Биология
ВЕЛИКИЙ СИМБИОЗ: ПРОИСХОЖДЕНИЕ ЭУКАРИОТНОЙ КЛЕТКИ
Владимир МалаховСто лет назад российский биолог К.С. Мережковский высказал предположение, что эукариотная клетка возникла в результате симбиоза нескольких самостоятельных организмов. Эта идея стала одной из главных парадигм современной биологии.
Все живые организмы, населяющие нашу планету, делятся на две большие группы: прокариоты (безъядерные) и эукариоты (ядерные). Прокариоты - это бактерии, у которых наследственный материал представлен простой кольцевой молекулой ДНК. Ядерными называются различные одноклеточные и многоклеточные организмы (простейшие, растения, животные и грибы), в клетках которых имеется оформленное ядро с хромосомами, в которых линейные молекулы ДНК связаны с особыми ядерными белками - гистонами. Помимо ядра в клетках эукариотных организмов есть и другие органеллы: митохондрии, жгутики, хлоропласты. Когда и как возникли эукариотные организмы, господствующие в современной биосфере?
 |
| Земля в первые полмиллиарда лет ее существования была лишена воды. Озера огненной лавы чередовались с застывшими лавовыми полями |
Согласно современным представлениям, наша планета сформировалась около 4,5 млрд. лет назад. Первоначально Земля была сухой, вода появилась в результате дегазации недр - выхода в атмосферу водяного пара и газов, составлявших древнюю атмосферу. По мере конденсации водяного пара появлялись сначала мелкие лужицы, которые понемногу становились все больше и больше. Однако понадобилось 500-700 млн. лет для того, чтобы на Земле возникли более или менее крупные водоемы, которые постепенно сформировали гидросферу - жидкую оболочку нашей планеты, занимающую в настоящее время около 70% ее поверхности. Затем в результате оседания на дно водоемов различных частичек образовались и осадочные породы.
Древнейшими осадочными породами считаются графитизированные сланцы из формации Исуа в Гренландии - их возраст составляет около 3,8 млрд. лет. Удивительно, что в этих породах обнаружены несомненные признаки некогда существовавшей жизни - следы деятельности организмов, осуществлявших процесс фотосинтеза. Дело в том, что в органическом веществе, созданном в процессе фотосинтеза, соотношение изотопов углерода 12С и 13С меняется в пользу более легкого изотопа 12С. И что бы с данным веществом ни происходило в дальнейшем, такое соотношение в нем будет сохраняться. Углерод в сланцах формации Исуа - явно органического происхождения. Это означает, что уже 3,8 млрд. лет назад в первичных водоемах планеты (скорее всего Мирового океана в то время еще не существовало) жили организмы, способные к фотосинтезу. Окаменевшие клетки, сходные с современными цианобактериями, обнаружены в породах возрастом 3,5 млрд. лет (формация Варравуна в Австралии). В чуть более молодых отложениях (более 3,1 млрд. лет) найдены остатки хлорофилла - фитан и пристан, а также специфические пигменты цианобактерий - фикобилины.
Разумеется, среди организмов той поры были не только фотосинтетики, использующие энергию солнечного света, но и хемосинтетики, получающие энергию за счет различных химических реакций. В первые миллиарды лет существования биосферы вследствие деятельности хемосинтетических бактерий сформировались многие (если не большинство) из рудных залежей, которыми до сих пор пользуется человечество, поэтому в рудных телах нередко находят окаменевшие остатки бактерий. Например, такое крупное месторождение железных руд, как Курская магнитная аномалия, по современным данным, образовалось в результате деятельности бактерий.
Нет сомнения в том, что на протяжении значительной части своей истории (не менее 2 млрд. лет) биосфера была прокариотной, то есть в ее состав входили только организмы, сходные с современными бактериями. Эукариотные организмы - разнообразные одноклеточные простейшие, а позднее (600-800 млн. лет назад) и многоклеточные организмы - заняли свое место в биосфере лишь около 1 млрд. лет назад.
 |
| Вода, выделившаяся с вулканическими газами, проливалась на поверхность планеты. Этот процесс сопровождался мощными электрическими разрядами, которые способствовали синтезу сложных органических молекул. |
Прокариоты и эукариоты - две главные разновидности живых существ на нашей планете. Биологи и медики, правда, активно изучают еще одну группу биологических объектов - вирусы, но они проявляют свойства живого организма только внутри клеток своих "хозяев". Размеры прокариотных клеток в большинстве случаев колеблются от 0,5 до 3 мкм, а самые мелкие (микоплазмы) не превышают 0,10-0,15 мкм. Гигантские клетки некоторых серобактерий достигают 100 мкм в длину, а клетки спирохет иногда вырастают до 250 мкм. Главная черта прокариот - отсутствие ядра. Их генетический материал (генофор) представлен единственной кольцевой молекулой двухцепочечной ДНК, закрепленной на цитоплазматической мембране, одевающей клетку. Прокариоты не имеют ядерной оболочки, эндоплазматического ретикулюма (иногда имеются впячивания поверхностной мембраны - так называемые мезосомы), митохондрий, пластид и других цитоплазматических органелл, свойственных эукариотам. У них отсутствуют и микротрубочки, поэтому они не имеют ни центриолей, ни веретена деления. Рибосомы прокариот лишены одного из типов рибосомальной РНК (так называемой 5,8S РНК) и имеют меньшую массу, чем у эукариот. Обычно масса рибосом оценивается так называемой константой седиментации (показателем скорости оседания при центрифугировании). Для рибосом прокариот она равна 70S, а для эукариот - 80S.
Прокариоты обладают громадным (по сравнению с эукариотами) разнообразием обменных процессов. Они способны к фиксации углекислоты, азота, различным вариантам брожения, окислению всевозможных неорганических субстратов (соединений серы, железа, марганца, нитритов, аммиака, водорода и др.). Среди прокариот немало фотосинтезирующих форм, прежде всего это часто встречающиеся в современной биосфере цианобактерии, которые еще называют сине-зелеными водорослями. Они (или родственные им организмы) были широко распространены и в далеком прошлом. Геологические постройки, созданные древними цианобактериями (вероятно, вместе с другими фотосинтезирующими прокариотами) - строматолиты, - нередко обнаруживаются в древнейших слоях земной коры, соответствующих архею и раннему протерозою. Начавшаяся около 4 млрд. лет назад деятельность фотосинтезирующих и других автотрофных прокариот имела несколько важнейших последствий.
 |
| Неглубокие водоемы, образовавшиеся около 4 млрд. лет назад. В них обитали первые живые организмы, близкие к современным цианобактериям. |
Первое связано с изменением атмосферы Земли. Дело в том, что в древности она была практически бескислородной. В результате фотосинтеза молекулярный кислород стал выделяться в атмосферу, но быстро связывался с неокисленными компонентами литосферы - железом и другими металлами. Поэтому, несмотря на наличие постоянного источника свободного кислорода, биосфера оставалась по преимуществу анаэробной. Живые организмы в этот период тоже были представлены в основном анаэробами. В литосфере между тем откладывались полосчатые железные руды (так называемые джеспилиты), в которых окисленное железо чередовалось с недоокисленным. В бескислородных условиях откладывались пириты (руды типа FeS2), которые не могли формироваться при наличии свободного кислорода. Находки подобных ископаемых позволяют установить, что, несмотря на обилие фотосинтетиков, анаэробный период в развитии биосферы длился почти 2 млрд. лет.
Однако около 2 млрд. лет назад содержание кислорода в атмосфере достигло 1% и продолжало повышаться, поскольку к тому времени большая часть находившегося на поверхности железа и других металлов оказалась окисленной. В то же время количество железа и других металлов, поднимающихся из глубин Земли, постепенно уменьшалось. При формировании планеты тяжелые и легкие компоненты оказались перемешаны случайным образом. В дальнейшем в процессе гравитационной дифференцировки металлы постепенно погружались к центру планеты, формируя ее железное ядро, а легкие компоненты - силикаты - поднимались вверх, образуя мантию.
Для анаэробных организмов повышение концентрации кислорода было катастрофой, поскольку кислород - очень агрессивный элемент, он быстро окисляет и разрушает органические соединения. Если в анаэробной биосфере, в толще строматолитов оставались аэробные карманы, откуда накапливающийся в результате фотосинтеза кислород диффундировал в атмосферу, то теперь биосфера, по меткому выражению академика Г.А. Заварзина, "вывернулась наизнанку" - она превратилась в кислородную с немногочисленными бескислородными карманами, где нашли убежище анаэробные микроорганизмы. В новой аэробной атмосфере могли выжить только те немногие прокариоты (оксибактерии), которые еще раньше в кислородных карманах в толще строматолитов приспособились к высокой концентрации кислорода.
Второе важное следствие деятельности автотрофных прокариот - накопление залежей органического вещества. Биотический круговорот веществ в биосфере, состоящей исключительно из прокариот, был очень несовершенен. Биомасса, созданная автотрофными бактериями, подвергалась разложению преимущественно под воздействием абиотических физических и химических процессов во внешней среде. Без сомнения, существенную роль в разложении биомассы, созданной прокариотными автотрофами, играли и гетеротрофные бактерии, однако их возможности были ограничены в силу особенностей организации клеток прокариот. Как известно, прокариоты принципиально не способны к заглатыванию своих жертв. Хищничество у бактерий встречается очень редко и выглядит весьма необычно. Хищная бактерия Bdellovibrio по размерам значительно меньше своих жертв, она проникает через клеточную стенку бактерии и размножается внутри организма несчастной.

Клетки ископаемых прокариотных организмов, близких к цианобактериям, в тонких шлифах архейских осадочных пород (слева и в центре). Справа - фотография ископаемых строматолитов, образованных древними фотосинтезирующими бактериями.
Почему же прокариоты неспособны к заглатыванию пищи? Дело в том, что у них отсутствуют актин и миозин - белки, обеспечивающие подвижность цитоплазмы у эукариот. Благодаря им при захвате пищевых частиц (фагоцитозе) и формировании пищеварительных вакуолей формируются псевдоподии (временные цитоплазмические выросты, служащие для передвижения и захвата пищи). Прокариоты этого делать не могут. Гетеротрофные бактерии выделяют ферменты во внешнюю среду, происходит своего рода "наружное переваривание" (экзоферментация), а низкомолекулярные продукты всасываются через цитоплазматическую мембрану. Все это обусловило низкую скорость разложения биомассы, созданной автотрофными прокариотами. Поэтому на ранних этапах эволюции биосферы огромные массы органического углерода выводились из биологического круговорота, сохранялись в осадке, подвергались химической трансформации, превращаясь в горючие сланцы, нефть и газ, которыми человечество активно пользуется до сих пор.
Усовершенствовать биологический круговорот, ускорить возврат в него углерода и других биогенных элементов могло только появление микроскопических аэробных хищников, которые заглатывали бы бактерий, переваривали их и возвращали в биосферу углерод (желательно в виде СO2), азот (в виде соединений аммония), фосфор и другие биогенные элементы. Такими хищниками стали первые эукариотные организмы.
Хищники
Эукариоты обладают двумя универсальными белками - актином и миозином, обеспечивающими разнообразные типы клеточной подвижности: амебоидную активность, движение органелл внутри клетки, а у высших организмов - мышечные сокращения. Актиново-миозиновая система позволяет образовывать псевдоподии, захватывать ими жертву и формировать пищеварительные вакуоли (даже вирусы проникают в клетку эукариот путем провокации так называемого "эндоцитоза" - клетка принимает их за нечто полезное, "проглатывает", и вирус, оказавшись в цитоплазме, начинает свою разрушительную работу). Приобретение актиново-миозиновой системы позволило эукариотам питаться путем фагоцитоза, активно захватывая крупные пищевые частицы.
Появление таких организмов необычайно ускорило биотический круговорот, поскольку они стали потребителями бактериальной биомассы. Переваривая клетки бактерий, фаготрофные эукариоты быстро возвращали в круговорот веществ элементы, которые до этого могли снова попасть в него только путем медленного разложения. Можно предположить, что появление эукариот повлекло за собой резкое уменьшение "бактериальных ископаемых", то есть отложений органических и неорганических веществ, возникших в результате деятельности бактерий.
Способность эукариот захватывать пищевые частицы подразумевала, что хищник должен быть крупнее жертвы. Действительно, линейные размеры мелких почвенных амеб или жгутиконосцов приблизительно в 10 раз превышают размеры бактерий, которыми они питаются. Таким образом, объем цитоплазмы эукариот примерно в 1000 раз больше, чем у прокариот, что требует и большого числа копий генов, чтобы снабжать цитоплазму продуктами транскрипции. Один из способов решения этой задачи - увеличение числа генофоров, то есть кольцевых молекул ДНК. По этому пути пошли крупные (так называемые "полиплоидные") бактерии и предки эукариот с большим объемом цитоплазмы. Множественные генофоры (первоначально одинаковые) стали зачатками хромосом, в которых постепенно накапливались различия.

Цианобактерии процветают и в современной биосфере (левая фотография). Кое-где они образуют постройки, напоминающие древние строматолиты. На правой фотографии показаны современные строматолиты из залива Шарк-бей на севере Австралии.
При амебоидном движении и питании путем фагоцитоза цитоплазма клетки (особенно периферическая) становится очень подвижной. Генофоры, прикрепленные к поверхностной мембране клетки, оказывались в зоне сильных токов цитоплазмы, поэтому в центральной цитоплазме появился защищенный мембранами участок, где хранились генофоры. Процесс мог происходить по-разному, но один из возможных путей - глубокие впячивания участков цитоплазматической мембраны с прикрепленными к ним генофорами (ведь ядерная оболочка - это часть эндоплазматического ретикулюма эукариотной клетки, который может быть связан с внешней средой).
Первичные эукариоты, таким образом, имели ядро, ограниченное двойной ядерной оболочкой - производным эндоплазматического ретикулюма, но имели еще кольцевое строение генофоров и были лишены специфических ядерных белков - гистонов. Как ни удивительно, подобное строение ядра сохранилось и у некоторых современных эукариот, например у динофлагеллят. У этих простейших ядро окружено двойной ядерной мембраной, но хромосомы содержат кольцевые молекулы ДНК, лишенные гистонов. По-видимому, ядро дино-флагеллят - реликтовая структура, сохранившая строение, характерное для первичных эукариотных организмов.
Симбиотическое происхождение митохондрий и жгутиков
Способность к фаготрофному питанию предопределила возможность появления у эукариот внутриклеточных симбионтов. Прокариоты этого делать не могли - лишенные способности заглатывать кого бы то ни было, они не приобрели внутриклеточных эндосимбионтов. Для эукариот, наоборот, включение в качестве внутриклеточных симбионтов различных прокариотных и эукариотных организмов весьма характерно. Эукариотная клетка возникла в результате симбиоза первичного амебоидного организма с различными прокариотными и эукариотными существами. Данное положение легло в основу так называемой концепции симбиогенеза, которая стала одной из парадигм современной биологии.
 |
 |
| "Хищничество" у современных прокариотных организмов. Вверху показано как "хищная" бактерия Bdellovibrio проникает в кишечную палочку и размножается внутри нее. Внизу - вирус-бактериофаг впрыскивает в бактерию свою ДНК, тогда как его белковая оболочка остается снаружи. Вирусная ДНК обеспечивает синтез новых вирусных частиц. | Благодаря актиново-миозиновой системе эукариотные организмы могут образовывать псевдоподии и фагоцитировать бактерии и другие частицы (вверху). Вирус использует это свойство эукариотных организмов и провоцирует эндоцитоз - поглощение вирусной частицы самой клеткой (внизу). |
Концепция симбиогенеза была сформулирована еще в начале ХХ в. двумя выдающимися российскими биологами - К.С. Мережковским (родным братом известного писателя Д.С. Мережковского) и Ф.С. Фаминицыным. Однако их идеи в то время не были оценены по достоинству и не получили широкого распространения. К идее симбиогенеза биологи вернулись только в последние десятилетия ХХ в., когда накопилось множество данных по структуре клеток эукариотных организмов. Современные положения симбиогенетической концепции разработаны в трудах американского биолога Лины Маргелис и отечественных исследователей А.Л. Тахтаджана и И.М. Мирабдуллаева.
Согласно нынешним представлениям, симбиотическое происхождение имеют такие важные органеллы эукариотной клетки, как митохондрии. Они обеспечивают синтез главного энергетического ресурса любой клетки - АТФ за счет окислительного фосфорилирования, которое возможно только в присутствии кислорода. Не имеют митохондрий лишь некоторые простейшие, обитающие в анаэробных условиях (например, в кишечнике животных или в лишенных кислорода болотных водах). Несомненно, отсутствие у них митохондрий - вторичный признак, связанный с существованием в бескислородных условиях, это подтверждается тем, что в геноме таких простейших найдены некоторые митохондриальные гены.
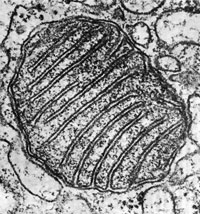 |
| Электронно-микроскопическая фотография митохондрии. |
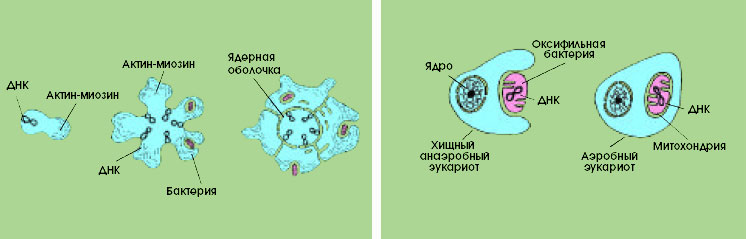 | |
| Схема происхождения эукариотной клетки. Слева - гипотетический прокариотный организм, обладающий актиново-миозиновой системой. В середине - крупный хищный прокариот с множественными генофорами, который 250 питается заглатывая бактерий. Справа - первичный эукариот, у которого за счет глубоких впячиваний поверхностной цитоплазматической мембраны сформировалась ядерная оболочка. | Происхождение митохондрий путем симбиоза хищного анаэробного эукариота и оксифильной бактерии. |
Какие же прокариоты могли быть предками митохондрий? Среди современных прокариот ближе всего к ним пурпурные альфа-протеобактерии (об этом свидетельствуют, в частности, и новые данные молекулярной филогении) - аэробные фотосинтезирующие бактерии, мембрана которых образует глубокие впячивания, похожие на кристы митохондрий. Прародители таких бактерий, вероятно, обитали в кислородных карманах анаэробной биосферы. Вступив в симбиоз с древними амебоидными эукариотами, протеобактерии утратили способность к фотосинтезу, поскольку все необходимые органические вещества они стали получать от хозяина - древнего эукариота, получившего свою выгоду: перестал бояться высоких концентраций кислорода, который утилизировали симбионты.
Первичные аэробные эукариоты, обладающие симбионтами, первоначально тоже заселяли кислородные карманы, но когда через 3 млрд. лет после образования биосферы концентрация кислорода стала нарастать, эукариоты получили возможность широко распространиться в биосфере. В слоях земной коры, относящихся к этому периоду, появляются так называемые акритархи - крупные сферические клетки диаметром 50-60 мкм. Они не могли принадлежать прокариотам, сферические клетки которых не превышают в диаметре нескольких микрон (нитевидные формы могут достигать значительно большей длины). В слоях, возраст которых составляет около 1,7 млрд. лет, найдены стеролы - вещества, синтезируемые в ядре эукариотных организмов. Таким образом, в период от 1 до 2 млрд. лет назад началась адаптивная эволюция эукариот.
Жгутики и реснички эукариотных клеток тоже считаются "потомками" симбиотических прокариот. Многие бактерии (например, вибрионы) имеют жгутики (так называемые "флагеллы") - особые образования, содержащие белок флагеллин. Но ни по строению, ни по химическому составу они не имеют ничего общего со жгутиками и ресничками эукариот. Зато у последних они устроены абсолютно идентично. Жгутики паразитического простейшего - трипаносомы, реснички инфузории-туфельки, сперматозоиды папоротника, реснички трахеи человека устроены одинаково. Внутри жгутика или реснички проходят правильно расположенные микротрубочки, состоящие из особого белка - тубулина (который никогда не встречается у прокариот). Располагаются они следующим образом: в центре проходят две одиночные микротрубочки, а по периферии - 9 дублетов. Микротрубочки жгутика присоединяются к базальному тельцу - кинетосоме, которая состоит из 9 триплетов микротрубочек. Даже если жгутики исчезают, кинетосомы остаются и функционируют в качестве центриолей. Например, все клетки (в том числе и безжгутиковые) многоклеточных животных обладают двумя рядом расположенными центриолями. Именно они организуют веретено деления, благодаря которому достигается правильное распределение хромосом в митозе и мейозе.
Жгутики не отделены от цитоплазмы мембранами, никаких препятствий для перехода белков из цитоплазмы в жгутик нет, поэтому большинство белков жгутика кодируются в ядре клетки. В то же время внутри базального тельца жгутика есть маленькая кольцевая молекула ДНК, которая содержит несколько генов, контролирующих формирование базального тельца. Дело в том, что центриоли (базальные тельца) не возникают в клетке на пустом месте. Перед делением две центриоли расходятся и рядом с каждой из них формируется новая. Таким образом, для синтеза очередного органоида необходима "затравка" в виде старого.
Предполагается, что предками жгутика были бактерии, напоминающие современных спирохет, подвижных бактерий, чьи узкие спирально закрученные клетки быстро движутся, как бы ввинчиваясь в пространство. Правда, сами они никак не могли быть предками жгутиков: в них нет микротрубочек, а тонкое строение совершенно иное. Но это вовсе не означает, что в далеком прошлом не было других спирохетоподобных организмов, которые и стали предком эукариотного жгутика. По-видимому, его прародители сначала были экзосимбионтами, то есть прикреплялись к цитоплазматической мембране примитивного эукариота снаружи. Симбионт использовал для своего питания выделяемые хозяином метаболиты, а взамен благодаря своей локомоторной активности способствовал его быстрому (по сравнению с формированием псевдоподий) перемещению. Именно такое взаимодействие сформировалось между спирохетами и некоторыми крупными простейшими. Симбиотические спирохеты сидят на поверхности жгутиконосца Myxotricha paradoxa (который имеет и обычные жгутики), их движения согласованны, как у настоящих ресничек, а локомоторная активность обеспечивает плавное и постепенное движение жгутиконосца, тогда как собственные жгутики позволяют ему совершать лишь быстрые движения вперед по спирали. Любопытно, что для большего удобства прикрепления спирохет клетка хозяина любезно образует специальные уплотненные "подставки", от которых внутрь цитоплазмы хозяина идут пучки фибрилл, напоминающие корешки настоящих жгутиков и ресничек. Этот пример показывает, что симбиоз между подвижными бактериями и эукариотами может возникать неоднократно.
 | |
| Происхождение эукариотной клетки путем симбиоза сначала с оксифильными бактериями (предками митохондрий), а потом - со спирохетами (предками жгутиков). | Происхождение хлоропластов красных водорослей путем симбиоза с цианобактериями. |
Происхождение эукариотных растений
Первичные эукариоты были одноклеточными животными. Они питались, захватывая и переваривая другие микроскопические организмы. Одним из магистральных направлений их эволюции стало приобретение фотосинтезирующих симбионтов, которые превратились в органеллы, обеспечивавшие синтез органических веществ из углекислого газа и воды за счет энергии солнечного света. Этот путь привел к появлению различных групп эукариотных растений, то есть автотрофных фотосинтезирующих организмов. Они не родственны друг другу и возникли в результате симбиоза хищных протистов (простейших или их колоний) с различными фотосинтезирующими организмами.
В нескольких случаях симбионтами хищных эукариот стали цианобактерии - сине-зеленые водоросли, самая распространенная (по крайней мере в современной биосфере) и, возможно, самая древняя группа фотосинтезирующих прокариот. Их несомненными потомками являются фотосинтезирующие органеллы (хлоропласты) красных водорослей. Они окружены только двумя мембранами, имеют собственную кольцевую ДНК и рибосомы прокариотного типа и содержат типичные для цианобактерий хлорофилл "а" и специфические пигменты цианобактерий - фикобилины. Красные водоросли в настоящее время широко распространены в морях нашей планеты. Они способны существовать на глубинах в несколько сот метров, но живут и в приливно-отливной полосе, а некоторые виды обитают и в пресных водах. Возможно, красные водоросли - самая древняя группа эукариотных растений. Об этом говорит полное отсутствие в их жизненном цикле жгутиковых стадий (даже их сперматозоиды - безжгутиковые), что позволяет предположить, что предки этих водорослей отделились от остальных эукариот еще до приобретения жгутиков.
Впрочем, красные водоросли - не единственная группа, использующая потомков цианобактерий в качестве симбионтов. У одноклеточных жгутиконосцев - глаукофитов (совсем не родственных красным водорослям) фотосинтезирующие органеллы так и называются - цианеллы. Они даже сохраняют характерную для цианобактерий муреиновую оболочку (т.е. механически прочный элемент клеточной стенки). Тем не менее цианеллы - настоящие симбионты, которые не могут жить отдельно от хозяина. Даже их геном - кольцевая ДНК - приблизительно в 10 раз короче, чем у свободноживущих цианобактерий. Это означает, что и в данном случае значительная часть белков цианелл кодируется в ядерном геноме хозяина.
 | |
| Происхождение хлоропластов зеленых водорослей и высших растений путем симбиоза с зелеными бактериями. | Происхождение хлоропластов криптомонад, золотистых и бурых водорослей путем двойного симбиоза: сначала хищный эукариот вступил в симбиоз с золотистыми бактериями, а потом сам стал симбионтом другого хищного организма. |
Хлоропласты зеленых ворослей (хлореллы, хламидомонады, вольвокса и др.) - тоже потомки фотосинтезирующих прокариот. Они окружены двумя мембранами, содержат кольцевую ДНК и собственные рибосомы прокариотного типа. Однако набор хлорофиллов у них совсем другой - это хлорофиллы "a" и "b", а фикобилинов нет. Значит, предками хлоропластов зеленых водорослей не могли быть цианобактерии. Долгое время свободноживущие бактерии с хлорофиллами "a" и "b" не были известны. Лишь в последние два десятилетия были обнаружены представители особой группы прохлорофитов - Prochloron и Prochlorotrix - с таким же набором хлорофиллов. Прохлорон представляет собой крупную шарообразную бактерию, живущую в тунике колониальных асцидий, а прохлоротрикс - нитчатая пресноводная форма. В настоящее время прохлорофиты - реликтовая группа, насчитывающая всего несколько видов, но в далеком прошлом они, вероятно, играли в биосфере значительную роль. Вполне возможно, что древние прохлорофиты участвовали (наряду с цианобактериями) в построении строматолитов. Тогда же они вступили в симбиоз с предками зеленых водорослей. Значение этого союза тем более велико, что потомки зеленых водорослей - высшие растения - унаследовали хлоропласты с двумя мембранами и хлорофиллами "a" и "b". Таким образом, в зеленой иголочке сосны или блестящем листе фикуса сохранились потомки древних прохлорофитов, превратившихся в хлоропласты.
Мир эукариотных растений отнюдь не ограничивается красными и зелеными водорослями. В современной биосфере процветают различные группы организмов с золотисто-бурыми хлоропластами. Одноклеточные и колониальные диатомовые водоросли, клетки которых защищены кремнеземным панцирем, господствуют в Мировом океане, населяют пресные воды и влажную почву. Прибрежная зона моря заселена бурыми водорослями - фукусами, ламинариями и саргассами (последние могут выживать и в открытом океане - вспомните Саргассово море). Среди бурых водорослей встречаются настоящие гиганты. Например, у тихоокеанского побережья Южной Америки обитает самый крупный растительный организм планеты - макроцистис, достигающий 150 м в длину. В планктоне морских и пресных вод распространены фотосинтезирующие жгутиконосцы - золотистые водоросли и криптомонады.
Хлоропласты золотистых, диатомовых и бурых водорослей содержат хлорофиллы "а" и "с" и почему-то окружены 4 мембранами. Их происхождение помогло понять строение криптомонад - небольшой группы жгутиконосцев, хлоропласты которых тоже имеют хлорофиллы "а" и "с", окружены 4 мембранами, причем между второй и третьей имеется маленькое эукариотное ядро - нуклеоморф, а внутри пространства, ограниченного последней, четвертой мембраной находится кольцевая ДНК. Такое строение позволяет предполагать, что хлоропласты криптомонад возникли в результате двойного симбиоза. Сначала некий хищный протист приобрел в качестве симбионта золотистую бактерию с хлорофиллами "а" и "с", а потом сам стал симбионтом криптомонады. В хлоропластах бурых, диатомовых и золотистых водорослей нуклеоморфа уже нет, хотя они по-прежнему окружены 4 мебранами, что говорит о более глубокой интеграции симбионта и хозяина.
Хлоропласты приобретены различными группами эукариотных растений независимо друг от друга, и предками хлоропластов были разные свободноживущие организмы: в одних случаях ими были бактерии (зеленые или сине-зеленые), а в других - эукариотные простейшие.
Вместо заключения
Эукариотные организмы - простейшие, различные группы растений, грибы и многоклеточные животные - доминируют в современной биосфере. Однако все они несут в своих клетках симбионтов - потомков древних свободноживущих бактерий. Только благодаря им эукариотные организмы способны жить в кислородной атмосфере и использовать энергию солнечного света для синтеза органических веществ. Так может быть, на самом деле эукариоты вовсе не доминируют в биосфере, а им это только кажется? Сторонник теории симбиогенеза американский биолог Л. Томас как-то сказал: "Обычно на митохондрии смотрят как на порабощенные существа, взятые в плен, чтобы снабжать АТФ клетки, и не способные дышать самостоятельно. С этой рабовладельческой точки зрения смотрят на дело и солидные биологи, которые сами - все эукариоты. Но с точки зрения самих митохондрий они - существа, которые давным-давно нашли для себя лучшее из возможных пристанищ, где можно жить, затрачивая минимум усилий и подвергаясь наименьшему риску".
Мы не должны забывать, что в каждой клеточке нашего тела живут крошечные потомки древних оксифильных бактерий, которые прокрались в организм наших далеких предков 2 млрд. лет назад и продолжают существовать в нас, сохраняя собственные гены и свою особую биохимию. Другая цитата Л. Томаса: "Вот они движутся в моей цитоплазме, дышат для нужд моего тела, но они - чужие. Мне жаль, что я не могу познакомиться с моими митохондриями поближе. Когда я сосредоточусь, я могу представить, что ощущаю их; не то чтобы я чувствовал, как они извиваются, но время от времени я воспринимаю какой-то трепет. Я не могу отделаться от мысли, что если бы я знал больше о том, как они достигают такой гармонии, я бы по-другому понимал музыку".
ОБ АВТОРЕ:
Малахов Владимир Васильевич, профессор кафедры зоологии беспозвоночных МГУ, автор 190 публикаций. Область научных интересов - сравнительная анатомия и эмбриология беспозвоночных. Действительный член Российской Академии естественных наук, член-корреспондент Российской Академии наук.