновой молекулы заметно увеличивает ее фотохимическую активность, а кроме того, и сильно увеличивает интенсивность окраски порфирина и тем самым его способность к поглощению видимого света. Дальнейшая эволюция шла по пути увеличения количества светочувствительных пигментов и усложнения их структур.
Серьезным завоеванием на пути к полной автотрофии явился анаэробный фотосинтез. Его представителями, сохранившимися до нашего времени, являются фотосинтезирующие бактерии (пурпурные, серные и несерные; зеленые серные бактерии). Они способны усваивать энергию света, но еще не способны к отрыву электрона от воды. Они используют в качестве восстановителя (источника электрона и водорода) различные органические или неорганические соединения. Например, для фотохимического отнятия электрона от сероводорода требуется значительно меньше энергии, чем для отнятия его от воды.
Главным эволюционным приобретением, лежащим в основе фотосинтеза, как в целом качественно нового этапа в развитии биоэнергетических систем на Земле, следует считать организацию электронного потока. Именно он оказался наиболее эффективным способом запасать энергию электронного возбуждения в виде химических связей.
Постепенное уменьшение содержания в среде восстановленных органических субстратов заставило в обостряющейся конкурентной борьбе расширять круг используемых источников углерода. Световая энергия из дополнительного источника энергии, облегчавшего фотоассимиляцию имевшихся органических соединений, превращалась в основной, более мощный поток. В клетках накапливалось большое количество пигментов, шел отбор наиболее эффективно работающих систем, происходило пространственное упорядочивание пигментных структур, совершенствовались механизмы миграции энергии возбуждения от всей массы пигментов к каталитически активно работающему пигменту — активному центру. (В современных организмах энергия, поглощенная большим количеством пигментов, находящихся в агрегированных структурах, очень быстро и эффективно передается к активному центру.)
При растущем дефиците органических соединений фотосинтезирующие бактерии приобрели способность усваивать в качестве источника углерода углекислоту, широко имевшуюся в наличии. Но, чтобы восстанавливать CO2 до уровня восстановленности углеродсодержащих соединений клетки (типа углеводов—(CH2O)n), потребовался постоянный источник электронов (протонов). Световая энергия стала расходоваться на образование АТФ и на образование восстановителя, и таким образом сформировался нециклический путь переноса электронов. Возникавшие электронные вакансии в возбужденных молекулах хлорофилла (дырки) потребовалось заполнить за счет организации непрерывного притока электронов. В окружающей среде шел поиск соединений, способных выполнять функцию внешних доноров электронов. Одной из таких находок и были соединения серы, о которых мы уже говорили. Использование соединений серы автоматически привязывало организмы к местам, где эти соединения имелись.
Самым распространенным веществом у поверхности Земли была вода. Поэтому организмам, способным использовать воду в качестве донора электронов, была гарантирована победа в борьбе за существование. Из тех древнейших форм прокариотных клеток, способных к фотолизу воды, до нас дошли современные варианты — цианобактерии, или синезеленые водоросли.
С развитием прокариотных фотосинтезирующих клеток (около 3 млрд лет назад) замкнулся биотический круговорот. Появилась возможность существования обеих ветвей, имеющих живую основу: ветви синтеза и ветви деструкции. Их энергетическая независимость от абиогенного химического синтеза может считаться одной из основных черт данного этапа. Жизнь в виде биотического круговорота вступила в свои права и стала перестраивать лик планеты.
Для нас наиболее важно, что в пределах группы цианобактерии сформировался и развился новый тип энергетики, который затем был «принят на вооружение» и высшими организмами. Это формирование фотосистемы II, обеспечивающей использование воды и выделение молекулярного кислорода. Развитие этой фотосистемы связано с появлением новой группы фоторецепторов (типа хлорофилла а и фикобилипротеидов) и образованием фотохимически активных реакционных центров, способных фотоокислять воду. Достройка новой фотосистемы к старой фотосистеме I позволяет оторвать электрон от молекулы воды и «подбросить» его на более высокий уровень, а дальше он уже может использоваться в энергетических превращениях фотосистемы I. Как конкретно поэтапно формировалась фотосистема II, пока неизвестно, ибо современные цианобактерии — это результат длительной эволюции.
Некоторые представления о функционировании первичного круговорота можно получить на примере изучения современных цианобактериальных сообществ, развивающихся «на задворках» биосферы: в термальных источниках, соленых морских лагунах или на берегах мелководных сильно засоленных озер. Следуя работе Г. А. Заварзина [1984], опишем, в качестве примера, сообщество, развивающееся в лагунах юго-восточной части Сиваша. Повышенная соленость этих мелководных водоемов глубиной несколько десятков сантиметров ограничивает развитие высшей растительности и эукариот вообще. Дно таких водоемов покрыто кожистой пленкой розоватого или серого цвета. Основной формообразующий компонент сообщества — это синезеленые водоросли рода микроколеус. Их трихомы переплетены в виде канатов, заключены в общее слизистое влагалище и даже способны мигрировать по вертикали. Самый верхний слой составляют слизистые бесцветные бактерии, они являются аэробными органотрофами. Днем под их слоем образуются пузырьки фотосинтетического кислорода. Находящийся под ним слой синезеленых водорослей имеет зеленую окраску. Он представляет собой плотную пленку толщиной несколько миллиметров. Под этим слоем находится оливковый слой трихомных бактерий. Под ним, в анаэробной зоне, где развиваются анаэробные бактерии, идет образование карбонатных материалов и гипса. Еще ниже — слой образования сероводорода и выпадения черного сульфида железа.
Автору этих строк довелось обнаружить цианобактериальное сообщество в Средней Азии, в пустыне, недалеко от г. Бухары. Около искусственного пресноводного оз. Тадакуль, за его насыпными берегами, есть места скопления соленой воды и солончаки. В соленых мелких лужах, берега которых покрыты коркой соли, очень чистая, прозрачная вода. Их дно покрыто серо-желтым слоем. Это — верхняя, бактериальная часть мата, похожая на войлок, с переплетением нитей толщиной 2–3 мм. Под ней — удивительно яркий, зеленый слой цианобактерий толщиной не более 1 мм. Еще ниже — сероватый 2–3-миллиметровый слой анаэробов, а затем — черная зона с запахом сероводорода. Видимо, так выглядела первичная биосфера.
Применение микроэлектродной техники показывает резкую вертикальную стратификацию метаболизма в таком цианобактериальном мате. Например, практически весь падающий свет может поглощаться слоем синезеленых водорослей около 0,3 мм. Здесь же и происходят процессы синтеза биомассы. Ниже идет деструкция органического вещества. И, что особенно важно отметить, она протекает анаэробно, без затраты O2. Это как раз и соответствует первичным условиям фотосинтеза и деструкции, которые имели место в бескислородной среде.
Остановимся чуть подробнее на другой части работы биотического цикла — на процессах деструкции. Как известно, в условиях отсутствия кислорода основным источником энергии для гетеротрофного звена является процесс брожения, или субстратного фосфорилирования. Замыкание биотического круговорота привело к тому, что органический углерод стал связываться во все более трудные для сбраживания формы в виде полимеров (белков, полисахаридов, нуклеиновых кислот и других). Часть соединений биотического или абиогенного происхождения оказалась вообще недоступной для сбраживания. К ним относятся прежде всего алифатические и ароматические углеводороды, составляющие основу нефти, сохранившейся до нашего времени.
В целом этот этап развития круговорота оказался «перекошенным» в пользу синтетической ветви, особенно по энергетике (второй энергетический кризис). Следующий шаг в развитии биотического круговорота и заключался в использовании побочного «ядовитого» продукта фотосинтеза — кислорода в цепях дыхания, т. е. в активизации звена деструкции, необходимой для обеспечения баланса круговорота.
Факт, что молекулярный кислород атмосферы имеет биогенное происхождение, в настоящее время почти не вызывает сомнений. Кислород является результатом нового этапа фотосинтеза, при котором в качестве донора электронов (протонов) служит вода. Усиление синтетической половины первичного биотического круговорота привело к связыванию углерода в биомассе автотрофов, к снижению концентрации CO2; в воде. Это могло вызывать защелачивание воды, а следовательно, и выпадение в осадок солей двухвалентных элементов типа кальция, т. е. еще большее обеднение гидро- и атмосферы углеродом. Гетеротрофное звено и с ним звено редукции явно отставали от звена синтеза. В атмосфере накапливался сильнейший яд — окислитель для существ, развивающихся в восстановительной атмосфере. Несомненно, что на этом этапе эволюция должна была быть связана с адаптацией к кислороду. Иначе — смерть и самим фотосинтетикам, его производящим. И мы можем проследить (теперь уже имеется достаточно данных палеонтологии, биохимии и физиологии), как живые организмы блестяще справились с этой эволюционной задачей. Сильнейший токсикант удалось не только обезвредить, но и явно использовать для ликвидации узкого места круговорота: в отборе получили преимущество те организмы, которые сумели использовать молекулярный кислород, прежде всего для своих энергетических потребностей.
Так как цианобактерий считаются первыми производителями кислорода, то им первым и должны были понадобиться защитные механизмы от его токсичности. Как конкретно происходило превращение нейтрализующих реакций в полезные, идущие с использованием молекулярного кислорода, остается неясным. Достаточно правдоподобные объяснения имеются, мы не будем их обсуждать детально (оставим это для субстратного подхода). Рассмотрим здесь только канву энергетических приобретений.
Накопление кислорода означало, что появился «идеальный» конечный акцептор электронов, т. е. открылась возможность передавать ему электроны с восстановленных при фотосинтезе органических соединений. Правда, разрыв по энергетике между донорами и акцептором был велик и ничем не заполнен. Развитие и совершенствование цепи переносчиков и ее связи с энергодающими системами клетки было основным направлением действия отбора в энергетическом смысле.
Чтобы наилучшим способом использовать открывшиеся энергетические возможности, связанные с переносом водорода («горячих» электронов) с субстрата на молекулярный кислород, гетеротрофным клеткам пришлось решать, как минимум, три сложные задачи. Во-первых, полностью отщепить водород от имевшегося органического субстрата. Это было сделано путем развития цикла трикарбоновых кислот (ЦТК). Во-вторых создать систему переноса электронов по электрохимическому градиенту, которую мы называем «дыхательная цепь». В-третьих, связать эту систему электронного транспорта с фосфорилированием, т. е. с образованием единой энергетической валюты — АТФ.
Отметим два очень существенных обстоятельства. Во-первых, окислительное фосфорилирование сходно с фотофосфорилированием, по крайней мере по энергетическому выходу они близки. И таким образом, гетеротрофное звено подтянулось до уровня фототрофного по энергетике, а биотический круговорот на основе прокариот сбалансировался. Во-вторых, для прокариот, сформировавшихся в «докислородную» эпоху, задача обезвреживания кислорода так и осталась одной из самых сложных. К примеру, у тех же цианобактерий фотосинтез и дыхание разобщены во времени, тогда как у эукариот-фотосинтетиков, сформировавшихся позднее, во времена кислородной эры, эти процессы могут идти одновременно из-за их пространственного разделения. Кроме того, у прокариот степень сопряжения электронного транспорта в дыхательной цепи с окислительным фосфорилированием невелика. У наиболее продвинутых в эволюционном отношении аэробных форм она достигает лишь 1/3 от эффективности сопряжения у эукариот. (Измерения проводятся по величине P/O, т. е. по числу потребленных молекул неорганического фосфата, или образовавшихся молекул АТФ, на один поглощенный атом кислорода. Для эукариот P/O = 3, для прокариот — около 1.)
Зато в биохимическом отношении прокариотные организмы отличаются громадным разнообразием: и по конечным акцепторам электронов, и по использованию разнообразных субстратов органической и неорганической природы, и по составу промежуточных переносчиков в дыхательных цепях. Действительно, поразителен тот факт, что в царстве прокариот имеется практически все, что достигнуто жизнью в области биосинтетических процессов. И «если бы мерой эволюционного прогресса служили только биосинтетические возможности, то многоклеточных животных следовало бы считать гораздо менее продвинутыми по пути эволюции, чем хемоавтотрофных бактерий...» [Маргелис, 1983, с. 131].
Можно сказать, что в настоящее время прокариоты заняли все те «пустые» и трудные для жизни места, которые эукариоты не смогли занять прежде всего из-за ограниченности метаболических возможностей. Но магистральное направление эволюции, биологический прогресс по вкладу в биотическии круговорот оказались на стороне эукариот.
Глава 8
Вторая ступень эволюции:
от амебы до человека
...Происхождение эукариотных клеток мыслится как частный случай общего феномена — эволюции микробных ассоциаций.
Л. Маргелис
Поведение представляет собой наиболее важный эволюционный определяющий фактор, особенно при возникновении новых тенденций.
Э. Майр
8.1. Появление эукариот
Ранее мы обсуждали вопросы развития биотического круговорота на основе прокариот. Оказалось, что главные достижения в совершенствовании обеих ветвей цикла как синтетической, так и деструктивной связаны с улучшением энергетики, повышением энергетической эффективности работы клеток. Можно сказать, что на этих этапах развития круговорота на энергетику работала биохимия и, по-видимому, практически «выдохлась» в своих возможностях, совершив все, что могла.
Различие в структурах прокариотных клеток и гораздо более сложно организованных эукариотных так велико, что считается одним из самых больших разрывов, скачков в эволюции. Случайный переход прокариотной клетки в эукариотную на основе накопления малых мутаций под действием отбора — менее вероятное событие, чем возникновение первичных протоклеток.
Если говорить о дивергентной эволюции, случайном расхождении признаков, то без привлечения представлений о целесообразности возникновения сложной эукариотной клетки обойтись нельзя. Приходится поневоле прибегать к телеологическим концепциям, типа «стремления к самосовершенствованию структур», самоусложнению и саморазвитию, от которых остается только небольшой шаг до скрытого витализма. Пожалуй, единственным направлением, твердо стоящим на материалистических позициях, является концепция последовательных эндосимбиозов, по которой эукариотные клетки есть комплексы прокариотных. Тогда эволюцию эукариот можно рассматривать как естественное продолжение эволюции прокариот, объединенных в ассоциации в эукариотной клетке.
Учет энергетического подхода позволяет снять всякий налет телеологичности с этого крупнейшего шага эволюции жизни на нашей планете. Рассматривая последовательные этапы симбиогенного развития, особое внимание мы будем обращать на два обстоятельства, очень важных с точки зрения энергетического подхода. Во-первых, в соответствии с ЭПЭР — расширение ареалов и увеличение потока энергии через биосистемы. Во-вторых, в соответствии с ЭПИР — улучшение качества функционирования биосистем: выполнение той же энергетической функции или увеличение потока энергии с помощью все меньшего количества вещества, которого всегда не хватает. Это связано: а) с замедлением размножения; б) с утратой ненужных структур; в) с организацией дополнительных циклов вещества. Итак, перейдем к рассмотрению последовательных эндосимбиозов.
1. Симбиоз первый — образование дигеномной структуры. В соответствии с принципом постепенности наиболее вероятно, что первая эукариотная клетка была гетеротрофной по своей функции в круговороте. (Хорошая аналогия с возникновением первых прокариот!) Для ее происхождения достаточно сожительства только двух типов клеток: малых прокариотных клеток, выполняющих энергетические функции и превратившихся в современных митохондрий, и большой клетки, анаэробной клетки хозяина типа современных ацидотолерантных термоплазм или микоплазм [Маргелис, 1983]. Возможно, что ближайший родственник протоэукариотной клетки — это ныне живущая крупная анаэробная амеба. Естественно, что симбиоз был выгоден обоим партнерам. Хозяйская клетка, создавая удобную среду обитания малых клеток, тем самым способствовала их сохранению и развитию, а малые клетки давали энергию (прежде всего в виде АТФ) для активного существования хозяина. С точки зрения интенсификации функции и сохранения структуры союз оказался «счастливым», по выражению Л. Маргелис [1983, с. 201]. Дальнейшее развитие симбионтов шло по пути коэволюции. И одним из существенных направлений этой совместной эволюции было избавление от избыточных структур, утрата ненужного дублирования. «Общая тенденция,— пишет Л. Маргелис,— состоит в том, что любой важный метаболит или фермент, необходимый как хозяину, так и симбионту и синтезируемый вначале обоими, будет со временем поставляться только одним из партнеров, обычно хозяином» [Там же, с. 186]. По мере развития симбиоза все больше метаболических функций «передоверяется» ядру клетки хозяина. Снижение доли сателлитной ДНК, или редукция генома органеллы, может даже считаться мерой совместного прогрессивного развития. Утрата автономии, таким образом, явно прогрессивна. Например, доля ДНК митохондрий у большинства эукариот по отношению к ядерной ДНК составляет лишь около 5%, то же может быть и для ДНК хлоропластов.
Одним из самых серьезных доводов против симбиотической теории считалось отсутствие фагоцитоза или пиноцитоза в мире прокариот. Полагалось несомненным, что внутрь прокариотной клетки не может проникнуть другая клетка. И вот в последние десятилетия были обнаружены бактерии рода бделловибрио, которые являются облигатными хищниками, питающимися другими бактериями! Причем оказалось, что они широко распространены в микробном мире и играют заметную роль в биосфере — выполняют задачи санитаров, разрушая микробные клетки в загрязненных водоемах. Переход от антагонизма к сосуществованию более «мирного» типа — вещь, часто встречающаяся и хорошо изученная.
Вполне возможно, что симбиотические отношения возникали в эволюции неоднократно. Симбиоз мог распадаться, особенно на ранних этапах сосуществования организмов, если отдельное развитие было более эффективным. Но союз митохондрий и нуклеоцитоплазмы оказался не только счастливым, но н «вечным». Особо подчеркнем его энергетические преимущества для обеих сторон. Анаэробная нуклеоцитоплазма была наименее специализированным и более крупным предшественником. Но для защиты ею ДНК от высокой температуры и кислотности у нее имелись белки особого класса — гистоны. Анаэробный хозяин мог использовать только экзогенные сахара, гликолитическим путем расщепляя их до трехуглеродных соединений и получая только две богатые энергией молекулы АТФ из каждой молекулы сахара. Эти трехуглеродные «отходы» стали пищей для митохондрий, которые способны расщеплять их до углекислоты и воды, получая гораздо больше энергии через цикл трикарбоновых кислот и систему транспорта электронов (по 18 молекул АТФ из каждого трехуглеродного фрагмента). Общий итог кооперации — 38 энергетических единиц на единицу субстрата (38=18·2+2) — оказался баснословно выигрышным. Такова энергетическая формула «счастья». Поэтому для хозяйской клетки было выгодно оберегать приобретенные фабрики энергии, давать им возможность эффективно трудиться, избавляя их от дополнительных функций.
2. Симбиоз второй — образование тригеномной структуры (наиболее спорный аспект последовательных симбиозов). По-видимому, он имел место после первого симбиоза крупной анаэробной амебоидной клетки с мелкими аэробными, т. е. с протомитохондриями. Второй акт симбиоза заключался в объединении такого митохондриально-цитоплазменного комплекса со спирохетами или спироплазмами, которые прикреплялись к этому комплексу для питания. Спирохеты обладали микротрубочками, которые в результате совместной эволюции составили основу структур клетки хозяина, появился митоз.
Многое в этой гипотезе спорно и многое не доказано, но привлекает в ней опять же полное отсутствие телеологичности. Мы не будем разбирать более подробно схемы развития этого типа симбиоза. Подчеркнем некоторые важные аспекты с позиций энергетического подхода. Во-первых, приобретение подвижности в результате этого симбиоза, способности к быстрому перемещению оказалось очень эффективным для поиска пищи и проникновения в новые места обитания, что соответствует ЭПЭР.
Во-вторых, что более существенно, мы уже говорили об энергетическом преимуществе первого симбиоза. Клетки, его осуществившие, получили возможность укрупниться, стать больше по размерам, чем каждый прокариотныи партнер, т. е. «улучшить» отношение поверхность — объем, и, в частности, резко повысить компартментализацию структуры. Это привело к возможности разделения функций, т. е. к повышению специализации. Но возрастание размеров клеток и разнообразие их компонентов потребовало и большего количества кодировавших их нуклеиновых кислот, особенно ДНК. Поэтому и понадобились эффективные механизмы равного распределения ее между дочерними клетками. Наиболее экономичной по веществу и энергетике оказывается стратегия выработки контролирующих деление клетки систем, и, таким образом, очевидно возрастание роли информационных систем на этом этапе.
3. Симбиоз третий — это очередной этап в серии последовательных симбиозов, в результате которого в клетке развился тетрагеном. Приобретение способности к фотосинтезу произошло в результате объединения сформировавшейся в двух первых симбиозах эукариотной клетки и фотосинтезирующих прокариотных клеток. Формула эукариот-фотосинтетиков проста: эукариот + фотосинтезирующая прокариота = водоросль или растение [Маргелис, 1983].
Улучшение энергетики, вернее, приобретение новой функции чисто энергетического плана — фотосинтеза в результате такого объединения совершенно очевидно.
4. Симбиоз четвертый, пятый и т. д.— образование полигеномпых организмов. К настоящему времени все яснее, что симбиозы имеют гораздо большее значение в эволюции, чем это было принято считать. Симбиоз, по утверждению Л. Маргелис, как объединение и слияние различных особей может рассматриваться в качестве одной из форм парасексуальности — объединения и слияния отдельных особей. Он не менее, а может быть, и более важен для эволюции, чем половой процесс. У организмов, возникших путем слияния двух особей (или их клеток) в результате полового процесса, родители имеют очень близких предков; а при симбиозе предки гораздо более отдалены. И самое главное — они могут выполнять несхожие, дополняющие друг друга функции. Очень существенно, что стабильные симбиотические ассоциации обладают приспособленностью большей, чем каждый партнер в отдельности. Это означает, что симбионты могут оставлять больше потомства при совместном, чем при раздельном, развитии и естественный отбор действует на их гены как на единый геном. Функциональная гибкость симбиоза заключается в том, что взаимодействие партнеров не является абсолютно жестким, если оно не столь выгодно, оно может регулироваться соотношением доли партнеров, вплоть до полного распада ассоциации. Особенно часто это возможно, если союз «молод» по времени и путь совместной эволюции был недолгим. Это хорошо видно на примере симбиозов последующих номеров, типа союзов эукариот с эукариотами. Кроме того, разделение функций может быть столь полезным и выгодным, что основу ассоциации может составлять обмен информацией, а не веществом.
В целом широко распространены и поддерживаются симбиозы, связанные с освоением новых источников пищи и энергии, недоступных ранее организму хозяина. Хорошо известны симбиотические микроорганизмы, культивируемые высшими животными, например жвачными или термитами. Это дает возможность использовать клетчатку как источник органического углерода. Симбиозы получаются уже пятого, шестого и последующих уровней кооперации. Используются симбионты и для информационных целей: так, светящиеся бактерии в определенных органах водных хищников хорошо выполняют функцию фонарей, освещающих возможные источники пищи.
Симбиотические ассоциации, перенос генов и удержание чужеродных органелл открывают колоссальные возможности для эволюции и для развития биотического круговорота как в глобальном биосферном варианте, так и в локальных экосистемах, осуществляющих местные круговороты вещества под влиянием потока солнечной энергии. В широком смысле слова можно утверждать, что весь круговорот в целом является (на каждый момент его существования) симбиозом видов, его осуществляющих. Взаимозависимость видов при их коэволюции возрастает. Достаточно напомнить, что человек и некоторые животные не способны синтезировать ряд витаминов и даже более существенных компонентов питания — незаменимых аминокислот. Как мы обсуждали в предыдущих главах, с точки зрения энергетической гораздо более выгодно не синтезировать эти аминокислоты самим хищникам, а «передоверить» это дело жертвам, способным к более быстрому биосинтезу.
8.2. Расцвет многоклеточных организмов
Для дальнейших ступеней развития биотического круговорота, связанных с образованием и повсеместным распространением эукариот, более характерны процессы дифференциации и специализации. На этих этапах эволюции жизнь освоила новые территории, и прежде всего сушу. Она распространялась не только вширь, но и вглубь. Многоклеточные эукариотные организмы стали доставать питательные вещества буквально «из-под земли», перекачивая их с потоками воды высоко вверх, в зону активного фотосинтеза с разветвленной фотопринимающей поверхностью. Удлинялись и разветвлялись трофические цепи, цели питания в гетеротрофной части цикла, заменяя недостаточно эффективные цепи разложения. Возрастал срок жизни биологических структур, но поток энергии, прокачиваемой через них, не только не уменьшался, но и заметно увеличивался. Этому способствовало совершенствование систем адаптации и прогнозирования, т. е. информационных систем (органов передачи и приема информации) , в конечном счете приведшее к возникновению и развитию разума, позволяющего не только адекватно реагировать на изменения окружающей среды, но и активно ее перестраивать.
Итак, мы уже знаем, что уже на уровне прокариот были исчерпаны возможности чисто биохимического совершенствования функционирования живых систем. С развитием эукариот открываются новые возможности ускорения метаболизма через специализацию функций внутри клетки, через развитие специфических органелл (митохондрии, хлоропласты, вакуоли и т. д.). Это объединяется одним сложным термином — «компартментализация», т. е. разделение клетки на отсеки, каждый со своей функцией.
Эукариотная клетка по структуре и по размерам сложнее и больше, чем прокариотная. Увеличение размеров оказалось возможным при накопления кислорода в атмосфере из-за энергетического скачка и в ветви цикла фотосинтеза, т. е. продукции, и в ветви дыхания, т. е. деструкции. Но с ростом размеров клеток все более существенным «узким местом» становятся диффузионные ограничения. Показателем дальнейшего прогресса выступает увеличение скорости диффузии, скорости переноса энергии и метаболитов. Как пишет С.Э.Шноль в книге «Физико-химические основы эволюции» [М., 1979, с. 142], «прогресс теперь можно измерять в сантиметрах в секунду».
Одним из наиболее эффективных путей увеличения потоков может служить относительное изменение поверхностей, через которые осуществляется диффузия. Поэтому изменение геометрии структур организма вместе с дифференциацией функций составляющих его клеток становится предметом действия естественного отбора. И это очевидно. Например, шар, наиболее обычная, но далеко не единственная для прокариот форма, оказывается теперь наименее эффективным, так как отношение его поверхности к объему минимально. Наиболее выгодна с этой точки зрения, нитевидная форма, но усложняются связи между ее концами из-за тех же диффузионных ограничений. По-видимому, плоские и кольцевые поверхности разнообразных проявлений могли удовлетворять большинству диффузионных ограничений. Перспективны также многолучевые структуры с разнообразными выростами. В этом параграфе мы и рассмотрим некоторые этапы морфологической эволюции, приведшей к современной биосфере, соблюдая как всегда принцип постепенности и заботясь о демонстрации энергетических преимуществ, получаемых организмами в эволюционном развитии.
Один из наиболее интересных этапов развития биотического круговорота связан с тем, что жизнь, возникшая в воде, захватила сушу. Первая принципиальная проблема, которую должна была преодолеть жизнь для распространения на сушу, это защита от губительных ультрафиолетовых лучей. Вода, где жизнь развивалась до сих пор, достаточно хорошо поглощала коротковолновое излучение Солнца. С увеличением содержания O2 в атмосфере за счет работы цианобактерий озоновый слой, поглощающий УФ-лучи, становился все более эффективным «щитом» для живых систем. Это позволило интенсифицировать фотосинтез у поверхности воды, на мелководьях, что дало еще более надежный озоновый экран, и т. д. Следовательно, реальные предпосылки использовать сушу появились, прибрежная жизнь стала искать пути ее захвата, перебирая варианты, делая множество попыток, чаще всего неудачных.
В соответствии с принципом постепенности очевидно, что колонизация суши происходила в направлении от берега к наиболее сухим, безводным участкам. Поскольку условия для жизни животных на суше создавались растениями, то мы и рассмотрим вначале развитие растений.
На поверхности практически не освоенной жизнью суши в протерозое в результате взаимодействия абиотических и биотических условий формировалась почва как особое биокосное тело. Биотическая часть создавалась бактериями и синезелеными водорослями. Возможно, имелось что-то типа цианобактериального мата. Существовал и развивался примитивный круговорот, менее эффективный, чем в толще воды. Неэффективность его связана с тем, что фотосинтезирующая пленка на суше очень тонка из-за ограничения подвижности фотосинтетиков (по сравнению с водой). Это же ограничение подвижности мешало снабжению фотосинтеза соответствующими биогенами. Таким образом, и поток солнечной энергии плохо использовался, и слабо включалось вещество в круговорот. Увеличить светоприемную поверхность и достать питательные вещества «из-под земли» могли только крупные многоклеточные организмы.
Каковы же были непосредственные задачи, которые должны были «решить» растения, чтобы завоевать манящую, богатую энергией сушу? Или точнее, чтобы снять налет телеологичности, скажем: какие направления их развития, связанные с умощнением и ускорением вращения вещества под влиянием потока энергии, были поддержаны естественным отбором? Перечислим несколько основных.
1. Увеличение светоприемпых поверхностей.
2. Развитие систем всасывания органических веществ из глубины почвы.
3. Обеспечение транспорта необходимых веществ и воды к специализированным системам.
4. Совершенствование прямостояния в условиях гравитации, которой практически не было в воде.
5. Борьба за воду, которая уже не омывает поверхность растения, а интенсивно испаряется с его развитых поверхностей.
Добавим к этим пяти задачам, направленным на интенсификацию процессов биосинтеза, еще несколько, связанных с размножением и сохранением организмов.
6. Повышение устойчивости к экстремальным колебаниям большинства факторов среды: температуры, влажности, ветра и т. д. Их размах гораздо выше, чем в водных условиях.
7. Обеспечение сохранения потомков в ряду поколений в условиях отсутствия воды.
8. Быстрое распространение генеративных клеток по поверхности земли в поисках новых подходящих условий.
На все это накладывалось требование экономии вещества, которого как всегда не хватало, в том или ином виде. Ведь даже возможные его запасы в почве могли быть практически мгновенно использованы, как только до них добирались живые системы, способные к автокатализу, т. е. к взрывному распространению.
Каковы же могли быть наиболее существенные шаги эволюции высшей растительности от момента ее выхода на сушу и до наших дней, кто оказался победителем и почему? Рассмотрим это чуть подробнее в соответствии с хронологией.
До выхода на сушу в водной среде существовали многоклеточные водоросли, среди которых имелись и прикрепленные формы. Однако их нельзя было назвать многоклеточными организмами, так как заметного разделения функций среди клеток не было, их прикрепление ко дну было чисто механическим, без подачи питательных веществ из донных отложений. Первые наземные растения — псилофиты — появились в конце ордовика и начале силура (500–440 млн лет назад). Понятно, что они могли существовать вблизи воды, окаймляя зеленым ковром влажные побережья морей и внутренних водоемов. По морфологии они занимают промежуточное место между сосудистыми растениями и водорослями. У них уже имеется примитивная система водоснабжения: появляется проводящая сосудистая система с примитивными флоэмой и ксилемой.
Но во многом они еще близки к водорослям, тело практически не дифференцировано на вегетативные органы, нет защиты от испарения, специализация только-только разворачивается.
В девонской флоре господствующее положение занимают леса хвощей, плаунов, папоротников. По сравнению с псилофитамш у них существенно развивается корневая система, усложняются и совершенствуются проводящие системы, имеются покровные и механические ткани. Они размножаются спорами, которые хотя и могут обходиться без воды, но нуждаются во влажных условиях. В карбоне и перми появляются и в дальнейшем процветают голосеменные, производящие семена и пыльцу. Их преимущество заключается в том, что в семенах зародыш надежно защищен от высыхания и обеспечен пищей для прорастания. А мельчайшая пыльца может разноситься ветром на большие расстояния. Их представители — хвойные. И в наше время в условиях большого города, в лесной зоне Сибири можно видеть после дождя на асфальте лужицы, окаймленные светло-желтой полоской. Это пыльца сосны.
Иссушение климата, захватившее мезозой (230–137 млн лет назад), привело к вымиранию папоротниковых, давших огромные толщи каменного угля, развитию различных форм голосеменных и появлению покрытосеменных растений (цветковых). Последние и оказались победителями в конечном счете: в наше время среди 500 тыс. существующих видов растений насчитывается более половины цветковых (300 тыс. видов). Появление цветковых растений, по мнению ряда ботаников, считается одним из важнейших этапов эволюции жизни на Земле. Время их массового развития — середина мелового периода — иногда называют началом новой эры в жизни нашей планеты. В чем же заключаются преимущества покрытосеменных с точки зрения функциональной? Это прежде всего интенсификация главной функции — фотосинтеза и, следовательно, быстрый рост и развитие. Они успевают завершить жизненный цикл в коротких благоприятных условиях ранней весны в пустыне и позднего лета в приарктической тундре. Основу этого прогресса составляет высокоразвитая система транспорта необходимых неорганических веществ от корней к листьям и раствора глюкозы от листьев к корням.
Не следует забывать и еще об одном преимуществе покрытосеменных (оно связано с самим названием), а именно об образовании семян, хорошо приспособленных к сохранению и распространению в разнообразных, подчас суровых условиях земной поверхности.
Заслуживает отдельного упоминания еще одна особенность цветковых растений, сумевших глубоко вписаться в круговорот: имеется в виду привлечение насекомых для опыления цветов. Это было выгодно по крайней мере в двух направлениях. Во-первых, перекрестное опыление и увеличение пластичности потомства, а во-вторых, уменьшение трат на выработку генеративных клеток (пыльцы) по сравнению, например, с использованием ненаправленных переносчиков (ветра) у других растений. Большую приспособленность цветковых хорошо продемонстрировали оледенения кайнозойской эры.
Палеонтологическая летопись истории животных известна лучше, чем для других таксонов, но следы царства животных в древних ископаемых остатках ранее 800 млн лет не обнаруживаются. Первые многоклеточные животные были исключительно морскими, так как растения еще не колонизировали сушу. Как мы знаем, для обеспечения энергией, прежде всего из-за слабой диффузии кислорода ,в воде, первичные животные должны были иметь хорошо развитые поверхности. Снабжение пищей было также одной из главных задач, поэтому необходимо было иметь отростки, способствующие перемещению либо самого животного в воде, либо воды относительно тела животного. Хорошим примером простых животных этого типа являются представители эдиакарской системы. Это мягкотелые морские беспозвоночные конца докембрийской эры и начала кембрия (670–550 млн лет назад). Вначале они были найдены в холмах Эдиакара в Австралии, а затем — на всех пяти континентах. В нашей стране детальное изучение ископаемых остатков этих животных проведено академиком Б. С. Соколовым [1984].
Для организмов этого типа характерны медузоподобпые плавающие формы либо прикрепленные колониальные формы типа морских перьев; имелись и морские черви, и беспапцирные членистоногие животные. Все они получали кислород из окружающей среды, поглощая его через эпителий. Это и ограничивало возможности их развития по энергетике. Может быть, расцвет их связан с тем, что хищных форм в то время не могло существовать все по тем же причинам: давление кислорода, около 7% от современного содержания в атмосфере, было недостаточным для развития активного, а потому и энергоемкого хищничества.
Поэтому активное снабжение кислородом было одной из главных задач эволюции в этой ветви круговорота, к ней же примыкают и развитие органов движения, и совершенствование скелета. Все это и приводило к разделению функций и специализации органов. Первые позвоночные, так называемые щитковые, появились около 450 млн лет назад. Дальнейшая специализация к плаванию и активному захвату пищи привела к развитию вначале бесчелюстных, а затем и челюстноротых рыб. В связи с развитием растительности в пересыхающих мелких водоемах появились и двоякодышащие рыбы. От них — прямой путь к амфибиям, а следовательно, и дорога на сушу.
Многие из первых амфибий были довольно крупными и занимали местообитания на суше и на мелководьях, среди них — и растительноядные, и хищники. Но они были тесно связаны с водой, где происходило их размножение. В карбоне (300 млн лет назад) появляются первые пресмыкающиеся, и можно считать, что суша окончательно «завоевана» жизнью. Рептилии, имеющие сухие прочные покровы, откладывающие яйца, покрытые твердой скорлупой с запасом питательных веществ, сделались независимыми от воды. В это же время жизнь начинает осваивать и воздушный бассейн, развиваются древнейшие группы беспозвоночных — насекомые, среди которых значительная доля — летающие.
Рептилии, вытеснившие своих предшественников — амфибий, дали много различных форм и долгое время были господствующей группой, освоившей воду, сушу и воздух. Только похолодания климата привели в кайнозое к расцвету теплокровных млекопитающих н птиц, а также насекомых. Кстати, по энергетике насекомые практически не уступают гомойотермным животным.
Оледенения кайнозоя приводили к неоднократному изменению видов, составляющих экосистемы каждого периода, однако в целом сложившийся тип биотического круговорота сохранился до нашего времени.
Отметим, какие магистральные направления развития животных имели место в макроэволюции.
1. Развитие многоклеточности и большая специализация функций различных органов многоклеточного организма; особо выделим развитие органов дыхания.
2. Повышение энергетической активности и подвижности и параллельное развитие опорной системы (наружный скелет у членистоногих и внутренний — у позвоночных) .
3. Совершенствование способности к терморегуляции, завершившееся развитием гомойотермии, что позволило теплокровным животным активно развиваться в условиях похолодания и распространиться в холодные зоны.
4. Развитие центральной нервной системы с основным регулирующим центром — мозгом.
5. Появление социальности в ряде ветвей древа животных, что привело в конечном счете к развитию социальной формы движения материи, относящейся лишь к одной из ветвей приматов — человеку.
Последние два пункта тесно связаны с информационным аспектом эволюции жизни, и мы их рассмотрим отдельно в следующем параграфе. Из первых трех пунктов, имеющих функционально-морфологическое значение, наиболее тесно связанным с энергетикой является третий. Он же и наиболее дискуссионный в оценках с различных точек зрения. Например, даже современная термодинамика открытых систем «не разрешила» бы появление и существование избыточного теплорассеяния при гомойотермии, если бы последняя не появилась раньше самой термодинамики. Возникновение гомойотермии — очевидное преимущество с точки зрения энергетики. Конкретные шаги становления теплокровности в эволюции остаются неизвестными. По-видимому, этот процесс во времени носил взрывообразный характер, так как медленное, постепенное развитие его натыкается на ряд запретов.
В эволюции животных очень важным оказалось развитие информационно-управляющих структур. По типу функционирования или, точнее, по месту организма в цепи питания его можно отнести к одному из четырех уровней [Шмальгаузен, 1946 (с добавлениями по энергетике)].
1. Нижний уровень занят организмами с наименьшими адаптивными способностями, у которых единственный способ защиты от хищников — это увеличение скорости размножения. Он характерен для одноклеточных и имеет корни в совершенной биохимии прокариот.
2. Следующий уровень образуют организмы с пассивными способами защиты. Это животные с прочными покровами типа специальных домиков или высшие растения, имеющие биохимическую защиту либо колючки.
3. На третьем уровне — способные к активному перемещению животные, животные-жертвы, спасающиеся от агрессоров благодаря способности к быстрому перемещению (и к активному восприятию и переработке информации).
4. Вершина занята животными, которые сами являются хищниками. На третьем и четвертом уровне очень многое зависит от индивидуальных способностей отдельных животных. Здесь элиминация в отличие от первых двух уровней становится высокоизбирательной. Она очень сильно зависит от адаптационных возможностей каждого отдельного организма, от его умения воспринимать информацию об окружающем мире и быстро реагировать в конкретных ситуациях. Отбор на этих уровнях связан с повышением адаптивности систем.
Что же такое адаптация? Этот термин очень емкий, его можно считать универсальным свойством живого, для организма он означает процесс непрерывного приспособления к меняющимся условиям среды. Сформированные в эволюции генетические программы обеспечивают адаптацию к постоянно действующим факторам среды, основываясь на биохимическом потенциале. Однако реакции, организуемые на основе этих программ, недостаточно гибки, и в эволюции отобрались более или менее универсальные механизмы, регулирующие их работу. Информационный аспект здесь более важен.
Коротко рассмотрим эволюционное развитие цепи обратной связи в одной из главных информационно-управляющих систем: рецептор — глаз; переносчики и переработчики информации—нервная система и мозг; исполнитель — мышца.
На примере развития глаза можно хорошо видеть, как функция «диктовала» свои условия совершенствованию структуры. С позиций только морфофизиологического подхода объяснение существования такого органа высокой степени сложности казалось очень непростым даже самому основателю дарвинизма. В одном из разделов книги «Происхождение видов...» (глава «Затруднения, встречаемые теорией») он писал:
«Предположение, что глаз со всеми его неподражаемыми приспособлениями для изменения фокусного расстояния по мере удаления предмета, для регулирования количества проникающего света, для поправки на сферическую и хроматическую аберрацию мог быть выработан естественным отбором, может показаться, сознаюсь в том откровенно, в высшей степени нелепым». Действительно, великолепное совершенство глаза (а электронная микроскопия в последнее время подтвердила это и для молекулярных структур) интуитивно связывается с его созданием по какому-то специальному предопределению. Тем более, что в живой природе глаз возникал неоднократно! Создается впечатление, что он был запланирован, предопределен ранее (может быть, чтобы жизнь могла увидеть самое себя?!).
Информационно-энергетический подход позволяет и в данном случае преодолеть ограниченность структурного подхода, снять налет телеологичности.
Следует обратить внимание на то, что для глаза, как и для других органов информации, характерно чрезвычайно малое потребление энергии при работе. Наш светоприемник при адаптации способен воспринимать даже отдельные кванты света.
Эволюция глаза как светопринимающего устройства характеризовалась постепенным накоплением усовершенствований. В самом начале на основе фоточувствительных молекул примитивные организмы умели лишь отличать свет от темноты. У некоторых одноклеточных имеется пигментированное светочувствительное пятно, как правило на переднем конце тела. Известна отрицательная реакция инфузорий на видимый свет и особенно на ультрафиолет. Имеющая хлоропласты эвглена, напротив, плывет к свету.
Развитие многоклеточности «позволило» усложнить и строение светопринимающих устройств. Уже у плоских червей имеются примитивные глаза — чашечки со зрительным пигментом родопсином. Примечательно, что функции светочувствительных клеток взяли на себя молекулы родопсина, которые являются ближайшими родственниками древнейшего белка бактериородопсина.
Строение глаза все более усложняется с ростом уровня организации животных. Образуется глазной пузырь с жидкостью и подвижным хрусталиком — линзой. Чувствительность и разрешающая способность глаза еще более возрастает с образованием сетчатки — слоя светочувствительных клеток. В дальнейшем светочувствительные возможности возрастают с увеличением кривизны хрусталика, развитием зрачка, глазной мускулатуры, разделением светочувствительных клеток на колбочки и палочки и т. д.
У членистоногих развился другой тип глаза — фасеточный. Отдельная фасетка имеет ряд светочувствительных клеток и неподвижную линзу. Увеличение числа фасеток в сотни и тысячи раз повышает разрешающую способность такого глаза. Для малых организмов, особенно для летающих форм, по-видимому, было выгоднее варьировать число мелких фасеток, чем создавать единую, довольно крупную структуру с глазным пузырем и хрусталиком.
Таким образом, можно видеть, какой путь прошел глаз в эволюционном развитии под действием естественного отбора на улучшение функции.
Может быть, еще более наглядным примером действия отбора на изменение структуры служит элиминация структур, если они не нужны. И здесь хорошо известны случаи упрощения или даже редукции (потери) глаза в тех условиях, когда он не нужен. Один из самых впечатляющих примеров был описан обозревателем газеты «Комсомольская правда» [6 июля 1985 г., № 18357] В. Песковым. В маленьком пруду (Бабынивский район Калужской области), вырытом 15 лет назад, были обнаружены безглазые караси, наряду с обычными. Имелись и одноглазые формы, и образцы с более выпуклыми глазами. Чем можно объяснить безглазие, развившееся так быстро? Прежде всего, именно этот вид карася имеет нестабильный генетический механизм передачи признаков и широкую вариабельность структур (от него выведены разнообразнейшие по форме и окраске аквариумные рыбки).
В данном случае безглазие оказалось даже не уродством, а выгодным преимуществом. В мелком илистом пруду зрение не нужно; для копания в иле важнее обоняние. По сведениям В. Пескова, обоняние у незрячих рыб хорошо развито. И главное: в пруду нет хищников — ни щуки, ни окуня, так что на всех фазах развития глаз карасям не требуется. Без этой лишней структуры безглазые караси развиваются быстрее зрячих. Преимущество освобождения от ненужной структуры здесь очевидно.
Продолжим описание нашей информационно-управляющей системы. Теперь очередь за рассмотрением переносчика информации — нервной системы и ее центра — мозга, перерабатывающего эту информацию и вырабатывающего сигнал управления. Для совершенствования адаптационных способностей организма это, пожалуй, самые важные системы.
Совершенно очевидно, что должен существовать центральный анализатор сигналов от зрительного и других рецепторов, который принимает решение и вырабатывает управляющий сигнал исполнителю (мышцам). Развитие мозга было прослежено еще в прошлом веке, и цефализация организмов считалась одним из важных критериев эволюции. Доля нервных и мозговых клеток в эволюции возрастала, причем характер этого роста во времени имел экспоненциальную природу.
Заметим, что работающий мозг потребляет энергии незначительно больше, чем отдыхающий, что говорит о его экономичности по энергетике.
И опять достойно упоминания то, что при ненужности функции даже эта совершенная структура может быть потеряна полностью. Примером тому служат паразитические черви, в условиях полного обеспечения пищей утратившие голову вместе с мозговыми клетками.
Теперь очередь за третьим компонентом системы — исполнительным органом. У высших животных это мышца. Совершенно очевидно, что чем быстрее организм достигнет источника пищи или избегнет опасности, тем больше вероятность его выживания, выше уровень его адаптации к условиям окружающей среды. И здесь природе пришлось вырабатывать разнообразные пути решения. Движение с помощью жгутиков было освоено уже на уровне одноклеточных прокариот. Сам бактериальный жгутик вращается в мембране, его основание — базальное тельце — крутится подобно ротору электродвигателя, что обусловлено градиентом концентрации ионов водорода. Аналогия с электродвигателем довольно глубокая, так как в обоих случаях имеется вращающееся электромагнитное ноле. Природа изобрела колесо давным-давно, только его размеры оказались малыми, и оно не смогло эффективно работать. Ускорение движения одноклеточного эукариотного организма могло быть обеспечено и обеспечивается увеличением числа ресничек, но предел тут достигается довольно быстро, и он имеет диффузионную основу (например, диффузия топлива к месту работы). Задача могла быть решена только на уровне многоклеточных, с развитием специализированных клеток. Удлинение движущегося рычага, помещенного в среду, содержащую топливо (типа АТФ), образование «сократительной нити» — вот путь совершенствования мышцы, нового аппарата перемещения организма в пространстве, по С.Э.Шнолю.
И здесь существенно представление о регрессе структуры. На собственном примере каждый из нас может убедиться, как быстро может потерять объем и эластичность мышца, если ее не нагружать. Проблема становится особо острой в условиях невесомости. Для длительных полетов пришлось разработать сложные программы интенсивных нагрузок.
Конечно, вся система обратной связи развивалась согласованно. Так, скорость физической реакции на раздражение не должна была сдерживаться, например, скоростью распространения нервного импульса, и той «пришлось» возрасти от 0,1 до 150 м/с, приблизившись к скорости звука.
Проведенное рассмотрение информационно-управляющей системы наглядно демонстрирует первичность функции перед структурой. Вывод этот не имеет оттенка телеологичности, если не забывать о движущих энергетических потоках, инициирующих круговороты. Сам вывод не противоречит заключениям эволюционистов, особенно если речь идет о конвергентном сходстве, казалось бы, далеких по происхождению структур. Примечательно в этом смысле высказывание известного эволюциониста Э.Майра [1968, с. 403]: «Мир животных, так же как и мир растений, полон конвергенции, когда сходные требования среды вызывают сходные фенотипические реакции у неродственных или по крайней мере не близкородственных организмов. Если существует только одно эффективное решение для данной функциональной проблемы, то весьма различные генные комплексы дадут одно и то же решение независимо от того, сколь бы ни были различны избранные пути. Поговорка, что „все дороги ведут в Рим”, столь же верна, как и в житейских делах».
И над всем этим витает «дух» энергетического (но не энтропийного) совершенствования. Можно считать, что развитие систем кодового управления хорошо соответствует действию энергетических принципов: увеличение активности организмов (в том числе и по захвату энергии, и по относительному уменьшению), экономии трат на поддержание структуры (вспомним потерю головы паразитом, если она не нужна).
В целом анализ развития информационно-управляющих систем животных хорошо демонстрирует важность энергетических характеристик в эволюции животных, особенно на верхних ступенях эволюционной лестницы. Для этого уровня становятся существенными социальные функции и структуры.
8.3. «Самоорганизация» на уровне популяций
Уделяя особое внимание морфофизиологическим структурам отдельного организма, изучая громадное, почти непостижимое разнообразие форм, очень легко оказаться в плену организмоцентризма, т. е., грубо говоря, считать, что структура организма и органа определяет его функцию. В предыдущем параграфе, говоря об особенностях морфологических и информационных структур организмов, мы старались подчеркивать ведущую роль функции. Но особенно ярко приоритет функции выступает, если подняться на следующую ступень и рассматривать популяционный уровень отношений организмов.
В связи с этим заслуживает отдельного рассмотрения поведенческий аспект в эволюции животных. Социальная сторона его очевидна, так как практически вся жизнь животных связана с их отношениями в популяциях. Роль поведения в эволюционных сдвигах отмечалась эволюционистами разных направлений, для пас важно проследить, как с его совершенствованием «улучшается» энергетика популяции (и количественно. и качественно). Как отмечает Э. Майр, выбор нового биотопа, использование новой пищи представляет собой, как .правило, новый аспект поведения и играет основную роль при переходе в новые адаптивные зоны. «Поведение представляет собой наиболее важный эволюционный определяющий фактор; особенно при возникновении новых тенденций... Большинство недавних переходов в новые экологические ниши вначале не сопровождалось структурными модификациями. В тех случаях, когда развиваются новые повадки, вслед за этим рано или поздно происходят структурные изменения» [Майр, 1968, с.401]. Таким образом, явно говорится о первичности поведения, связанного с функцией, и вторичности структуры.
Еще в начале нашего века высоко оценивал адаптивность поведения А. Н. Северцов. Он отмечал, что при быстрых изменениях условий существования в неблагоприятную сторону выживают животные — «изобретатели новых способов поведения».
Рассмотрим некоторые особенности популяционных структур высших животных. В любой группе животных, обитающих совместно на одной территории, всегда имеется определенная иерархическая структура. Она проявляется в комплексе постоянно поддерживаемых отношений доминирования и подчинения. Существуют две крайние формы иерархии. В одном случае какое-либо одно животное — вожак доминирует над остальными, между которыми нет различий по рангу. В другом устанавливается линейная иерархия: одна особь доминирует над всеми, следующая подчиняется первой, но доминирует над оставшимися вплоть до последней, которая подчиняется всем (хорошо известен быстро устанавливающийся у цыплят линейный порядок клевания при недостатке корма: вначале ест самый сильный цыпленок, за ним всегда второй и т. д.). Обычно устанавливается комбинация из этих форм, иногда возможны кольцевые связи.
Иерархия устанавливается в результате агрессивного «выяснения отношений», побеждает наиболее сильная агрессивная особь, как правило взрослый самец. Для группы это выгодно, если ее возглавляет самая сильная или самая умная особь. Для доминирующей особи это тоже очевидно. Для подавляемых особей выгода не столь очевидна, но и тут есть преимущество: нет слишком частого «битья», т. е. возможна спокойная жизнь, открывается путь к эмиграции, поиску новых местообитаний.
Можно сказать, что в целом иерархическая структура позволяет уменьшить траты на поддержание (на популяционном уровне), без нее слишком много энергии уходило бы на постоянное выяснение отношений внутри группы. После установления иерархии животные мирно сосуществуют в единой группе, занимаясь своими делами. Итак, иерархия способствует повышению активности группы, группа ведет себя как единое целое, что увеличивает ее потенциал в экологической нише, в захвате пищи (энергии) и пространства.
В чем же заключаются преимущества стадного образа жизни? Почему он так широко распространен во многих таксонах животного царства? Перечислим некоторые, имеющие функциональный аспект.
1. Повышение способности к конкуренции. Группа совместно держащихся животных уже своими размерами способна подавить конкурентов, предпочитающих индивидуальный образ жизни. В большинстве групп или локальных популяций существует разделение труда. Хорошо известны, к примеру, коллективные действия хищников во время совместной охоты (волки, львы), когда каждый зверь выполняет свою «задачу».
2. Повышение эффективности добывания пищи. Это преимущество тесно связано с предыдущим.
3. Защита от хищников. Очевидно, что защита от нападения может быть в группе гораздо более эффективной. Существуют целые системы групповой круговой обороны у копытных животных, служащие надежным средством даже при нападении крупных хищников. Высока информационная роль оповещения о приближении хищника, его «окрикивание» и т. д. Это значит, что в среднем каждая особь может больше времени заниматься активным видом деятельности, в том числе и питанием.
4. Изменение среды вплоть до активного. Самый простой пример — совместное согревание в куче животных при похолодании. Более сложный, а подчас вызывающий изумление — это совместная достройка очень сложных, с совершенной термо- и влагорегуляцией жилищ, широко распространенных у общественных насекомых. К этому же типу относится постройка бобрами плотин с регулируемым уровнем воды.
5. Повышение эффективности размножения. Прежде всего, если в стаде имеется активный самец (вожак), то он и оставит больше потомков, что выгодно и для популяции в целом. Для многих групп насекомых характерно роение, в котором участвуют особи, прилетающие издалека. Велика роль стадности и в коллективной защите потомства. Известны целые «детские сады» у стадных млекопитающих, где дети находятся под присмотром не только своих родителей. У тех же общественных насекомых имеются особые категории «нянек», кормящих потомство.
6. Обучение. Возможность ускоренного обучения в группе, пожалуй, является одним из наиболее важных преимуществ для большинства высших животных. Теперь это все более осознается специалистами-этологами и эволюционистами в целом. Развитие способности к обучению дает преимущества по всем выше рассмотренным пунктам, особенно при передаче накопленного опыта потомству. Большая информационная емкость и низкая энергетическая стоимость обучения очевидны. Можно сказать, что действие энергетических принципов проявляется здесь в полной мере. Во-первых, передается новая функциональная возможность, т. е. открывается новая ниша (пища, пространство). Это иллюстрация экспансии, работа по экстенсивному типу. Но это все совершается без образования новых структур, что является показателем интенсивного типа развития. В данном случае практически никаких трат вещества и энергии не требуется, а информация, как мы знаем, передается почти бесплатно. Она без потерь и даже с усилением циркулирует по контурам обратной связи у одной особи или переходит от особи к особи в пределах группы, популяции и даже выходит за границы одного вида. В настоящее время известны интересные примеры быстрого распространения нового «знания», изобретения, навыка. Стоило, к примеру, одной из британских синиц научиться открывать бутылки с молоком и выпивать сверху сливки, как это умение практически мгновенно распространилось среди синиц разных видов.
Все обсуждавшиеся адаптивные признаки, связанные с групповым поведением, имеются у приматов и особенно ярко выражены в ветви высших обезьян и гоминид. В середине и конце третичного периода изменение климата привело к сокращению лесных местообитаний, развитию саванн и степей. Переход от древесного образа жизни к наземному у предков гоминид привел к развитию способности перемещения на двух конечностях. Только групповой образ жизни позволил первым гоминидам спасаться от опасных нападений со стороны крупных наземных хищников. Отдельным особям это было не под силу. То же можно сказать и о коллективной добыче пищи при совместной охоте, групповой охране примитивного жилища и т. д. Следовательно, селективное преимущество стадного образа жизни, повышение организованности группы, обучение в ней, разделение труда становятся очевидными. Все это требует совершенствования средств коммуникации, выработки языка и в конечном счете интеллекта у человека. Эволюция гоминид, приведшая к развитию человека,— процесс сложный и многосторонний. Ясно, что наиболее существенное различие между человеком и животным лежит в сфере разума, а не тела.
Обсуждение эволюционной природы человека, конкретных шагов морфологической эволюции не входит в нашу задачу. Оставаясь на позициях главным образом энергетического подхода, мы только подчеркнем, что такой показатель, как использование энергии человеком, в эволюции рос с наибольшей скоростью. Об этом и пойдет речь в следующей главе.
Глава 9
Человек и энергетика
... Прогресс техники в том и заключается, что человеческий труд все более и более отступает на задний план перед трудом машин.
В. И. Ленин
9.1. Роль энергии в истории человечества
Весь длительный процесс освоения энергии человеком можно разделить, хотя бы для удобства обсуждения, на четыре-пять этапов [по Алексееву, 1983, с модификациями].
Первый — этап мускульной энергии, он уходит в глубь тысячелетий и длится до V–VII в. н. э. Одним из самых замечательных достижений этого периода является овладение огнем: вначале поддержание костра, а затем добывание огня и запасание первого энергетического ресурса — дров.
Второй этап (VII–XVII вв.) относится к использованию энергии движущей воды и ветра, он связан с изготовлением специальных, порой очень непростых сооружений, требовавших коллективного труда и творчества. Техническая основа разработок этого времени — колесо.
Третий этап (с XVIII в. до начала XX в.) соответствует все более широкому применению «движущей силы огня», источником которого является химическая энергия топлива, накопленного в былых биосферах:
каменного угля, нефти, газа, горючих сланцев и т. д.
Четвертый этап (с «XX в. наравне») не зря называют «золотым веком электричества». Благодаря его открытию, а главным образом созданию многочисленных приборов и движителей для человечества оказалось возможным освоить и энергетически обеспечить практически все уголки нашей планеты, более или менее пригодные для жизни.
Пятый этап развития энергетики, основанной на использовании энергии распада атома и синтеза ядра, практически станет определяющим только в следующем веке или (более осторожно) в следующем тысячелетии. Рассмотрим развитие энергетики человека в историческом плане.
Вспомним, что иссушение климата и уменьшение площадей лесов в конце третичного — начале четвертичного периода вынудили предков-гоминид перейти от древесного к наземному существованию. Только социальный образ жизни, развитие систем коммуникации, обучения и приготовления примитивных орудий защиты и труда позволили некоторым линиям гоминид, не имевшим ни густого меха, ни мощных когтей, твердо обосноваться в степях и саваннах, используя естественные укрытия и пещеры как убежища. Примитивные орудия труда, обнаруженные археологами вместе с человеческими останками, относятся к дальней дали веков. (Например, орудия, найденные при раскопках вблизи древнего греческого города Птолемаис в Македонии, изготовлены более 3 млн лет назад.) Конечно, первые орудия очень примитивны. Это камень с острым сколом, дубина, очищенная от сучьев, и позднее — копье, сильное смертоносное оружие. Но со временем они все более совершенствовались, с их помощью первобытный человек мог охотиться на диких животных, выкапывать съедобные корни, строить укрытия из деревьев. Сто лет назад были проведены сравнения, какой топор лучше: каменный или железный. Оказалось, что каменный топор ненамного хуже; дерево толщиной 17 см было им срублено за 7 мин, а железным — за 5.
Самым великим событием в развитии человечества на этом этапе было «приручение» огня. Точной даты нам не узнать, да ее, видимо, и не было: в различных регионах земли это происходило неоднократно и в разное время.
Недаром в мифах и легендах всех народов мира говорится о божественном происхождении огня. Огню очень долго поклонялись как божеству; не умея его добывать, старались сохранить в одном месте на долгие годы. В одной из пещер вблизи Пекина археологи обнаружили следы костра, который непрерывно горел в течение 500 тыс. лет на одном месте! Огонь зажигали от лесных пожаров, возникших от ударов молний, реже —от извержения вулканов.
Следующий огромный шаг в овладении человеком силами природы — это умение добывать огонь. Ф. Энгельс писал, что добывание огня трением впервые доставило человеку господство над определенной силой природы и тем окончательно отделило человека от животного царства. Постоянное применение огня резко изменило жизнь человека. Учеными доказано, что уже неандертальцы ели жареное мясо. Приготовление хлеба, вареного и жареного мяса существенно облегчило пищеварение, высвободив время и энергию для активных действий (т. е. в соответствии с ЭПИР доля активной энергии у человека возрастала по сравнению с другими млекопитающими; непосредственное увеличение потока использованной энергии по ЭПЭР в связи с использованием огня очевидно).
Однако прямые энергетические возможности человека определялись главным образом мощью его мускулов и крепостью костей. Средняя мощность мужчины около 0,1 л.с. Первыми машинами, преобразующими и запасающими энергию, по-видимому, можно считать самодействующие ловушки. В них использовалась сила тяжести животных (ямы) либо упругостные силы отогнутых ветвей, согнутых деревьев. Самый яркий пример облегченного варианта такой «машины» с дистанционным действием — это лук и стрелы.
На протяжении 3–4 млн лет технологическая эволюция человека протекала довольно медленно. Сотни тысяч лет, согласно данным раскопок в разных регионах Земли, традиции изготовления каменных орудий сохранялись почти неизменными. Но около 40 тыс. лет назад скорость эволюции значительно возросла. Неоценимую помощь человеку, прежде всего по энергетике, оказали первые прирученные животные и среди них — собака. Она помогала и выследить добычу, и охранять жилье, могла использоваться как тягловая сила, а в тяжелых случаях — и как подручный запас еды. Приручение различных животных, а с ними и разведение домашнего скота резко изменило весь образ жизни человека: от простого собирания «благ» дикой природы он перешел к их производству. Постепенно вместо собирания злаков он начинает их охранять, выращивать, а потом и сеять.
Десять — двенадцать тысяч лет назад с переходом к земледелию и скотоводству произошла так называемая неолитическая революция. Местом наиболее раннего ее проявления считается Передняя Азия. В этом регионе были не только необходимые дикорастущие злаки, но и животные, легко поддающиеся одомашниванию,— свиньи, козы, овцы, коровы. Сезонный сбор урожая однолетних растений вынудил человека делать запасы зерна, фуража, а отсюда и появилась возможность содержать животных. Потребовались новые орудия труда, не только каменные топоры, но серпы, мотыги, потребовались могучие быки, чтобы тянуть повозки с урожаем или перевозить домашний скарб при переселении с места на место, когда земля переставала родить. Неолитическая эволюция в течение нескольких тысячелетий распространилась по всему миру.
Используя действие огня, человек научился выплавлять из руды твердые металлы, на смену каменному веку пришел бронзовый, а за ним и железный. Неолитическая эволюция и прогресс энергетики привели к тому, что впервые в истории производство пищи и орудий труда стало постоянно превышать минимальные жизненные потребности. Появился прибавочный продукт, а с ним — собственность и государство. Уже не требовалось каждому человеку участвовать в добывании и производстве пищи. Появились квалифицированные специалисты, ремесленники, целиком занятые изготовлением определенных орудий труда и производства. Возросла роль знания, умения, специализации, расширился обмен товарами и идеями, улучшалось энергетическое обеспечение.
Изобретение колеса было одним из самых значительных изобретений этого времени (5–6 тыс. лет назад). Можно сказать, что первые государства и почти вся техническая цивилизация въехали в историю «на колесах». С их развитием потребовались новые источники энергии — простых мускульных сил не хватало. Государство, этот «аппарат насилия», позволяло решать задачу просто: заставить работать на себя другого, сделать его своим рабом, использовать его силу, брать от него больше, чем давать (таков только один из аспектов развития рабовладения — энергетический).
Мускульная сила рабов резко увеличила энергетические возможности рабовладельческих государств. Меньшинство захватило орудия производства, власть и стало эксплуатировать большинство, используя его труд и энергию,— образовались классы рабов и рабовладельцев. С помощью рабов прокладывались каналы для орошения земель и для отвода воды с затопляемых территорий. Создавались искусственные плотины, изменяющие течение крупных рек. Широко известны грандиозные сооружения, «чудо света», техники и рабовладения того времени,— египетские пирамиды. Удивляет количество труда, вложенного в них. Пять тысяч лет назад за 20–30 лет была построена одна из самых знаменитых пирамид — пирамида египетского фараона Хеопса. Ее высота соответствует высоте современного 50-этажного дома, длина 230 м. Ее возвели сто тысяч рабов из 2,3 млн блоков со средним весом 1,5 т, а некоторые—до 10–15 т. Щели между блоками меньше 0,5 см, грани пирамиды точно обращены на четыре стороны света. Какой яркий пример огромной энергетической мощности государства и... бессмысленного ее применения.
Труд рабов широко использовался в ткачестве, которое постепенно становилось одним из наиболее распространенных ремесел вплоть до создания ткацких мастерских — первых коллективов специалистов. На рабском труде было основано и тяжелое горное дело.
Рабство с течением времени стало тормозить процесс развития энергетики, как источник энергии оно изживало себя. Человек стал искать новые источники, и, естественно, что он обратил внимание на те, что всегда были перед ним: текущую воду и ветер. Мы знаем, что источником этой энергии, движителем круговорота воды и воздуха является поток солнечной энергии, но для древних людей первопричина не была особенно важной. Они уже давно эпизодически пользовались силой движущейся воды и ветра. Так мы переходим к описанию второго этапа развития энергетики, используемой человеком.
Этот этап, как и полагается по законам диалектики развития, давно вызрел в недрах первого периода. Доподлинно доказано старыми документами, что парус применялся не менее 4 тыс. лет назад, а водяное колесо, вращаемое потоком воды, насчитывает более чем двухтысячелетнюю историю.
Но широкое использование энергии воды и ветра относится к фазе повсеместного перемещения народов в Европе, к V—VII вв. н.э.
С гибелью Римской империи и с фактическим затуханием рабовладения физический труд и энергия стали цениться гораздо дороже и старая энергетическая основа — мускульная сила потеряла ведущую роль. Становление нового феодального строя связано и с развитием новой техники. Если первое документальное упоминание о водяной мельнице относится к IV в., то к XI в. их насчитывались десятки тысяч. Добавим к этому, что если лошадь в технической установке заменяла 10 рабов, то хорошее водяное или ветряное колесо — до 100. Ветряные мельницы, хотя и появились позднее водяных, тоже быстро получили широкое распространение, но из-за непостоянства энергоносителя — ветра не могли заменить более непрерывно действующие водяные. Водяные колеса совершенствовались со временем, и к XI в. для их работы использовалась даже сила приливов (в Англии, Франции и позднее, при Иване Грозном, в России, на берегу Белого моря).
Средние века как раз и характеризуются переходом от ручного производства к машинному. Создаются прядильные и ткацкие станки, маслобойные и бумагоделательные машины, металлический сельскохозяйственный инвентарь, лесопильные установки. На все это требовалось огромное количество металла, а добыча руды и угля все усложнялась. Из-за выработки древесного угля, необходимого при выплавке стали (до изобретения кокса), сводились на нет огромные площади лесов. В наиболее промышленно развитой Англии практически не оставалось лесов. Можно говорить о первом серьезном экологическом кризисе, связанном с развитием промышленности.
Но гораздо более серьезным и угрожающим был энергетический кризис. Всем новым машинам нужны были мощные, постоянно действующие движители, независимые ни от положения, ни от сезона в отличие от ветряных и водяных колес. Идея надежного двигателя недаром занимала умы мыслителей того времени.
Своеобразным отражением энергетического кризиса являются многочисленные в то время попытки создать вечный двигатель. Видя кажущееся «самодвижение» воды и воздуха (реки, приливы — отливы, ветра), легко можно было представить, что хитроумная комбинация машин способна к вечному движению, а следовательно, и к постоянному совершению работы.
Естественно, что наибольшее число «изобретений» относилось к использованию энергии воды и воздуха. Среди них наиболее популярны комбинации спирального подъемника воды — архимедова винта и обычного водяного колеса, которые вращают друг друга; колесо, вращающееся под действием неуравновешенных грузов; и т. д. Попытки создания вечного двигателя крайне заманчивы. Они не прекращаются до сих пор, правда, уже на других сочетаниях движущих сил. Еще в 1775 г. Парижская академия приняла решение не рассматривать утопических проектов вечных двигателей из-за невозможности их создания. Это — крупное достижение науки того времени, очень важна его гносеологическая роль. По сути оно означает отказ от самодвижения во всех формах, необходимость учета внешней накачки энергией всех преобразователей энергии. Не мешает напомнить, что идея самодвижения и саморазвития любых систем автоматически приводит, с энергетической точки зрения, и к возможности существования самоисточников энергии, т. е. вечных двигателей, чего, как известно, в природе не обнаружено.
Выход из энергетического кризиса средневековья был найден с помощью приручения «движущей силы огня», использования перехода химической формы энергии в тепловую, применения силы сжатого пара. Это — третий этап развития энергетики человечества. И опять мы не знаем, когда была построена первая паровая машина. Может быть, это был эолопил Герона или одна из первых паровых пушек Архимеда. Хотя древние греки и были знакомы с действием паровых машин, но объяснения принципа их действия тогдашняя схоластическая наука дать не могла. Не была известна сущность происходящих при этом физических процессов (считалось, например, что воздух превращается в пар), а без этого создать серьезную, эффективно работающую машину было нельзя.
Только научная революция XVI–XVII вв., вызванная требованиями развивающихся капиталистических отношений, привела к возникновению опытной науки, сформулировавшей правила разработки и создания разнообразных энергетических движителей.
На стыке XVII и XVIII вв. были созданы первые длительно работающие паровые машины, вначале пригодные лишь для откачивания воды из шахт (одной из самых тяжелых задач того времени). Они были громоздкими и неэффективными, с к.п.д. не выше 0,3%! Фактически это были паровые насосы. Настоящая паровая машина непрерывного действия была разработана в Англии знаменитым изобретателем Джеймсом Уаттом во второй половине XVIII в. (Параллельно в России был разработан двухцилиндровый паровой двигатель умельцем-механиком с Урала Иваном Ползуновым, но со смертью автора изобретение было забыто.) В Англии, этой мастерской мира того времени, где две трети населения работали в промышленности, паровые машины распространились необычайно быстро; к началу XIX в., т. е. через 25 лет после изобретения Уатта, их насчитывалось более 1500, они заменяли работу 180 тыс. лошадей. За Англией поспешили континентальная Европа и Северная Америка. В России первая после двигателя И. Ползунова машина заработала на Урале в 1799 г. Паровая машина, по словам Энгельса, оказалась поистине интернациональным изобретением. И это неудивительно, так как она была единственным в то время средством решения проблемы энергетического кризиса. Паровые машины повышенного давления можно было поставить на колеса и получить самодвижущиеся по рельсам повозки; довольно быстро по рекам и внутренним водоемам пошли пароходы, а в 1838 г. Атлантический океан пересекли два парохода, использующие только паровую тягу. Таким образом, к середине XIX в. паровые машины практически везде пришли на смену естественным источникам энергии — воде и ветру. Наступил «золотой век пара», который, казалось бы, мог длиться очень долго. Но... чем больше возможностей, тем быстрее растут потребности. Быстрый количественный рост числа паровых машин, их непрерывные модификации (хорошая аналогия с ЭПЭР и ЭПИР в биологии) уже за хронологических полвека не смогли удовлетворять потребности в энергетических мощностях экспоненциально растущей экономики. Перечислим самые существенные недостатки паровых машин: низкий к.п.д. при увеличении числа и мощности машин приводил к громадному расходу топлива; передача движения от машины к станкам осуществлялась через целые системы трансмиссий, сложные и ненадежные; атмосфера городов с тысячами заводских дымовых труб становилась непригодной для жизни горожан.
В недрах XIX в. зрели новые способы преобразования и использования энергии, но только в XX в. электричество вступило в права основного энергодателя, энергопреобразователя и энергопереносчика. Существует рассказ о том, что когда Майкла Фарадея, открывшего явление электромагнитной индукции, спросили: «А зачем это надо?», он ответил: «Не знаю, но когда-нибудь вы это обложите налогом». Имелось в виду, что это явление будет широко применяться на практике. Но вряд ли и сам великий экспериментатор и все исследователи, изучавшие природу электрических и магнитных явлений, могли предвидеть, как широко войдет электричество в нашу экономику, в быт каждой семьи. Применение электричества резко повысило энергообеспеченность человечества, в том числе и удельную. Электрическая энергия имеет большие преимущества перед другими видами: она быстро и с малыми потерями передается на большие расстояния; может легко преобразовываться в другие виды энергии; к. п. д. электропреобразователей может быть очень высоким, вплоть до 100%. Источником ее может служить как энергия падающей воды, так и энергия органического топлива. Отметим, что около 80% получаемой в мире энергии, большая часть которой превращается в электрическую на огромных ТЭЦ и ГРЭС, производится на основе паровых турбин. Схема превращения энергии органического топлива (угля, нефти, газа, мазута) в электрическую энергию многоступенчата. Например, тепло сгорающего топлива нагревает воду в котле, вода превращается в пар высокого давления, он приводит в движение паровую турбину, турбина — ротор электрического генератора, находящийся в сильном магнитном поле, тоже создаваемом током.
Интересно отметить, что и для пятого этапа развития энергетики, основанного на использовании атомной энергии, основным энергоносителем тоже является пар. Современная атомная и, возможно, будущая термоядерная электростанция — это типичные тепловые станции. В них тапка парового котла заменяется на атомный или термоядерный реактор, а «тепловое тело» — пар — остается. А это значит, что к. п. д. таких станций, как и ранее, не будет высоким. Характерно резкое критическое высказывание по этому поводу профессора А. Китайгородского: «...Сегодняшняя атомная электростанция напоминает мне телегу, которую движет великолепный восьмицилиндровый двигатель» (цит. по [Чирков, 1981, с. 75]). «Дедовские» способы превращения тепла в электроэнергию через посредство пара действительно резко тормозят развитие новых методов производства энергии в наше время. Вот почему в нашем веке остается невысоким вклад атомной энергетики в общую энергетику человечества, не более 5% по прогнозам к 2000 г., хотя ее экологическая безвредность очень привлекательна при безаварийной работе.
Заканчивая обзор развития энергообеспеченности человечества в его эволюции, обратим внимание на то, что к настоящему моменту человек использует и рассеивает энергию, в десятки раз большую, чем среднее млекопитающее его размера. Это означает, что рост энергетических показателей является одним из важнейших факторов в эволюции человека и развитие всех его технологий связано с совершенствованием энергетики. По воспоминаниям соратника К.Маркса В.Либкнехта, познакомившись с работой одного из первых электродвигателей и действием модели железной дороги, К.Маркс сказал, что теперь результаты необозримы: за экономической революцией должна последовать политическая, так как вторая служит только выражением первой (цит. по [Карцев, Хазановский, 1984, с. 135]).
9.2. Будущее человека и перспективы энергетики
Вопрос о будущем развитии энергетики человечества можно по праву считать одним из самых главных в настоящее время. Для этого имеются две основные причины. Во-первых, наше время можно считать переломным из-за близкого истощения сырья, накопленного в былых биосферах. Вскоре, действительно, мы уже не сможем паразитировать на несовершенствах круговоротов прошлых биосфер и будем должны побеспокоиться о более падежных (возобновляемых или имеющих большие запасы) источниках. Во-вторых, следует подумать о гармоническом «вписывании» человечества в глобальный круговорот, пока он не нарушен в его основе (об этом—подробнее в следующей главе). Под этим углом зрения мы и рассмотрим некоторые возможные варианты совершенствования энергетики человечества.

Рис. 14. Рост народонаселения и удельного энергопотребления в XX в. [Скалкин и др., 1981].
1 — народонаселение, млрд чел.; 2 — удельные энергозатраты (% к 1900 г.).
К теперешнему моменту разработано несколько вариантов прогнозов развития энергетики на период до конца века и более отдаленные времена. Все они сходятся в том, что рост энергетики будет наблюдаться в любом случае, несмотря на призывы к его ограничению. И характер этого роста, по крайней мере в ближайшем будущем, остается экспоненциальным. Несмотря на рост народонаселения (рис. 14), и это следует подчеркнуть особо, (удельные расходы (или производство) энергии все же будут возрастать опережающими темпами [Скалкин и др., 1981]. Человек с мощностью основного обмена около 100 Вт рассеивает в 50—100 раз больше энергии в результате развития техники и энергетики, и этот показатель явно возрастает.
Приведем данные прогноза развития энергетики на XXI в. (рис. 15). Если к концу XXI в. атомная и ядерная энергия станут основными в энергопотреблении человечества (на рис. 15 область 4 заштрихована), то конец XX в. будет характеризоваться главным образом еще применением органического топлива, накопленного в прошлых биосферах. Рассмотрим подробнее некоторые аспекты роста и изменения структуры энергетики.
Ископаемым топливом № 1 нашего времени является нефть. Производимые из нее бензин, керосин, дизельное топливо используются в большом числе энергетических машин в качестве основного горючего. Добыча нефти к концу XX в. может достигнуть громадных величин — до десятка миллиардов тонн. И хотя разведанные ее запасы растут, нет сомнения, что более чем на 100 лет ее не хватит. По оценкам экспертов, скорость извлечения нефти из земных глубин п миллион раз превышает возможные максимальные скорости ее накопления в прошлом. Все труднее добывать нефть из недр, все сложнее до нее добираться, да и доля ее экономически выгодного изъятия не превышает 30—60% от имеющейся. Падают и энергетические характеристики. Хотя еще Д. И. Менделеев говорил, что сжигать нефть — это «все равно, что топить ассигнациями».

Рис. 15. Прогноз развития мировой экономики и изменение ее структуры [по Легасову и др., 1985, модифицировано].
Области: 1 — уголь; 2 — нефть; 3 — природный газ; 4 — ядерное и атомное топливо; 5 — остальные источники энергии, включая возобновляемые источники.
Согласно простому, но очень показательному расчету, проведенному Линдсеем Грантом, консультантом Экономического фонда США, нефти при теперешних темпах ее потребления не хватит надолго. Даже если предположить, что вся масса Земли состоит из нефти (?!), то она будет исчерпана за срок около 350 лет, если скорость увеличения ее потребления останется такой же, какой она была до 1973 г. (цит. по [Литературная газета, 1984, 18 апр., № 16 (4978)]).
Примерно то же самое можно сказать об угле, невозобновляемом горючем этого же типа. В начале XX в. он был лидером по использованию в энергетике мира. Запасы его примерно в 10—20 раз больше, чем нефти. Однако он, особенно его наиболее распространенный вариант — бурый уголь, обладает пониженной калорийностью. При сжигании от него остается много отходов и сильнее загрязняется окружающая среда, главным образом ядовитыми оксидами серы и азота, а также зольными выбросами.
Перспективен в настоящее время природный газ, широко используемый в быту и промышленности, но его запасы не превышают запасов нефти, и он тоже заслуживает внимания как химическое сырье, но не как топливо. То же самое относится и к горючим сланцам, торфу и другим ископаемым «подаркам» прошлых биосфер. Сроки их использования ограничены даже в не столь отдаленном будущем.
Принципиально новым видом энергии, перспективным для использования человеком, является ядерная. Основы для ее применения были разработаны в результате достижений одной из самых развитых наук — теоретической и экспериментальной физики. К сожалению, первой серьезной демонстрацией мощи атома были взрывы в военных целях в конце второй мировой войны в японских городах Хиросиме и Нагасаки.
Пионером мирного применения энергии атомного распада стал СССР. 27 июня 1954 г. вступила в строй первая в мире атомная электростанция (АЭС) в г. Обнинске. Ее мощность достигала всего 5 тыс. кВт, но главное достоинство заключалось в практической проверке мирной атомной технологии. В настоящее время мощность одного блока стандартной АЭС составляет 1 млн кВт (т. е. в 200 раз больше первенца), а общее производство энергии в мире с помощью атома достигло в 1980 г. 8 млрд МВт·ч, что приблизилось к 5% от ее общего производства. В 70-х годах более сорока стран пришли к решению о создании у себя атомной энергетики. По некоторым оценкам, к 2000 г. на атомных станциях будет производиться до 30—50% всей электроэнергии. Преимущества применения урана ясны: 1 кг этого горючего дает энергии почти столько же, сколько железнодорожный состав каменного угля.
При использовании энергии атома на одно из первых мест выдвигаются проблемы безопасности. Действительно, опасность атомного взрыва очевидна и, увы, уже продемонстрирована на людях. На АЭС взрывного деления тяжелых ядер урана или плутония не происходит, скорость атомного распада строго регулируется с помощью графитовых поглотителей избыточных нейтронов. Теоретически опасность нерегулируемого взрыва почти нулевая (по расчетам — один случай на миллион лет). Более опасно радиоактивное загрязнение среды при возможных авариях, что и показала трагедия Чернобыля. Далеко не просты проблемы надежного захоронения радиоактивных отходов реакции. Радиоактивные шлаки после извлечения из них ценных урана и плутония для повторного использования необходимо тщательно упрятать поглубже, так как они остаются радиоактивными в течение целого ряда лет.
При должных мерах безопасности средний радиационный фон около АЭС ниже, чем около обычных промышленных предприятий. Правда, цена этому немалая: около 1/3 всех отпускаемых средств должно тратиться на природоохранительные цели, и экономия здесь недопустима.
Упомянем еще одну положительную сторону применения мирного атома. Работа АЭС не связана со сгоранием топлива, а это значит, что не потребляется кислород и не образуется избыточный углекислый газ, как при работе тепловых электростанций. Кроме того, не требуется подачи и нет загрязнения большого количества воздуха (подробнее об этом — в следующей главе).
Имеются ли серьезные ограничения на широкое распространение атомной энергии? Поскольку источник невозобновляем, то надолго ли хватит урана? По самым осторожным расчетам, его хватит на несколько столетий. Особые перспективы в этом смысле сулит разработка реакторов на «быстрых нейтронах». В реакторах этого типа с повышенным удельным тепловыделением балластный уран-238 превращается в плутоний, который тоже является атомным горючим. Параллельно с расходом топлива может идти его накопление в удобной для использования форме и даже в нарастающих количествах. Пока реакторы такого типа сложны и дороги, но это уже обычные задачи совершенствования технологий.
Еще более эффективным, а главное химически и радиационно безопасным источником энергии для человечества может стать термоядерный синтез. Наверное, это — одна из самых сложных технологических задач, стоящих перед человечеством. Академик И. В. Курчатов называл ее «величайшей». С конца 50-х годов по инициативе Советского Союза работы по управляемому термоядерному синтезу стали вестись по международным программам, были сняты все завесы секретности с этой поистине интернациональной задачи. Поступательное движение по пути к управляемой термоядерной реакции становится все более ощутимым. Чтобы «зажечь» реакцию термоядерного синтеза, надо сжать плазму до плотности 10 триллионов ядер дейтерия и трития в 1 см3 и удерживать ее при температуре не ниже 200 млн °С в течение одной секунды. На современных токамаках, тороидальных камерах с аксиальным магнитным полем, удерживающим плазму, удалось достичь в начале 80-х годов либо нужного времени удержания плазмы с температурой около 20 млн °С, либо разогрева плазмы до 100 млн °С при более коротком времени удержания.
Имеется и другой путь к управлению термоядом — это инерциальное удержание плазмы. По этому способу на поверхность таблетки из смеси дейтерия и трития «обрушивается» огромная энергия в виде импульса лазерного излучения. Это приводит к гигантским температурам и давлениям в толще таблетки, при которых может начаться термоядерная реакция. На установках с мощными лазерами получены температуры в 100 млн °С. Для использования термоядерной энергии этим способом потребуется решить ряд дополнительных технологических задач, так как по сути, в данном случае, будет работать серия микровзрывов, в отличие от стационарного состояния плазмы в токамаках. Независимо от способа в начале 90-х годов нашего века можно ожидать разработки демонстрационного термоядерного реактора, т. е. такого, в котором полученная энергия будет не ниже энергии приложенной. И тогда путь к овладению «идеальным» источником энергии будет открыт. В самом деле, этот источник практически неисчерпаем, дейтерия в морской воде хватит на миллионы лет. Химического и радиоактивного загрязнения тоже практически нет, так как работа идет с водородом и гелием. Однако одна из самых сложных проблем, связанных с воздействием человека на биосферу, все же остается. Она относится к применению и атомного, и ядерного горючего. Это — проблема теплового загрязнения, или избыточного тепловыделения.
В конечном счете вся энергия, полученная человеком, превращается в тепло. А это означает, что возможен «перегрев» оболочки Земли со всеми последствиями: опустынивание в теплых регионах, таяние антарктических льдов и подъем уровня Мирового океана и т. д. По оценкам академика Н. Н. Семенова и ряду прогнозов, производство дополнительной энергии можно довести лишь до 3—5% от поступающей на Землю энергии Солнца. Это превышает современный уровень лишь в несколько сот раз. Следовательно, стабилизация должна наступить уже в ближайшие 200 лет, если мы будем использовать ядерное или ископаемое горючее.
Есть и другая перспектива роста энергетики, совершенно безопасная экологически: это более полное использование солнечной энергии, непосредственно поступающей на Землю. В этом случае не будет никакого загрязнения, включая и тепловое, так как работает энергия, уже пришедшая от Солнца и ранее бесполезно терявшаяся в виде сразу выделившегося тепла.
Широкое применение солнечной энергии, или развитие гелиоэнергетики, связано с преодолением ряда трудностей. В основе их лежит низкая концентрированность потока солнечной энергии. Несмотря на огромное общее количество энергии, поступающей от Солнца (более чем в 1000 раз выше энергопотребления человека), на каждый квадратный метр поверхности Земли приходится 100—200 Вт, в зависимости от географических координат. К этому надо добавить нерегулируемую облачность, изменения в течение дня и перерывы на ночное время. Низкая итоговая плотность потока солнечной энергии делает ее неконкурентоспособной с нынешними источниками, по крайней мере в ближайшие 15—20 лет. По современным экономическим оценкам энергия, полученная от солнечных батарей, пока в 100 раз дороже, чем поступающая с теплоэлектростанций. Однако, даже при теперешних способах преобразования солнечной энергии в электрическую, для удовлетворения СССР в энергии нужно «всего лишь» 10 тыс. км2. Много это или мало? Это — квадрат пустыни со стороной 100 км. Это меньше 1% территории, занятой под сельское хозяйство, и меньше площади, запятой в настоящее время угольными шахтами, нефтяными промыслами, нефтепроводами. Есть смысл приложить усилия для овладения солнечной энергией. В настоящее время существует несколько путей ее прямого использования. На физических и химических способах мы не будем останавливаться.
Заслуживают особого внимания направления работ, связанные с получением энергии из органики, накопленной не в прошлых биосферах, а образуемой в нашей биосфере под влиянием непосредственного потока энергии от Солнца. Пример такой возобновляемой органики давно известен — это древесина. Не зря ее доля в общем энергопотреблении резко упала уже к середине нашего века. Низкая калорийность — главная тому причина. А нельзя ли выращивать или получать продукт, близкий по теплотворной способности к самой нефти? Оказалось, что идея далеко не фантастическая, а вполне осуществимая. Есть целый ряд тропических деревьев, обычных кустарников и сорных трав, которые могут выделять в больших количествах соки — легкие углеводороды, близкие по энергетическим свойствам к типичным бензинам. Самые перспективные среди них — растения семейства молочаевых, чертополохи и многие сорта кактусов. Культивирование «нефтеносов» очень перспективно: при культивировании бразильских деревьев, и прежде всего знаменитого каучуконоса — гевеи, с одного гектара можно получить в год до железнодорожной цистерны жидкого топлива. При этом деревья практически не повреждаются, а древесина может использоваться в строительстве и для производства бумаги. Эксперименты показали, что себестоимость «выращенной» нефти приближается к себестоимости традиционной нефти.
Большие надежды возлагаются также на получение водорода с помощью биологических процессов. Водород привлекает внимание энергетиков из-за своей огромной энергоемкости. Не зря его называют топливом помер один для будущего, так как экологическая его чистота очевидна. Если его получать из воды электролизом, то при его сгорании, т. е. соединении с кислородом, снова образуется вода. Получается цикл. Если его осуществлять с помощью химического расщепления воды, то
это энергетически невыгодно. Мы знаем, что растения умеют это делать быстро и хорошо, надо только научиться забирать у них хотя бы часть водорода до того, как он вступит в реакцию синтеза. Наибольшие надежды в этом направлении связываются с хорошо знакомыми нам синезелеными водорослями. Разрабатываются также синтетические аналоги живых фотосинтезирующих систем. Трудно сказать, на каком пути ждет наибольший выигрыш, но уже по современным оценкам с квадратного метра поверхности, освещаемой Солнцем, можно получить за день около 20 г фотоводорода, т. е. около 20 т с квадратного километра. И снова, по ориентировочным расчетам, участок пустыни 140×140 км сможет удовлетворить все энергетические нужды страны. До реального осуществления таких проектов очень далеко, сделаны лишь первые шаги. В самое последнее время удалось увеличить выделение фотоводорода у цианобактерий более чем в 20 раз, используя новые штаммы. Развитие методов генной инженерии позволяет надеяться на быстрое продвижение на этом пути.
Заканчивая анализ развития энергетики человечества, подчеркнем несколько наиболее существенных моментов. Прежде всего это ускорение роста энергообеспеченности человека, непосредственно связанное с развитием общественно-экономических отношений. Если политика — это концентрированная экономика, то экономика — это концентрированная энергетика. Производительность труда как главный показатель уровня экономики базируется на энергообеспеченности. Однако экспоненциальный рост энергетики не может продолжаться, как мы видели, даже в течение ближайших столетий.
Второй показатель, который, в отличие от первого, уже связан с качественными изменениями,—это переход на все более энергетически емкое топливо. Если принять теплотворную способность нефти за единицу, то для угля она составит менее 0,5, а дрова имеют коэффициент только 0,2, по зато водород в 3 раза более калориен, а атомное и термоядерное горючее более энергоемки уже в миллионы раз.

Рис. 16. Тенденции развития энергетики человечества.
1 — энергия, вовлеченная в орбиту деятельности человека; 2 — потери энергии во времени. Внизу заштрихована и отдельно построена область, соответствующая энергии, непосредственно используемой человеком.
Заботы о повышении к.п.д. энергетических машин также можно считать одним из важнейших аспектов развития энергетики человечества на всем ее протяжении. История человечества показывает, что переходы с одного типа источников энергии на другой никогда не происходят плавно, процесс этот полон драматизма и противоречий: периоды быстрого прогресса и количественного роста (гигантизма) сменяются периодами замедления, в которые очень важное значение приобретает экономичность использования источников. Общие тенденции роста энергетики человечества качественно можно представить в виде возрастающей негладкой кривой (рис. 16) с фазами быстрого развития и замедления роста. Видно, что с использованием нового источника общая величина энергии резко возрастала, но при этом возрастали и потери. По мере освоения источника данного типа (и его исчерпания) рост замедлялся, но снижались потери, а значит, росла экономичность. Наиболее гладкая кривая роста соответствует величине использованной энергии, т. е. разнице между всей вовлеченной в деятельность человека энергией и бесполезно потерянной. Эта тенденция имеет характер общебиологический, мы ее отмечали раньше в виде энергетических принципов.
Возможно, что в настоящее время, с крупномасштабных позиций, мы находимся перед очередным скачком — овладением термоядерной энергией. Но и здесь проявятся все те же принципы: наряду с количественным ростом будет наблюдаться и качественный. К снижению теплопотерь нас настоятельно призывает еще более насущная, чем рост энергетики, проблема нашего сосуществования с биосферой, нашего гармоничного вписывания в глобальный круговорот. Этому и посвящена следующая глава.
Глава 10
Человек в циклах биосферы
Культура, если она развивается стихийно, а не направляется сознательно... оставляет поело себя пустыню...
К. Маркс
Есть такое твердое правило: утром встал, умылся, привел себя в порядок — и сразу же приведи в порядок свою планету.
А. Сент-Экзюпери
10.1. Воздействие на биосферу в прошлом
Первое серьезное воздействие человека на окружающую среду непосредственно связано с его «энергетическими достижениями»: с овладением огнем и выработкой простых, но эффективных орудий охоты и лова. Уже более 250 тыс. лет назад первой серьезной жертвой человека стали крупные млекопитающие. Наиболее вероятно, что именно человек истребил мамонтов, гигантских оленей и других крупных травоядных животных во времена верхнего палеолита.
Особенно резко возросло воздействие человека на природу с последующим развитием технологий во времена неолита, с переходом вначале к скотоводству, а затем к земледелию. Выжигание растительного покрова производилось и для увеличения пастбищ, и для повышения плодородия земель. Широко развилось так называемое подсечно-огневое земледелие, сохранившееся в некоторых тропических странах до сих пор! По этому методу вырубали участки леса, сжигали деревья, а почва обогащалась биогенными элементами и давала высокие урожаи даже при плохой обработке. Но все это длилось год-другой, а затем требовалась новая территория. Правда, заброшенная площадь потом зарастала лесом, по это либо происходило за десятки-сотни лет, либо не происходило никогда вследствие иссушения территории. Особенно уязвимыми оказались тропические леса из-за их хорошо сбалансированного круговорота, а следовательно, и из-за слабых почвенных покровов.
К последнему времени накапливается все больше сведений о том, что судьба первых цивилизаций в истории человеческого общества во многом определялась их неразумной экологической политикой. Раньше в каждом учебнике говорилось о благоприятном географическом положении каждой из древних цивилизаций, связанных с обширными лесами, плодородными землями, полноводными реками. И это способствовало развитию и процветанию данной цивилизации. Теперь все чаще говорится, что и конец некоторых цивилизаций был связан с окружающей средой, вернее, с ее резким ухудшением в результате развития самой цивилизации: культура в буквальном смысле оставляла после себя пустыню.
Нельзя сказать, что древние люди не пытались бороться с изменением среды; мы восхищаемся великолепными ирригационными сооружениями Азии и Америки. Но что-то не удавалось, возможно, слишком поздно принимались меры, и потому, например, «козы съели Грецию», а там, где Ганнибал разводил слонов, остались только раскаленные зыбучие пески.
Резкий скачок в развитии техники и особенно энергетики в средние века нашего тысячелетия, переход к использованию машин привели к еще более резкому воздействию на природу. Ощутимые результаты этого давления человека на биологическое окружение появились в Европе, самой густонаселенной и технически передовой. (В гл. 9 мы говорили об Англии, имевшей самые развитые технологии и уже в XVII в. лишившейся почти всех лесов.)
Интересны меры по охране природных ресурсов, предпринятые в России Петром I. Жестокость этих мер определялась не только русским деспотизмом, но и их насущностью. От сельскохозяйственных палов и вырубки лесов страдала оборонная мощь государства. Изданный в 1703 г. указ об охране лесов под угрозой физической расправы запрещал рубки на расстоянии до 50 верст от берегов крупных рек и до 20 верст от малых. Сбросы нечистот в Неву создавали опасность для северной столицы, и Петр издал указ об их запрещении. Виновные в нарушениях солдаты присуждались к порке и ссылке, а офицеры — к разжалованию в рядовые. Было даже запрещено обрабатывать на льду бревна и доски, чтобы не загрязнять воду. Меры давали эффект: так в Москве были очищены старые пруды, служившие местом свалок, и после того они превратились из Поганых в Чистые, что соответствовало истине не только на словах. К сожалению, преемники Петра но были столь дальновидными по отношению к природе: один из результатов—превращение части Черноземья в Нечерноземье из-за эрозии почв в средней полосе России.
С 1600 г. на Земле вымерло 74 вида (1,23%) птиц и 63 вида (1,43%) млекопитающих. Еще больше погибло подвидов птиц и зверей, из них не менее 80% погублено человеком. Пока еще ни один другой вид в биосфере не был столь смертоносным для других: ведь многие виды не были на стадии вымирания, наоборот, целый ряд из них находился на стадии биологического прогресса, т. е. был крайне многочисленным, а необходимости в истреблении этого вида не было.
Особенно трагичной с этой точки зрения выглядит история с истреблением странствующего голубя в Северной Америке, ярко демонстрирующая всю неграмотность и жестокость человека как хищника. В 1810 г. американскими орнитологами наблюдалась огромная стая странствующих голубей в долине р. Огайо (описание дано по [Риклефс, 1979]). В течение нескольких дней колонна птиц шириной значительно больше километра пролетала над головой, закрывая небо. По подсчетам орнитологов, число птиц в этой стае превышало 2 млрд особей. А примерно через 100 лет, в 1914 г., умер последний странствующий голубь в мире — это была старая голубка, долго жившая в Цинциннатском зоопарке. Последний голубь на свободе был убит в 1899 г. И единственной причиной его истребления стал человек. Его стреляли, ловили сетями, сшибали на землю шестами, рубили деревья с гнездами, выпускали свиней, поедавших птенцов. На всех рынках мира за бесценок продавалось огромное количество голубей. Но уже через 10 лет странствующие голуби стали редкостью и вскоре были полностью истреблены. В одном из парков Висконсина осталась бронзовая мемориальная доска в память последней убитой птицы с печальным добавлением: «Этот вид вымер из-за алчности и легкомыслия человека». И после таких деяний мы называем себя Homo sapiens!
Почти столь же трагична история с американским бизоном. Сходна история и его близкого родственника — европейского зубра. Он был широко распространен почти по всей Европе, Кавказу, Закавказью. Но в 1920 г. даже в Беловежской пуще, где он охранялся, а в 1927 г. и на Кавказе в естественном состоянии он перестал существовать. Международное общество по спасению зубров провело инвентаризацию животных, содержавшихся в зоопарках, и получило печальный итог — 56 особей. Интенсивные меры по восстановлению популяции зубра привели к тому, что численность чистокровных зубров в мире достигла 2000. Это уже неплохо. Но трагедийной ситуации здесь могло и не быть.
Отметим очень важное обстоятельство: к настоящему времени заработал глобальный «бумеранг-эффект» в наших отношениях с природой. За свое неправильное отношение к ней, за наше незнание законов природы мы начинаем получать ответы во все возрастающих масштабах. Как правило, они негативны, а мы к этому не готовы.
10.2. Современная проблема: человек и биосфера
Рост воздействия человека на биосферу непосредственно связан с ростом его численности. Разговоры о демографическом взрыве не просто красивые фразы. Увеличение числа людей на планете за последнее столетие носит именно взрывообразный характер (рис. 17). Еще более быстрыми темпами растет загрязнение окружающей среды, связанное с деятельностью человека. Есть данные, что около 80% всех видов загрязнения биосферы обусловлено энергетическими процессами, включая добычу, переработку и использование топлива. Резко возрастает с развитием разнообразных химических технологий количество трудноокисляемого органического мусора, производимого человеком. Уже к началу 70-х годов оно превысило астрономическую цифру — 2 · 1010 т/год [Ковда, 1975]. Этот показатель удваивается раз в 6—8 лет и к середине 80-х годов уже должен оцениваться величиной около 1011 т/год. Если принять среднее время распада этого трудно разложимого в природных условиях мусора за 10 лет, что явно превышает возможности биосферы, не «умеющей» разлагать незнакомые ей соединения типа пластмасс, то количество имеющегося мусора антропогенного происхождения составит 1012 т. А эта цифра уже приближается к общей массе живых организмов биосферы, уступая ей лишь в 2 раза, но зато в 5 раз превышает производство этой биомассы в год. Это первые данные, по которым активность человечества сравнялась с активностью биосферы. (В то время как биомасса человечества не превышает 0,01% от биомассы биосферы, а поток энергии, им используемый, достигает десятых долей процента.) Жаль, что глобальная активность человечества является лишь мусоропроизводящей, но таково состояние дел.

Рис. 17. Демографический взрыв [Агесс, 1982].
1 — Северная Америка; 2 — Латинская Америка; 3 — Австралия и Океания; 4 — Африка; 5 — СССР; 6 — Индия; 7 — Китай; 8 — остальная часть Азии.
Если нарисовать рост производства грязи человеком во времени, по аналогии с рис. 17, то «гриб» взрыва будет гораздо шире, да еще и с тлетворным душком.
Если говорить о воздействии человека на первичную продуктивность биосферы, то по ряду оценок в результате отчуждения земель для непроизводительных нужд (производство, города, дороги и замены лесных экосистем на сельскохозяйственные монокультуры, менее продуктивные по первичной продукции) уменьшение составляет 15—20% [Титлянова, 1979].
Рассмотрим подробнее влияние человека на основные резервуары и аккумуляторы веществ в биосфере: атмосферу, гидросферу и литосферу (точнее, ее верхнюю часть на суше — почву).
Состояние атмосферы. Что же происходит в атмосфере, а проще говоря, с тем самым воздухом, без которого нам не обойтись и пары минут? К сожалению, точный анализ этой сложной задачи невозможен. Поскольку наш подход связан с изучением потоков энергии, то большее внимание мы будем уделять влиянию энергетики на состояние и динамику процессов в атмосфере.
Из-за высокого уровня развития промышленности в Северном полушарии около 93% всех газовых выбросов в атмосферу сосредоточено именно здесь. Более того, около 90% этих выбросов приходится на 8— 10% поверхности суши, включающей часть Европы, Северной Америки и Японии. Следовательно, основная часть продуктов сгорания всех видов топлив выбрасывается в атмосферу на площади лишь около 3% от всей поверхности планеты. И на этой территории темными шлейфами дымов выделяются крупные города.
Одними из наиболее токсичных соединений, поступающих в атмосферу из технических источников, являются оксиды серы и азота. Согласно различным оценкам, техногенный выброс в атмосферу оксидов серы к началу 80-х годов достигал 70—100 млн т, а оксидов азота — около 20 млн т. Эти показатели примерно равны величине естественных выбросов этих элементов в атмосферу [Курьер ЮНЕСКО, 1985, № 5, с. 21]. Наиболее токсичным из газообразных оксидов является сернистый ангидрид. (По некоторым оценкам, если бы его не перерабатывали высшие растения, то за 20 лет все высшие животные погибли бы.) К сожалению, он составляет до 99% от выбросов сернистых соединений энергоустановками. Время жизни его в атмосфере может достигать нескольких десятков суток при сухом воздухе или уменьшаться до нескольких часов в присутствии воды. Он участвует в различных каталитических, фотохимических реакциях, переходит в сульфаты и, растворяясь в парах воды, образует серную кислоту. Сходным образом оксиды азота дают азотную кислоту. Получающиеся из-за этого кислотные дожди стали бичом Северного полушария.
Еще большую опасность представляет техногенный выброс в атмосферу металлов, особенно тяжелых. В естественных условиях поступление металлов в атмосферу — это ветровые переносы с поверхности и выбросы вулканов. Но все эти потоки отступают перед техногенными. Антропогенные источники приносят в атмосферу почти в 20 раз больше свинца, почти в 10 раз больше кадмия, более чем в 7 раз — цинка. По величине выбросов цинка, меди, кадмия всем вулканам Земли далеко до мусоросжигательных печей. Особую тревогу вызывает свинцовое загрязнение планеты. Связано оно также с энергетикой, но с малой, точнее, с выхлопами автомобильных двигателей.
Концентрация свинца растет во льдах не только Гренландии, но и Антарктики, в иле рек, морей, в теле животных, не исключая человека. Если в скелете первобытного человека было лишь 2 мг свинца, то в наших скелетах его в 50—100 раз больше. А ведь избыток свинца в организме очень опасен для здоровья.
Примеры накопления ядовитых веществ в атмосфере и из нее попадания в другие сферы можно, к сожалению, множить почти беспредельно. Достаточно вспомнить об авиационных методах применения различных химикатов в сельском и лесном хозяйстве. Неудивительно, что ДДТ был обнаружен в заметных количествах в снегах Антарктиды, где он никогда не применялся. Известно, что многие пестициды способны концентрироваться в пищевых цепях. Так, в цепях питания от морской воды через планктон, рачков и рыб смытые в воду химикаты (тот же ДДТ) в яйцах морских птиц могут аккумулироваться в миллион (!) раз.
Приведенных примеров вполне достаточно, чтобы видеть, что загрязнение атмосферы приняло угрожающие масштабы, и угрожают они прежде всего самому человеку. Чтобы избежать уже упоминавшегося бумеранг-эффекта, необходимо принимать серьезные меры по очистке газовых выбросов. По оценочным расчетам, снижение выбросов серы вдвое на обычных тепловых электростанциях в Европе стоило бы не менее 10% издержек на производство самой электроэнергии. Может показаться дорого, но что может быть дороже здоровья человека и биосферы?
Хорошим обнадеживающим примером того, что человечество в состоянии справиться с глобальным загрязнением, служит снижение радиоактивности на поверхности Земли. В начале 60-х годов, к моменту запрещения испытаний ядерного оружия в атмосфере, космосе и под водой, содержание радиоактивных стронция-90 и цезия-137 в продуктах питания возросло по сравнению с естественным фоном в несколько, вплоть до десятков, раз. Концентрация этих элементов в мышечной и костной тканях человека и высших животных тоже возросла и в некоторых регионах даже более чем в 100 раз. После прекращения испытаний, примерно в течение десятилетия, радиоактивный фон и содержание радиоактивных соединений в живых организмах вернулись к обычному естественному уровню, кроме отдельных мест, связанных с повышенной техногенной радиоактивностью.
Загрязнение гидросферы. Вода составляет основу жизни, и мы неоднократно обсуждали на страницах этой книги, что и в циклах вещества, и в энергетике всех живых существ она занимает ведущее место. Она пронизывает нашу жизнь на всех ее уровнях. И вдруг неожиданно для человечества оказалось, что мы практически не имеем чистой воды на планете. Недаром 80-е годы объявлены ООН десятилетием борьбы за чистую воду. Громадный Мировой океан на пути превращения в бессточную мусорную яму. Особенно загрязнен Атлантический океан.
С ростом энергетики, промышленных и сельскохозяйственных технологий резко возрастает потребность в чистой воде. Если ежегодно мировое потребление энергетических ресурсов составляет около 10 млрд т, то человек расходует около 10 млрд т воды ежедневно. И это не удивительно. Каждая тысяча киловатт мощности, выработанной тепловыми электростанциями, требует миллионы кубических метров воды в год. Для производства 1 т стали нужно более 100 т воды, 1 т произведенной соды «потребляет» 300 т воды, искусственного шелка — 400, бумаги — более тысячи, резины — 4000.
Одним из основных потребителей воды является сельское хозяйство. Например, чтобы вырастить 1 т пшеницы, требуется за вегетационный период 1500 т воды, 1 т риса — более 7 тыс. т, 1 т хлопка — около 10 тыс. т воды. С ростом технологий к 2000 г. без их качественного изменения может понадобиться практически весь речной сток. И здесь сельское хозяйство является одним из главных поставщиков различных ядовитых соединений типа пестицидов: гербицидов, инсектицидов, фунгицидов и пр.
Ухудшение качества воды уже теперь резко отрицательно сказывается на здоровье человека. По оценкам ВОЗ, 80% всех болезней в мире связано с неудовлетворительным качеством воды и нарушением санитарно-гигиенических норм из-за ее нехватки.
То самое «самоочищение» биосферы, которое выручало человека с незапамятных времен, начинает давать опасные сбои и именно в глобальном масштабе. Как бы ни казался велик наш «мокрый шарик», он не справляется с потоком загрязнения.
В последние годы как будто удалось остановить рост одного из наиболее опасных загрязнений Мирового океана — загрязнения нефтью. Нефть способна разливаться тонкой пленкой по поверхности воды, изолируя атмосферу от океана, образовывать комочки различных размеров, а наиболее тяжелые фракции могут опускаться на дно. Килограмм нефти может разлиться тонкой пленкой на площади до 1 га и тем самым погубить свыше 100 млн личинок рыб и многие виды зоопланктона. Тяжелое впечатление производят аварии супертанкеров при перевозках нефти. Но главный поток загрязнения связан со сбросами балластных и промывных вод танкеров. Этот поток был значительно уменьшен в связи с принятием международных постановлений и организацией контроля за их исполнением. В ряде больших портов построены специальные очистные станции для промывки танкеров, которые даже приносят прибыль. Однако угроза увеличения нефтяного загрязнения остается, так как в последние годы быстро увеличивается подводная добыча нефти и газа на шельфе.
Особую тревогу вызывает загрязнение внутренних водоемов, даже если они связаны непосредственно с океаном. Так, самым загрязненным из морей является Средиземное. Турецкие ученые пришли к выводу, что «Мраморное море гибнет». Вызывает обоснованную тревогу уровень загрязнения Балтийского и Северного морей. Без принятия эффективных мер результат может быть печальным. Медлить нельзя.
Определенный оптимизм внушают положительные примеры как в Европе, так и в Северной Америке, самых неблагополучных с этой точки зрения регионах. Достаточно вспомнить о том, что несколько чище стали Волга, Днепр, Великие озера, Темза. Разработка и использование специальных очистных систем, как показывает опыт, не только требуют затрат, но и могут самоокупаться и приносить прибыль.
Состояние литосферы (почвы). Верхний плодоносящий слой суши — почва — по площади, а по объему тем более существенно уступает гидросфере. Толщина слоя достигает всего лишь нескольких десятков сантиметров и только в черноземной зоне может возрастать до нескольких метров. Но именно этим слоем, его состоянием определяется жизнь человечества с самых ранних этапов развития.
В настоящее время практически все пригодные для земледелия места заняты человеком и переезжать просто некуда.
Действительно, площадь засеянных угодий составляет около 10% наземной части биосферы. А для сбора урожая и выпаса используется уже около 30% суши. Если учесть, что для хозяйственных нужд пригодно менее половины поверхности суши, а для интенсивного земледелия только около 25%, то и по этим оценкам человек практически занял всю пригодную территорию. Эра экстенсивного земледелия кончилась, оно постепенно переходит на рельсы интенсивного развития. Но этот путь требует другого подхода и другого типа мышления. Простая переэксплуатация земель быстро приводит к истощению почвы, ее эрозии, вплоть до полного уноса с пылевыми бурями. Хорошо известны пыльные бури на Великой американской равнине, в Средней Азии, на Северном Кавказе. В засушливых песчаных районах разрушение поверхностного слоя поразительно быстро приводит к появлению зыбучих песков и полному опустыниванию территорий. Идет «наступление» песков Сахары, растут пустыни Юго-Восточной Азии, Северной и Южной Америки. Ежегодно потери продуктивных земель по земному шару только вследствие опустынивания составляют 50—70 тыс. км2, а общая площадь «искусственных» пустынь, возникших в результате деятельности человека,— более 9 млн км2. (Эта величина уже сравнима с площадью нынешних посевных угодий.)
Может быть, еще большую опасность для решения продовольственных задач населения Земли представляет разрушение самых плодородных почв — черноземов в результате их переэксплуатации. Поскольку не менее половины всех черноземных земель расположено в Советском Союзе, то для нашей страны эта проблема приобретает самое злободневное звучание (из 300 млн га черноземных почв в Советском Союзе находится более 150 млн га). Если сопоставить со всей пахотной площадью (225—227 млн га), то черноземы составят около 70% ее. По разным оценкам, около 80% всей продовольственной продукции страны производится и выращивается на черноземах.
Учитывая большие запасы гумуса в черноземах, можно подсчитать, что количество энергии, запасенной в черноземах, в 20 раз и более выше, чем в суммарной биомассе высших и низших растений и животных этих ландшафтов. Впечатляющие данные. Можно провести некоторые аналогии с запасом в океане тепловой энергии, который тоже в 20 раз больше, чем ежегодный приход от Солнца. Следовательно, можно говорить о том, что, используя черноземы, и по энергетике, и по веществу мы живем за счет прошлых биосфер, как и при использовании органических ископаемых типа нефти и угля. Правда, в данном случае это не столь отдаленные времена — на образование чернозема требуется «всего лишь» сто-двести лет. Недаром великий русский почвовед В. В. Докучаев называл русский чернозем «главным основным богатством России, стоящим неизмеримо выше богатств Урала, Кавказа, богатств Сибири».
В настоящее время много энергии и вещества уносится с урожаями из черноземов. Например, чтобы компенсировать унос 30 ц зерна с 1 га или 100 ц картофеля, или 200 ц сахарной свеклы, нужно вносить эквивалентное количество органики, которое должно исчисляться тоже десятками центнеров на 1 га. При посевах монокультур необходимо именно внесение органики извне, но откуда ее столько взять? Для бездефицитного баланса гумуса по стране в целом требуется 1,5—3 млрд т органики в год, всего удобнее ее применять в форме твердого подстилочного навоза, а его «производство» в животноводстве достигает лишь 300—400 млн т в год, т. е. в 5—10 раз и меньше. Явное несоответствие потоков налицо. Поэтому необходимо иметь ориентацию на естественное возобновление плодородия почв, запасов гумуса «биологическим путем», а именно за счет сбалансированных севооборотов. Применение в них бобовых растений позволяет накапливать в почве столь дефицитный органический азот, другие компоненты питательных веществ пополняются корневой массой, которая остается в поле. Честь разработки севооборотов с использованием азотфиксирующих бобовых растений принадлежит нашей стране, достаточно вспомнить В.В.Докучаева, Б.Б.Полынова, В.Р.Вильямса. Однако практическое использование таких севооборотов явно недостаточно [Ковда, 1985]. Монокультуры имеют явное предпочтение. Однако не следует забывать грозного, но верного предостережения основоположника научного земледелия Ю. Либиха: «Нет более прямого пути к абсолютному обнищению народа, чем беспрерывная культура однолетних растений» (цит. по [Усольцев, 1985, с. 152]).
По-видимому, не меньше вещества, а с ним и энергии уносится из-за смывов черноземных почв в результате применения неправильных агротехнических приемов. Весенние смывы и изобильные поливы приводят к уносу с дождевой и поливной водой до 1,5— 1,6 млрд т почв ежегодно.
Избыточный полив или даже просто интенсивное орошение земель приводит к еще одному страшному «бичу» орошаемого земледелия — к засолению почв. (Проблема эта древняя, существовавшая еще у вавилонян.) Даже если вода, используемая для орошения, не очень соленая и относится к слабо минерализованной, все равно при большом ее расходе с орошением полей в почве накапливаются соли, и она быстро приходит в негодность, теряя плодородие. Образуемые человеком «вторичные» солончаки, к сожалению, широко распространены по всему миру.
С энергетической точки зрения интенсификация сельского хозяйства прежде всего означает привлечение дополнительных потоков энергии, повышение расхода энергии, затраченной на производство единицы продукции. Это дополнительные потоки энергии прежде всего от добываемого энергетического сырья. Так что при полной оценке по энергии производимой сельскохозяйственной продукции общие энергетические затраты возрастают многократно. «При интенсивном ведении сельского хозяйства большая часть энергии для производства картофеля, мяса и хлеба берется не от Солнца, а из ископаемого топлива. Широкая публика плохо себе это представляет. Например, многие думают, что большие успехи в сельском хозяйстве объясняются только умением человека создавать новые генетические варианты. Но использование этих вариантов рассчитано на большой расход дополнительной энергии. Деятели, пытающиеся помочь развивающимся странам поднять эффективность их сельского хозяйства, не обеспечив значительных дополнительных вложений энергии, просто не понимают положения дел. Основанные на опыте высокоразвитых стран рекомендации для развивающихся стран могут иметь успех только в том случае, если они сопровождаются подключением к богатым источникам дополнительной энергии...» [Odum H., 1967; цит. по Одум Ю., 1975, с. 64]. Эта довольно длинная цитата из работы известного американского эколога Г. Одума убедительно иллюстрирует необходимость правильного учета энергетических потоков при интенсификации процессов биосинтеза.
Одним из наиболее негативных явлений интенсификации сельского хозяйства является резкое ухудшение окружающей среды, и прежде всего ее загрязнение избыточными удобрениями и ядовитыми пестицидами. Чем больше вносится удобрений, тем меньшая доля их используется непосредственно растениями, тем большая часть уходит со смывами в водоемы, загрязняя реки, озера и даже моря и океаны.
Загрязнение внутренних водоемов материков биогенными соединениями типа нитратов и фосфатов, смываемыми с полей и культивируемых пастбищ, является одним из главных загрязнений гидросферы планеты, и об этом мы говорили в предыдущем разделе. Но еще большую опасность представляет загрязнение почвы и воды пестицидами, одним из главных источников которых также является сельское хозяйство. Обработка почвы за последние 100 лет становилась все более активной. Это означает, что мобилизовывалось естественное плодородие почвы и повышалась урожайность, по в то же время снижалось содержание почвенного гумуса и усиливались эрозионные процессы. Для восполнения потерь стали применяться искусственные удобрения, с помощью мощной сельскохозяйственной техники росли производительность труда, урожайность и эксплуатация земель. Но в целом индустриализация сельского хозяйства с очевидностью приводит к серьезным негативным последствиям. Возрастает энергетическая цена, которую надо платить, чтобы ликвидировать эти последствия: рекультивировать и восстанавливать эрозированные почвы, очищать водоемы, усложнять водозаборные и водораспределительные сооружения, совершенствовать системы очистки и т. д. Это и есть одна из важных причин снижения биоэнергетических показателей.
Для разрешения этих противоречий анализируются различные пути. Один из них связывается с созданием систем минимальной обработки почвы, чтобы не нарушать ее структуру и минимизировать кажущиеся энергетические траты (до 50% общих трат энергии при отвальной обработке почвы идет на вспашку и посев). Если, например, вносить гербициды в почти не обработанную почву (вариант нулевой обработки почвы), то затраты энергии уменьшаются в 3,5 раза. Но с одной стороны, количество гербицидов, необходимых для минимальной и нулевой обработки, столь велико, что увеличивает энергетическую цену продукции почти до тех же величин, что и при традиционных методах плужной обработки. А с другой — большое количество применяемых гербицидов представляет такую опасность для потребителей продукции сельского хозяйства, перед которой бледнеют все прочие. Гербициды, как и всякие другие «...циды», это убийцы живого. Их избирательность далеко не абсолютна, как у всех химических соединений; большинство из них имеют длительный срок жизни, канцерогенны, мутагенны и т. д. Достаточно напомнить о том же ДДТ, распространившемся по всей планете и накопленном в теле антарктических пингвинов. Сельское хозяйство имеет гораздо большие масштабы и распространенность на планете, чем другие виды деятельности человека, связанные с использованием биоцидов, поэтому приходится считаться с тем, что его «цидовая» политика представляет серьезную опасность в настоящее время. И прежде всего это относится к здоровью самого человека.
Итак, резюмируя современное состояние всех сфер (атмо-, гидро- и лито-) биосферы, мы можем еще раз подчеркнуть, что оно вызывает серьезную тревогу, так как загрязнение приняло глобальный характер. Работа по старым технологиям становится невозможной. Основные антропогенные загрязнители: энергетика, промышленность, индустриализованное сельское хозяйство — должны быть в корне перестроены с учетом сохранения окружающей среды.
Известный французский эколог Франсуа Рамад заключает свой обстоятельный труд «Основы прикладной экологии» [Л., 1981] весьма примечательными словами: «Сейчас, в конце 20-го столетия, никто не станет отрицать, что только радикальное изменение взаимоотношений между человеком и природой позволит нам избежать судьбы динозавров» (с. 514).
10.3. Перспективы сосуществования
Идея о том, что биосфера, в которой развивается разумная деятельность человека, превращается в ноосферу (сферу разума), наиболее глубоко развита в трудах В. И. Вернадского. Одним из условий ноосферы как единого организованного целого является гармоничная связь всех частей на разных уровнях и их согласованное взаимодействие. Что касается современной биосферы, то она пока только частично охвачена преднамеренными и целенаправленными воздействиями носителя разума — человечества. Поэтому в настоящее время можно говорить лишь о начальных этапах, о самом возникновении или рождении ноосферы, а до полного становления ее еще далеко [Будыко, 1984]. И увы, как все новое, рождается ноосфера в муках, и, по-видимому, мы сейчас переживаем один из наиболее тяжелых периодов ее рождения (см. материал предыдущего раздела).
Вселяет определенный оптимизм разработка международными организациями Всемирной стратегии охраны природы (в Советском Союзе она оглашена 5–6 марта 1980 г.). Она предназначена для правительственных, неправительственных, общественных и международных организаций в качестве руководства к мероприятиям по охране природы. На ее основе рекомендуется разработка национальных стратегий охраны природы в каждой стране. Особое место должно отводиться охране типичных для данного региона экосистем, центров эндемизма и эндемичных видов, существование которых находится под угрозой.
Так как главной причиной ухудшения окружающей среды и загрязнения биосферы являются технологии, то мы и рассмотрим перспективы их изменения в будущем по трем основным типам: энергетика, промышленность, сельское хозяйство. Сделаем попытку оценить, какие направления развития технологий являются наиболее экологичными, способными гармонично вписываться в окружающую среду.
Как всегда начнем с энергетики. В предыдущей главе мы достаточно подробно описали общие перспективы развития энергетики на ближайшие 20–100 лет. Предпочтение отдавалось «большой» энергетике термоядерного синтеза как самой экологичной, не дающей ни радиоактивного, ни химического загрязнения и не выделяющей CO2, приводящего к дополнительному тепловому загрязнению. Дело за «немногим»: нужно научиться управлять термоядерной редакцией. Однако, по имеющимся оценкам, стоимость решения этой задачи в десятки раз ниже расходов на вооружение; не будем забывать, что мы входим в ноосферу, т. е. сферу разума. «Малая» энергетика должна активно использовать энергию солнечного излучения, абсолютно экологичную.
Промышленное производство, включая разнообразнейшие химические синтезы, является одним из самых сильных загрязнителей. Эффективность современного производства, с точки зрения использования сырья остается крайне низкой: в готовом продукте содержится всего 2—10% от сырья по весу. Таким образом, до 98% от исходного сырья промышленность выбрасывает в окружающую среду, отсюда и понятно: чем интенсивнее хозяйство, тем больше рассеивается вредных веществ и тем выше их концентрация в среде обитания. Так, в начале 70-х годов развитые страны с населением, составляющим около половины человечества, имели долю в мировом загрязнении свыше 85%, т. е. почти в 7 раз более активно (на человеческую душу) загрязняли окружающую среду, чем развивающиеся страны. Семидесятые годы можно назвать годами упорядочивания отношений промышленности с биосферой. Если раньше отходы, не особенно беспокоясь, выбрасывали «за ворота» предприятия, то теперь часть их, особенно наиболее токсичная, перерабатывается, остальное тщательно рассеивается в окружающей среде. (Мы знаем из предыдущего, что грязь растет по всей биосфере.) Мечты о безотходных технологиях становятся все более настоятельными, идет лихорадочный поиск повышения степени замыкания производств. На этом пути имеется ряд метаморфоз, которые образно названы «мифом о безотходной технологии», или «псевдобезотходностью».
Рассмотрим один из примеров. Шлаки и шламы цветной металлургии, а их накапливается несколько сот миллионов тонн ежегодно, могут повышать качество стройматериалов. Казалось бы, выход найден: отходы одного производства стали ценным сырьем для другого. Но увы, опасность вредного биологического действия отходов цветной металлургии отнюдь не уменьшается от того, что они входят в состав строительных блоков. Просто опасность появляется в другом месте. Канцерогенность и аллергенность кадмия, никеля и других тяжелых металлов остается, и стройматериалы, особенно в жилищном строительстве, с повышенным содержанием тяжелых элементов просто недопустимы.
Можно привести примеры «революционных» идей этого типа и в других областях, в частности попытки использовать почвы под видом орошения как место утилизации неочищенных промышленных и бытовых стоков. Емкость почвы гораздо выше емкости воды, но и ее очистка и восстановление тоже гораздо сложнее. Недаром академик ВАСХНИЛ В.В.Егоров [1985] назвал такие предложения «дичайшими». Такие примеры, к сожалению, можно приводить еще и еще. Главное — не перебрасывать отходы с места на место, а организовать их глубокую переработку до соединений, безвредных для человека и биосферы. Может быть, самый главный вред от «псевдобезотходности» в том, что она тормозит сам технологический прогресс. В самом деле, отрасль, которая наработала горы отходов и сумела их «сбагрить в чужие руки», выступает уже не в качестве отравителя природы, а в достойной и благородной роли производителя и поставщика ценного сырья и даже получает за это экономические блага. Потребители отходов заинтересованы в бесперебойной поставке этого сырья, без изменения его свойств. «Так создается порочный круг, в основе которого лежит неверно понятый принцип безотходности... Безотходная технология необходима человеку. Но к ней ведет крутая лестница научных, технических и промышленных решений, которую нам ступень за ступенью еще предстоит одолеть»,— пишут Н.Ф.Реймерс и И.А.Роздин [1981, с. 15]. Очень верные слова! Мы не должны обольщаться безотходностью, она пока недостижима, и точнее говорить сейчас о «малоотходных» технологиях или об «экологически безвредных».
Ближайшей непосредственной задачей, стоящей перед человечеством, является интенсификация имеющихся и разработка новых путей и методов борьбы с загрязнением среды, включая активное очищение.
Наиболее существенным способом борьбы с загрязнением промышленного и индустриализованного сельского хозяйства является разработка специально создаваемых очистных сооружений, так называемых систем интенсивной очистки. Основную нагрузку в этих системах, особенно при очистке водных стоков, несут ассоциации микроорганизмов, способные утилизировать широкий спектр активных загрязнителей различного рода. При этом степень очистки особо ядовитых специализированных промышленных стоков с небольшим количеством ингибиторов сильного действия заметно возрастает, если используются специально отселекционированные штаммы микроорганизмов, способные инактивировать сильно ядовитые соединения. Однако, несмотря на высокие скорости метаболических процессов и широкие возможности регулирования обмена у микроорганизмов, практически невозможно организовать абсолютно полную очистку загрязнений в выходящих потоках жидкости, газов даже с учетом повышения степени замкнутости технологий. С приближением к полному очищению стоимость процесса очистки возрастает экспоненциально. Дополнительную доочистку, таким образом, приходится перекладывать на естественные, т. е. экстенсивные, процессы самоочищения в биосфере. Как уже неоднократно отмечалось, биосфера не справляется с ростом загрязнения и по ряду параметров происходит довольно быстрая его аккумуляция.
Эмпирически к настоящему времени нащупывается выход из очень сложной ситуации с растущим загрязнением среды. Он заключается в разработке промежуточных систем, играющих роль буфера между выходом интенсивной системы очистки и «входом» биосферы. Обычно это выделенный участок ранее существовавшей экосистемы, довольно большой по размерам, с ярко выраженной модификацией, произведенной человеком (например, в нашей стране мелиорированные лиманы на Черном море, рукава Волги, Дона, старицы Оби, Енисея с примыкающей территорией и т. д.). Увеличенный размер и уменьшенные скорости деструкции загрязнителей отличают такую сложную систему от интенсивных специализированных систем. Кроме того, такие буферные системы характеризуются разветвлением потоков, наличием циклов по ряду веществ и организацией биотического круговорота в гидро- и педосфере. Это сближает их с природными экосистемами, однако заданные функции самоочищения в них более специализированы и более интенсифицированы, чем в природе. Такие системы можно отнести к управляемым экологическим системам (УЭС), функционирование которых направлено на выполнение определенных функций, задаваемых человеком.
Сельское хозяйство, являясь одним из наиболее опасных загрязнителей окружающей среды, видимо, еще долго будет оставаться в этой неприглядной роли. Помимо эрозии почв почти по всей планете сельскохозяйственное производство широким потоком «распыляет» в биосфере специальные ядовитые соединения типа гербицидов и пестицидов. Увы, пока без таких соединений не обойтись, а химизация сельского хозяйства приносит большую выгоду. Выгода несомненна и сиюминутна, вред от применения пестицидов не столь очевиден, но, к сожалению, он долговременен. Мы знаем, как пестициды накапливаются в цепях питания, концентрируясь в организме человека в сотни тысяч и миллионы раз. Есть ли выход из создавшегося положения? Конечно, он связан с использованием биологических методов борьбы с вредителями. Эти методы имеют неоспоримые преимущества перед химическими. Вот главные: высокая избирательность действия, а следовательно, и безвредность для человека; возможность длительного существования действующего агента (например, растущей популяции организмов, паразитирующей на вредителе); меньшая вероятность появления устойчивых форм вредителя к патогенным организмам, чем к химикатам. Это последнее свойство особенно интересно в эволюционном плане, так как паразитирующая на вредителе и тем самым работающая на человека популяция быстро адаптируется к новым устойчивым вариантам хозяина. Это позволяет надолго сохранять вирулентность патогена [Печуркин, 1978].
Биологические методы пока намного дороже, чем химические, и не столь эффективны, производство биологических препаратов не налажено в больших масштабах, но и эти затруднения не принципиальны. Как мы говорили, эти сложности имеют технологическую природу, а значит, преодолимы. В наше время наиболее перспективно использование комбинированных способов борьбы: химия + биология с постепенным и неотвратимым наращиванием вклада биологии.
Очень сходна ситуация с использованием гербицидов: и здесь возникают сложности с химическим загрязнением среды. Хотя применение гербицидов для безотвальной обработки почвы позволяет избегать эрозии (но энергетически недешево!), их накопление в окружающей среде грозит большими неприятностями. Эффект аккумуляции более опасен, так как гербицидов для полного уничтожения целых армий сорняков требуется во много раз больше, чем пестицидов против вредителей. И здесь выход — в переходе к биологической системе земледелия (это мы обсуждали в предыдущем разделе). Потребуются более высокая культура земледелия, строгое выполнение правил агротехники и другие очевидные вещи.
Большой интерес для будущего представляет нетрадиционная форма ведения сельского хозяйства. Современное сельское хозяйство потому и неэффективно и громоздко, что оно рассеяно по поверхности планеты, «размазано» по большим площадям. К настоящему времени разработано несколько схем гигантских биофабрик (биотронов) с почти замкнутыми экологическими системами и с практически безотходным производством.
Другой вариант развития нетрадиционных вариантов сельского хозяйства связан с заменой дефицитного животного белка на белок одноклеточных или соевых растений. Производство дрожжевого белка вышло на рубеж 1 млн т/год в 80-е годы и продолжает нарастать. Перспективы его производства высоки из-за чрезвычайно больших скоростей прироста биомассы, которые в тысячи раз выше, чем скорости прироста животного белка. И в то же время аминокислотный состав, особенно по квоте незаменимых аминокислот, может быть аналогичен составу животного белка. Но пожалуй, одно из главных преимуществ — возможность его наработки на непищевом сырье: это сопутствующие парафины нефти; отходы древесины и сельскохозяйственных растений; низкокалорийные для сжигания бурые угли, торфы и др. Одна из проблем, связанных с очищением микробного белка от избыточных нуклеиновых кислот, тоже может быть отнесена к разряду технологических, т. е. решаемых. Энергетическая стоимость готового продукта на основе белков микроорганизмов, пока еще довольно высокая, может быть снижена в несколько раз по сравнению со стоимостью белков говядины, производимой традиционными путями. То же относится и к выработке белка из бобовых растений. Поэтому в 90-е годы в ряде развитых стран планируется заменить 10–25% мясо-молочных продуктов растительными и микробными белками, по виду, вкусу и качеству близкими к изделиям, сейчас выпускаемым из молока и мяса.
Со второй половины нашего столетия возросла активность математического прогнозирования глобального развития эколого-экономических процессов на нашей планете. И это не удивительно. Очень резко поднялись темпы изменения лика биосферы в наше время. Каждый год конца нашего века в этом смысле стoит десятилетия его начала, столетия средних веков и тысячелетий палеолита. Поэтому необходимость количественных прогнозов очевидна. Быстрое развитие вычислительной техники позволило осуществлять расчеты динамики развития биосоциальных систем в глобальных масштабах. До сих пор мы могли изучать закономерности биосферы как уникального объекта главным образом в ретроспекции. Экспериментировать с биосферой мы не можем и не имеем права. Имитационные эксперименты на ЭВМ являются единственной возможностью системных исследований биосферы.
Первые попытки формализовать глобальное описание экологических процессов предприняты по инициативе «Римского клуба» — неофициальной организации, одним из создателей которой стал известный итальянский предприниматель Аурелио Печчеи. В первых докладах «Римскому клубу» были проанализированы модели развития общества и среды в многомерном фазовом пространстве, компонентами которого были производственные, социальные и экологические процессы (модели Форрестера и Медоузов). Если результаты расчетов по первым моделям показались обескураживающими, типа полной остановки роста экономики и снижения числа людей на планете, то в дальнейшем удалось выявить условия сбалансированного развития экологии и экономики. При этом совершенно необходимым требованием было существенное увеличение трат на охрану окружающей среды во всех вариантах положительных прогнозов (модель «ГЕЯ», СССР).
Не имея возможности проанализировать детально результаты прогнозов развития человечества в биосфере, коротко остановимся на одном из них, самом ужасном — глобальном термоядерном конфликте. С точки зрения действия энергетических принципов этот вариант не биологичен, т. е. противоречит тенденции постоянного роста энергетики и умощнения круговорота в живой природе, так как связан с глобальными разрушениями и уничтожением большого числа живых и промышленных объектов. Однако законы социального развития могут иметь свое, в том числе и трагическое для человечества, обоснование.
И по радиоактивному, и по химическому загрязнению, и по изменению климатических условий (резкое похолодание) глобальный ядерный конфликт окажется гибельным для человека и ряда высших животных и растений если не в первом, то в последующих поколениях (см. [Природа, 1985, № 6, ряд статей советских ученых]). Сама жизнь на планете не будет уничтожена, но ей придется отступить на уже пройденные позиции. Хочется верить, что вступление в ноосферу состоится в ближайшем будущем и что оно будет связано с видом Homo sapiens.
Глава 11
Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»?
Многие вещи нам не понятны не потому, что наши понятия слабы, но потому, что сии вещи не входят в круг наших понятий.
Козьма Прутков
11.1. Почему жизнь дискретна
О дискретности жизни и смене поколений образно сказал А. С. Пушкин в «Евгении Онегине»:
«Увы! на жизненных браздах
Мгновенной жатвой поколенья,
По тайной воле провиденья,
Восходят, зреют и падут;
Другие им вослед идут...»
Можно сказать, что естествознание нашего века началось с осознания дискретности мира. В физике «сплошная, гладкая» материя — континуум, с непрерывными превращениями, сменилась на нечто дискретное в пространстве и времени, с квантованными превращениями. В биологии представления о слитой наследственности, многим казавшиеся самыми естественными, с 1900 г., после «переоткрытия» законов Менделя, стали интенсивно вытесняться концепцией дискретного наследования.
Дискретность организменного уровня не требовала доказательств: настолько она очевидна, достаточно взглянуть на самих себя. Она и легла в основу эволюционных представлений об изменении во времени, эволюции форм. Дискретность на клеточном уровне была доказана экспериментально с применением микроскопов; дискретность видов лежала в основе дарвинизма; пространственная отграниченность и дискретность локальных экосистем активно изучаются в настоящее время. Итак, всю биологию пронизывает идея дискретности. И все же... И все же почему не может существовать непрерывная живая плазма, вечная во времени (типа живого мыслящего океана С. Лема)? Почему мы должны рождаться и умирать, сменяя друг друга в поколениях, а наши внуки должны нас «вытеснять из мира»?
Этот вопрос «почему» для традиционной биологии непрост, а для субстратного подхода, опирающегося на неопределенную и рыхлую идею «саморазвития», он во многих случаях просто убийственный. Действительно, зачем «саморазвивающейся» системе рвать себя на куски, отмирать, теряя при этом почти все? Разве не хватило бы простой «косметики», сохраняющей непрерывность структур? Посмотрим, как можно объяснить дискретность с позиций С + Э подхода, рассмотрев ее на разных уровнях биологической организации.
Обратимся к клеточному уровню. Для этого вернемся к самому началу, к происхождению жизни и появлению фазово-обособленных частиц в абиогенном органическом бульоне, образовавшемся под влиянием накачки энергией. Отметим: в фазово-обособленных, т. е. дискретных, единицах резко ускоряются процессы переноса энергии (электронов) и под их действием начинают работать первые циклы вещества. Таким образом, дискретность усиливает взаимодействие: энергия — вещество, ускоряя его при многократном использовании вещества, которое без циклов быстро израсходовалось бы, в то время как накачка энергией со стороны может идти вечно. Законы здесь только физико-химические, ни о каком «саморазвитии» нет смысла говорить (эволюцию клеток мы обсуждали подробно в гл. 7 и 8).
Существует ли оптимальный размер клеток? Или: почему клетке выгодно быть определенной величины? Нижний предел ясен — он определяется минимальными размерами, при которых возможно самостоятельное существование, т. е. метаболизм и воспроизводство. А каковы верхние границы? Для определения верхних размеров издавна использовался энергетический подход. С прошлого века известны правила соотношения поверхности и объема клеток, получившие название законов Рубнера. Суть рассуждений сводится к тому, что с увеличением размеров клетки энергетическая эффективность ее функционирования падает. Это очевидно из самых простых соображений: расход энергии на рост (метаболизм) клетки пропорционален массе клетки или ее объему, а приток энергии пропорционален ее поверхности, так как питание она получает через поверхность. Если представить клетку в виде шарика, то отношение поверхности к объему шарика падает с ростом радиуса по обратному закону. Следовательно, масса (m) клетки изменяется во времени таким образом:
![]()
где α и β — константы питания и метаболизма. Остановке роста соответствует ![]() . Отсюда максимальный X (радиус клетки-шарика) определяется в виде
. Отсюда максимальный X (радиус клетки-шарика) определяется в виде ![]() т. е. верхний размер определяется соотношением констант питания и метаболизма. Константа питания зависит как от внутренних характеристик, так и от внешнего потока энергии.
т. е. верхний размер определяется соотношением констант питания и метаболизма. Константа питания зависит как от внутренних характеристик, так и от внешнего потока энергии.
В самом общем виде с увеличением потока энергии константа питания возрастает вначале линейно, затем по гиперболической кривой с насыщением, а дальнейшее увеличение потока энергии может оказаться вредным для клетки. Эти качественные рассуждения хорошо демонстрируют идею дискретности на клеточном, а вообще говоря, и на организменном уровнях. Они применимы для определения максимальных размеров представителей двух основных типов живых организмов: растений и животных.
Известно, что растения, независимо от природы, растут сначала быстро, затем рост постепенно замедляется и, наконец, прекращается совсем. Интуитивно ясно, что с увеличением размеров растения увеличивается приток энергии благодаря фотосинтезу, но зато увеличиваются трудности, связанные с переносом питательных веществ, особенно с подъемом неорганических солей от корней к листьям. В конечном счете притока энергии перестает хватать для покрытия расходов, и дерево останавливается в росте. Несложные расчеты показывают, что предельное значение высоты дерева определяется энергетическими константами фотосинтеза, фотодыхания и транспорта.
Аналогичны рассуждения о предельных значениях размеров животных. Расходы энергии у животного связаны с основным обменом (аналог фотодыхания), с перемещением тела (аналог переноса вещества в растении) и с ростом. Форма уравнения для роста животного соответствует таковой для растения или отдельной клетки. Могут получаться более сложные зависимости максимального размера от концентрации корма или интенсивности энергетического потока, но дискретность везде имеет место.
Для оценки размеров водных организмов особый интерес представляет сопоставление размеров пищи и типа питания (фильтрация или активное хватание) с учетом энергетики. Из самых общих соображений понятно, что должна существовать размерная граница между пассивной фильтрацией и активным поиском и захватом пищи; очень мелкие частицы энергетически невыгодно разыскивать и хватать, так как полученная энергия не компенсирует ее расхода. Действительно, по различным оценкам, самые мелкие частицы (до 0,1—1 мкм) отфильтровываются животными, образующими слизистую сеть, через которую движениями ресничек прогоняется вода; частицы более 50 мкм преимущественно потребляются животными-хватателями, а промежуточные размеры соответствуют обоим типам питания. При этом соблюдается соответствие размеров животного-хищника и его жертвы. Пример с фильтрующими китообразными, по форме не укладывающийся в изложенную схему, хорошо подтверждает неуниверсальность С + Э подхода без учета информационных аспектов. Хорошо известна большая роль информации у этих высокоорганизованных высших животных (млекопитающих). Они буквально «пасут» стаи криля — их основной пищи, мигрируя за ними, имея громадный выигрыш прежде всего по энергетике, связанной с поиском и перемещением. Само появление хищничества оказалось возможным, начиная лишь с определенного нижнего размера тела животных, так как оно требует избыточного расхода энергии на поиск, преследование и захват жертвы. С энергетикой связано и соотношение размеров прокариотных и позднее появившихся, в сотни раз более крупных, эукариотных клеток.
Дискретность более высоких уровней тоже определяется функциональными отношениями в системе: «живое — неживое», среди которых энергетические потоки играют существенную роль. Для популяционного уровня, например, широко распространены представления об оптимальном размере популяции. Причем ограничения на численность популяции сверху непосредственно связываются с ограничением по ресурсам: на этом основано одно из наиболее известных уравнений математической экологии — логистическое. Нижний критический уровень для популяции (если он перейден сверху вниз, то популяция вымирает) определяется разнообразными причинами, но он существует и для микробных популяций, и для популяций высших организмов. В литературе представление об оптимальных размерах популяций связывается с «принципом Олли».
Для чего нужна дискретность на следующем видовом уровне? Почему она отобралась в эволюции, так как нескрещиваемость видов приводит к вымиранию целых таксонов, среди которых было немало находок, безвозвратно потерянных живой природой? И здесь можно оценить энергетическую невыгодность полного скрещивания на генном уровне из-за усложнения структуры генома и энергетической дороговизны ее содержания (см. гл. 6).
Вопрос о дискретности экосистем (биогеоценозов, биоакваценозов) хотя и дискуссионен, но, по-видимому, тоже очевиден. Дискретность этого уровня наглядна с точки зрения структуры: мы видим хорошо выраженные типы растительности, животного мира, относящиеся к определенным регионам. Границы определяются географическими особенностями, пространственным положением, а также зависят от истории развития биоты в данном месте. Нас более интересует функциональный аспект, тесно связанный с энергетикой. По функции соседние экосистемы именно тем и отличаются, что потоки энергии и круговороты вещества в них гораздо интенсивнее, чем обмены между ними. Казалось бы, дискретность экосистемам ни к чему, по крайней мере с точки зрения структурного содержания.
Однако дискретность экосистем, а с ними и биотического круговорота ярко выражена не столько в экологическом, сколько в эволюционном плане. При длительном функционировании круговорота в каком-либо месте, как мы знаем с позиций энергетики, должна происходить подгонка составных элементов круговорота в двух основных направлениях: в количественном, т. е. в увеличении потока захваченной энергии, и в качественном, т. е. в ускорении циклов лимитирующего вещества, в том числе и путем упрощения структур. И здесь мы можем видеть удивительные примеры согласованной эволюции «соучастников». Просто перечислим некоторые примеры, характеризующие разные трофические уровни: растения и их ризосфера, специфическая в каждом случае, особо интересен здесь симбиоз азотфиксирующих бактерий и растений типа бобовых; цветковые растения и насекомые-опылители; многоярусные леса; птицы, обитающие в каждом ярусе таких лесов; и т. д. Возможно, одним из самых забавных вариантов такой подгонки может служить «запрещенный» по прямой энергетике случай животного с зеленой, фотосинтезирующей поверхностью. Это ленивец, в шерсти которого во влажном тропическом лесу размножаются водоросли. Они не дают ему питания (и здесь все сходится по энергетике с расчетами), но зато маскируют его: зеленоватый оттенок спасает малоподвижного животного от крупных хищных птиц.
Итак, дискретность всех уровней биологической организации выгодна прежде всего с энергетических позиций. Поэтому для понимания ее необходимости в развитии и эволюции живого достаточно привлечь С + Э подход. Для строгих количественных оценок дискретности структур (их старения и замен) нужно привлекать и информационные аспекты, т. е. использовать в комплексе С + Э + И подход.
11.2. Почему «процветают» примитивные паразиты
Одним из главных материалистических достижений дарвинизма явился отказ от признания ламарковского внутреннего стремления к совершенству как одной из причин эволюции. (В гл. 1 мы говорили об отношении Ч. Дарвина к прогрессу, который считал, что нет направленного прогресса, нет повышения организации в эволюции.) Однако в целом повышение организации живых структур в макроэволюции явно прослеживается, о чем говорит развитие жизни от пробионта до высшего животного или растения. Отсюда и возникает неудовлетворенность дарвинизмом, поскольку повышение приспособленности, как правило, связано с усложнением структур, а в дарвинизме этого не объясняется. В то же время хорошо известны и широко распространены случаи дегенерации форм, потери ими тех сложных надстроек, которые приобретались в процессе длительного действия отбора на выживание наиболее приспособленных. Самоорганизация жизни, лежащая в основе представлений дарвинизма о движущих силах развития, никак не может объяснить явление дегенерации форм организмов в эволюции. (Зачем нужно упрощение на общем пути к самоусложнению?) Отсюда—прямой путь к агностицизму и к признанию непредсказуемости действия естественного отбора. Получается, что отбор делает, что хочет («выступает в роли господа бога» — по А. А. Любищеву), то усложняя, то, наоборот, резко упрощая структуру. Остается только post factum фиксировать все то, что он «напридумывал» в результате «мгновенного принятия решения», т. е. перейти на описательный путь и попытаться классифицировать имеющееся. Это и было тщательно проделано дарвинистами при разработке учения о разных формах прогресса. Наиболее обстоятельно учение о прогрессе освещено в трудах А. Н. Северцова [1922,1967].
Он был вынужден признавать, что общая дегенерация является полной противоположностью морфофизиологического прогресса, или ароморфоза. Действительно, у многих дегенерировавших форм целиком редуцируются органы движения, т. е. вся мускульная система, а также скелет конечностей, редуцируется центральная нервная система со всеми нервами и органами чувств и органы активного питания и, наоборот, прогрессивно развивается половая система [Северцов, 1967]. Очень существенным замечанием А. Н. Северцова является то, что он говорит о понижении и самой «интенсивности проявления жизнедеятельности при дегенерации вместе с понижением сложности организации» [Там же, т. 138]. Таким образом, дегенерация и ароморфоз полностью противопоставляются по направлению изменений как структуры, так и функции. Но тем не менее оба типа изменений могут приводить к процветанию вида, а это и означает, что их следует включить в эволюционный процесс как равноценные направления, приводящие к биологическому прогрессу (вместе с идиоадаптациями и ценогенезами). Что же, на этом пути остается еще более абстрагироваться, признать единство противоположностей и перестать искать естественно-научное, в данном случае биологическое, объяснение различия форм прогрессивного развития. По-видимому, в пределах только субстратного подхода не удается сделать большего, чем констатация имеющегося. Об этом говорил и А. Н. Северцов [1967], ограничивая рассмотрение эволюции животных тем, «как она происходит в действительности» и не рассматривая, «почему происходит именно то, а не другое изменение» (с. 125).
Рис. 18. Схема возможных изменений в системе «структура — функция» организма (I) и разрешенные (с позиций С + Э подхода) изменения структуры и функции в эволюции: А — направление дегенерации, Б — идиоадаптации и ценогенезы, В — ароморфозы (II). Заштрихована запрещенная зона.
Попробуем, в дополнение к субстратному, применить энергетический подход, т. е. использовать С + Э концепцию. Само собой разумеется, что всякое эволюционное изменение — это изменение и структуры, и функции организма. Схематически все возможные сочетания таких изменений представим в фазовой плоскости «структура—функция» (рис. 18). Стрелка 3 означает, что усложнение структуры связано с усложнением функции, в нашем случае — с увеличением потока энергии, перерабатываемого данным типом биологической структуры. Это как раз и соответствует прогрессивному изменению с позиций структурного подхода. Направлениями от стрелки 2 до стрелки 4 с разными углами наклона и будут иллюстрироваться прежде всего ароморфозы. Заметим, что сюда же следует отнести и идиоадаптации, так как энергетический подход требует хоть и небольшого, но все же усиления функции. Начиная со стрелки 4, включая 5, 6, 7 и 8, мы имеем явное ухудшение функции, что запрещается энергетическими принципами и потому не должно реализовываться. «Самыми запрещенными» являются направления, близкие к стрелке 5, т. е. усложнение структуры и ослабление функции.
Направления, примыкающие к стрелке 1, и будут характеризовать явление дегенерации, так как структура явно упрощается. Но при этом энергетический подход требует усиления функции и не разрешает ее ослабления (по крайней мере в удельном выражении). Термин «общее понижение жизнедеятельности» требует здесь некоторого уточнения. Если организм, популяция вынуждены переходить в новые, часто более простые условия существования (пещеры, большая глубина воды, отсутствие света, глубокие илы и т. д.), то по сравнению со старым местообитанием его функционирование ухудшается. И по-видимому, только упрощение структуры, избавление от ненужных на новом месте старых приспособлений позволяет сохранить и одновременно интенсифицировать функцию, т. е. повысить жизнедеятельность. Таким образом, учет энергетического подхода резко сужает разрешенные варианты развития в системе «структура — функция». Кстати, снимаются и противоположные направления, т. е. тот произвол, что был в рамках структурного подхода (см. рис. 18,II).
Очень существенно то обстоятельство, что эволюция «вниз» по функции не разрешается, если учитывать энергетические принципы. Поэтому говорить о том, что «микроорганизмы эволюционируют в стороны и вниз, но не вверх», с позиций С + Э подхода некорректно, ибо функция улучшается — такая трактовка допустима только с позиций субстратного подхода. Современные микроорганизмы отличаются от древних форм не только разнообразием структур (и иногда их серьезным упрощением), но и существенным усложнением функций: сюда входят и использование различных источников энергии и вещества, недоступных высшим формам, и интенсификация функции до уровней, тоже недостижимых для высших организмов. Но, совершенствуя и во многих случаях упрощая структуру, они под действием естественного отбора интенсифицируют функцию, т. е. эволюционируют «вверх». Как мы обсуждали в предыдущем разделе, питание растворенными веществами делает невыгодным увеличение размеров именно с позиций энергетики. Подчиняясь этому, прокариотные формы выполняют свою миссию в круговороте, «не беспокоясь» об усложнении структур, а наоборот, в соответствии с энергетическими принципами, «заботясь» об освобождении от любой структурной избыточности. Именно на микроорганизмах может быть ярче всего продемонстрирована способность необычайно быстро избавляться от лишних структур.
Как замечает Э. Брода в книге «Эволюция биоэнергетических процессов» [М., 1978, с. 61–62]: «Организмы имеют тенденцию утрачивать ненужные гены и ферменты. Это явление играет огромную роль в эволюции, и особенно в эволюции микробов...
...Автотрофы имеют тенденцию превращаться обратно в гетеротрофы, если дать им подходящие питательные вещества. И вообще, готовность к утратам — это свойство клеток, которое было хорошо изучено на простых системах». Одним из наиболее изученных и показательных фактов «гибкости» приспособления на уровне микробных популяций является смена прототрофных форм в популяции на ауксотрофные. Обычно на обедненных средах прототрофные варианты, способные производить для себя все необходимые метаболиты, быстро вытесняют ауксотрофные формы, требующие для своего развития дополнительно какого-либо определенного соединения. Если же это соединение добавлено в среду, то ауксотроф получает преимущество и способен расти быстрее, так как более экономен и не производит этого соединения в отличие от прототрофов. Этот пример с победой «дефективного» ауксотрофа при развитии на обогащенной среде может служить хорошей моделью явления дегенерации формы при переходе к паразитизму. (Дефективность, т. е. упрощение структуры ауксотрофа, может быть связана с потерей гена (делецией) в ДНК клетки, т. е. иметь генетическую природу.)

Рис. 19. Общая схема относительного изменения структуры и функции организмов в макроэволюции.
1 — прокариоты; 2 — эукариоты; 3 — многоклеточные; 4 — теплокровные; n — млекопитающие. Буквенные обозначения те же, что и на рис. 18.
Покажем схему усложнения структуры и функции организмов, начиная с пробионтной клетки и кончая млекопитающими, в том числе и человеком (рис. 19). В этом ряду несомненно усложнялась структура, но закреплялись только те варианты, которые приводили к существенному усилению функции, как того требует С + Э подход. Практически на каждом этапе, связанном с крупным ароморфозом, помимо усложнения структуры наблюдалась и дегенерация формы (структуры). На рисунке это отмечено стрелками А, загнутыми влево и вверх. Заметим, что к стрелкам А, указывающим на дегенерацию структуры млекопитающих, по-видимому, должна примыкать и стрелка Ачел, соответствующая человеку. Действительно, что касается структуры, то мы в эволюции немало потеряли: шерсть, спасающую от холода; острые клыки хищника; возможно, и хвосты, помогающие предкам лазать по деревьям. Но энергетика человека как вида возросла в десятки раз (см. гл. 9).
Общая тенденция увеличения угла наклона кривой отражает тот факт, что с повышением организации функция растет быстрее, и это во многом связано с цефализацией структуры. Следовательно, для более точных представлений о темпах эволюции необходимо привлекать и информационный подход. К этому же можно добавить, что возможность какого-либо ароморфоза у млекопитающих, связанного с сильным изменением структуры, мало вероятна. Эволюция из биологической трансформировалась в технологическую, особенно если иметь в виду энергетику человечества. Став глобальной силой, человечество способно вмешиваться в ход эволюции, переведя ее на направленный отбор, селекцию и регулируемую перестройку генотипов.
Хотелось бы обратить внимание на всеобщность и распространенность явления конвергенции, во многих случаях тесно связанного с явлением регресса формы. Конечно же, общими физическими законами определяются особенности и конкретные проявления конвергентности признаков на различных уровнях организации. Среди этих законов хорошо прослеживается действие обоих энергетических принципов. Как правило, конвергентное развитие приводит к наилучшему структурному варианту, позволяющему организму как можно больше изымать энергии (пищи) из окружения, при этом минимизируя собственные траты энергии на перемещение и ориентацию в пространстве и его использование.
Именно сходство условий существования определяет сходство форм и структур различных, а иногда и очень отдаленных таксонов. Это значит, что сходные признаки контролируются очень разными генами или генными комплексами. А это, в свою очередь, значит, что концепция: «Гены хотят жить и размножаться» — явно не состоятельна. Структура оказывается вторичной, подчиняясь функции, которая определяется местом организма (популяции, вида) в круговороте. Это заявление не имеет оттенка телеологичности, ибо с позиций С + Э подхода хорошо известно, что движущая сила развития — поток энергии — имеет физическую природу (а значит, и не имеет каких-либо целей).
Для оценки количественных аспектов изменения формы во времени требуется привлечение информационного подхода, т.е. использование С+Э+И концепции. При этом субстратный подход определит преемственность и модификации структур, энергетический — одну из главных функций, а информационный — скорости перестроек и развития. Можно надеяться, что на этом пути мы сумеем хотя бы в целом вычислять и прогнозировать эволюцию формы и функции и не только на малых отрезках времени.
11.3. Почему не выгодно прямое наследование приобретенных признаков
Проблема «наследование — ненаследование приобретенных призаков», примыкающая к задачам отношения наследственности и определенной — неопределенной изменчивости, стояла в самом центре внимания эволюционистов всех времен и направлений.
Еще Аристотель правильно угадал действие основных факторов эволюции, позднее названных мутациями и отбором: «Те органы и организмы, у которых все случайно оказалось целесообразным, выжили, а прочие погибли как неприспособленные» (цит. по [Берг, 1977, с.67]). Более глубоко идея о приспособительной эволюции была проработана Ж.Б. Ламарком, который достаточно убедительно показал, что все организмы в течение жизни приспосабливаются к окружающей среде. При этом изменяются и физические характеристики организма, и поведение — организм приобретает новые свойства. Приобретенные признаки передаются по наследству. Причиной всего этого является, по Ж.Б. Ламарку, «внутреннее стремление совершенству», заложенное Творцом.
Стоявший на материалистических позициях Ч. Дарвин резко отрицал какое-либо «стремление к совершенству», но и ему приходилось делать уступки сторонникам определенной изменчивости, преувеличивать роль «упражнения органов» в эволюции и передаче признаков по наследству. Однако абсолютизация случайности плохо увязывается с очевидностью прогрессивного развития жизни от пробионта до высшего растения или животного, включая человека.
Субстратный подход к изучению эволюции жизни, в основе которого лежит представление о самоорганизации, саморазвитии и самоусложнении живого, оставляет богатую пищу для различных толкований проблемы наследования-ненаследования. Поэтому, поднимаясь до «высоких абстракций» типа «единства организма и среды», очень просто взять на вооружение тезис о наследовании приобретенных признаков как адекватном реагировании организма на изменение условий среды, рассматривая его как адаптивную подстройку генотипа под оптимальный фенотип. (Речь идет о печальных временах доминирования псевдомарксистской диалектики Т.Д. Лысенко в нашей биологии.) Ненамного богаче в естественно-научном смысле и тезис «единства популяции и среды», хотя в философско-методологическом плане он более обоснован.
Что же в данном случае может дать учет энергетического подхода, а вернее, его сочетание с упомянутыми субстратным и информационным подходами? Можно ли попытаться объяснить, почему в эволюции «запрещено» прямое воздействие фенотипа на генотип, чем оно невыгодно? Почему справедлива центральная догма биологии — одна из крупнейших находок биологии нашего века, согласно которой информация с белка на нуклеиновую кислоту передаваться не может? Ведь совершенно очевидно, что адекватное реагирование организма, популяции, экосистемы на условия среды крайне необходимо для сохранения и эволюции всего живого! Например, из-за отсутствия обратной связи от фенотипа к генотипу для эволюции потеряно столько «блестящих» фенотипов, случайно появившихся гениальных находок, не переданных последующим поколениям. В чем же основа для существования такого запрета? Попробуем рассмотреть возможные ответы на основе комплексного подхода, т.е. С+Э+И концепции.
Вспомним, что при формулировке и обсуждении энергетического принципа интенсивного развития мы говорили как об одном из важных путей эволюции об экономии расходования энергии на образование и поддержание биологической структуры. При этом мы подчеркивали, что в прогрессивной эволюции размер генома не возрастает по абсолютной величине и что у старых таксонов он может быть гораздо больше, чем у молодых (достаточно вспомнить земноводных и птиц). Кроме того, акцентировалось внимание на том факте, что относительная доля основного носителя структуры — ДНК (и примыкающей к ней РНК) в клетках более древних прокариот выше, чем у эукариот. И наконец, мы неоднократно обсуждали примеры быстрой потери структуры (и на уровне ДНК, и на уровне целых органов), если она почему-то сталовилась ненужной. Еще раз напомним, что общая тенденция развития в сторону специализации, т.е. интенсификации определенной функции, как правило, сопровождается ослаблением других функций и потерей избыточных структур. Иначе геном раздувался бы до таких размеров, что на его воспроизведение и поддержание уходила бы большая часть энергии, захваченной организмом. А это уже прямое противоречие энергетическим принципам, согласно которым в итоге возрастает именно активная энергия и организм распоряжается ею по своему усмотрению.
Итак, запрет на разбухание генома исходит из энергетики эволюционного развития. И одним из первых под этот запрет попадает наследование приобретенных признаков. Действительно, попробуем оценить, что нужно иметь организму, чтобы унаследовать в поколениях приобретенное родителями. Прежде всего нужно уметь воспринять информацию, суметь оценить ее, принять решение о необходимости запоминания, записать в генотип, а затем подключить к системе воспроизводства информации и проявления в фенотипе.
Рассмотрим для конкретности известный пример — зеленую гусеницу на зеленом листе. По-видимому, зеленая окраска была когда-то приобретена предками гусеницы как полезный признак. Предположим, что это было передано через прямое наследование данного благоприобретения. Что для этого потребовалось бы? Во-первых, нужно было установить полезность приспособительной окраски и выбрать ее тип (т. е. зеленая на зеленом). Но гусеницы не имеют глаз и не встречаются с бабочками: т. е. и оценить, и передать информацию крайне непросто. Отрицательный опыт контакта с птицей тоже не передать в поколениях, ибо такую гусеницу уже съели и потомства от нее не будет. Но пусть все же решение приобрести зеленый пигмент принято. Как его осуществить? Надо «знать» непростую схему реакций синтеза данного пигмента, какие ферменты будут катализировать эти реакции, как синтезировать эти ферменты (скорее всего модифицировать имевшиеся белки) и т. д. Поистине сортирующий признаки демон Максвелла должен быть «дьявольски» изощрен: все знать, оценивать и уметь делать а потому — он должен быть крайне громоздок и являться тяжелой нагрузкой, прежде всего по энергетике для организма. Следовательно, организму для прямого наследования по петле обратной связи необходимо иметь сложную систему иерархически связанных структур типа: воспринимающих информацию, перерабатывающих ее и запоминающих; на следующих этапах — управляющих и исполняющих, а затем вновь оценивающих результат: надо ли закрепить данный признак или уже можно его выбрасывать. Такие сложные информационно-управляющие и исполняющие системы мы и обсуждали на примере высших животных имеющих все перечисленное: рецепторы, нервную систему, высокоорганизованный мозг и исполняющие органы. Все это постепенно развивалось под действием естественного отбора.
Правда, в данном случае необходимо подчеркнут! одно очень существенное обстоятельство: сама сложная система — мозг не имеет сверхстрогого наследования. К примеру, головной мозг человека содержит более 10 млрд нервных клеток. Эти клетки связаны друг с другом, каждая — с десятками и сотнями других — как длительными, так и динамичными, кратковременными связями. Этими связями и определяется работа мозга. Ясно, что закодировать каждую такую связь в генотипе невозможно, хотя бы потому, что у человека число генов составляет несколько миллионов, а числе связей между клетками мозга может превышать сотни миллиардов! Поэтому генетически кодируется не структура мозга во всех деталях, а способ ее образования, да и то лишь в самых общих чертах (подробности см. [Нейфах, 1978]).
И все же, несмотря на это, генотип высших животных в тысячи раз больше по абсолютной величине, чем генотипы прокариот. Считается, что лишь малая часть генома высших организмов содержит структурные гены, кодирующие белок (около 2% у мыши и человека), а остальная часть связана с работой регуляторных генов. Именно такое изменение генома можно принимать за один из немногих сформулированных принципов его эволюционных изменений в макроэволюции: «...при прогрессивной эволюции фенотипа, т. е. при возрастании морфофизиологической сложности организма, доля структурных последовательностей в геноме снижается за счет увеличения неструктурных, очевидно, регуляторных» [Медников, 1982, с. 80].
Только благодаря развитию сложной системы иерархически управляемых и управляющих структур, возглавляемых мозгом, высшие организмы сумели преодолеть запрет центральной догмы биологии: они с успехом передают свой накопленный опыт новым поколениям через воспитание и обучение как индивидуальное, так и коллективное. Энергетическую выгоду такой передачи признаков (известный теоретик эволюции Э. Майр называет ее «странным образом ламаркистской») мы обсуждали в гл. 8 и далее. С позиций С + Э + И подхода она ничуть не странна: она позволяет организму (популяции) более адекватно реагировать на условия среды, а значит, и совершенствоваться, и, что особенно важно,— без избыточного раздувания генотипа.
В связи с этим неизбежно возникает вопрос: а как же обходятся «бедные», примитивные прокариоты (и низшие эукариоты), каким образом «ухитряются» они адекватно реагировать на динамику условий существования? Ведь не имея сложных иерархических структур, способных автономно и слаженно работать по схеме: восприятие, оценка, исполнение, для передачи приобретенных признаков по наследству понадобилось бы закодировать в генотипе каждый возможный шаг в индивидуальном развитии клетки. А это и означает, что геном прокариотпой клетки или низшей эукариотной с таким типом наследования был бы гораздо больше генома высших организмов, чего не отмечалось ни разу. Более того, известно, что у прокариот большая часть генома состоит из структурных генов и связанных с ними, а у вирусов эта доля может превышать 90%. Итак, каким же образом обходился и обходится запрет центральной догмы биологии, начиная с самых низших уровней организации? Перечислим некоторые «маленькие хитрости» (а иногда и немалые), которые были взяты на вооружение живой природой.
Первым (и очень изящным!) способом приспособительного реагирования генотипа через фенотип на условия окружающей среды можно назвать способ плазмидной (вирусной) передачи генов на уровне прокариот. Его можно считать в определенном смысле универсальным: он работает и по вертикали (передача от предков к потомкам), и по горизонтали (в пределах одного поколения). Им могут быть охвачены как ближайшие родственники, так и отдаленные соседи из других семейств. Можно даже утверждать, что этот путь передачи генетической информации от клетки к клетке способен осуществляться не только «вверх» по вертикали, по и «вниз», т. е. если информация не нужна, то она не передается.
И здесь не надо придумывать ничего экстраординарного — здесь везде работает естественный отбор, который достаточно строго подчиняется действию энергетических принципов. А плазмиды являются основным поставщиком разнообразия и создают материал для работы естественного отбора. Они представляют собой внехромосомные кольцевые молекулы ДНК, способные автономно реплицироваться (воспроизводить себя) и передаваться в дочерние бактерии при делении клеток. Для многих плазмид (но не для всех) характерна способность самостоятельно переходить из одних бактериальных клеток в другие, от донора к реципиенту. Это свойство называется транспортабельностью, или трансмиссивностью. Плазмиды могут ассоциироваться друг с другом, и тогда нетранспортабельная плазмида способна «переехать» в другую клетку на плазмиде-перепосчике.
Плазмиды могут находиться в бактерии как в автономном состоянии, так и интегрироваться с хромосомой. При выходе из хромосомы (дезинтеграции) они способны «прихватить» с собой часть хромосомных генов.
Плазмиды, принося новые блоки генов в клетку, позволяют ей осваивать новые экологические ниши и успешно развиваться при ухудшении условий среды, например при действии внешних ядов — ингибиторов. Само обнаружение плазмид в 50-е годы нашего века как раз и было связано с неожиданной для человека быстро возникшей устойчивостью патогенных бактерии к антибиотикам. Практически мгновенное распространение лекарственной устойчивости среди бактерий, да еще множественные ее формы (т.е. устойчивость к нескольким антибиотикам), буквально посеяли среди химиотерапевтов панику, которая в наше время сменилась тихой растерянностью. Пока, вплоть до настоящего времени мы пытаемся побеждать очень дорогой ценой — создаем все новые и новые антибиотики. Это не лучший путь, и его недостатки мы обсуждали на страницах этой книги. Сейчас лишь подчеркнем, что С+Э подход позволяет использовать общебиологические законы для разработки стратегии более выгодной (менее энергоемкой) борьбы с лекарственной устойчивостью патогенных бактерий. Она основывается на использовании стабилизирующей формы естественного отбора: R-плазмиды резко уменьшаются в численности в популяциях бактерий, если они не нужны. Например, антибиотик карбеницилин оказалось возможным вновь использовать через два года, так как после прекращения его применения резко упала частота появления (выделения) резистентных к нему штаммов синегнойной палочки [Lowbury, 1973; цит.по.: Гольдфарб, 1980].
Другой способ перегруппировки генетического материала и его обновления связан с наличием в клетках прокариот (и эукариот) мигрирующих генетических элементов, способных к самостоятельному перемещению в пределах клеточного генома. Называют их по-разному: подвижные, мобильные, прыгающие гены и пр. Эти элементы могут включаться как в главные хромосомные репликоны, так и в дополнительные (плазмиды, эписомы фаги), вызывая их мутации, а также могут осуществлять обмен генов между различными генетическими системами. Это приводит к резкому увеличению рекомбинационных возможностей генотипа клеток.
Что касается эукариотных клеток, то они имеют мигрирующие генетические элементы как с малым (100–300) числом пар нуклеотидов, так и с большим числом повторов (до 10000 оснований). В последнее время они обнаружены у большинства таксонов: от одноклеточных грибов-дрожжей до человека. Например, в геномах млекопитающих широко распространены так называемые вездесущие короткие повторы. Они на самом деле короткие — до 100–300 нуклеотидых пар. Обнаружены они в геномах мыши и человека. Поскольку становится очевидным, что подвижные генетические элементы не экзотика, а обыденное явление, что хромосомы буквально «пестрят» этими фрагментами, то естественно возникает вопрос: а не слишком ли они «дороги» для клетки? Иногда на мобильные гены приходится по 1/4 от всей синтезируемой клеткой РНК. Избавиться от них клетка не может. Одно из распространенных мнений, что это — та самая «экзотическая» нуклеиновая кислота или «генетический паразит», который хочет размножаться даже во вред клетке. Однако, по видимому, более корректно говорить не о генетическом паразите, а о симбионте или, еще точнее, о дополнении к геному, который потому и удерживается клеткой, что помогает ей в определенных условиях выжить. Если бы эти элементы были бы только лишней нагрузкой, то они быстро элиминировались бы из популяции. И неважно, что несущая их клетка не может избавиться от них. Популяция под действием естественного отбора быстро очистилась бы от клеток с избыточной нагрузкой. И для таких микроэволюционных событий (очищений от избыточных структур в соответствии с энергетическими принципами), особенно в популяциях быстро размножающихся про- и эукариот, понадобились бы считанные недели и месяцы. А коль этого не происходит, то следует обратить внимание на полезность мобильных элементов для клетки популяции.
Как и бактериальные транспозоны, мобильные гены эукариот способны влиять на активность генов, в соседство с которыми они попали, перепрыгивая с места на место и меняя свою численность. Эти мобильные гены иногда обнаруживают большое сходство с прокариотными транспозонами и ретровирусами птиц и млекопитающих. При перемещениях в геноме они также могут «прихватить с собой» соседние структурные гены. Известно, что большая часть нестабильных мутаций (мутаций с высокой частотой возврата к нормальному фенотипу) связана с прыгающими генами.
Таким образом, мобильные генетические элементы и про- и эукариот, резко увеличивая рекомбинационные способности, позволяют клетками и популяциям адекватно реагировать на условия среды без прямого наследования приобретенных признаков. И адекватность эта, как мы видели, достигается «малой кровью», т.е. относительно низкой долей затрат на дополнительные структуры.
Мы не говорили о способе рекомбинации генетического материала с помощью половых процессов не потому, что он несущественен, а потому, что он наиболее изучен и общеизвестен. Ознакомление с другими механизмами, обеспечивающими приспособленность популяций низших организмов к условиям среды, показывает, что адекватное реагирование возможно и без прямого наследования полезных признаков. Эффективность его обеспечивается у популяций низших организмов за счет быстрой смены поколений (т.е. через действие отбора) и рекомбинации прежде всего дополнительного генетического материала и с его помощью — основного генома. На этом уровне вполне хватает тех механизмов неопределенной изменчивости, которые мы только что рассматривали. Определенная, т.е. направленная изменчивость оказывается ненужной, ибо она привела бы к резкому разбуханию генотипа. И потому она здесь запрещена, хотя и обеспечивала бы наиболее быстрое и адекватное реагирование, приспособление организмов к окружающей среде.
На магистральном направлении эволюции хищников она проявляется в виде воспитания и обучения потомков. Удлинение сроков жизни и уменьшение числа потомков, как мы знаем, характерно для магистрального направления эволюции, так как на этом пути экономится лимитирующее вещество и интенсивность работы биологических структур возрастает. Но именно длинные интервалы жизни и малое число потомков резко снижают эффективность действия отбора, а с ним и обнаруживает недостаточность неопределенной изменчивости. И в этих условиях возрастает роль информационно управляющих структур, прежде всего мозга, которые способны оценивать и запоминать информацию без строгого генетического контроля. Отсюда и возрастание роли направленной определенной изменчивости, «такой ламаркистской по сути». Наиболее ярко она проявляется у человека в его общественном поведении, в системе коллективного образования и воспитания. Естественно, что здесь работают уже не биологические, а социальные законы, человек перешел от биологической к культурной и экономической эволюции, он не пассивно подстраивается к среде, а активно ее перестраивает (это уже выходит за рамки нашего биологического рассмотрения).
Глава 12
Перспективы отдаленного будущего:
человеческий разум и судьбы Вселенной
... В каждой мимолетности вижу я миры,
полные изменчивой радужной игры.
Не кляните, мудрые. Что вам до меня?
Я ведь только облачко, полное огня...
К.Д.Бальмонт
Итак, с позиций структурно-энергетического подхода к изучению эволюции мы можем подчеркнуть три важнейших положения. Во-первых, неизбежность возникновения жизни на нашей планете (т.е. в определенных условиях). Во вторых, наличие магистрального направления ее развития, ведущего к появлению и совершенствованию информационно-энергетических структур вплоть до появления разума в некоторых звеньях биологического круговорота. В третьих, рост «энерговооруженности жизни», и прежде всего разума. Рассмотрим эти положения чуть подробнее.
Жизнь на нашей планете развилась как результат ускорения вращения вещества под влиянием накачки энергией со стороны. Фазово-обособленные комочки делали это быстрее гомогенного окружения и победили в отборе на встраивание в круговорот. (В добиогенном этапе прошел отбор на самый энергоемкий и быстрый носитель — воду, на основе которой и развилась жизнь.) Напомним, что ускорение потока вещества в живой природе не просто сопутствующее явление. Во многих реакциях в живых организмах скорости трансформации веществ возросли по сравнению с абиотическим окружением в сотни миллионов — десятки миллиардов раз! К случайностям относить такие события, конечно, нельзя; использовать туманные представления о том, что это нужно для самоорганизации, в данном случае просто не требуется.
Наличие магистрального направления жизни, ее развития в сторону совершенствования энергетики естественным образом ведет к совершенствованию все более сложных информационно-управляющих структур (путь к разуму). Но это не путь их самосовершенствования или саморазвития (по своей или чужой воле), а путь вынужденного развития, встраивания в круговорот. Выживают те структуры, которые способны наиболее быстро перерабатывать информацию, чтобы перераспределять и увеличивать поток энергии, организовывать разнообразнейшие циклы. Напомним данные, определяющие среднюю энергетическую активность среднего млекопитающего — человека. Количество энергии, которую расходует в секунду взрослый человек весом 70 кг, около 100 Дж, т.е. примерно 1,5 Вт/кг. Сравним с удельным излучением Солнца (E = 3,8·1026 Вт; M=2·1030 кг) — это чуть меньше 2·10–4 Вт/кг. Таким образом, удельная энергетическая активность млекопитающего почти в 10 тыс. раз выше, чем у нашего светила. Это тоже не случайно!
И наконец, обратимся к динамике роста энерговооруженности коллективного разума. Современный человек в совей экономике вовлекает в оборот в среднем в 20 раз большее количество энергии, чем показатели основного обмена, приведенные выше. В развитых странах эта величина к началу 70-х годов возросла в 50–100 раз, а обоснованный прогноз говорит о ее удвоении к концу этого столетия. (Это — удельная характеристика; общий поток энергии возрастет в 4 раза, так как численность населения удвоится.) И хотя экспоненциальный рост энергетики человечества не может длиться долго (см. рисунки в гл. 9), не стоит забывать, что мы на пути к управляемому термоядерному синтезу.
Таким образом, общий итог рассмотрения частного случая развития жизни на нашей планете указывает на неизбежность ее возникновения, энергетического совершенствования и развития осознающих себя структур. Каковы необходимые (и, осмелимся сказать, достаточные) условия для ее появления и эволюции? Главное условие — поток энергии, который извне должен непрерывно «возмущать» косное вещество, организуя циклы вещества. Твердотельные или газовые циклы не могут привести к появлению сложных активных структур: в первом случае — из-за слишком сильных, а во втором — из-за слишком слабых связей между взаимодействующими единицами. Оба этих крайних случая могут приводить лишь к упрощению структур, давая два крайних типа: чистый кристалл или гомогенный газ. Молекулы или атомы твердого тела слишком связаны, а газа — слишком независимы друг от друга. Поэтому «популяционного взаимодействия» особей и отбора наиболее приспособленных, частично измененных вариантов здесь ожидать практически невозможно.
Такого рода возможность (отбор и его полезность для всей популяции) реализуется только в промежуточном случае, т.е. в жидкой фазе. Но не обязательно это — вода и не обязательно привязываться к полимерам на углеродной основе, такой «водно-углеродный шовинизм» характерен для субстратного подхода. Возможны, например, варианты с жидким аммиаком и кремниевыми полимерами. Следовательно, близкая нам, т.е. наша субстратная, основа жизни может быть случайной и характерной только для Земли. Не случайным должно быть лишь ускорение циклов вещества и энергетическое (плюс информационное) совершенствование такого же типа, который мы наблюдаем, изучая эволюцию нашей жизни.
Как ни «прискорбно», но приходится осознавать, что субстрат жизни на других планетах с высокой степенью вероятности не похож на наш ни по составу, ни по форме организмов. Но такой вывод можно считать обнадеживающим. Он снимает налет «уникальности» жизни и освобождает от антропоцентризма и других «шовинизмов». Действительно, трудно представить себе нашу исключительность. Напомним, что в нашей Галактике примерно две сотни миллиардов (2·1011) звезд. И где-то в одном из отдаленных спиральных рукавов светит не очень яркая, желтая звезда — Солнце (звезд такого класса в Галактике — миллионы) . Планетные системы при таких звездах, согласно расчетам на современных ЭВМ в США и СССР, должны быть весьма обычным явлением.
Можно ли утверждать, что мы одиноки во Вселенной, потеряны «как иголка в стоге сена», что именно «наша жизнь — уникальное явление» (по И.С. Шкловскому). Эту гипотезу можно защищать, если оставаться на методологических позициях только субстратного подхода с его идеей внутреннего саморазвития и самосовершенствования жизни (грубо говоря: хочу — развиваюсь, хочу — нет).
Субстратно-энергетический подход определяет концепцию «презумпции естественности», и даже более — неизбежности возникновения жизни. Для этого нужно немного: потоки энергии, которые пронизывают в виде излучений (полей) всю Вселенную, и достаточный их уровень, чтобы устроить в скоплениях вещества, в жидких фазах планет «игры» отбора структур, способных наименьшим веществом с высокой скоростью взаимодействовать с этими потоками.
Попробуем оценить число планет, на которых имеются цивилизации с высокоразвитыми (по энергетике) технологиями. Для этого используем широко известную формулу Дрейка в применении к нашей Галактике:
N = n · P1 · P2 · P3 · P4 · t/T.
Здесь n — полное число звезд в Галактике; множители Pi имеют вероятностный характер: P1 — вероятность того, что звезда имеет планетную систему; P2 — вероятность наличия на планете жизни; P3 — вероятность существования разума; P4 — вероятность наличия высокоразвитой технологии, прежде всего по энергетике; t — средняя продолжительность технологической эры; T — возраст Галактики. Определенные численные значения имеют n и T. Для нашей Галактики это 2·1011 звезд и 15·109 лет ее существования. С позиций субстратного подхода вероятностные величины совершенно не определены и, пожалуй, неопределяемы. Особенно не ясно, что такое t — время существования технологической эры, так как предполагается, что цивилизации (как всякие структуры: организмы, виды, государства) должны существовать конечное время. Применение С+Э подхода позволяет существенно уточнить обсуждаемые характеристики.
Прежде всего, для оценки величин P2, P3, P4, вероятностей наличия жизни, разума и развитой технологии соответственно нужно взять довольно высокие цифры. Весь предыдущий анализ показал, что жизнь возникает с неизбежностью и быстро развивается в сторону умощнения энергетики при наличии возмущающих потоков энергии и определенных (не очень узких) условий среды на планете. Если доля звезд с подходящими планетами составляет 1% (т.е. P1 ~ 0,01), то можно положить Pi = 0,1÷1 (где i = 2, 3, 4). Величина 0,1 (или 10%-ная вероятность) явно занижена, но не будем фаталистами. Тогда число развитых цивилизаций будет
N = 2 · 1011 · 10–2 (0,1÷1) (0,1÷1) (0,1÷1) t/T.
Оценим время существования технологически развитой цивилизации. Совершенно не требуется, чтобы она отмирала со временем. С позиций субстратно-энергетического подхода это — нонсенс! Не особая структура государства, не отдельный этнос определяют технологию: от палеолита, рабовладельческого строя и далее энергетика цивилизаций росла, несмотря на смены ее конкретных носителей, народов и государств. Факел энергетики разгорался со временем все ярче, хотя иногда и потрескивал и помигивал. Поэтому энергетическая технология не имеет тенденций к отмиранию, напротив, она только совершенствуется — того требует С+Э подход к изучению эволюции на нашей планете. Технически развитая цивилизация, таким образом, никак не обречена. Время ее существования зависит только от продолжительности ее зарождения и развития. Для нашей планеты оно составляет 4 млрд лет. Округляя в «худшую» сторону, время задержки можно положить в среднем равным половине T, т.е. 7–8 млрд лет. («Ухудшение» действительно явное, если не забывать, что планета наша и светило заметно периферийны) Таким образом, отношение t/T будет равно примерно половине единицы. Теперь найдем и границы числа возможных развитых цивилизаций:
![]()
Итак, мы насчитываем огромное количество развитых цивилизаций, среди которых многие должны быть более продвинутыми в своем развитии, чем наша. Отсюда возникает другой, прямо противоположный аспект: если жизнь типична, то почему мы, «провинциалы», не столкнулись с ее проявлениями из «метрополий», например из центра нашей Галактики? Вопрос существенный: иногда на него отвечают гипотезой «направленной панспермии», как это сделал Ф.Крик; иногда его называют одним из основных «парадоксов» (по И.С. Шкловскому). Ответ на вопрос о контактах с разумными существами мы попытаемся обосновать чуть позднее, а пока еще раз констатируем, что с позиций С+Э подхода (с добавлением И) жизнь, в том числе осознавшую себя, следует рассматривать как обычное, типичное, распространенное явление, где разум служит посредником в трансформациях энергии и вещества.
Один из способов наиболее полного использования энергии звезды для нужд разумной жизни описан Ф. Дайсоном более 30 лет назад. Например, чтобы планета Земля использовала не одну двухмиллиардную долю солнечного излучения, а все излучение целиком, необходимо построить гигантскую сферу радиусом около 150 млн км. Поверхность ее будет в миллиард раз больше поверхности земного шара; ясно, что население также может увеличиваться в миллиарды раз. Для увеличения потоков энергии можно использовать вещество крупных планет типа Юпитера (в термоядерных реакциях) и т.д. Интересно отметить, что звезда при сооружении такой искусственной биосферы становится невидимой. В этом месте возникает сильный источник инфракрасного, теплового излучения. (Такие источники можно обнаружить только за пределами нашей атмосферы, непрозрачной для слабых тепловых лучей.)
Итак, будем считать жизнь ординарным феноменом, имеющим всегалактическое распространение. Конечно, это не значит что каждая частичка материи участвует в живых структурах. Мы утверждаем только то, что жизнь, с высокой степенью вероятности, можно встретить в различных местах Вселенной, на разных фазах ее развития. Тогда можно спросить, если жизнь не случайна, а естественна, то какова ее роль во Вселенной? Для ответа на такой «сакраментальный» вопрос (типа: «кому нужна жизнь», «зачем мы живем», «зачем живу я» и т.д.) нужно принять во внимание, что мы живем в нестабильной, нестационарной Вселенной.
В настоящее время наша Вселенная расширяется в пространстве и неясен конечный результат этого расширения. Однако по крайней мере практически ясно, что данный этап ее развития начался с Большого взрыва (Big Bang) около 15 млрд лет назад с бесконечных величин температур, давлений и плотностей вещества. Усилиями теоретиков удалось нарисовать более или менее полную картину такого взрыва, начиная с миллионных долей секунды до его начала.
По последним оценкам того же Ф.Дайсона, даже если земная жизнь уникальна, то для широкой экспансии в космос ей нужно будет преодолеть три барьера. Он образно пишет: «Нужно научиться жить и быть счастливыми при g-нуле, T-нуле и P-нуле, что означает: при нулевой гравитации, при нулевой температуре и нулевом давлении» [Дайсон, 1982, с.67]. Не останавливаясь на механизмах адаптации жизни к этим трем нулям (по Дайсону это возможно для нашей формы жизни), отметим, что такая адаптация очень важна, если нас ждет судьба «замерзающей» Вселенной. Этот вариант соответствует открытой модели вечно расширяющейся Вселенной, в случае если в ней слишком мало вещества, чтобы (на основе гравитации) остановить расширение. Отрадно, что, по расчетам физиков, даже в таком варианте энергетического умирания жизнь сможет долго просуществовать, хотя все-таки прекратится.
Другой случай, если масса вещества больше критической (около 10 атомов на 1 м3, или 10–29 г/см3), относится к более «опасному» для жизни варианту — закрытой модели Вселенной. Это значит, что расширение затормозится силами тяготения, прекратится и затем начнется сжатие Вселенной до тех бесконечных температур и давлений, при которых мыслимая нами жизнь абсолютно невозможна, разрушаются даже атомы и ядра элементов. Представим оба этих случая (рис. 20 а). Очевидно, что за сжатием должно последовать очередное расширение.
Теперь рассмотрим наиболее интересную возможность, определяемую промежуточным вариантом. По современным оценкам, плотность материи во Вселенной примерно равна критической, если принять во внимание массы нейтрино, черных дыр, межзвездного газа и прочих (см., например, [Хокинг, 1985]). Трудно себе представить, что она в точности равна критической, что означало бы крайне невероятный случай абсолютного совпадения. Отсюда следует одна очень важная возможность. Законы сохранения вещества и энергии для Вселенной объединяются в единый закон сохранения вещества и энергии, так как они взаимопереходят друг в друга в соответствии с формулой Эйнштейна (E=mC2). Изменяя соотношение между E и m, т.е. изменяя гравитационную массу, можно повлиять на гравитационные силы притяжения. А это и означает, что можно управлять процессом расширения-сжатия Вселенной, удерживая ее размеры (радиус) в квазистационарном состоянии. И именно эту роль регулятора сможет выполнять разум, который способен осознавать окружающий мир и прогнозировать его развитие (рис. 20, б). Такова возможная галактическая роль разума, вытекающая из анализа развития частного случая жизни на маленькой планете с «обочины» нашей Галактики. Теперь можно попытаться ответить на вопрос: почему мы не вышли на контакты с другим, более мощным цивилизациями? Мы им пока просто не нужны, у них заботы помасштабнее. Контакт через огромные пространства очень энергоемок, ничего особенного от нас они не ждут, ибо мы развиваемся по обычному сценарию (может быть, только с лишними ошибками). Если мы и хотим чего-то получить, то это — исключительно наша забота. И за это ничтожно короткое время наших поисков мы пока не вправе надеяться на находку, а тем более на активный поиск нас. «Спасение утопающих — дело рук самих утопающих», — этот лозунг, помимо саркастического, имеет и деловой оттенок.

Рис. 20. Схематическая картина изменения размеров Вселенной во времени.
а — закрытая и открытая модели [Хокинг, 1985]; б — добавлена волнистая линия квазистационарной Вселенной, регулируемой разумом.
В самом деле, если нам так хочется выйти на связь с внеземными цивилизациями, то мы должны прежде всего разработать строгую теорию эволюции жизни на нашей планете. Это — долг, а точнее, задолженность естествоиспытателей. Мы должны уметь построить количественный прогноз развития жизни, уметь вычислить темпы ее эволюции и не только описывать прошлое (как это делает современная биология), но и точно рассчитать будущее развитие. Второе условие связано с более обстоятельным изучением структуры и динамики Вселенной, начиная с нашей Галактики. Здесь наши возможности пока не очень велики: они почти не выходят за рамки простых наблюдений. И загадок пока — огромное количество. Например, известен факт, что вблизи центра нашей Галактики расположено огромное, гораздо больше Солнца, облако с почти земной температурой (около 0°C). Поскольку излучение ядра Галактики, т.е. потока энергии, в миллион раз мощнее солнечного, а само ядро гораздо старше (на миллиарды лет) нашего светила, то где же, как не там, следует ожидать существования сверхцивилизации? [Мухин, 1980]. Так же, как не следует ожидать развития жизни на холодном, замороженном Марсе, если конечно, она не попала с Земли. Возможно, что цивилизации центра уже имеют точный график прибытия представителей различных периферийных цивилизаций с рапортами об уровне развития и о собственных находках (не только ошибках).
Для оценки возможностей разума по влиянию на судьбы Вселенной приведем космологическую гипотезу Я.Б. Зельдовича, связанную с объяснением причин рождения и разбегания Вселенной. В соответствии с этой гипотезой при определенных, очень малых размерах Вселенной может возникать сильнейшее отрицательное давление, или расталкивание силами тяготения. Детальное рассмотрение этой интересной идеи не входит в нашу задачу, но один из главных выводов гипотезы сводится к тому, что Вселенная замкнута. Следовательно, суммарная плотность вещества ней больше критической и Вселенная обречена на сжатие и очередной коллапс. По оценкам Я.Б.Зельдовича, средняя плотность превышает критическую на очень малую величину, порядка 10–10, т.е. на одну стомиллионную долю процента. Известно, что энергия фотонов в современной Вселенной составляет в эквивалентных единицах выше 10–4 от критической массы (2·10–4 ρкрит). Таким образом, для относительно разумного регулирования (надо, чтобы осталось место и для существования самих регулирующих структур) имеется приличный запас, т.е. миллион раз.
Конечно, гипотеза о разумном регулировании расширения — сжатия Вселенной кажется фантастической, но она не беспочвенна. При ее обсуждении не требуется вводить новых сущностей, достаточно только напомнить, что биология и физика — это часть единого естествознания, а земной разум — не уникальное явление. Сама по себе она, например, гораздо менее фантастична, чем довольно широко обсуждаемая гипотеза о множественности Вселенной, согласно которой объект типа нашей Вселенной является как бы отдельной молекулой.
Хотелось бы верить в то, что земной разум может внести свою (пусть скромную) лепту в регулирующую космическую функцию вселенского разума, и надеяться, что этот вклад будет достоин вида, который присвоил себе определение — разумный.
Использованная и рекомендуемая литература
- Агесс П. Ключи к экологии. — Л.: Гидрометеоиздат, 1982.
- Айала Ф. Введение в популяционную и эволюционную генетику. — М.: Мир, 1984.
- Алексеев Г.Н. Энергоэнтропика. — М.: Знание, 1983.
- Берг Л.С. Труды по теории эволюции. — Л.: Наука, 1977.
- Бернал Дж. Возникновение жизни. — М.: Мир, 1969.
- Блюменфельд Л.А. Проблемы биологической физики. — М.: Наука, 1977.
- Борзенков В.Г. Биология и физика // Знание. Сер. биол. — 1982. — № 10.
- Борзенков В.Г., Северцев А.Н. Теоретическая биология: размышления о предмете // Знание. Сер. биол. — 1980. — №9.
- Будыко М.И. Глобальная экология. — М.: Мысль 1977.
- Будыко М.И. Эволюция биосферы. — Л.: Гидрометеоиздат, 1984.
- Вернадский В.И. Биосфера // Собр. соч. — М., 1960. — Т.5.
- Вернадский В.И. Химическое строение биосферы Земли и ее окружения. — М.: Наука, 1965.
- Вернадский В.И. Живое вещество. — М.: Наука, 1978.
- Вильямс В.Р. Агрономия // Собр. соч. — М., 1949. — Т. 10.
- Винберг Г.Г. Энергетический принцип изучения трофических связей и продуктивности экосистем // Зоол. журн. — 1962. — Т. 41, вып. 11. — С.1618–1630.
- Волькенштейн М.В. Сущность биологической эволюции // УФН. — 1984. Т. 143, вып. 4. — С.429 — 466.
- Воронцов Н.Н. Теория эволюции: истоки, постулаты и проблемы // Знание. Сер. биол. — 1984. — № 7.
- Гленсдорф П., Пригожин И. Термодинамическая теория структуры, устойчивости и флуктуации. — М.: Мир, 1973.
- Гольдфарб Д.М. Генетическая коммуникация у бактерий // Журн. ВХО им. Д.И. Менделеева. — 1980. — № 4. — С.372–382.
- Дайсон Ф. Дж. Будущее воли и будущее судьбы // Природа. — 1982. — № 8. — С. 60–70.
- Дарвин Ч. Происхождение видов путем естественного отбора. — 2-е изд. — Спб.: изд. В.И.Губинского, 1912.
- Дикерсон Р.Е. Химическая эволюция и происхождение жизни // Эволюция. — М.: Мир, 1981. — С.67–107.
- Егоров В.В. Землю надо беречь // Наш современник. — 1985. — № 7. — С. 129–130.
- Заварзин Г.А. Бактерии и состав атмосферы. — М.: Наука, 1984.
- Залыгин С.П. Поворот. — М.: Мысль, 1987. — 78 с.
- Зотин А.И., Зотина Р.С. Экспериментальная и теоретическая основы качественной феноменологической теории развития // Термодинамика биологических процессов. — М.: Наука, 1976. — С. 47–77.
- Израэль Ю.А. Кислотные дожди. — Л.: Гидрометеоиздат, 1983.
- Кадомцев Б.Б., Рыдник В.И. Волны вокруг нас. — М.: Знание, 1981.
- Карцев В., Хазановский П. Тысячелетия энергетики. — М.: Знание, 1984.
- Кедров Б.М., Серебровская К.Б. Проблема происхождения и сущности жизни и ее философский аспект // Журн. ВХО им. Д.И. Менделеева. — 1980. — № 3. — С.225–266.
- Ковда В.А. Биогеохимические циклы в природе и их нарушение человеком. — М.: Наука, 1975.
- Ковда В.А. Как помочь нашим черноземам // Наш современник. — 1985. — № 7. — С. 117–128.
- Левонтин Р. Генетические основы эволюции. — М.: Мир, 1978.
- Легасов В.А., Феоктистов Л.П., Кузьмин И.И. Ядерная энергетика и международная безопасность // Природа. — 1985. — № 6. — С.6–16.
- Майр Э. Зоологический вид и эволюция. — М.: Мир, 1968.
- Майр Э. Эволюция // Эволюция. — М.: Мир, 1981. — С.11–31.
- Маргелис Л. Роль симбиоза в эволюции клетки. — М.: Мир, 1983.
- Медников Б.М. Происхождение и эволюция нуклеиновых кислот // Журн. ВХО им. Д.И. Менделеева. — 1980. — № 4. — С.425–431.
- Медников Б.М. Закономерности эволюции генома // Молекулярные механизмы генетических процессов . — М.: Наука, 1982. — С.76–85.
- Медников Б.М. Монофилия органического мира и эволюция экосистем // ЖОБ. — 1985. — № 4. — С.462–470.
- Мигдал А.Б. Поиски истины. — М.: Молодая гвардия, 1983.
- Морозов Л.Л. Поможет ли физика понять как возникла жизнь? // Природа . — 1984. — № 5. — С.35–42.
- Мухин Л.М. Внеземной разум // Химия и жизнь. — 1980. — № 5. — С.89–93.
- Нейфах А.А. Слишком много ДНК, Слишком много активных генов, слишком много признаков // Химия и жизнь. — 1978. — № 12. — С.23–28.
- Ничипорович А.А. Фотосинтез // Природа . — 1967. — № 6. — С.21–27.
- Новиков И.Д. Эволюция Вселенной. — М.: Наука, 1983.
- Одум Г., Одум Э. Энергетический баланс человека и природы. — М.: Прогресс. — 1978.
- Одум Ю. Основы экологии. — М.: Мир, 1975.
- Певзнер Л. Основы биоэнергетики. — М.: Мир, 1977.
- Перельман А.И. Биокосные системы Земли. — М.: Наука, 1977.
- Печуркин Н.С. Популяционная микробиология. — Новосибирск: Наука, 1978.
- Печуркин Н.С. Энергетические аспекты развития надорганизменных систем. — Новосибирск: Наука, 1982.
- Полищук В. Третье начало биодинамики? — Химия и жизнь. — 1978. № 8. — С.26–33.
- Поннамперума С. Происхождение жизни. — М.: Мир, 1977.
- Реймерс Н.Ф., Роздин И.А. Ступени к безотходному хозяйству // Химия и жизнь. — 1981. № 1. — С.12–15
- Риклефс Р. Основы общей экологии. — М.: Мир, 1979.
- Розен Р. Принцип оптимальности в биологии. — М.: Мир, 1969.
- Свирежев Ю. М., Логофет Д.О. Устойчивость биологических сообществ. — М.: Наука, 1978.
- Северцов А.Н. Эволюция и психика. — М.: Изд-во братьев Сабашниковых, 1922.
- Северцов А.Н. Главные направления эволюционного процесса. — М.: Изд-во МГУ, 1967.
- Сент-Дьёрдьи А. Введение в субмолекулярную биологию. — М.: Наука, 1964.
- Скалкин Ф.В., Канаев А.А., Копп И. З. Энергетика и окружающая среда. — Л.: Энергоиздат, 1981.
- Соколов Б.С. Вендский период в истории Земли // Природа. — 1984. — № 12. — С. 3–18.
- Татаринов Л.П. Направленность филогенетических процессов и прогнозируемость эволюции // ЖОБ. — 1985. — № 1. — С.3–19.
- Татаринов Л.П. Суждение палеонтолога об эволюции. — Знание-сила. 1987. — № 2. — С. 55–61.
- Тимофеев-Ресовский Н.В. Генетика, эволюция и теоретическая биология // Природа. — 1980. — № 9. — С.62–65.
- Титлянова А.А. Биологический круговорот углерода. — М.: Наука, 1979.
- Уиттекер Р. Сообщества и экосистемы. — М: Прогресс, 1980.
- Усольцев Н.В. Время возвращать долги // Наш современник. — 1985. — № 7. — С. 150–152.
- Хокинг С. Край Вселенной // Природа. — 1985. — № 4. — С.21–28.
- Чирков Ю.Г. Занимательно об энергетике. — М.: Молодая гвардия, 1981.
- Шварц С.С. Экологические закономерности эволюции. — М.: Наука, 1980.
- Шкловский И.С. Вселенная, жизнь, разум. — М.: Наука, 1987. — 320 с.
- Шмальгаузен И.И. Факторы эволюции. — М.: Советская наука, 1946.
- Шредингер Э. Что такое жизнь (с точки зрения физика). — М.: Атомиздат, 1972.
- Эйген М. Самоорганизация материи и эволюция биологических макромолекул. — М.: Мир, 1973.
- Яблоков А.В., Юсуфов А.Г. Эволюционное учение. — М: Просвещение, 1976.
- Яншин А. Без ссор с природой. — Знание-сила. — 1987. — № 4. — С. 1–8.