(учебник по теории эволюции)
Эволюция жизни
Содержание
Предисловие
ЧАСТЬ I. ИЗ ИСТОРИИ ЭВОЛЮЦИОНИСТИКИ: ДАРВИНИЗМ И ЛАМАРКИЗМ
Глава 1. Ранние этапы развития эволюционных представлений
Глава 2. Теория эволюции Ламарка
Глава 3. Теория эволюции Дарвина
Глава 4. Неоламаркизм
Глава 5. Формирование современной теории эволюции
ЧАСТЬ II. МИКРОЭВОЛЮЦИЯ И ВИДООБРАЗОВАНИЕ
Глава 1. Элементарные предпосылки эволюционного процесса: формы изменчивости организмов
Глава 2. Биологический вид
Глава 3. Факторы эволюции
Глава 4. Видообразование и межвидовые отношения
Глава 5. Развитие приспособлений
ЧАСТЬ III. ИСТОРИЧЕСКОЕ РАЗВИТИЕ (ФИЛОГЕНЕЗ) ОРГАНИЗМОВ
Глава 1. Развитие жизни в криптозое и загадка нижнего кембрия
Глава 2. Жизнь в палеозойской эре
Глава 3. Мезозойская эра - век рептилий
Глава 4. Кайнозой - век млекопитающий и птиц
Глава 5. Происхождение и эволюция человека
ЧАСТЬ IV. МАКРОЭВОЛЮЦИЯ
Глава 1. Индивидуальное развитие и эволюция
Глава 2. Соотношения макроэволюции и микроэволюции
Глава 3. Направления макроэволюции и проблема ее направленности
Глава 4. Эволюционный прогресс
Глава 5. Темпы макроэволюции
Cписок рекомендуемой литературы
В 1979-1981 гг. были опубликованы книги автора "Основы теории эволюции" и "Развитие жизни на Земле", получившие благоприятные отзывы и рецензии. Тогда и возник замысел объединить их в одну, коренным образом переработав. Результатом этой работы стала предлагаемая книга, в которую включены новые данные, накопившиеся за последние десятилетия в разных областях стремительно развивающейся науки. Многие разделы полностью написаны заново, в частности четвертая часть книги, посвященная проблемам макроэволюции.
Книга предназначена для студентов биологических факультетов различных университетов и институтов, изучающих проблемы эволюционной теории и филогенеза организмов.
Характер издания, к сожалению, не позволяет привести полный список использованной литературы, включающий около 800 наименований. Некоторые ссылки на литературные источники сделаны в подстрочных примечаниях, а в конце книги приведен краткий список основной литературы.
Автору приятно выразить свою искреннюю признательность профессору, доктору биологических наук Н.М.Черновой и Л.Г.Прилежаевой за прочтение рукописи, доброжелательную критику и ценные советы.
Хотя теория эволюции исторически сложилась на основе анализа данных различных биологических наук (палеонтологии, сравнительной анатомии, эмбриологии, генетики, систематики, экологии, биогеографии, со второй половины XX в. молекулярной биологии и др.), в современной биологии выделилась область, специально изучающая механизмы и общие закономерности эволюционного процесса и представляющая собой, в сущности, особую биологическую науку. Эту дисциплину называют по-разному: эволюционное учение, дарвинизм (подчеркивая особое значение трудов Ч.Дарвина в создании эволюционной теории), эволюционистика. Последнее название представляется нам наиболее удачным, ближе всего соответствуя по своему звучанию наименованиям других биологических наук. Эволюционистику можно определить как биологическую науку, изучающую причины, движущие силы, механизмы и закономерности эволюции организмов.
Необходимо подчеркнуть, что эволюция организмов представляет собой исторические преобразования всех уровней организации биологических систем - от молекулярного до экосистемного, включая и биосферу в целом. Взаимосвязи разных уровней иерархической организации биологических систем имеют сложный и неоднозначный характер - даже в пределах единого целостного организма, не говоря уже о надорганизменных системах (популяции, виды, биоценозы). Эволюционные преобразования разных уровней биологической организации происходят в определенной степени независимо друг от друга. В связи с этим в современной эволюционистике сформировались три основных направления исследований эволюционного процесса, связанных с разными уровнями организации биологических систем и использующих методы разных биологических наук: 1) молекулярно-биологическое (анализ молекулярной эволюции, т.е. процессов эволюционных преобразований биологических макромолекул, в первую очередь нуклеиновых кислот и белков, методами молекулярной биологии); 2) генетико-экологическое (исследования микроэволюции, т.е. преобразований генофондов популяций, и процессов видообразования, а также эволюции биологических макросистем - биоценозов и биосферы в целом - методами популяционной генетики, экологии, систематики, фенетики); 3) эволюционно-морфологическое (изучение макроэволюции -эволюционных перестроек целостных организмов и их онтогенезов методами палеонтологии, сравнительной анатомии и эмбриологии).
Основные задачи эволюционистики менялись на разных этапах ее развития. В очень упрощенной форме можно сказать, что в XIX в. важнейшей задачей было доказательство реальности эволюции органического мира; в XX в. на первый план вышло причинное объяснение механизмов и эмпирически установленных закономерностей эволюционного процесса. При этом в первой половине XX в. внимание исследователей концентрировалось главным образом на процессах микроэволюции, тогда как во второй все более развивались исследования молекулярной эволюции; на очереди анализ макроэволюции и новая интеграция всех областей эволюционистики.
Всякая претендующая на полноту и последовательность эволюционная теория должна решить ряд принципиальных проблем, среди которых: 1) общие причины и движущие силы эволюции организмов; 2) механизмы развития приспособлений (адаптации) организмов к условиям их обитания и изменениям этих условий; 3) причины и механизмы возникновения поразительного разнообразия форм организмов, а также причины сходств и различий разных видов и их групп; 4) причины эволюционного прогресса - нарастающего усложнения и совершенствования организации живых существ в ходе эволюции - при одновременном сохранении более примитивных и просто устроенных видов.
Для анализа эволюционного процесса часто используются понятия "эволюционные факторы", или "факторы эволюции", и "эволюционные механизмы". Эволюционные факторы - это любые явления или процессы, оказывающие какое-либо воздействие на эволюцию организмов (например, изменчивость организмов - наследственная и ненаследственная, гибридизация, изоляция популяций и др.). Эволюционные механизмы представляют собой способы действия и взаимодействия эволюционных факторов, которые определяют возникновение определенных эволюционных событий и закономерностей (например, механизмы видообразования, формирования адаптации, эволюционного прогресса и др.).
Структурными элементами эволюционного процесса являются микроэволюционные события - изменения генофондов популяций, факторы и механизмы которых рассматриваются во второй части книги. Третья часть посвящена обзору исторического развития (филогенеза) органического мира и является основой для анализа направлений и закономерностей макроэволюции в завершающей, четвертой части книги. В первой же части мы кратко рассматриваем некоторые основные вехи в истории эволюционистики с особым вниманием к тем концепциям, которые оказали наибольшее влияние на последующее развитие науки. В первую очередь это относится к теориям Ж. Б.Ламарка и Ч.Дарвина.
Из философов Античности наибольшей известностью и авторитетом среди натуралистов в последующие эпохи (в частности, в период Средневековья) пользовался Аристотель (384-322 до н.э.) Аристотель не поддерживал, во всяком случае в достаточно ясной форме, идею изменяемости окружающего мира. Однако многие его обобщения, сами по себе укладывавшиеся в общую картину неизменности мира, сыграли в дальнейшем важную роль в развитии эволюционных представлений. Таковы мысли Аристотеля о единстве плана строения высших животных (сходство строения соответствующих органов у разных видов было названо Аристотелем "аналогией"), о постепенном усложнении ("градации") строения в ряду организмов, о многообразии форм причинности (Аристотель выделял 4 ряда причин: материальную, формальную, производящую, или движущую, и целевую).
Эпоха Поздней античности и особенно последовавшая за ней эпоха Средневековья стали временем затянувшегося почти на полторы тысячи лет застоя в развитии естественно-исторических представлений. Господствовавшие догматические формы религиозного мировоззрения не допускали идеи изменяемости мира. Соответствующие представления античных философов были преданы забвению. Возможности для развития эволюционных идей появились лишь после эпохи Возрождения (XV-XVI вв.), когда европейская наука сделала значительные шаги вперед в познании окружающего мира.
Креационизм и трансформизм
Постепенно были накоплены многочисленные данные, говорившие об удивительном разнообразии форм организмов. Эти данные нуждались в систематизации. Важный вклад в этой области был сделан знаменитым шведским естествоиспытателем К.Линнеем (1707-1778), которого справедливо называют создателем научной систематики организмов. Следует отметить, что Линней последовательно придерживался точки зрения о неизменности видов, созданных Творцом.
В XVII-XVIII вв. наряду с господствовавшим мировоззрением, основанным на религиозных догмах о неизменности созданного Творцом мира и получившим название "креационизм" (от лат. creatio - созидание, порождение), постепенно начали вновь формироваться представления об изменяемости мира и, в частности, о возможности исторических изменений видов организмов. Эти представления именовались "трансформизмом" (от лат. transformatio - преобразование). Наиболее яркими представителями трансформизма были естествоиспытатели и философы Р. Гук (1635-1703), Ж.Ламеттри (1709-1751), Ж.Бюффон (1707-1788), Д Дидро (1713-1784), Эразм Дарвин (1731-1802), И.В.Гёте (1749-1832), Э.Жоффруа Сент-Илер (1772-1844).
У трансформистов еще не сложилось целостной концепции эволюции органического мира; их взгляды были во многом эклектичными и непоследовательными, объединявшими материалистические и идеалистические представления. Общим для всех трансформистов былочпризнание изменяемости видов организмов под воздействием окружающей среды, к которым организмы приспосабливаются благодаря заложенной в них изначально способности целесообразно реагировать на внешние воздействия, а приобретенные этим путем изменения наследуются (так называемое "наследование приобретенных признаков"). При этом изменения видов не столько доказывались, сколько постулировались трансформистами, что делало слабыми их позиции в дискуссиях со сторонниками креационизма. Не случайно в 1830 г. на знаменитом диспуте во Французской академии наук между Э.Жоффруа Сент-Илером и научным лидером креационизма Ж.Кювье первый потерпел сокрушительное поражение, не сумев доказать реальности исторических изменений организмов.
Ж.Кювье (1769-1832) обладал огромными знаниями в области сравнительной анатомии и палеонтологии, будучи, в сущности, создателем последней науки, и легко опроверг аргументы Э.Жоффруа, пытавшегося доказать несуществующее "единство плана строения" у позвоночных и головоногих моллюсков. Изменяемость же видов организмов Кювье отрицал на том основании, что мумифицированные животные из древнеегипетских захоронений принадлежат к тем же видам, что и современные. Эти аргументы казались тогда неопровержимыми, поскольку еще не были известны реальные масштабы геологического времени, и время постройки пирамид считали одной из самых ранних эпох в истории Земли.
Честь создания первых эволюционных теорий принадлежит великим естествоиспытателям XIX в. Ж.Б.Ламарку (1744-1829) и Ч.Дарвину (1809-1882). Эти две теории почти во всем противоположны: и в своей общей конструкции, и в характере доказательств, и в основных выводах о причинах и механизмах эволюции, и в своей исторической судьбе. Выход в свет гениального труда Ч.Дарвина "Происхождение видов" (1859) справедливо рассматривается как начало новой эпохи в развитии естественной истории, или биологии, в современном понимании. Дарвинизм стал основой эволюционистики XX в. Напротив, теория Ламарка не получила признания у современников, была надолго забыта, но позднее вновь привлекла внимание ученых, и некоторые ее положения с Удивительным постоянством продолжают воскресать в концепциях различных эволюционистов вплоть до нашего времени.
Таким образом, обе классические теории XIX в. продолжают сохранять актуальность, хотя и по-разному. Поэтому нам необходимо хотя бы вкратце остановиться на основных положениях этих теорий, чтобы разобраться в особенностях их исторической судьбы и месте в современной науке.
Изменчивость организмов
При этом, конечно, нельзя сказать, что Ламарк совсем не опирался на факты: именно наблюдения над некоторыми проявлениями изменчивости организмов были стимулом для переосмысления господствовавших креационистских представлений. Среди этих проявлений изменчивости наиболее очевидными были приспособительные изменения организмов, подвергавшихся воздействию новых условий (например, развитие из одинаковых семян при их выращивании в разных условиях растений различного облика; усиление мускулов у человека и животных при их усиленном упражнении и ослабление этих мускулов при отсутствии соответствующей физической нагрузки и т.п.). Ламарк обратил внимание и на существование разновидностей, выглядящих как промежуточные формы между разными видами, и на изменения организмов в результате процессов одомашнивания, и на отличия ископаемых форм организмов от современных.
Общим выводом Ламарка из этих наблюдений было признание исторической изменяемости, трансформации организмов во времени, т. е. их эволюции. Однако этот вывод уже не был оригинальным: исторические преобразования видов организмов под влиянием изменений внешней среды признавали, как уже было отмечено, все трансформисты.
Учение о градации
Своеобразие концепции Ламарка придало объединение идеи изменяемости органического мира с представлениями о градации - постепенном повышении уровня организации от самых простых до наиболее сложных и совершенных организмов. (Идея градации, понимаемой как последовательный ряд постепенно усложняющихся форм организмов, восходит к Аристотелю. Эти представления о "лестнице существ", образующих непрерывную цепь от неорганических тел до человека, развивал до Ламарка Ш.Боннэ).
Разнообразие видов живых существ, по Ламарку, не является просто хаосом всевозможных форм - в этом разнообразии можно усмотреть определенный порядок, как бы ступени последовательного и неуклонного повышения уровня организации. Из этого Ламарк сделал важнейший вывод, что изменения организмов имеют не случайный, а закономерный, направленный характер: развитие органического мира идет в направлении постепенного совершенствования и усложнения организации. На этом пути жизнь возникла из неживой материи путем самозарождения, и после длительной эволюции организмов появился человек, произошедший от "четвероруких", т.е. от приматов.
Движущей силой градации Ламарк считал "стремление природы к прогрессу", которое изначально присуще всем живым существам, будучи вложено в них Творцом, т.е. Богом. Ламарковское объяснение прогрессивной эволюции, очевидно, является телеологическим (от греч. телео - цель), поскольку оно приписывает организмам стремление к совершенствованию, т.е. к определенной цели (любопытно, что в другом месте своей книги сам же Ламарк утверждает, что приписывать природе какую-нибудь цель есть несомненное заблуждение). С другой стороны, прогрессивное развитие живой природы, по Ламарку, представляет собой процесс саморазвития - автогенез. В осуществлении этого процесса (градации) организмы совершенно независимы от внешнего мира, от окружающей среды. Хотя Ламарк утверждал, что природа развивается "сама", без непосредственного участия Творца, но именно Высший Творец, по Ламарку, создал "материю и природу" и тем самым косвенным образом является создателем всех продуктов саморазвивающейся природы. (Философская концепция, приписывающая Богу роль "первопричины" или "перводвигателя", "заводящего часы" мироздания, после чего Вселенная развивается сама в соответствии с законами, заложенными Творцом, называется Деизмом). Идеалистичность концепций Ламарка вполне очевидна; в различных формах неоламаркизма (см. ниже) она обычно так или иначе затушевана, но сущность их всегда остается той же.
Влияние на организмы внешних условий
Впервые признав прогрессивную эволюцию закономерной и придав ей телеологическое истолкование, Ламарк должен был далее дать ответ на остальные вопросы, стоявшие перед эволюционной теорией, - о приспособлениях, разнообразии, сходствах и различиях разных видов, а также о том, почему же при наличии у всех организмов изначального стремления к прогрессу, к градации в природе существуют одновременно сложные, высокоразвитые и гораздо более простые, примитивные организмы.
Ответ на все эти вопросы Ламарк видел во влиянии на организмы внешних условий, которое нарушает правильность градаций. "Нарастающее усложнение организации подвергается то здесь, то там на протяжении общего ряда животных отклонениям, вызываемым влиянием условий места обитания и усвоенных привычек". Градация, так сказать, "в чистом виде" проявляется при неизменности, стабильности внешней среды; всякое изменение условий существования понуждает организмы приспосабливаться к новой обстановке, чтобы не погибнуть. Этим нарушается равномерное и неуклонное изменение организмов на пути прогресса, и различные эволюционные линии уклоняются в сторону, задерживаются на примитивных уровнях организации. Так Ламарк объяснял одновременное существование на Земле высокоорганизованных и простых групп, а также разнообразие форм животных и растений. Любопытно, что Ламарк в отличие от Э.Жоффруа Сент-Илера отвергал возможность "естественного", без истребления человеком, вымирания видов организмов. По его мнению, все виды древних животных были предками тех или иных современных видов, при всем их несходстве.
Вторая часть теории Ламарка - об изменениях организмов под воздействием изменяющихся внешних условий - в позднейшее время получила значительно большую известность, чем первая (учение о градации). Чтобы объяснить сущность приспособления организмов к различным условиям среды, Ламарк постулировал, что организмы изначально обладают способностью целесообразно реагировать на любые изменения внешних условий. Сущность этой способности Ламарк оставляет без внимания, рассматривая ее просто как свойство живых существ - такого же рода, как их "стремление к прогрессу".
Изменения животных и растений под воздействием внешних условий, по Ламарку, происходят по-разному. Растения воспринимают изменения условий, так сказать, непосредственно - через свой обмен веществ с внешней средой (с усваиваемыми минеральными соединениями, водой, газами и светом). Согласно Ламарку, если семя какого-либо лугового растения, растущего в низинах, будет случайно занесено на сухую каменистую возвышенность, то, приспосабливаясь к новым условиям, потомство этого семени превратится в новую разновидность. У лютика Ranunculus aquatilis, растущего в воде, подводные листья рассечены на тонкие волосовидные доли, а листья, находящиеся на поверхности воды, широкие и округлые, разделенные на простые лопасти (рис. 1). У родственного вида Ranunculus hederaceus, растущего на влажной почве, листья не разделены на тонкие доли. Ламарк считал, что при попадании семян первого вида не в воду, а на влажную почву из них вырастают растения второго вида. Однако в действительности этого не бывает.

В этом и других подобных примерах Ламарк принимает модификационную ненаследственную изменчивость организмов, представляющую собой реакцию данного индивида на различные условия внешней среды, за наследственные изменения. В действительности такие модификационные изменения, как таковые, не наследуются (подробнее см. ниже). Ламарк понимал, что восприятие животными изменений внешних условий не может быть столь простым, как он предполагал это для растений. Поэтому для животных Ламарк разработал более сложный механизм преобразований: 1) всякая значительная перемена во внешних условиях вызывает изменение в потребностях животных; 2) это влечет за собой новые действия животных и возникновение новых "привычек"; 3) в результате животные начинают чаще употреблять органы, которыми они раньше мало пользовались; эти органы значительно развиваются и увеличиваются, а если требуются новые органы, то они под влиянием потребностей возникают "усилиями внутреннего чувства". С обоснованием этого механизма эволюционных изменений животных под влиянием изменений внешних условий связана формулировка Ламарком двух так называемых законов:
I. Во всяком животном, не достигшем предела своего развития, более частое и постоянное употребление какого-либо органа приводит к усиленному развитию последнего, тогда как постоянное неупотребление органа ослабляет его и в конце концов вызывает его исчезновение.
II. Все, что организмы приобретают под влиянием преобладающего употребления или утрачивают под влиянием постоянного неупотребления каких-либо органов, в дальнейшем сохраняется в потомстве, если только приобретенные изменения являются общими для обеих родительских особей.
Как примеры, иллюстрирующие эти положения, Ламарк называл утрату способности к полету у домашних птиц, утрату зубов у китов (вследствие привычки глотать пищу не пережевывая), удлинение шеи и передних конечностей у жирафов (в результате постоянного вытягивания этих органов при срывании высоко растущих листьев), удлинение шеи у водоплавающих птиц (из-за постоянного ее вытягивания при извлечении добычи из-под воды) и т. п.
Странным образом именно эти "законы" Ламарка чаще всего вспоминают, говоря о его теории. В действительности эти обобщения занимают в теории Ламарка второстепенное положение и, более того, не являются, в сущности, его оригинальными идеями: представления о влиянии употребления и неупотребления органов на степень их развития и о наследовании признаков, приобретенных организмами таким путем, разделялись всеми трансформистами и были, так сказать, их общим достоянием.
Первый "закон" Ламарка описывает модификационную изменчивость животных, в принципе сходную с модификационной изменчивостью растений; второй - утверждает наследование приобретенных этим путем признаков взрослых животных. Так же, как и для растений, подобные особенности строения животных в действительности ненаследственны и не могут служить основой для наследственных преобразований.
Если первую часть теории Ламарка (учение о градациях) мы охарактеризовали как автогенетическую, то вторая ее часть представляет собой эктогенетическую концепцию. Согласно этой концепции, внешняя среда активно изменяется, а организм пассивно следует за ее изменениями, приспосабливаясь к ним (эктогенез). Хотя эктогенез и автогенез выглядят полностью противоположными друг другу принципами, у них есть общее, заключающееся в противопоставлении, разрыве организма и внешней среды: в одном случае организм развивается независимо от среды, стремясь к прогрессу, в другом - среда односторонне влияет на организм, изменяя его. И то и другое представляет собой метафизическое понимание взаимоотношений организма и среды, поскольку эти компоненты рассматриваются не в их диалектическом взаимодействии, а как существующие рядом, совместно, в отрыве друг от друга.
Метафизичность концепций Ламарка проявляется также в отсутствии каких бы то ни было объяснений как "стремления организмов к прогрессу", так и свойства организмов целесообразно реагировать на внешние воздействия изменениями, которые Ламарк считал наследственными. И то и другое для Ламарка - "законы природы", изначальные свойства организмов, отличающие живое от неживого.
Отметим также, что для теории Ламарка (как и для взглядов других трансформистов) характерна концентрация внимания на отдельном организме, рассматриваемом вне его связей с другими особями того же вида, т.е., говоря современным языком, отсутствие популяционного подхода, непонимание эволюционной роли биологического вида и составляющих его популяций.
Подводя итоги, следует прежде всего еще раз отдать должное Ламарку как создателю первой последовательной эволюционной теории. Как справедливо отметил К.М.Завадский, особой заслугой Ламарка является то, что он впервые выдвинул эволюционный прогресс как одну из основных закономерностей эволюции организмов. Подчеркнем еще раз, что основные положения теории Ламарка были не столько выведены и доказаны исходя из научных фактов, сколько постулированы, так что теория в целом представляет собой, в сущности, умозрительную логическую схему. Метод доказательства, использованный Ламарком, Л.Я.Бляхер удачно назвал методом "подбора непротиворечащих примеров", которые при возможности другой их интерпретации сами по себе, разумеется, не могут служить доказательствами. Ламарк не доказывал эволюцию организмов, а постулировал ее.
Объяснение Ламарком сущности и движущих сил эволюционного процесса телеологично, метафизично и идеалистично. Теория Ламарка не была принята большинством ученых его времени; ее слабые стороны, противоречивость и шаткость аргументов были слишком очевидны, чтобы эта теория смогла преодолеть господствовавшие креационистские взгляды.
Как первая последовательная и цельная эволюционная концепция, теория Ламарка была прогрессивной для своего времени. Однако возрождение метафизических и телеологических взглядов Ламарка на сущность эволюционного процесса в позднейшее время различными неоламаркистскими концепциями является уже шагом назад в развитии науки. О характере этих неоламаркистских концепций и причинах, вызывающих их возникновение, мы будем говорить ниже, после рассмотрения теории Ч.Дарвина, заложившей основу современных эволюционных взглядов.
Теория Дарвина противоположна теории Ламарка не только по своим последовательно материалистическим выводам, но и по всему своему строю. Она представляет собой замечательный образец научного исследования, основываясь на огромном количестве достоверных научных фактов, анализ которых ведет Дарвина к стройной системе соразмерных выводов. Данные для обоснования своей теории Дарвин собирал в течение многих лет. Первый очерк теории был написан уже в 1842 г., но (поразительный пример научной осторожности и добросовестности!) не был опубликован в течение многих лет, на протяжении которых Дарвин продолжал собирать и анализировать новые данные. Великий труд Дарвина "Происхождение видов путем естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь" вышел в свет лишь в 1859 г.
Известно, что стимулом, ускорившим опубликование Дарвином его работы, был труд А.Уоллеса (1823-1913), независимо пришедшего к близким эволюционным выводам. Обе работы были совместно доложены в 1858 г. на заседании Линнеевского общества в Лондоне, и Уоллес, ознакомившись с трудом Дарвина, полностью признал его приоритет. Дарвин анализировал эволюционный процесс гораздо шире и глубже, чем Уоллес, и, отдавая дань должного уважения последнему, мы с полным основанием называем автором теории естественного отбора Ч.Дарвина.
Мы, разумеется, не можем на страницах этой книги дать полное изложение грандиозного труда Дарвина и ограничимся кратким обзором основных положений его теории, необходимым для уяснения ее соотношений с современной теорией эволюции.
Изменчивость организмов в одомашненном состоянии
Прежде всего Дарвин собрал многочисленные доказательства изменяемости видов животных и растений. Ко времени Дарвина практикой селекционеров были созданы многочисленные породы различных домашних животных и сорта сельскохозяйственных растений. Поскольку работа селекционеров, ведущая к изменению породных и сортовых качеств организмов, была сознательной и целенаправленной и было очевидно, что по крайней мере многие из пород домашних животных созданы этой деятельностью в относительно недавнее время, Дарвин обратился к изучению изменчивости организмов в одомашненном состоянии.
Прежде всего важен был сам факт изменений животных и растений под влиянием одомашнивания и селекции, что, собственно говоря, уже является доказательством изменяемости видов организмов.
По мнению Дарвина, стимулом для возникновения этих изменений является воздействие на организмы новых условий, которому они подвергаются в руках человека. При этом Дарвин подчеркнул, что природа организма в явлениях изменчивости важнее природы условий, поскольку одинаковые условия нередко приводят к разным изменениям у разных особей, а сходные изменения последних могут возникнуть при совершенно разных условиях. В связи с этим Дарвин выделил две основные формы изменчивости организмов под влиянием изменения условий среды: неопределенную и определенную.
Изменения могут быть признаны определенными, если все или почти все потомство особей, подвергшихся известным условиям, изменяется одинаковым образом (так возникает ряд неглубоких изменений: рост зависит от количества пиши, толщина кожи и волосистость - от климата и т. п.).
Под неопределенной изменчивостью Дарвин понимал те бесконечно разнообразные слабые различия, которые отличают друг от друга особей одного вида и которые не могли быть унаследованы ни от родителей, ни от более отдаленных предков. Дарвин заключает, что неопределенная изменчивость является гораздо более распространенным результатом изменения условий, чем определенная, и сыграла более важную роль в образовании пород домашних животных. В этом случае изменения внешних условий играют роль стимула, усиливающего неопределенную изменчивость, но никак не влияющего на ее специфику, т. е. на качество изменений.
Организм, изменившийся в каком-либо направлении, передает потомству тенденцию изменяться далее в том же направлении при наличии условий, вызвавших это изменение. В этом заключается так называемая длящаяся изменчивость, которая играет важную роль в эволюционных преобразованиях.
Наконец, Дарвин обратил внимание на существование у организмов определенных соотношений (корреляции) между различными структурами, при изменении одной из которых закономерно изменяется и другая - соотносительная, или коррелятивная, изменчивость. Примерами таких корреляций являются, по Дарвину, глухота белых котов, имеющих голубые глаза; ядовитость для белых овец и свиней некоторых растений, безвредных для черных особей тех же пород, и т.п.
Искусственный отбор
Так как основной формой изменчивости, по Дарвину, является неопределенная, очевидно, что признания наследственной изменчивости организмов было еще недостаточно для объяснения процесса выведения новых пород животных или сортов сельскохозяйственных растений. Необходимо было указать еще силу, которая на основе незначительных различий особей формирует устойчивые и важные породные признаки. Ответ на этот вопрос Дарвин нашел в практике селекционеров, которые производят искусственный отбор на племя только тех особей, которые обладают интересующими человека признаками. В результате такого отбора от поколения к поколению эти признаки становятся все более ярко выраженными. Отбор представляет собой творческую силу, преобразующую частные различия отдельных особей в признаки, характерные для данной породы или сорта.
Если в XIX в. селекционеры вели уже методический и целенаправленный отбор, то на более ранних этапах развития человеческого общества, когда люди только начинали одомашнивать полезных животных и возделывать некоторые съедобные растения, первобытные скотоводы и земледельцы вели бессознательный искусственный отбор, оставляя на племя тех животных, которые лучше уживались с человеком и лучше служили его целям. Так формировались первые породы домашних животных.
Дарвин указал условия, благоприятствующие проведению искусственного отбора: а) высокая степень изменчивости организмов; б) большое число особей, подвергаемых отбору; в) искусство селекционера; г) устранение случайных скрещиваний среди подвергаемых отбору особей; д) достаточно высокая ценность данных животных или растений для человека.
Необходимо отметить, что, подчеркивая важнейшую эволюционную роль отбора, основывающегося на неопределенной изменчивости, Дарвин допускал также возможность преобразований на базе определенной изменчивости и действия упражнения и неупражнения органов, объясняя именно этими факторами относительно меньший вес костей крыла и больший вес костей ног у домашних уток по сравнению с дикими или большее развитие вымени у коров и коз в тех странах, где этих животных используют для получения не только мяса, но и молока.
Наконец, анализируя породы домашних животных, Дарвин пришел к выводу, что все породы данного вида (например, около 150 пород домашних голубей), как ни велики различия между ними, происходят от одного (или немногих) видов диких предков. Следовательно, путем отбора человек значительно увеличивает разнообразие форм организмов.
Если искусственный отбор был основной силой, используя которую человек смог за относительно короткие сроки создать многочисленные породы домашних животных и сорта растений, существенно отличающиеся от своих диких предков, логично предположить, что подобные процессы могут обусловливать эволюционные преобразования также и в природе.
Изменчивость организмов в природе
Дарвин собрал многочисленные данные, свидетельствующие о том, что изменчивость самых различных видов организмов в природе очень велика, а ее формы принципиально сходны с формами изменчивости домашних животных и растений.
Разнообразные и колеблющиеся различия между особями одного вида образуют как бы плавный переход к более устойчивым различиям между разновидностями этого вида; в свою очередь, последние столь же постепенно переходят в более четкие различия еще более крупных группировок -- подвидов, а различия между подвидами - во вполне определенные межвидовые различия. Таким образом, индивидуальная изменчивость плавно переходит в групповые различия. Из этого Дарвин сделал вывод, что индивидуальные различия особей представляют собой основу для возникновения разновидностей. Разновидности при накоплении различий между ними превращаются в подвиды, а те, в свою очередь, - в отдельные виды. Следовательно, ясно выраженная разновидность может рассматриваться как первый шаг к обособлению нового вида (разновидность - "зачинающийся вид").
Дарвин считал, что между видом и разновидностью нет качественной разницы - это лишь разные этапы постепенного накопления различий между группами особей разного масштаба.
Большей изменчивостью характеризуются более широко распространенные виды, обитающие в более разнообразных условиях. В природе так же, как и в одомашненном состоянии, основной формой изменчивости организмов является неопределенная, служащая универсальным материалом для процесса видообразования.
Если породы домашних животных и сорта растений, выведенные человеком с помощью искусственного отбора, целесообразно приспособлены к потребностям человека, то любые виды организмов в природе приспособлены к жизни в определенных условиях внешней среды; особенности строения и функций организмов способствуют их выживанию.
Здесь необходимо подчеркнуть, что Дарвин впервые поставил в центре внимания эволюционной теории не отдельные организмы (как это было характерно для его предшественников-трансформистов, включая Ламарка), а биологические виды, т.е., говоря современным языком, популяции организмов. Только популяционный подход позволяет правильно оценить масштабы и формы изменчивости организмов и прийти к пониманию механизма естественного отбора.
Борьба за существование и естественный отбор
Сопоставляя все собранные сведения об изменчивости организмов в диком и прирученном состоянии и о роли искусственного отбора для выведения пород и сортов одомашненных животных и растений, Дарвин подошел к открытию той творческой силы, которая движет и направляет эволюционный процесс в природе, - естественного отбора. "Так как рождается гораздо более особей каждого вида, чем сколько их может выжить, и так как, следовательно, постоянно возникает борьба за существование, то из этого вытекает, что всякое существо, которое в сложных и нередко меняющихся условиях его жизни, хотя незначительно, изменится в направлении, для него выгодном, будет иметь более шансов выжить и, таким образом, подвергнется естественному отбору. В силу строгого принципа наследственности отобранная разновидность будет стремиться размножаться в своей новой и измененной форме".
Другими словами, естественный отбор, или переживание наиболее приспособленных, представляет собой сохранение полезных индивидуальных различий или изменений и уничтожение вредных. Изменения, нейтральные по своей ценности (неполезные и невредные), не подвергаются действию отбора, а представляют непостоянный, колеблющийся элемент изменчивости.
Разумеется, отдельные особи, обладающие каким-то новым полезным признаком, могут погибнуть, не оставив потомства, по чисто случайным причинам. Однако влияние случайных факторов уменьшается, если полезный признак появляется у большего числа особей данного вида - тогда возрастает вероятность того, что по крайней мере для части этих особей достоинства нового полезного признака сыграют свою роль в достижении успеха в борьбе за существование. Отсюда следует, что естественный отбор является фактором эволюционных изменений не для отдельных организмов, рассматриваемых изолированно друг от друга, но лишь для их совокупностей, т. е. популяций. Понимание этого вероятностного аспекта действия естественного отбора, эффективного только на достаточно больших выборках, и является важнейшим результатом популяционного подхода Дарвина, о котором мы уже упоминали выше. Естественный отбор мог быть открыт только при таком популяционном подходе, учитывающем количественную, вероятностную сущность эволюционных изменений.
Естественный отбор сам по себе не вызывает изменчивости организмов, которая стимулируется изменениями внешних условий.
Важнейшее место в теории естественного отбора занимает концепция борьбы за существование. Согласно Дарвину, борьба за существование является результатом тенденции любого вида организмов к безграничному размножению. Приведя многочисленные примеры невозможности выживания всего потомства у различных видов организмов, Дарвин заключает: "Так как производится более особей, чем может выжить, в каждом случае должна возникать борьба за существование либо между особями того же вида, либо между особями различных видов, либо с физическими условиями жизни".
Термин "борьба за существование" (struggle for existence) не вполне точно соответствует тому значению, которое вкладывал в него сам Дарвин, предлагая понимать этот термин "в широком и метафорическом смысле". Во-первых, Дарвин включал в понятие "существование" не только жизнь данной особи, но и успех ее в оставлении потомства. Во-вторых, словом "борьба" обозначалась не столько борьба как таковая (т.е. как прямое столкновение), сколько конкуренция, часто происходящая в пассивной форме. В сущности, Дарвин понимал под борьбой за существование совокупность всех сложных взаимодействий между организмом и внешней средой, определяющих успех или неудачу данной особи, в ее выживании и оставлении потомства.
Подчеркивая роль перенаселения как фактора, обусловливающего борьбу за существование, Дарвин сделал вывод, что наиболее ожесточенной должна быть внутривидовая борьба как конкуренция между особями одного вида, которые обладают сходными жизненными потребностями. Если, например, рассматривать модель взаимоотношений между видом-хищником и видом-жертвой (скажем, лисы и зайцы), то, по Дарвину, важнейшим фактором, определяющим отбор, будет для лис конкуренция между разными лисами, а для зайцев - между самими зайцами.
Действию естественного отбора благоприятствуют: 1) длительность времени; 2) высокая изменчивость; 3) большое число особей данного вида; 4) обширность его ареала (области распространения); 5) изоляция групп особей данного вида, уменьшающая их скрещивание с обитателями других частей ареала. Дарвин подчеркнул, что естественный отбор должен действовать с гораздо большей эффективностью, чем искусственный, поскольку, во-первых, природа располагает неизмеримо большим временем, чем человек; во-вторых, человек, ведущий искусственный отбор, обращает внимание главным образом на внешние признаки животных и растений, тогда как для естественного отбора важна любая особенность организмов; в-третьих, искусственный отбор ведется для нужд человека, а естественный отбирает признаки, важные для самого организма; наконец, в-четвертых, естественный отбор действует гораздо более жестко, так как человек обычно не истребляет всех менее пригодных домашних животных, сохраняя их для различных нужд. Все это в совокупности подчеркивает огромные творческие возможности естественного отбора, если человек при всех этих ограничениях сумел посредством искусственного отбора создать столь многочисленные и разнообразные породы домашних животных и сорта сельскохозяйственных растений.
Результаты действия естественного отбора
Естественный отбор является неизбежным результатом борьбы за существование и наследственной изменчивости организмов. По Дарвину, естественный отбор представляет собой важнейшую творческую силу, которая направляет эволюционный процесс и закономерно обусловливает возникновение приспособлений организмов, прогрессивную эволюцию и увеличение разнообразия видов.
Возникновение приспособлений (адаптации) организмов к условиям их существования, придающее строению живых существ черты "целесообразности", является непосредственным результатом естественного отбора, поскольку самая сущность его - дифференцированное выживание и преимущественное оставление потомства именно теми особями, которые в силу своих индивидуальных особенностей лучше других приспособлены к окру-жаюшим условиям. Накопление отбором от поколения к поколению тех признаков, которые дают преимущество в борьбе за существование, и приводит постепенно к формированию конкретных приспособлений.
Дарвин подчеркивал, что любая конкретная степень приспособленности организмов относительна - обычно возможны и более совершенные формы приспособлений к данной среде обитания. Это доказывается многочисленными примерами чрезвычайно быстрого размножения и широкого распространения целого ряда видов животных и растений в совершенно новых для них районах земного шара, куда они были случайно или преднамеренно завезены человеком (кролики в Австралии, крысы, кошки, собаки, свиньи на островах Океании, канадская элодея в водоемах Европы и т. п.). Все эти виды, возникшие в совершенно других географических районах, оказались лучше приспособленными к условиям новых для себя областей, чем виды животных и растений, издавна населявшие эти области и обладавшие достаточно совершенными приспособлениями к их условиям.
Теория естественного отбора позволяет объяснить развитие самых сложных и совершенных приспособлений, в том числе таких, как взаимное приспособление друг к другу двух разных видов, взаимодействие которых выгодно для обоих. Таковы, например, взаимные приспособления цветковых растений и опыляющих их насекомых.
Действуя в видовых популяциях, естественный отбор способствует формированию и распространению особенностей, важных и полезных для вида в целом: при этом может возникнуть противоречие между "интересами" вида и отдельных особей. В этом случае закрепляются признаки, полезные для вида, несмотря на их отрицательную роль для отдельных особей. Так развиваются взаимные приспособления особей в общинах и колониях различных организмов, причем во многих случаях особь теряет свое прежнее значение дискретной и самостоятельной единицы, становясь частью целого (колониальные организмы гидроидных полипов, сифонофор и т.п.; "семьи" общественных насекомых - термитов, муравьев, пчел и др., в которых наблюдается сложнейшая дифференциация особей, выполняющих различные функции и неспособных существовать и давать потомство как самостоятельные индивиды).
Вторым (после возникновения адаптации) важнейшим следствием борьбы за существование и естественного отбора является, по Дарвину, закономерное повышение разнообразия форм организмов, носящее характер дивергентной эволюции. Поскольку наиболее острая конкуренция ожидается между наиболее сходно устроенными особями данного вида в силу сходства их жизненных потребностей, в более благоприятных условиях окажутся наиболее уклонившиеся от среднего состояния индивиды. Эти последние получают преимущественные шансы в выживании и оставлении потомства, которому передаются особенности родителей и тенденция изменяться дальше в том же направлении (длящаяся изменчивость). При преимущественном сохранении в каждом поколении самых крайних вариантов изменчивости, очевидно, что эволюция пойдет в направлении разделения вида на разновидности, которые со временем превратятся в новые (дочерние) виды. По Дарвину, предковая и промежуточная формы имеют худшие шансы для выживания по сравнению с наиболее уклонившимися дочерними формами, поскольку первые более сходны друг с другом, и конкуренция между ними должна быть наиболее ожесточенной. В итоге от общего предка в ходе эволюции должны происходить все более разнообразные и отличающиеся друг от друга потомки (рис. 2).

Идея дивергентной эволюции объясняет не только повышение разнообразия форм организмов, но и существование в природе групп сходных видов - родов, группировок сходных родов - семейств и таксонов более высоких категорий. Все эти группировки сходных форм представляют собой совокупности родственных видов, возникших от общего предка; роды, семейства, отряды, классы - разные этапы дивергентной эволюции. Сходство видов, принадлежащих к одному таксону ("единство типа"), является результатом родства, общие признаки унаследованы от общего предка. Заметим, что в концепции Ламарка этот важный вопрос, в сущности, просто обойден.
Наконец, третье важнейшее следствие естественного отбора - постепенное усложнение и усовершенствование организации, т.е. эволюционный прогресс. Согласно Ч.Дарвину, это направление эволюции является результатом приспособления организмов к жизни в постоянно усложняющейся внешней среде. Усложнение среды происходит, в частности, благодаря дивергентной эволюции, увеличивающей число видов. Усовершенствование реакций организмов на усложняющуюся среду приводит к постепенному прогрессу организации. При этом Дарвин подчеркивал, что естественный отбор сам по себе отнюдь не предполагает обязательно прогрессивного развития, действуя "только посредством сохранения и накопления изменений, благоприятных для организма при тех органических и неорганических условиях, в которых он существует во все периоды своей жизни". Если для данного вида совершенствование организации почему-либо невыгодно, естественный отбор отнюдь не будет способствовать его прогрессивной эволюции. "При очень простых жизненных условиях высокая организация не оказала бы никакой услуги, пожалуй, даже оказала бы дурную услугу, так как, вследствие своей хрупкости, была бы более подвержена повреждению и порче". Одновременное существование на Земле сложно построенных, далеко ушедших по пути прогресса организмов и примитивных форм, сохранивших простое строение, является результатом эволюции различных групп организмов под контролем естественного отбора в разной жизненной среде. В одних условиях для организмов выгодно совершенствоваться, в других - сохранять простое строение; и то и Другое обеспечивается отбором.
Частным случаем естественного отбора является половой отбор, который связан не с выживанием данной особи, а лишь с ее воспроизводительной функцией. По Дарвину, половой отбор возникает при конкуренции между особями одного пола в процессах размножения. Важность воспроизводительной функции самоочевидна; поэтому в некоторых случаях даже само сохранение данного организма может отступить на второй план по отношению к оставлению им потомства. Для сохранения вида жизнь данной особи важна лишь постольку, поскольку она участвует (прямо или косвенно) в процессе воспроизводства поколений. Половой отбор как раз и действует на признаки, связанные с различными аспектами этой важнейшей функции (взаимное обнаружение особей противоположного пола, половая стимуляция партнера, конкуренция между особями одного пола при выборе полового партнера и т. п.).
Половой отбор определяет различное участие разных особей в размножении: некоторые индивиды получают лучшие шансы оставить потомство (и передать ему свои признаки, обеспечившие этот успех), чем другие. Механизмами полового отбора являются, с одной стороны, активное соперничество между особями одного пола (обычно между самцами), с другой же - предпочтение, оказываемое определенным особям другого пола при выборе полового партнера. Следствием полового отбора является развитие внешних признаков, различающих самцов и самок (вторичные половые признаки). Среди наиболее ярких примеров действия полового отбора можно назвать развитие рогов оленей, используемых как "турнирное оружие" при столкновениях самцов во время гона, ярких окрасок и причудливого оперения самцов фазанов, знаменитого хвоста самцов павлинов и т. п.
Заканчивая это - по необходимости очень краткое - рассмотрение теории Дарвина, перечислим важнейшие ее положения.
1. Организмам как в прирученном, так и в диком состоянии свойственна наследственная изменчивость. Наиболее обычной и важной формой изменчивости является неопределенная. Стимулом для возникновения изменчивости организмов служат изменения внешней среды, но характер изменчивости определяется спецификой самого организма, а не направлением изменений внешних условий.
2. В центре внимания эволюционной теории должны находиться не отдельные организмы, а биологические виды и внутривидовые группировки (популяции).
3. Все виды организмов в природе вынуждены вести жестокую борьбу за свое существование. Борьба за существование для особей данного вида складывается из их взаимодействия с неблагоприятными биотическими и абиотическими факторами внешней среды, а также из их конкуренции между собой. Последняя является следствием тенденции всякого вида к безграничному размножению и огромного "перепроизводства" особей в каждом поколении. По Дарвину, важнейшей является именно внутривидовая борьба.
4. Неизбежным результатом наследственной изменчивости организмов и борьбы за существование является естественный отбор - преимущественное выживание и обеспечение потомством лучше приспособленных особей. Хуже приспособленные организмы (и целые виды) вымирают, не оставляя потомства.
5. Следствиями борьбы за существование и естественного отбора являются: развитие приспособлений видов к условиям их существования (обусловливающее "целесообразность" строения организмов), дивергенция (развитие от общего предка нескольких дочерних видов, все большее расхождение их признаков в эволюции) и прогрессивная эволюция (усложнение и усовершенствование организации).
6. Частным случаем естественного отбора является половой отбор, который обеспечивает развитие признаков, связанных с функцией размножения.
7. Породы домашних животных и сорта сельскохозяйственных растений созданы посредством искусственного отбора, аналогичного естественному отбору, но ведущегося человеком в своих интересах.
Таким образом, теория Дарвина дала логически последовательное и строго материалистическое объяснение важнейшим проблемам эволюции организмов и сложившейся в результате эволюционного процесса обшей структуре органического мира. Дарвин первым доказал реальность эволюционных изменений организмов. Взаимоотношения организма и внешней среды в его теории имеют характер диалектического взаимодействия: Дарвин подчеркивал роль изменений среды как стимула изменчивости организмов, но, с другой стороны, специфика этих изменений определяется самими организмами, и дивергентная эволюция организмов изменяет среду их обитания. Учение о естественном отборе и борьбе за существование представляет собой, в сущности, анализ этих сложных взаимоотношений организма и среды, в которых организм не противопоставляется среде как саморазвивающаяся автономная единица, но и не следует пассивно за изменениями среды (как трактуются взаимоотношения организма и среды в теории Ламарка). Согласно теории Дарвина, эволюция представляет собой результат взаимодействия организма и изменяющейся внешней среды.
Современная эволюционная теория сложилась на основе теории Дарвина. В принципе признавая это и оценивая конкретное место идей Дарвина в совокупности современных эволюционных взглядов, нередко впадают в одну из двух крайностей: либо считают, что ныне концепция Дарвина как таковая имеет уже только исторический интерес --от нее в современной науке осталась лишь сама идея естественного отбора; либо, напротив, утверждают, что со времени Дарвина основы теории не претерпели существенных изменений. В действительности, как это нередко бывает, истина находится между этими крайними точками зрения. Хотя, несомненно, современная теория эволюции является развитием основных идей Дарвина, но в процессе их развития выявлены многие неизвестные во времена Дарвина факторы, механизмы и закономерности эволюционного процесса и сформировались новые представления, значительно отличающиеся от классической теории Дарвина.
В дальнейшем, рассматривая современную эволюционную теорию, мы будем специально оговаривать ее основные различия со взглядами Дарвина. Здесь же необходимо упомянуть о некоторых нечеткостях и отдельных ошибочных утверждениях Дарвина. К ним относятся: 1) признание возможности эволюционных изменений на основе определенной изменчивости и упражнения и неупражнения органов; 2) переоценка роли перенаселения для обоснования борьбы за существование; 3) преувеличенное внимание к внутривидовой борьбе в объяснении дивергенции; 4) недостаточная разработанность концепции биологического вида как формы организации живой материи, принципиально отличающейся от подвидовых и надвидовых таксонов; 5) непонимание специфики макроэволюционных преобразований организации и их соотношений с видообразованием. Однако все эти не вполне отчетливые или даже неверные представления по некоторым вопросам отнюдь не умаляют исторической значимости гениального труда Дарвина и его роли для современной биологии. Указанные неточности соответствуют уровню развития науки во время создания теории Дарвина.
Так, допущение Дарвином прямой эволюционной роли определенной изменчивости и наследования результатов упражнения и неупражнения органов было связано с господствовавшими в его время представлениями о наследственности. Тогда наследственность рассматривали как своего рода "общее свойство" всего организма, присущее ему как целому ("слитная наследственность"). С этим связано одно из возражений против теории естественного отбора, выдвинутое Ф.Дженкином и казавшееся самому Дарвину очень серьезным. Суть этого возражения сводилась к следующему. Пусть Дарвин прав, и эволюция организмов основывается на незначительных изменениях отдельных особей, первоначально неопределенных по отношению к изменениям внешней среды. Однако поскольку новые полезные признаки первоначально появляются у немногих особей и притом слабо выражены, то при скрещивании с другими особями того же вида, подавляющее большинство которых еще не имеет данного нового признака, неминуемо должно происходить как бы "разбавление" полезного новоприобретения вдвое в каждом последующем поколении. И постепенно новый признак практически исчезнет, распределившись между множеством особей, и будет поглощен консервативным состоянием. Возражения Дженкина были одной из причин, побудивших Дарвина в более поздних изданиях "Происхождения видов" и других сочинениях в большей мере, чем прежде, склоняться к признанию эволюционной роли определенной изменчивости, т.е. наследования признаков, приобретаемых организмами под прямым воздействием изменяющихся внешних условий.
Еще раз подчеркнем, что все указанные неточности и ошибки Не искажают основных идей Ч.Дарвина. Остановиться на них следовало для того, чтобы, во-первых, понимать их действительное место в теории Дарвина и, во-вторых, чтобы яснее видеть корни ряда ошибочных концепций позднейшего времени, авторы которых нередко ссылаются на авторитет Дарвина. К рассмотрению ряда таких концепций, являющихся в действительности развитием идей ламаркизма, мы теперь и перейдем.
Прежде всего это простота и кажущаяся очевидность (особенно для малоосведомленного в вопросах теории эволюции человека) некоторых постулатов ламаркизма. Особенно это относится к идее наследования изменений (модификаций), приобретенных организмами под прямым воздействием изменяющихся внешних условий, и последствий упражнения и неупражнения органов. Эти модификации, являющиеся целесообразной приспособительной реакцией организма на различные внешние условия, представляют собой самые обычные и легко наблюдаемые изменения организмов. Стоит лишь допустить их наследование - и готов чрезвычайно простой "механизм эволюционных изменений", вполне удовлетворяющий профана.
Изучая ряды последовательных форм по ископаемым остаткам различных групп организмов, палеонтологи обращали внимание на существование некоторых общих тенденций и закономерностей филогенеза (параллельная эволюция родственных филогенетических линий, пульсирующий характер темпов макроэволюции, эволюционный прогресс и др.). Для неодарвинистской синтетической теории эволюции характерна концентрация внимания на процессах микроэволюции и видообразования, с чем связана тенденция рассматривать макроэволюцию просто как сумму микроэволюционных изменений, игнорируя ее специфику. Однако попытки такого редукционистского решения проблем макроэволюции наталкивались на серьезные трудности. Это создавало благоприятную почву для их истолкования в духе неоламаркизма.
Наконец, еще одной причиной, определяющей склонность некоторых исследователей к идеям ламаркизма, является отсутствие "популяционного мышления", т. е. ясных представлений о сущности и структуре биологического вида и понимания вероятностного характера действия естественного отбора. В связи с этим такие исследователи склонны недооценивать эффективность естественного отбора и рассматривать модификационные изменения, наблюдаемые в онтогенезе организмов, как "направленную изменчивость", принимаемую за основу эволюционного процесса.
Основные направления неоламаркизма
Неоламаркизм не представляет собой единой целостной концепции: различные ученые, склонные по тем или другим причинам разделять ламаркистские идеи, чаще всего принимают лишь одну из двух сторон теории Ламарка - либо автогенетический подход к объяснению прогресса и направленности эволюции, либо эктогенетические взгляды на развитие приспособлений организмов к изменяющимся условиям внешней среды. При этом у многих авторов неоламаркистские взгляды маскируются использованием собственной терминологии и принятием некоторых отдельных положений теории Дарвина. Такие авторы иногда называли себя "дарвинистами", а систему своих взглядов - "творческим дарвинизмом", претендуя на дальнейшее развитие дарвиновской теории, хотя на деле эти взгляды представляют собой эклектическое соединение теории естественного отбора с ламарковской эктоге-нетической концепцией, включающей постулат изначально целесообразной реакции организмов на изменения внешних условий и "закон" наследования приобретаемых признаков.
Автогенетические идеи в духе ламаркизма выдвигали Э. Коп (1840-1897) ("батмогенез" -- стремление организмов к прогрессу), Г.Осборн (1857-1935) ("аристогенез"), О.Шиндевольф (1896-1971), П.Тейяр де Шарден (1881-1955). К этому же кругу идей принадлежат теории ортогенеза Т. Эймера (1843-1898) и номогенеза (или "эволюции на основе закономерностей") Л.С.Берга (1876-1950). Во всех этих концепциях в разных вариантах организмам приписывается "стремление" к прогрессу, к усложнению, к усовершенствованию и т.п. Сущность этих взглядов принципиально не отличается от идеи Ламарка о градации: это телеологические и метафизические концепции, подменяющие объяснение наблюдаемых в природе процессов и явлений декларированием неких "изначальных свойств" живой материи. Вопросы о сущности и первопричине "стремления" организмов к прогрессу обычно просто обходятся авторами подобных автогенетических теорий, которые при логически последовательном своем развитии неминуемо смыкаются с идеализмом. Мы вернемся к оценке этих концепций в четвертой части книги, посвященной проблемам макроэволюции.
Гораздо большее число сторонников находят эктогенетические взгляды, согласно которым эволюционные изменения организмов являются результатом целесообразной их реакции на изменения внешних условий. Как мы уже отмечали, эта вторая часть теории Ламарка не является его оригинальной идеей и принималась тогда в той или иной форме всеми трансформистами. В более позднее время наивные формулировки Ламарка и его предшественников обычно заменяют более "современными", хотя сущность их остается прежней. Таково, например, учение Т.Д.Лысенко, согласно которому организмы в процессе обмена веществ "ассимилируют условия внешней среды", которые после этого сами становятся частью наследственности, понимаемой в духе представлений середины XIX в. - как некое общее свойство организма, как "эффект концентрирования воздействий условий внешней среды, ассимилированных организмами в ряде предшествующих поколений". Если отбросить словесную шелуху, очевидно, что здесь в "оригинальных" терминах излагается ламарковское объяснение изменений растений под влиянием изменений внешних условий. Что, собственно, такое "ассимиляция внешних условий", как она происходит и что именно изменяется при этом в организме, Лысенко и его последователи не разъясняли.
В разных вариантах эктогенетические идеи высказывали такие известные ученые, как Э.Геккель, Э. Коп, Г. Спенсер, К. Негели, Ф. Вайденрайх, Е.С.Смирнов, Л.Ш.Давиташвили и некоторые другие.
Адекватная соматическая индукция
Сущность воззрений ученых, стоящих на позициях эктогенеза, сводится к признанию так называемой адекватной соматической индукции, под которой понимается способность организмов передавать по наследству модификационные изменения, возникающие в онтогенезе как приспособления к изменившимся внешним условиям. Адекватную соматическую индукцию можно описать по следующей схеме (рис. 3). Изменение внешней среды, например Устойчивое повышение температуры, воздействует на организм. Тот реагирует на это воздействие соответствующим модификационным изменением тела (сомы) - например увеличением густоты меха. Это изменение каким-то образом передается половым клеткам ("индукция" со стороны соматических клеток, и притом адекватная, т.е. соответствующая изменениям последних), и в следующем поколении новый признак (в нашем примере - большая густота меха) будет выражен с самого начала, без дополнительного воздействия внешних условий.

Анализируя эту схему, нетрудно убедиться, что первый ее этап, безусловно, представляет собой самое обычное явление. Всякий организм обладает определенными возможностями саморегуляции, перестройки своих функций и (в несколько меньшей степени) формы в соответствии с изменениями внешних условий. Особенно пластичен молодой, развивающийся организм, и в разных условиях он может приобретать различный облик (фенотип): сравните, например, сосну, ель, дуб или другие деревья, выросшие на открытом месте, с такими же деревьями, но растущими в глубине леса. Сюда же относится пример Ламарка с подводными и надводными листьями лютика Ranunculus aquatilis, а также результаты усиленного упражнения или неупражнения органов (это общеизвестно, например, для мышц) и т. п. Все это - примеры модификационной изменчивости, представляющей собой комплекс реакций данного организма (в конечном счете наследственного аппарата, или генотипа организма) на разные внешние условия. Каждый генотип обладает определенной нормой реакции, под которой понимается его способность в разных условиях обеспечивать формирование разного состояния признаков организма. Все модификационные изменения, следовательно, не есть что-то новое для вида; все они входят в норму реакции генотипов данного вида на различные комбинации внешних условий. В связи с этим модификационные изменения соматических структур не оказывают никакого воздействия на половые клетки (т. е. соматической индукции вообще не происходит).
В более редких случаях под влиянием внешних воздействий могут возникнуть изменения аппарата наследственности соматических клеток - соматические мутации. Однако появляющиеся в результате соматических мутаций изменения фенотипа не являются приспособительными к внешним факторам, вызвавшим возникновение этих мутаций. По отношению к этим факторам фенотипические изменения носят случайный характер (например, появление пигментных пятен или опухолевое перерождение тканей в ответ на действие ионизирующего облучения). Поэтому соматические мутации не соответствуют основной для неоламаркистов идее о целесообразной реакции организма на внешние воздействия. Зато соматические мутации в отличие от модификаций могут в принципе обусловить соматическую индукцию: изменения соматических структур в этом случае являются новообразованиями и могут так повлиять на внутреннюю среду организма, что эти изменения отразятся и на половых клетках. Хотя такое воздействие сомы на гаметы возможно, но отнюдь не обязательно, так как половые клетки в организме обычно имеют максимально возможную защиту от всякого рода внешних воздействий.
Необходимо подчеркнуть, что в организме нет специальных систем, которые обеспечивали бы передачу неискаженной наследственной информации от изменившихся соматических клеток к половым (подробнее см. в первой главе следующей части книги). Более того, такие системы были бы крайне вредны для организмов: ведь при их наличии наследственный аппарат каждого организма в течение его жизни подвергался бы непрерывным перестройкам, наследственность вида потеряла бы свой консерватизм и не смогла бы сохранять и передавать последующим поколениям постоянные признаки; вид как целостная система, состоящая из принципиально сходных скрещивающихся друг с другом особей, Не смог бы существовать. Никакой достаточно сложный организм, требующий для своего индивидуального развития точного соответствия разнообразнейших морфогенетических процессов и взаимных влияний развивающихся структур, не смог бы возникнуть в эволюции на основе подобной лабильной, неустойчивой к внешним воздействиям наследственности.
Иногда термин "адекватная соматическая индукция" заменяют более простым "наследованием приобретаемых свойств (признаков)". Такая замена неудачна: строго говоря, любое наследственное изменение является именно "наследованием приобретаемых свойств" - если никакое новоприобретение не наследуется, нет и никакого эволюционного изменения организмов. Поэтому термин "наследование приобретаемых свойств" по отношению ко взглядам ламаркистов также нуждается в уточнениях (следует говорить об адекватном наследовании новоприобретенных признаков соматических структур).
Попытки экспериментального доказательства неоламаркизма
Таким образом, адекватная соматическая индукция, на признании которой основываются неоламаркистские концепции, представляется в принципе невозможной. Однако неоламаркисты пытались доказать свои взгляды разного рода наблюдениями и экспериментами. Собранные ими данные довольно многочисленны и разнородны (см. Бляхер Л.Я. Проблема наследования приобретенных признаков.-М.,1971), но все они при последующем научном анализе либо оказывались результатом ошибок в постановке экспериментов и не подтверждались при проверке, либо представляли собой неверно истолкованные факты, отнюдь не доказывающие того, что желали доказать с их помощью неоламаркисты.
Попыткам доказать наследственные изменения организмов под прямым воздействием изменяющихся внешних условий были посвящены многие работы П.Каммерера, выполненные в 1910-1923 гг. Упомянем из их числа достаточно показательные эксперименты с протеями и пятнистыми саламандрами.
Пещерные земноводные - протеи в норме обладают недоразвитыми глазами и слабо пигментированной кожей. Каммерер выращивал протеев в лаборатории на свету и при более высокой температуре, чем в природных местообитаниях этих животных. В первом же поколении у протеев в лабораторных условиях кожа становилась темной, а глаза развивались в значительно большей степени, чем в природе. При этом протеи переходили к откладке яиц вместо живорождения, обычного для них в пещерных водоемах, где температура воды ниже 15 °С. Эти результаты Каммерер интерпретировал как пример изменений организма, обусловленных изменениями внешней среды и наследуемых. Однако на деле все обстоит как раз наоборот, что было справедливо отмечено Э.Гийено и А.Н.Северцовым. Способ размножения, недоразвитие глаз и пигментации покровов - признаки, развившиеся у протея в результате приспособления к жизни в пещерах в течение многих тысяч поколений. Опыты Каммерера показали, что эти признаки как таковые ненаследственны, поскольку в первом же поколении, выращенном при условиях "наземной" жизни они модифицировались к состоянию, которое было, вероятно, присуще предкам протея до их перехода к пещерной жизни. Очевидно, сильная или слабая пигментация покровов, большая или меньшая степень развития глаз, откладка яиц или живорождение представляют собою разные модификации, т.е. фенотипические варианты, развивающиеся в разных условиях на основе одной и той же нормы реакции остающегося неизменным по этим признакам аппарата наследственности. Таким образом, наблюдения Каммерера показали, вопреки его собственному мнению, как раз ненаследственный характер рассмотренных им признаков протея.
Наиболее известны опыты Каммерера с пятнистыми саламандрами. Окраска этих земноводных состоит из черных и желтых пятен неправильной формы. При содержании личинок саламандр на желтом фоне субстрата у животных развивалось преобладание желтой, а на черном фоне - более темной окраски. Эти особенности сохранялись и усиливались в следующем поколении при содержании саламандр на том же фоне, но изменялись в обратном направлении при помещении на другой фон. Последнее показывает, что и в этом случае имеет место модификационное изменение окраски в соответствии с фоном внешней среды (вообще нередкое явление среди земноводных), но, кроме того, в данной работе Каммерера проявилась характерная методическая ошибка, которую часто допускают неоламаркисты. В подобных случаях экспериментатор использует не чистую линию организмов, состоящую из генетически однородных особей, а случайную выборку, включающую организмы с разным наследственным состоянием изучаемого признака. В природных популяциях саламандр имеется значительная индивидуальная изменчивость окраски с наследственно обусловленным преобладанием черного или желтого цвета у различных особей. В эксперименте, подобном поставленному Каммерером, используются генетически разнокачественные особи, которые по-разному реагируют на фоновую окраску субстрата. Экспериментатор выбирает для дальнейших исследований тех животных, которые "лучше реагируют" на изменение условий (т.е. в данном случае тех. которые с самого начала наследственно предрасположены к большему развитию черной или желтой окраски в соответствующих вариантах опыта). Естественно, что эти особи передают потомству свой наследственный признак, который отнюдь не приобретен ими в ходе эксперимента, но существовал уже в природных популяциях как вариант изменчивости данного вида. Во всех подобных случаях экспериментатор производит своего рода "бессознательный искусственный отбор" в первоначально гетерогенной популяции, который и обусловливает изменение среднего состояния признака в следующем поколении в желательном для экспериментатора направлении. Налицо серьезная методическая ошибка.
Следует отметить еще, что Каммерер не регистрировал каких-либо точных количественных характеристик, все его описания сделаны на обобщенном качественном уровне (более темная или более светлая окраска). К. Гербст, повторивший опыты Каммерера, применил более точную регистрацию изменений цвета и пришел к выводу, что наметившиеся у личинок саламандр модифи-кационные изменения окраски не сохраняются после метаморфоза у взрослых особей.
Обратимся к нескольким более поздним работам. В 50-е гг. энтомологи Е.С.Смирнов, С.И.Келейникова и Г.В.Самохвалова выкармливали тлей Neomyzus circumflexus на малопригодных для питания этого вида растениях (вместо вики - на красном перце, горчице, гречихе и др.). Это сначала приводило к снижению плодовитости, а в опытах Самохваловой - также к повышенной смертности тлей, но начиная с седьмого-девятого поколений происходило постепенное "привыкание" насекомых к новым кормам, выражавшееся в увеличении плодовитости (и снижении смертности в опытах Самохваловой). При этом проявлялись морфологические различия с исходной популяцией тлей в рисунке кутикулы спинки, что Самохвалова трактовала как возникновение новой формы тлей. Совершенно очевидно, что и в этом случае речь идет о генетически гетерогенном исходном материале, включавшем тлей с разной наследственной способностью использовать новые корма, скоррелированной с особенностями склеротизации спинки насекомого. В ряду последовательных поколений происходил отбор именно этих изначально наиболее устойчивых к новым условиям особей, и в ходе эксперимента отнюдь не была получена какая-либо новая форма тлей, а просто выделена посредством отбора из разнородных особей линия насекомых, лучше усваивающих необычные корма. Характерно, что Е.С.Смирнов отрицал роль отбора в полученных им результатах на том основании, что примененные в его опытах воздействия не повышали смертности тлей. Однако отбор вовсе не требует обязательной гибели особей -- для эффективного отбора совершенно достаточно снижения плодовитости одних и более высокой плодовитости других особей в новых условиях!
Неоламаркисты привлекали также в качестве примера наследования особенностей, вызванных влиянием среды на организм, возникновение так называемых длительных модификаций у простейших, впервые описанных В.Йоллосом в 1920-е гг. Длительные модификации представляют собою постепенные изменения, развивающиеся, например, в популяции инфузорий при длительном воздействии экстремальных условий (высокая температура, различные химические воздействия) и повышающие устойчивость простейших к этим неблагоприятным условиям. Устойчивость к неблагоприятным температурам или химическим веществам сохранялась у инфузорий в течение многих десятков (до сотни) поколений и, следовательно, была наследственной. Позднее при отсутствии новых воздействий экстремального фактора, т. е. в нормальных условиях, длительная модификация исчезает. Подчеркивается, что, во-первых, вся экспериментальная популяция в этих опытах представляла собой клон, полученный путем бесполого размножения от одной исходной особи, и, во-вторых, в эксперименте отсутствовала смертность инфузорий, вызванная действием экстремального фактора. Это истолковывали как "исключение возможности действия отбора".
Анализ развития длительных модификаций у простейших был сделан Ж. Женермоном. Женермон указал, что прежде всего, хотя экспериментальная популяция инфузорий и представляет собой клон, но у инфузорий, на которых сделано большинство экспериментов, это отнюдь не исключает генетической гетерогенности популяции. Такая гетерогенность возникает у инфузорий на основе специфичности макронуклеусов, приобретаемой в моментдиф-ференцировки макронуклеусов из микронуклеуса при автогамии или даже в течение вегетативной жизни особей. Во-вторых, как и в разобранных выше опытах Е.С.Смирнова, в процессе развития длительных модификаций, очевидно, основную роль играет отбор, влияющий не на смертность, а на темпы размножения: преобладание в популяции получает более многочисленное потомство более устойчивых к действию неблагоприятного фактора особей. Наконец, в развитии длительных модификаций могут играть роль и обратимые изменения цитоплазматических структур, передающиеся следующему поколению через наследственный аппарат цитоплазмы (плазмотип, см. главу 1 ч. II).
Сторонниками Т.Д.Лысенко много говорилось и писалось о вегетативной гибридизации организмов как доказательстве возможности адекватного влияния соматических клеток на половые. При этом под вегетативной гибридизацией понимали возможность получения гибрида не путем гибридизации геномов соматических клеток, что в принципе вполне возможно (например, такой "соматический" (вегетативный) гибрид двух видов табака Nicotiana glauca и N. langsdorfli был получен П.Карлсоном путем слияния изолированных протопластов, извлеченных из мезофилла листьев), а посредством прививок черенков одного сорта или вида растений на растение другого сорта или вида. Это подразумевает обмен организмов наследственными свойствами без непосредственного объединения каких бы то ни было ядерных и вообще внутриклеточных структур через посредство "пластических веществ", выделяемых клетками. Такая гибридизация невозможна, поскольку в "пластических веществах" отсутствует наследственная информация. Изменения растений, возникшие в результате прививок черенков другого сорта, сами по себе, конечно, вполне реальны и хорошо известны садоводам, но не передаются по наследству. "Вегетативные гибриды" представляют собой генетические химеры - так называют мозаичные организмы, объединяющие в себе клетки, ткани или органы разных организмов. В организмах-химерах клетки с разным наследственным аппаратом сосуществуют, но не обмениваются наследственной информацией. При половом размножении в этом случае половые клетки получают наследственный аппарат только одного типа и фенотипические свойства организма-химеры не передаются потомству. Именно поэтому сортовые качества плодовых деревьев, полученных путем прививок и также являющихся генетическими химерами, не передаются при размножении семенами: из семян вырастает дичок, снова нуждающийся в прививке черенками культурных сортов для приобретения соответствующих сортовых качеств.
Данные, приводившиеся в доказательство возможности передачи наследственных свойств "вегетативных гибридов", при проверках не подтвердились (подробнее см. в кн.: Краева С.Я. "Возможна ли вегетативная гибридизация растений посредством прививки?", 1967). Подлинная же гибридизация соматических клеток, основанная на объединении их наследственных аппаратов, очевидно, не имеет ни малейшего отношения к адекватной соматической индукции.
Таким образом, наблюдения и эксперименты, которые пытались использовать для подтверждения неоламаркистских идей, либо являются примерами модификационной изменчивости, ошибочно принимаемой за появление новых наследственных признаков, либо представляют собой результат отбора в генетически неоднородной популяции, либо недостоверны. В последнее десятилетие появились попытки использовать для доказательства возможности адекватной соматической индукции некоторые новейшие достижения молекулярной биологии. Эти работы мы рассмотрим в первой главе следующей части книги.
Неоламаркизм и основные проблемы эволюционной теории
Ламаркизм в любой его форме не дает объяснения ни прогрессивной эволюции, ни возникновению адаптации (приспособлений) организмов, поскольку "стремление к прогрессу", "эволюция на основе закономерностей", "изначальная способность организмов к целесообразной реакции", "ассимиляция условий внешней среды" и другие подобные концепции подменяют научный анализ постулированием неких метафизических свойств, якобы присущих живой материи.
Ламаркизм не способен объяснить появление в эволюции организмов каких бы то ни было качественных новообразований: никакое "прямое влияние измененных внешних условий" не может объяснить, например, замену хорды позвоночником в эволюции позвоночных животных, или развитие легких, или возникновение сердца, если, конечно, не считать объяснением ламарков-ское "усилие внутреннего чувства". Ламаркизм беспомощен и в анализе причин, обусловивших возникновение колониальных и общественных организмов и развитие приспособлений у нераз-множаюшихся особей таких видов (например, у рабочих пчел или муравьев). В самом деле, как понять с ламаркистских позиций "воздействия измененных условий" или архаического "упражнения или неупражнения органов" развитие и усовершенствование сложных приспособлений для добывания пищи, строительства гнезда, заботы о потомстве у тех категорий особей данного вида, которые сами не оставляют никакого потомства (рабочие пчелы и т.п.) и уже поэтому не могут передать следующему поколению какие бы то ни было приобретенные ими признаки?
Необъяснимы для последовательно ламаркистских концепций и многие другие эволюционные явления - такие, например, как возникновение любого рода покровительственных окрасок. Покровительственные окраски делают незаметными жертву для хищника и хищника для его добычи (например, зимняя белая окраска зайцев, песцов, горностаев, куропаток и т. п., рис. 4). Своеобразный вариант покровительственной окраски представляет собой мимикрия - подражательное сходство какого-либо незащищенного вида животных (имитатор) с хорошо защищенным от врагов видом (модель). Таково, например, сходство мух-пчеловидок Eristalis tenax с пчелами, самок бабочек-парусников Papilio dardanus с несъедобными бабочками-данаидами рода Amauris (рис. 5) и т.п. Все эти и подобные им приспособления никак не могут возникнуть под влиянием "измененных внешних условий" или "упражнения органов", если только опять-таки не предполагать таинственных "усилий внутреннего чувства" у вида-имитатора.


Точно так же необъяснимо с ламаркистских позиций развитие сложных структур, образованных мертвыми тканями (различные панцири, раковины и другие защитные образования; роговые структуры, подобные, например, перьям птиц). Для решения всех этих проблем некоторые неоламаркисты вынуждены прибегать к теории естественного отбора, в результате чего взгляды многих из них (Т.Д.Лысенко и его последователи, Л.Ш.Давиташвили и др.) представляют собой эклектическое объединение ламаркистских (или, правильнее, трансформистских) и дарвинистских идей.
Подводя итоги, мы с полным основанием можем констатировать, что ламаркизм в лице самых различных его вариантов представляет собой идеалистическую и метафизическую концепцию. Ламаркизм теоретически несостоятелен, поскольку подменяет научный анализ декларированием неких изначальных свойств организмов (стремление к прогрессу, способность целесообразно реагировать на изменения внешней среды, адекватная соматическая индукция). Ламаркизм принципиально не способен решить важнейших проблем эволюционной теории. Наконец, ламаркистские взгляды не подтверждаются никакими достаточно достоверными научными фактами. Следовательно, по удачной формулировке И.И.Шмальгаузена, "ламаркизм является ложной теорией эволюции, дающей лишь мнимое объяснение развитию органического мира".
Дарвинизм и генетика
В то же время, при очевидном торжестве эволюционной идеи и всеобщем увлечении эволюционными исследованиями, многих ученых по различным причинам не удовлетворял классический дарвинизм. Некоторые из этих причин были рассмотрены в предыдущей главе в связи с обсуждением неоламаркистских концепций. При этом мы отмечали, что одной из причин, способствовавших проникновению идеи наследования приобретенных соматическими структурами новых признаков в работы самого Ч.Дарвина, были представления о так называемой "слитной наследственности" ("кошмар Дженкина"). Неразработанность проблемы наследственности организмов вызвала к жизни в конце XIX в. целый ряд умозрительных гипотез, среди которых наиболее существенную роль в истории эволюционистики сыграла концепция А.Вейсмана.
В противоположность представлениям о слитной наследственности как общем свойстве всего организма, А. Вейсман предположил, что за передачу наследственных свойств отвечает особое вещество наследственности - зародышевая плазма, состоящая из дискретных единиц - наследственных детерминантов, которые локализованы в хромосомах клеточного ядра. Представления о корпускулярной дискретной наследственности снимали опасность "растворения" новых признаков в общей "слитной наследственности" вида, поскольку наследование различных признаков определялось обособленными детерминантами. Вейсман резко выступал против неоламаркистских представлений о наследовании новоприобретаемых соматических признаков, утверждая, что в соматических клетках в отличие от половых нет полного набора наследственных детерминантов (это впоследствии оказалось ошибочным). Вейсман рассматривал естественный отбор как важнейший фактор эволюции и категорически отрицал эволюционную роль ненаследственных соматических модификаций. Взгляды А.Вейсмана во многих отношениях были ближе ко взглядам А.Уоллеса, чем к классическим представлениям дарвинизма. Концепция Вейсмана-Уоллеса получила название "неодарвинизм" (отметим, что этот термин используют и для обозначения современной синтетической теории эволюции, которая существенно отличается от вейсманизма).
Первые эксперименты, показавшие дискретность аппарата наследственности, были проделаны Г.Менделем еще в 1865 г., но его работы долго не привлекали внимания других исследователей. Лишь на рубеже XIX и XX вв. в изучении наследственности организмов были достигнуты первые существенные успехи. На основании работ Л.Кено, С.И.Коржинского, В.Иогансена, Г.де Фриза и некоторых других исследователей сложились представления о дискретной наследственности. Структурно-функциональные единицы аппарата наследственности - гены - контролируют наследование различных признаков. Новые признаки не могут быть "поглощены" консервативным состоянием потому, что их возникновение обусловлено изменениями отдельных генов, которые хотя и взаимодействуют друг с другом, но не сливаются и не разбавляют друг друга. Всякое наследственное изменение (мутация), раз появившись и выдержав "проверку на жизнеспособность", сохраняется и постепенно распространяется в популяциях данного вида, если новому признаку благоприятствуют внешние условия. Новый признак (новый вариант гена) входит в генофонд вида - сумму всей наследственной информации всех особей данного вида.
В 1906 г. Д.Харди и В. Вайнберг математически доказали, что при свободном скрещивании (панмиксии) особей в бесконечно большой популяции организмов данного вида частота встречаемости различных генов остается из поколения в поколение постоянной при отсутствии влияния каких-либо внешних факторов (например, действия отбора). Следствием этого правила Харди - Вайнберга является невозможность бесследной потери каких бы то ни было новых мутаций из генофонда бесконечно большой панмиктической популяции без воздействия извне и, разумеется, при жизнеспособности мутантов. Бесконечно большая панмикти-ческая популяция представляет собой абстракцию, реально не встречающуюся в природе, но достаточно крупные популяции со случайным скрещиванием составляющих их особей вполне обычны, и в таких популяциях проявляется тенденция к сохранению генетического равновесия в соответствии с правилом Харди-Вайнберга. С появлением в таких популяциях новых генов частота их встречаемости стабилизируется (при отсутствии отбора) уже в следующем поколении. Тем самым правило Харди-Вайнберга математически опровергает возражение Ф.Дженкина Ч.Дарвину.
В основу современных представлений о наследственности легла хромосомная теория Т. Моргана, согласно которой основными носителями генов являются хромосомы клеточного ядра; гены в хромосомах располагаются в линейном порядке.
Однако большинство пионеров научной генетики не были сторонниками теории Ч.Дарвина. Увлеченность успехами своей науки, в частности открытием мутаций, склонила их к мнению о второстепенной роли естественного отбора в эволюционном процессе. По их мнению, для объяснения эволюции организмов достаточно мутационного процесса: мутации скачкообразно приводят к формированию новых видов (мутационная теория эволюции). Мутационисты еще не знали, что подавляющее большинство признаков организма контролируется не одним геном, а несколькими (часто многими), а сложные фенотипические структуры формируются в результате взаимодействия сложных комплексов генов. Отдельные мутации разных генов сами по себе принципиально не могут сформировать гармоническое целое, тем более обеспечить развитие и преобразование сложных приспособлений организмов. Эволюция организмов в значительной степени связана именно с развитием адаптации, причем преобразование сложных многокомпонентных адаптивных комплексов контролируется многими независимыми генетическими системами. Следовательно, для объяснения эволюции мутационный процесс совершенно недостаточен - необходимо указать фактор, интегрирующий мутации и формирующий на их основе фенотипические приспособления любой сложности. Таким фактором является естественный отбор. Осознание этого пришло к генетикам не сразу.
Формирование синтетической теории эволюции
В конце XIX - начале XX в. начали формироваться представления, которые в дальнейшем легли в основу биологической концепции вида. Хотя "происхождение видов" занимает центральное положение в теории естественного отбора, Ч.Дарвин не сделал определения вида, подчеркивая отсутствие естественных границ между подвидовыми категориями (разновидностями) и видами. В связи с этим основным критерием для различения видов Дарвин считал отсутствие между ними промежуточных форм. К.Джордан, Е.Пултон и В.Ротшильд исследовали различные формы внутривидовой изменчивости (возрастную, половую, сезонную, географическую, модификационную и полиморфную) и разработали представлений о видах как совокупностях скрещивающихся друг с другом особей, которые в то же время не скрещиваются с особями, принадлежащими к другим видам, при симпатрии (т. е. совместном обитании в одном районе). Скрещиванию разных видов препятствуют различные изоляционные механизмы (стерильность гибридов, физиологическая и морфологическая несовместимость особей как половых партнеров и т. п.). В конце прошлого века М.Вагнер показал важную роль географической изоляции в процессах видообразования.
В 1926 г. была опубликована работа С.С.Четверикова "О некоторых моментах эволюционного процесса с точки зрения современной генетики", в которой была показана высокая генетическая гетерогенность природных популяций организмов, их насыщенность разнообразными мутациями. При этом многочисленные рецессивные мутации не проявляются фенотипически -фенотипы особей в популяции зачастую различаются меньше, чем их генотипы. Четвериков подчеркивал, что фенотипические проявления активности любого отдельного гена определяются его взаимодействиями с другими генами в целостном генотипе, представляющем своего рода "генетическую среду". Генетическая гетерогенность популяции является основой для ее эволюционных преобразований. Работы С.С. Четверикова заложили основы попу-ляционной генетики.
Интеграция дарвинизма и генетики произошла в 30-е гг. XX в. Основными вехами этого процесса принято считать работы Р.Фишера, С.Райта, Н.И.Вавилова, Н.П.Дубинина, Д.Холдейна, Д.Хаксли и др. В 40-е гг. синтез данных и выводов различных биологических наук (прежде всего - генетики, систематики, экологии, биогеографии) привел к формированию обобщающей концепции, которую в 1942 г. Д.Хаксли назвал синтетической теорией эволюции. Основным ядром этой концепции стала теория микроэволюции, представляющая собой анализ эволюционных преобразований популяций и процессов видообразования. Развитие этой теории связано с работами Т.Добжанского, Э.Майра, Б.Ренша, Н.В.Тимофеева-Ресовского и др. В последующие годы синтетическая теория эволюции в определенной степени интегрировала также данные эволюционной морфологии (в первую очередь палеонтологии, в которой особо следует выделить работы Д.Симп-сона), однако макроэволюционные исследования в основном остались все же за рамками синтетической теории эволюции. Важнейшим вкладом в развитие этой области стали основополагающие труды А.Н.Северцова и И.И.Шмальгаузена, положившие начало системному анализу эволюционного процесса.
В последние десятилетия стремительно развиваются исследования эволюции на молекулярно-биологическом уровне, где были получены важные результаты, требующие осмысливания с позиций общей теории эволюции.
Подводя краткие итоги, отметим основные достижения современной эволюционистики по отношению к классической теории Ч.Дарвина.
В области микроэволюции (разрабатывавшейся в рамках синтетической теории эволюции, иногда называемой также "неодарвинизмом"):
1) выяснена сущность изменчивости и наследственности организмов;
2) исследована природа биологического вида, показана его сложная популяционная структура, выяснена роль популяций в эволюционном процессе;
3) открыты новые факторы и механизмы эволюционного процесса (дрейф генов, полиплоидизация, гибридизация и др.);
4) получила дальнейшее развитие теория естественного отбора.
В области макроэволюции (анализируемой главным образом в русле формирующейся системной теории эволюции):
1) выяснена сущность макроэволюции и ее соотношения с элементарными эволюционными изменениями;
2) установлен ряд эмпирических закономерностей макрофилогенеза;
3 ) показана эволюционная роль преобразований онтогенеза;
4) проделан анализ причин направленности и неравномерности темпов макрофилогенеза;
5) выяснены сущность и причины прогрессивной эволюции.
Рассмотрение современной эволюционной теории мы начнем с проблем микроэволюции.
Необходимыми предпосылками эволюционного процесса являются возникновение элементарных изменений аппарата наследственности - мутаций, их распространение и закрепление в генофондах популяций организмов. Направленные изменения генофондов популяций под воздействием различных факторов представляют собой элементарные эволюционные изменения -микроэволюцию. Микроэволюция завершается формированием новых видов организмов (процесс видообразования). Рассмотрение предпосылок, структурных элементов, факторов и механизмов всех этапов микроэволюции является основным содержанием этой части книги.
Чтобы не нарушать последовательность изложения, в нее пришлось включить также ряд проблем, выходящих за рамки собственно микроэволюционных процессов. Это относится прежде всего к различным межвидовым отношениям, которые оказывают значительное влияние на процессы видообразования. Межвидовые отношения реализуются в природных сообществах разных видов организмов -- биоценозах, представляющих собой целостные биологические макросистемы с механизмами авторегуляции. Их преобразования могут иметь уже макроэволюцион-ное значение.
Заключительная глава этой части посвящена развитию приспособлений организмов. Эта проблема также занимает пограничное положение между микро- и макроэволюцией. Формирование приспособлений организмов (адаптациогенез) начинается на уровне микроэволюции, но общий приспособительный характер эволюционного процесса и ряд специфических проблем адаптациогенеза в полной мере проявляются лишь в масштабе макроэволюции.
Таким образом, разделы второй части, посвященные межвидовым отношениям и развитию приспособлений, предваряют переход к проблемам макроэволюции, которым будут посвящены третья и четвертая части книги. Соотношения микро- и макроэволюции как двух уровней системной организации эволюционного процесса и свойства разных категорий биологических систем будут рассмотрены в начале четвертой части.
Современные представления о наследственности организмов
Наследственность организмов, под которой понимается способность передавать от поколения к поколению основные структурные и функциональные свойства, обеспечивающие сходство организации потомков и их родителей, представляет собой одно из фундаментальных качеств живых организмов. Само по себе явление наследственности чрезвычайно давно известно людям, но сущность этого важнейшего свойства организмов стала понятной лишь в середине XX в., когда была доказана роль хромосомной дезоксирибонуклеиновой кислоты (ДНК) в передаче наследственных свойств, а в 1953 г. Дж. Уотсоном и Ф. Криком была расшифрована структура молекулы ДНК.
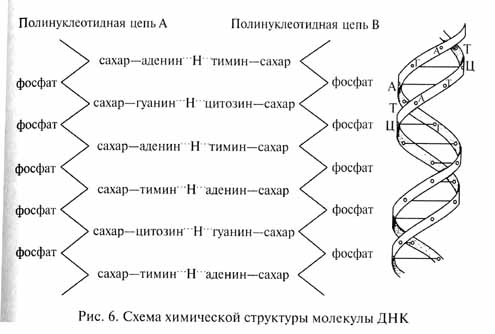
Хромосомы состоят из молекул ДНК, рибонуклеиновой кислоты (РНК) и некоторых типов белков. Основную роль в аппарате наследственности играет ДНК. Согласно модели Уотсона и Крика, молекула ДНК состоит из двух полинуклеотидных цепочек (рис. 6), спирально закрученных вокруг общей оси. Каждый отдельный нуклеотид включает молекулу (точнее, молекулярный остаток) сахара-циклопентозы (в ДНК - дезоксирибоза) и связанные с ней молекулярные остатки фосфорной кислоты и одного из азотистых оснований (пуриновых - аденина и гуанина, или пиримидиновых - тимина и цитозина). Нуклеотиды связаны друг с другом в полинуклеотидную цепочку, а две соседние полинуклеотидные цепочки связаны друг с другом в одну дву-спиральную молекулу ДНК водородными связями между пури-новыми и пиримидиновыми основаниями разных цепочек. При этом возможны только соединения аденин-тимин и гуанин-цитозин. Нуклеотиды с соответствующими парами оснований могут чередоваться в молекуле ДНК в любом порядке. Именно эта последовательность чередования разных пар азотистых оснований молекулы ДНК и представляет собою запись наследственной информации. При этом различные комбинации оснований в последовательных тройках (триплетах) нуклеотидов кодируют разные аминокислоты в молекулах полипептидов, синтезируемых в клетке. Отдельные гены представляют собой участки молекулы ДНК, обладающие определенной биохимической функцией (например, ответственные за синтез определенного типа белковых молекул).
Молекулы ДНК обладают способностью к редупликации, т.е. они могут удваиваться, причем новые молекулы в норме совершенно идентичны старой по строению и расположению всех нуклеотидов. На этом свойстве редупликации ДНК основана точная передача наследственных признаков от поколения к поколению.
Второе важнейшее биологическое свойство молекул ДНК - их способность контролировать синтез белков в клетке, причем, естественно, специфика молекул ДНК у данного вида организмов определяет специфику белкового синтеза у этого вида. Перс-дача информации с ДНК в цитоплазму для последующего синтеза белков на рибосомах осуществляется через посредство РНК -более простой одноцепочной нуклеиновой кислоты, в молекуле которой место тимина занимает другое пиримидиновое основание - урацил. Процесс синтеза молекул РНК на матрице ДНК (так называемая транскрипция) происходит при участии внутриклеточного фермента транскриптазы (или ДНК-зависимой РНК-полимеразы).
Точность редупликации молекул ДНК в процессах клеточного деления (митоза) обеспечивает полную генетическую эквивалентность всех клеток многоклеточного организма. Каждая клетка содержит полный геном, т.е. минимальный полный набор наследственных факторов, включающий 100% генетической информации. Однако в разных типах соматических клеток (например, эпителиальных, нервных, мышечных и т. п.) синтезируются совершенно определенные белки, специфичные для каждого типа клеток. Это обусловлено инактивацией (репрессией) большей части генов в хромосомах соматических клеток хромосомными белками (по одной из гипотез - белками-гистонами). Таким образом, специфичность белкового синтеза в клетках, образующих разные ткани и органы, определяется активностью различных участков хромосом.
Непосредственно кодируют последовательность аминокислот в молекулах полипептидов, синтезируемых на матрицах нуклеиновых кислот, далеко не все гены, а лишь некоторые из них, называемые структурными генами. Другие гены - регуляторы -контролируют процессы считывания наследственной информации со структурных генов, определяя синтез белков, репрессирующих те или иные структурные гены.
Новейшие молекулярно-генетические исследования показали значительную сложность организации генома. Оказалось, что в геномах большинства организмов содержится гораздо больше ДНК, чем необходимо для обеспечения биосинтеза всех белков данного организма. Значительная часть генома представлена так называемой "молчащей ДНК", не транслируемой в белки (трансляцией называется процесс синтеза полипептидных цепей белка по матрице информационной РНК; трансляция происходит на рибосомах). Она включает участки с многократными повторами одних и тех же последовательностей нуклеотидов. Кроме того, у эукариот (т. е. организмов, клетки которых имеют обособленное ядро, см. с. 149) гены обладают своего рода мозаичной структурой: транслируемые при синтезе белков участки (экзоны) чередуются с нетранслируемыми (нитронами).
С другой стороны, в геноме многих организмов были обнаружены так называемые мобильные генетические элементы - фрагменты молекул ДНК, которые могут перемещаться по геному, "встраиваясь" с помощью синтезируемых на их матрицах специальных ферментов в разные места хромосом и вызывая разнообразные наследственные изменения, подавляя или усиливая проявления активности других генов и способствуя повышению мутагенеза (см. ниже). Показано, что у эукариот мобильные генетические элементы составляют не менее 5-10% всего генетического материала.
Мобильные генетические элементы имеют различные размеры (порядка 1000-20000 нуклеотидов) и структуру. Наиболее крупные и сложные из них - плазмиды - способны не только интегрироваться в состав хромосом, образуя так называемые эписомы, но и самостоятельно функционировать в клетке. Полагают, что мобильные генетические элементы могут перемещаться из одной клетки в другую и даже переносить наследственную информацию между разными видами организмов (подробнее см. кн.: Xесин Р.Б. Непостоянство генома. - М., 1984). Подобный так называемый "горизонтальный" (или "латеральный") межвидовой перенос генетической информации, видимо, достаточно широко распространен у прокариот (низшие безъядерные организмы - бактерии, синезеленые водоросли); для эукариот хорошо доказанных данных такого переноса пока нет.
Помимо клеточного ядра специфическая ДНК локализована также в небольшом количестве в митохондриях и пластидах. По-видимому, ДНК этих органоидов обладает такими же основными свойствами, как и ядерная ДНК (способность к редупликации и контролю за синтезом определенных белков через посредство РНК), и представляет собой основу аппарата цитоплазматической наследственности (или плазмотипа), который дополняет основной, ядерный, наследственный аппарат -- генотип. Плазмотип состоит из относительно небольшого (30-300) количества плаз-могенов, кодирующих синтез некоторых белков, важных для функционирования соответствующих органоидов.
ДНК является основной, но, возможно, не единственной формой передачи наследственной информации. Дополнительными источниками информации, передающейся от клетки к клетке, могут быть некоторые надмолекулярные структуры (особенно система клеточных мембран). По мнению К. Маркерта и Г.Уршпрунга, они могут служить матрицами для сборки аналогичных структур из отдельных молекул. Вместе с ДНК митохондрий и пластид эти механизмы передачи наследственной информации входят в состав плазмотипа. В некоторых случаях плазмотип обеспечивает проявление так называемой "материнской наследственности", которая заключается в преобладании у гибридного организма материнских особенностей. Известный пример материнской наследственности - различия мулов (потомство от скрещивания жеребца с ослицей) и лошаков (потомство от скрещивания осла с лошадью). В подобных случаях преобладающее сходство потомства с матерью обеспечивается передачей наследственной информации цитоплазмой яйцеклетки (сперматозоид по сравнению с яйцеклеткой практически лишен цитоплазмы).
В принципе передача наследственных свойств плазмотипом вполне сходна с таковой, осуществляемой генотипом. Очевидно, цитоплазматическая наследственность не имеет никакого отношения к "адекватной соматической индукции" по той же самой причине, по которой фенотипические изменения, как таковые, не передаются следующим поколениям через генотип (см. выше).
Норма реакции, модификационная изменчивость и проблема обратной транскрипции
Необходимо подчеркнуть, что наследственный аппарат сам по себе кодирует только синтез специфических белковых молекул, а отнюдь не признаки целого сформированного фенотипа (т.е. конкретные особенности строения и функционирования тех или иных органов), как это иногда упрощенно представляют. Синтез специфических белков в зародышевых клетках и взаимодействие этих клеток друг с другом определяют общее направление морфогенетических процессов в ходе эмбрионального развития организма. Сущность явлений, лежащих в основе перехода наследственной информации с молекулярного уровня на клеточный в развивающемся организме, во многом еще не ясна. Однако вполне очевидно, что все морфогенетические процессы протекают в разных условиях несколько по-разному. Генотип программирует лишь общее направление морфогенетических процессов, а условия внешней среды оказывают существенное влияние на их конкретную реализацию. В результате каждый ген имеет несколько разных вариантов своего фенотипического проявления, т. е. разных состояний того признака или признаков, которые контролируются этим геном. Всякий конкретный генотип в разных условиях обеспечивает развитие разных фенотипов, т.е. разных совокупностей морфологических, физиологических и поведенческих признаков организма. Все эти фенотипы составляют норму реакции одного и того же генотипа на различные внешние условия. По удачной формулировке М.М.Камшилова, фенотипы наследственно обусловлены, но, как таковые, не наследуются: наследственна лишь норма реакции всякого конкретного генотипа на внешние воздействия.
Поскольку норма реакции обеспечивает формирование тех или иных фенотипов в соответствии с определенными условиями среды, она имеет огромное значение для выживания вида и, следовательно, подвергается действию естественного отбора. Поэтому фенотипические признаки, проявляющиеся у всякого организма в зависимости от условий его сушествования, обычно имеют черты "целесообразности", являясь приспособлениями к данным условиям.
Эти разные фенотипические проявления одних и тех же генов представляют собой изменчивость, но изменчивость, не являющуюся для вида чем-то новым, поскольку все ее варианты запрограммированы в норме реакции генотипа, чтобы обеспечить выживание организмов данного вида в разных условиях. Это и есть модификационная (или фенотипическая) изменчивость, на которой концентрируют внимание ламаркисты, принимающие ее за основу эволюционного процесса. Этой форме изменчивости соответствует определенная изменчивость в теории Дарвина. Из всего сказанного совершенно очевидно, что эта изменчивость хотя нередко и приобретает в результате отбора приспособительный характер, но не может сама по себе привести ни к каким эволюционным преобразованиям.
Если организм подвергается воздействию необычных для данного вида экстремальных условий, возникающие при этом модификации нередко теряют приспособительный характер, получая облик уродства; такие модификации называются морфозами.
Неоламаркисты пытались привлечь для аргументации своих позиций некоторые новые достижения молекулярной генетики, в частности - возможность обратной передачи наследственной информации - с РНК на ДНК. Эта так называемая обратная транскрипция происходит в специфических условиях инфицирования клеток эукариот особыми вирусами (онко-РНК-вирусы), у которых геном образован РНК. При действии вирусного фермента РНК-зависимой ДНК-полимеразы, или ревертазы, на матрице вирусной РНК синтезируется молекула ДНК, которая затем внедряется в геном инфицированной клетки. Сторонники неоламаркистских взглядов полагают, что этот механизм может быть использован Для объяснения адекватной соматической индукции. При этом не обращают внимания на крайнюю специфичность ситуации, при которой происходит обратная транскрипция - вирусная инфекция. Но даже если и допустить подобную возможность, это отнюдь не докажет желаемого для неоламаркистов унаследования "целесообразных" изменений соматических структур, возникших как приспособительные реакции на изменения внешних условий.
Вариации строения и функций соматических структур приобретают приспособительный характер только в результате эволюции под контролем естественного отбора. Все целесообразные приспособительные реакции фенотипа являются модификациями - и никакой механизм передачи наследственной информации не закрепит этих признаков просто потому, что они уже входят в норму реакции данного генотипа, не являясь, в сущности, новыми признаками.
Конечно, изменения внешних условий могут вызвать и такие нарушения в геноме соматических клеток, которые приведут к возникновению качественных новообразований - соматических мутаций. Однако эти изменения, как и все мутации, не являются приспособительными (целесообразными) -- их фенотипические проявления имеют случайный характер по отношению к факторам, вызвавшим их появление. Подробнее наследственную изменчивость мы рассмотрим в следующем разделе этой главы.
Мутационная изменчивость
Эволюционные преобразования организмов основываются на изменениях аппарата наследственности, приводящих к соответствующим перестройкам нормы реакции генотипа; такие изменения получили название мутаций. Мутации могут возникнуть (и неизбежно возникают в определенном проценте случаев) в процессе редупликации генома, предшествующем клеточному делению, или в процессе самого клеточного деления. Точные причины естественно происходящих (спонтанных) мутаций обычно неизвестны, но, очевидно, они кроются в каких-то нарушениях нормального режима сложных процессов редупликации и расхождения хромосом, связанных в конечном итоге с внешними воздействиями на клетку. Возникновение спонтанных мутации связывают, например, с мутагенной активностью некоторых метаболитов (продуктов обмена веществ) или с термическими флюктуаииями атомов в процессах редупликации и т. п.
Размножение и связанная с ним редупликация наследственного аппарата представляют собой важнейшие и неотъемлемые свойства живых организмов. Поскольку эти сложные процессы протекают на фоне постоянных изменений внешней среды и поскольку всякий механизм и всякая структура обладают конечной степенью устойчивости, нарушения процесса редупликации, ведущие к мутациям, совершенно неизбежны. Мутации представляют собой элементарные изменения, являющиеся материалом для действия естественного отбора. Неизбежный результат естественного отбора - эволюционные изменения организмов. Таким образом, как отметил Н.В.Тимофеев-Ресовский, неизбежность эволюции следует из основных свойств живой материи.
Хотя стимулом мутаций являются изменения внешних условий, между этими последними и происходящими под их влиянием мутациями нет адекватности. Это означает, что возникающие мутации сами по себе не являются приспособлениями к изменениям условий, вызвавшим их появление. Аппарат наследственности не обладает и принципиально не может обладать свойством целесообразно перестраиваться в ответ на изменения внешней среды. Это отчетливо проявляется при искусственном мутагенезе, когда внешние воздействия, вызвавшие возникновение мутаций, точно известны. Например, хорошо известно мутагенное действие проникающей радиации, но возникшие под ее влиянием мутации могут затрагивать самые различные особенности организмов (цвет глаз, степень развития и особенности строения крыльев у мух и т. п.), которые в подавляющем большинстве случаев никак не связаны с защитой организма от радиации. Точно так же действуют и другие, более "мягкие" мутагены (различные химические факторы, изменения температуры и др.), причем разные мутагены могут вызывать у данного вида организмов одни и те же мутации.
Этот факт важен в двух отношениях. С одной стороны, он подчеркивает неопределенный, неприспособительный характер мутаций, с другой - многократное появление у разных особей данного вида организмов одних и тех же мутаций. Из этого следует чрезвычайно важный вывод: всякий генотип имеет хотя и большие, но вполне конкретные возможности мутационных изменений, определенный спектр изменчивости. Одни и те же мутации с определенной регулярностью появляются в пределах популяций данного вида вновь и вновь. И с другой стороны, некоторые изменения, кажущиеся столь же вероятными, никогда у этого вида не наблюдаются. Например, среди различных видов мух-Дрозофил, вероятно наиболее изученных в генетическом отношении животных, никогда не наблюдались особи с зелеными или синими глазами. Поскольку генетиками изучено уже около двух миллиардов (!) мух, вероятность обнаружения указанных мутаций в будущем выглядит ничтожной. По-видимому, в геноме дрозофил нет каких-то предпосылок, необходимых для возникновения таких мутаций, хотя подобные окраски глаз нередко наблюдаются у других видов насекомых. Для дрозофил эти мутации являются по каким-то причинам невозможными ("запрещенными").
Мутации могут происходить на разных уровнях организации наследственного аппарата: на уровне полного хромосомного набора (генома) клетки, на уровне его морфологически обособленных частей -- хромосом и на уровне его структурно-функциональных единиц - генов.
Геномные мутации представляют собой изменения количества хромосом, которые могут происходить либо путем кратного увеличения числа целых (гаплоидных) наборов хромосом, либо посредством изменения числа отдельных хромосом в геноме, в результате чего общее число хромосом становится не кратным гаплоидному.
Увеличение числа целых хромосомных наборов (плоидности) в клетке называется полиплоидизацией, изменение числа отдельных хромосом в геноме - анеуплоидией, или гетероплоидией. Фенотипический эффект геномных мутаций основан на взаимодействии генов или их комплексов, расположенных в соответственных (гомологичных) хромосомах, число которых изменилось в результате данной мутации. Вероятно, это взаимодействие осуществляется через посредство продуктов белкового синтеза (ферментов) или РНК, транскрибированной с ДНК соответствующих хромосом.
Геномные мутации обычно приводят к стерильности мутантов в результате невозможности осуществления полового процесса с представителями родительской формы, сохранившими нормальное количество хромосом. Поэтому у животных относительно редко геномные мутации дают начало новым формам - это возможно только при способности данного вида размножаться бесполым способом, или посредством партеногенеза, или гиногенеза (гиногенез - способ размножения, при котором сперматозоид проникает в яйцеклетку и стимулирует ее развитие, но его ядро не сливается с ядром яйца и не участвует в развитии зародыша). Среди растений бесполое размножение распространено гораздо шире, чем среди животных. В связи с этим у растений полиплоидизация, вероятно, играла существенную роль в эволюционном процессе.
Обычно различают автополиплоидию и аллополиплоидию. Автополиплоиды образуются посредством увеличения числа хромосомных наборов у отдельных особей одного вида. У автополиплоидов каждая хромосома представлена одним и тем же числом гомологов, например в клетках тетраплоидов имеется по четыре гомологичные хромосомы, у октоплоидов - по восемь и т. д. Ал-лополиплоиды образуются посредством гибридизации организмов разных видов. У аллополиплоидов соответствующие хромосомы уже в определенной степени утратили свою гомологичность. Поэтому процесс мейоза, в котором осуществляется редукция числа хромосомных наборов перед половым процессом, у аллополиплоидов обычно нарушен. Аллополиплоиды могут сохраниться как новый вид, если способны к бесполому размножению.
Половой процесс может восстановиться у полиплоидной формы, когда в популяции накапливается достаточное количество полиплоидных особей. У аллополиплоидов, кроме того, необходимо еще дополнительное кратное удвоение числа хромосом (посредством эндомитоза в соматических клетках или нерасхождения хромосом при образовании гамет). (Эндомитоз - удвоение числа хромосом внутри ядра без образования веретена деления клетки). Тогда в аллополиплоидном наборе появляются гомологичные хромосомы, и в дальнейшем мейоз может протекать нормально. Известно несколько примеров возникновения новых форм растений этим путем; среди них можно упомянуть рафанобрассику - межродовой гибрид, полученный Г.Д.Карпеченко путем скрещивания редьки (Raphanus sativus) с капустой (Brassica oleracea).
Хромосомные мутации являются структурными перестройками отдельных хромосом, изменяющими расположение в них генов, фенотипическое проявление подобных мутаций зависит от изменений пространственной локализации генов в хромосомах (эффект положения). Выделяют несколько основных типов хромосомных мутаций: а) транслокация - взаимный обмен участками между двумя разными (не гомологичными друг другу) хромосомами; б) делеция - утрата хромосомой какого-либо ее участка; в) инверсия - поворот внутри хромосомы какого-либо ее участка на 180°; г) дупликация - удвоение тех или иных участков хромосомы; д) транспозиция - перестановка генов внутри хромосомы.
Среди указанных типов хромосомных мутаций особую роль играют дупликации, которые (так же как и полиплоидизация) позволяют увеличить количество генетического материала в клетках. Как подчеркнул С. Оно, возникновение избыточности генетического материала может играть важную роль в эволюционных перестройках генетической системы вида, открывая новые возможности для прогрессивной эволюции, в частности для развития качественных новообразований. Дупликации ведут к появлению в наследственном аппарате генов-дублеров, отвечающих за одну и ту же функцию. В таких дублерах могут в скрытой форме сохраняться и накапливаться новые генные мутации. С другой стороны, инверсии способствуют образованию устойчивых генных комплексов (групп сцепления), которые редко разрушаются кроссинговером при мейозе и обеспечивают совместное наследование различных признаков.
Наконец, генные или точковые мутации представляют собой изменения самих генов, т.е. элементарных функциональных единиц аппарата наследственности. Сущность генных мутаций заключается в изменении порядка пуриновых и пиримидиновых оснований в молекуле ДНК. Для возникновения одной точковой мутации достаточно изменения хотя бы одной пары оснований. Если такие мутации происходят в структурных генах, может измениться порядок считывания наследственной информации при транскрипции ("мутация рамки" - смешение рамки считывания наследственной информации), что приведет к изменению порядка аминокислот в полипептидной цепи, т. е. к синтезу нового типа белковых молекул. Мутации генов-регуляторов изменяют соотношения между репрессированными и активными структурными генами, что неизбежно влечет за собой изменения белковых комплексов, синтезируемых в различных типах соматических клеток, несмотря на оставшиеся неизменными структурные гены.
Генные мутации являются основным источником наследственной изменчивости организмов. В результате мутирования возникают различные варианты генов - аллели, обусловливающие разные состояния контролируемого данным геном признака (или признаков). Число аллельных состояний отдельных генов может быть очень большим (более ста).
Поскольку мутационная изменчивость неопределенна по отношению к характеру изменений внешней среды, вызвавших появление мутаций, подавляющее большинство возникших наследственных изменений не улучшают, а ухудшают приспособленность мутантных организмов, и тем в большей мере, чем больше масштаб фенотипического проявления мутации. Поэтому в среднем наибольшие шансы на сохранение имеют мутации с наименьшим фенотипическим выражением ("малые мутации"), которые и служат основным материалом эволюционных изменений.
Фенотипическое проявление действия генов
В гаплоидном состоянии клетка обладает лишь одним полным хромосомным набором, и каждый ген представлен каким-то одним аллелем. В этом случае любая мутация сразу проявляется фенотипически и сразу же подвергается действию отбора. Гаплоидия как состояние, обеспечивающее возможность в первом же поколении опробовать новые мутации фенотипически, убыстряет эволюционный процесс и повышает скорость развития новых приспособлений, но требует огромных потерь мутантных особей в каждом поколении, так как подавляющее большинство мутантов оказываются хуже приспособленными к существующим условиям. Поэтому гаплоидное состояние как основная форма существования вида возможно только у низших одноклеточных организмов, для которых характерны гигантская численность особей и огромные темпы размножения (бактерии, синезеленые водоросли, простейшие и др.). У таких организмов выбраковка непригодных мутантов не угрожает сохранению вида как целого, и в каждом поколении новые мутации сразу испытываются отбором. Это и обусловливает поразительную быстроту приспособления подобных организмов к изменениям условий (например, приспособления многих болезнетворных микроорганизмов к действию новых лекарственных препаратов).
Однако выгоды гаплоидного состояния не могут быть использованы более сложными многоклеточными организмами, популяции которых состоят из меньшего числа особей (численность многих видов позвоночных животных измеряется лишь тысячами особей), имеют длительный онтогенез и низкие темпы размножения. Большая выбраковка особей в каждом поколении для таких форм была бы гибельной. Вместе с тем наследственная изменчивость и неизбежна у любого вида организмов, и необходима для его выживания в изменяющихся условиях. Макроорганизмы нашли оптимальное решение этой проблемы в диплоидном состоянии, при котором в каждой соматической клетке имеется по два полных хромосомных набора, состоящих из гомологичных хромосом, которые несут гомологичные гены, отвечающие за одни и те же признаки. Наличие в обеих гомологичных хромосомах одинаковых аллелей какого-либо гена называется гомозиготным состоянием (по данному гену), наличие же разных аллелей - гетерозиготным состоянием. При гетерозиготности фенотипически проявляется лишь один из аллелей - доминантный, или же в фенотипе наблюдается особое состояние признака, отличающееся от гомозиготных фенотипов обоих аллелей (неполное доминирование). Благодаря таким соотношениям разных аллелей в диплоидном генотипе в гетерозиготном состоянии могут сохраняться в скрытой, не проявляющейся фенотипически (рецессивной) форме любые новые аллели. Так, новые мутации могут постепенно накапливаться в популяции данного вида в форме рецессивных аллелей, время от времени проявляясь фенотипически в гомозиготном состоянии и подвергаясь в этом случае действию отбора.
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но, как указано выше, вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что Доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к Доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вари ант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путем отбора малых мутаций как структурных генов, так и генов-модификаторов.
Так или иначе степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название "индустриальный меланизм"; подробнее см. в гл. 3).
Соотношение фенотипической изменчивости, входящей в норму реакции генотипа, с изменчивостью наследственной, вообще говоря, достаточно сложно. Это соотношение носит в известном смысле парадоксальный характер: наследуются изменения генотипа (мутации), но отбор не может непосредственно их контролировать - отбор имеет дело с фенотипами организмов, в каждом из которых наследственная основа осуществила лишь один из многих возможных вариантов своего выражения. Другими словами, генотипическая изменчивость проявляется и подвергается отбору только в форме фенотипических признаков. При этом важно подчеркнуть, что отбор, действуя на фенотипы организмов, способствует выживанию или же элиминирует целостные генотипы, обладающие определенной нормой реакции.
Взаимодействие различных генов в целостном генотипе существенно влияет на общую норму реакции генотипа. В зависимости от этого физиологическое проявление одного и того же гена в одних и тех же внешних условиях может быть различным у особей с разными генотипами, т.е. в разной генетической среде. Данный ген может проявляться как доминантный у одних особей и не проявляться фенотипически у других. Н.В.Тимофеев-Ресовский назвал это свойство пенетрантностью проявления гена. Пенетрантность мутантного гена измеряется процентным соотношением особей, имеющих мутантный фенотип, и общего количества особей-носителей данного аллеля в популяции. Если этот фенотип обнаруживается у всех носителей данного гена, говорят о полной (100%-ной) пенетрантности, если же лишь у некоторых особей - о неполной пенетрантности данного гена.
Пенетрантность генов зависит как от взаимодействия генов в генотипе, так и от внешних условий. При разных условиях у особей со сходным генотипом фенотипическое выражение данного гена может быть различно. Так, мутация дрозофилы "abnormal abdomen" (фенотипически проявляется в неправильном расположении полосок окраски на брюшке) обнаруживается в фенотипе только при выращивании личинок мух на влажном корме; при использовании сухого корма мутанты ничем не отличаются от нормальных мух.
Кроме того, может варьировать (даже при 100%-ной пенетрантности) степень фенотипического проявления гена. Признак может быть выражен сильнее или слабее, проявляться более или менее отчетливо в зависимости от внешних условий или от взаимодействия генов друг с другом. Изменчивость степени выраженности признака в фенотипе называют его экспрессивностью. Если данный признак сходно выражен у разных особей в данных условиях, говорят о постоянной экспрессивности контролирующего этот признак гена, если же степень выраженности признака варьирует - об изменчивой экспрессивности. Недавно было показано, что на экспрессию структурных генов могут оказывать влияние повторяющиеся нуклеотидные последовательности "молчащей ДНК", которая, таким образом, также воздействует на признаки фенотипа.
Р. Гольдшмидт обратил внимание на удивительный параллелизм между фенотипическими проявлениями модификационной и наследственной изменчивости. Одно и то же состояние фенотипических признаков у разных особей данного вида может сформироваться по-разному: у одних особей - как модификацион-ная вариация, проявляющаяся лишь в определенных условиях, у других - как единственно возможный вариант признака (т.е. для Данного аллеля со 100%-ной пенетрантностью и постоянной экспрессивностью). Если в популяции, где данный признак первоначально был одним из модификационных вариантов нормы реакции, появляется мутация, обусловливающая более высокую Пенетрантность и экспрессивность развития этого признака, возникает иллюзия "превращения" ненаследственной модификации в наследственный признак ("эффект Болдуина"), т. е. "унаследования приобретенного соматического признака" в духе ламаркизма. о Действительности здесь имеет место просто возникновение новой мутации (рис. 7) - либо структурного гена, либо гена-модификатора, влияющего на фенотипическое проявление данного признака.

Модификации, сходные с фенотипическим проявлением определенных мутаций, получили название фенокопии мутаций; соответствующие мутации, "дублирующие" модификации, называются генокопиями последних. Так, у плодовой мушки-дрозофилы мутация tetraptera, при которой вместо обычной пары крыльев развиваются две пары (рис. 8), является генокопией модификации-морфоза, возникающей при обработке ранних личиночных стадий эфиром или высокой температурой. Соответственно этот морфоз является фенокопией указанной мутации.

И.И.Шмальгаузен показал, что глубокий параллелизм фенотипического проявления наследственной и ненаследственной изменчивости (мутаций и модификаций) основан на общности путей их реализации в индивидуальном развитии. И мутации, и модификации проявляются в фенотипе как результат изменений процессов клеточного метаболизма (чаще всего - простых количественных изменений, затрагивающих сроки или интенсивность синтеза определенных белков, реже - синтез нового белка). Разница лишь в том, чем непосредственно "запускаются" перестройки онтогенеза - изменениями генома (мутациями) или внешними воздействиями (при возникновении модификаций).
Подчеркнем еще раз, что мутационная изменчивость неопределенна по отношению к характеру изменений внешней среды. Мутационная изменчивость входит в состав дарвиновского понятия неопределенной изменчивости. Однако последнее понятие шире: оно включает не только вновь возникающие мутации, но также индивидуальные вариации особей, которые определяются независимым распределением разных генов при образовании половых клеток (в процессе мейоза) и последующим их комбинированием в половом процессе, приводящим к возникновению огромного количества всевозможных комбинаций генов в генотипах разных особей. Число генов в генотипе данного вида, помноженное на число аллелей каждого гена, при независимом комбинировании большинства генов в мейозе и половом процессе обеспечивают колоссальное разнообразие особей в каждом поколении. У видов, размножающихся половым способом, практически каждая особь обладает неповторимой, только ей одной присущей комбинацией генов (за исключением однояйцевых близнецов), хотя те же самые гены, но в других комбинациях широко рассеяны у других особей того же вида. Эти индивидуальные различия особей, основанные на разных комбинациях многочисленных генов и их аллелей, получили название комбинативной изменчивости.
В дарвиновское понятие неопределенной изменчивости входят также некоторые модификационные изменения. Учитывая указанные выше сложные взаимоотношения разных форм изменчивости организмов, совершенно естественно, что Дарвин не мог дифференцировать эти формы, объединяя их под названием "неопределенная изменчивость", в соответствии с уровнем развития науки его времени.
Мутационный процесс
Частота мутаций может колебаться в широких пределах и зависит как от характера внешних воздействий, так и от особенностей генотипа. Конкретные формы жизни различных видов организмов (численность и характер динамики популяций, определяемые способом питания, особенностями размножения, продолжительностью жизни, врагами данного вида и т.п.) обусловливают в каждом отдельном случае определенный уровень частоты мутаций, оптимальный в данных условиях для выживания вида. Хотя сами по себе спонтанные мутации (как результат нарушений процессов редупликации ДНК в клетках, кроссинговера и расхождения хромосом при клеточном делении) могут возникнуть только под влиянием каких-то изменений внешних условий, но генотип может быть в различной степени устойчив к внешним воздействиям, и разные гены обладают разной мутабильностью. В этом смысле возможен контроль над мутированием - точнее, над большей или меньшей его частотой - со стороны самого генома. Доказано, что существуют специальные гены-мутаторы, влияющие на частоту мутирования других генов. Например, у мух-дрозофил описан ген-мутатор Hi. При гомозиготности по этому гену общая частота точковых мутаций и хромосомных инверсий возрастает примерно в 10 раз, при гетерозиготности - в 2-7 раз. Частота спонтанных мутаций может значительно возрастать при инсерциях мобильных генетических элементов, вызывающих высокому-табильное состояние генома. (Инсерция - вставка мобильного фрагмента ДНК в хромосому). Частота мутирования, как всякий признак, важный для сохранения вида в данных условиях, подвергается естественному отбору.
Общая частота мутаций обычно выражается в проценте половых клеток одного поколения, содержащих какую-либо мутацию. Этот показатель весьма изменчив и может варьировать от нескольких единиц до 20-30 % на поколение. Частота мутирования отдельных генов, разумеется, много ниже и составляет, например, у дрозофилы 0,001-0,004% при общей частоте мутаций до 25%, (табл. 1).
Важно подчеркнуть, что, поскольку генотипы разных особей одного вида состоят из гомологичных генов и одни и те же аллели, хотя и в разных комбинациях, широко рассеяны в популяциях, неизбежным результатом является неоднократное возникновение тождественных мутаций в потомстве разных особей данного вида. Всякая конкретная мутация характеризуется определенной частотой своего появления в данной популяции организмов. Совокупность всех реально возможных мутаций составляет спектр изменчивости данного генотипа и генофонда данной популяции как системы из всех составляющих его генотипов.
Таблица 1. Частота спонтанных мутаций разных генов у разных видов организмов (по М. Е.Лобашеву, 1967)
| Виды организмов | Прямая мутация | Число изученных гамет | Частота на 10000 гамет |
| Кукуруза | Wx - > wx | 1503744 | 0,000 |
| Рг -> рг | 647102 | 0,110 | |
| Sh -> sh | 2469285 | 0,012 | |
| Sll - > SLI | 1678731 | 0,024 | |
| Дрозофила | ct+ -> ct | 60000 | 1,5 |
| y+ - > у | 70000 | 0,29 | |
| w+ - > w | 70000 | 0,29 | |
| lz+ -> Iz | 70000 | 0,29 | |
| Человек | Гемофилия | 0,32 | |
| Хондроди-строфия | 0,427 | ||
| Ретино-бластома | 0,23 |
Возможны и обратные точковые мутации, которые обычно происходят с меньшей (иногда с такой же) частотой, что и "прямые" мутации. Частота обратных мутаций также является характеристикой спектра изменчивости видового или популяциейного генофонда.
В генофондах родственных видов сохраняются гомологичные гены (их количество в целом пропорционально степени родства). Поэтому в популяциях родственных видов могут появляться и гомологичные мутации, приводящие к сходным фенотипическим Результатам. Это явление было описано Н. И. Вавиловым в 1935г. как закон гомологических рядов в наследственной изменчивости. Согласно Н.И.Вавилову, "виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, Можно предвидеть нахождение параллельных форм у других видов и родов" (рис. 9).

Это обобщение подчеркивает определенную направленность мутационного процесса, обусловленную для каждого конкретного вида организмов исторически сложившейся организацией его генома. Интересно, что явление параллельной изменчивости было открыто еще Ч.Дарвином, подчеркивавшим, что "близко параллельные разновидности часто получаются от самостоятельных рас или даже от самостоятельных видов".
Молекулярная эволюция
В связи с накоплением данных об особенностях структуры ДНК у разных видов и о различиях строения ряда белков, имеющих сходные биохимические функции, М.Кимура, М.Кинг и Т.Джукс в 60-е гг. выдвинули предположение о возможности эволюции без участия естественного отбора, или так называемой нейтральной ("недарвиновской") эволюции. Сущность этой гипотезы сводится к тому, что многие из возможных изменений структуры ДНК адаптивно нейтральны, т.е. не имеют никакой приспособительной ценности (ни положительной, ни отрицательной) для организмов. Поэтому сохранение этих вариантов никак не зависит от естественного отбора.
Гипотеза "нейтральной эволюции" основана на следующих данных. Во-первых, известно, что генетический код имеет так называемый вырожденный характер. Это означает, что в процессах транскрипции и трансляции несколько разных триплетов азотистых оснований в нуклеотидах кодируют одну и ту же аминокислоту. Например, синтез аминокислоты аланина в равной мере кодируется триплетами гуанин - цитозин-урацил (ГЦУ), гуанин- цитозин -аденин (ГЦА), а также ГЦЦ и ГЦГ. Точковые мутации, преобразующие один из этих триплетов в другой, не могут привести к изменениям синтезируемых в клетке белков. С другой стороны, даже синтез разных аминокислот не всегда приводит к изменениям биохимической функции белков, в состав которых они входят: разные аминокислоты могут быть функционально эквивалентны. Это подтверждают данные о широком распространении в природных популяциях многих видов организмов полиморфизма (т.е. множественности форм) определенных белков, отвечающих за одну и ту же биохимическую функцию. По данным Ф.Аялы, в генофондах природных популяций различных видов организмов полиморфны 20-50% локусов. Наконец, нейтральными по отношению к отбору представляются и мутации, происходящие в "молчащей ДНК" (повторяющиеся последовательности нуклеотидов).
Нейтральные мутации любого рода могут постепенно накапливаться в геноме, приводя к изменению структуры белков без изменений их биохимических функций (синонимический, или случайный, дрейф). (Его неудачно называют также "генетическим дрейфом", что приводит к путанице с так называемым дрейфом генов в изолированных малых популяциях – см. гл.3). Повторяющиеся и нетранслируемые последовательности нуклеотидов могут подвергнуться и более крупным, чем точковые мутации, перестройкам - типа хромосомных мутаций (дупликации, транслокации) и неравному кроссинговеру. Такие перестройки были названы Г.Доувером "молекулярным Драйвом".
Логическое развитие представлений о нейтральной по отношению к отбору молекулярной эволюции привело к выводу о постоянной (в среднем) скорости нуклеотидных замещений и соответствующих изменений белковых молекул ("молекулярные часы"). В соответствии с концепцией молекулярных часов, определив скорость молекулярной эволюции, можно по степени молекулярных различий ("генетическое расстояние") между различными современными видами организмов установить время эволюционной дивергенции соответствующих филетических линий.
Однако попытки практического применения постулата о постоянстве хода молекулярных часов эволюции для анализа хода филогенеза организмов встретились с серьезными трудностями. Некоторые из полученных этим методом филогенетических выводов разительно противоречили сложившимся в науке представлениям, хорошо обоснованным традиционными методами филогенетики и имеющим надежную геологическую и палеонтологическую датировку. Например, среди грызунов дивергенция родов Крыса (Rattus) и Мышь (Mus), по данным молекулярной биологии, определена в интервале 17,5-67 млн лет назад (этот огромный разброс датировок, обусловленный применением разных методов, сам уже говорит против концепции молекулярных часов). По данным же палеонтологии, дивергенция этих родов произошла не далее чем 8-12 млн лет назад. В связи с этим высказывалось предположение, что расхождение филетических линий на молекулярном уровне происходит раньше, чем это проявляется на уровне организма, однако в любом случае трудно согласиться со столь значительными масштабами этого различия.
Затем было обнаружено, что молекулярные часы во многих случаях "идут" весьма неустойчиво: в разных филетических линиях скорость нуклеотидных замещений значительно различается. Например, среди млекопитающих эта скорость у грызунов оказалась в 2-4 раза выше, чем у копытных, и в 4-8 раз выше, чем у высших приматов. Отмечено замедление скорости молекулярной эволюции у гоминид по сравнению с другими видами приматов и т. п. С другой стороны, разные белки в одной и той же филетичес-кой линии эволюционируют с разной скоростью. Вероятно, скорость преобразований определенных белков также изменяется на разных этапах филогенеза.
Гипотеза нейтральной эволюции была подвергнута серьезной критике многими учеными (Ф.Аяла, B.C.Кирпичников и др.). Прежде всего было отмечено, что сторонники нейтрализма исходят из ошибочных представлений, что отбор действует на отдельные гены, тогда как в действительности отбор влияет на целостный генотип. Благодаря этому вероятно, что многие мутации, сами по себе кажущиеся нейтральными, в действительности таковыми не являются. Пусть, например, данная точковая мутация - изменение какой-либо пары оснований в молекуле ДНК - представляется нейтральной (либо из-за вырожденности генетического кода, в котором несколько разных триплетов оснований кодируют одну и ту же аминокислоту, либо благодаря тому, что изменение кода повлекло за собой включение в молекулу кодируемого белка такой новой аминокислоты, которая имеет ту же биохимическую функцию, что и прежняя аминокислота, и т. п.). Но даже такая мутация для целостного организма не может с полной уверенностью рассматриваться как нейтральная, поскольку любое изменение кода влечет за собой по крайней мере изменение типа транспортной РНК (тРНК), доставляющей аминокислоту на рибосомы для синтеза белка. Для подлинной нейтральности мутаций необходимо, чтобы разные типы тРНК и разные аминокислоты имелись в клетке в одинаковых количествах, а для синтеза каждого типа тРН К требовалось бы одинаковое количество энергии, поступающей в клетку извне. Такая ситуация чрезвычайно маловероятна. Различно содержание в клетке разных аминокислот, и также различно соотношение количества разных аминокислот в клетках различных тканей. Поэтому сомнительно, чтобы использование новой тРНК (пусть даже не приводящее к изменениям молекулы синтезируемого белка) или функционально нейтральная замена аминокислоты в молекуле белка никак не сказались бы на сложнейшем балансе внутриклеточных процессов, а следовательно - и на балансе межклеточных связей в морфогенезе.
Полиморфизм белков в природных популяциях различных видов может контролироваться отбором через полигенные системы, связанные с клеточной энергетикой, соотношением концентраций разных аминокислот и типов тРНК. В пользу этого говорят, например, результаты лабораторных экспериментов, в которых создавались искусственные популяции мух-дрозофил со случайным соотношением частот различных аллелей, контролирующих синтез некоторых полиморфных белков. Но спустя несколько поколений в этих популяциях устанавливались такие же соотношения частот этих аллелей, которые характерны для популяций мух в природе. Это указывает на неслучайный характер белкового полиморфизма, хотя приспособительный смысл соотношений различных аллелей пока остается неясным.
Новые данные о влиянии повторяющихся нуклеотидных последовательностей на экспрессию структурных генов показывают, что и мутации, происходящие в "молчащей ДНК", не являются нейтральными, поскольку они оказывают влияние на состояние фенотипических признаков. Это ставит под сомнение реальность "молекулярного драйва", основанного на аберрациях хромосом в области нуклеотидных повторов.
А.Г.Креславский полагает, что значительная часть вариаций, обычно рассматриваемых как адаптивно нейтральные, в действительности могут соответствовать так называемым псевдонейтральным мутациям. Их фенотипические проявления становятся при оптимальных внешних условиях адаптивно нейтральными благодаря действию различных регуляционных механизмов (морфоге-нетических, физиологических и поведенческих), нейтрализующих вредный эффект этих мутаций. Однако при неблагоприятных внешних условиях выявляется отрицательное воздействие таких мутаций на приспособленность. Псевдонейтральные мутации легко проникают в популяционный генофонд, и общая картина попу-ляционной изменчивости при благоприятных условиях создает ложное впечатление соответствия теории нейтральной эволюции.
Но еще более важно, что никакое нейтральное на молекулярном уровне изменение не может само по себе привести к развитию какого бы то ни было приспособления организмов к изменяющимся условиям внешней среды, поскольку по определению "нейтральные изменения" лишены всякой приспособительной ценности. Естественный отбор является единственным фактором, направляющим наследственные изменения организмов в русло приспособительной эволюции.
Разумеется, это не означает, что все эволюционные изменения (как молекулярные, так и макроструктурные) обязательно должны иметь приспособительный характер. Возможность неприспособительных, нейтральных по отношению к отбору изменений отмечал еще Ч.Дарвин. (Поэтому, кстати, не следует именовать нейтралистскую концепцию "недарвиновской" - тем более что ее основатель, М.Кимура, не отрицает естественный отбор, хотя пытается ограничить его роль лишь устранением неблагоприятных мутаций). Важно лишь не впадать ни в одну из двух крайностей: основой эволюции не может быть случайное комбинирование неприспособительных изменений, но нельзя полагать также, что абсолютно все эволюционные изменения приспособительны.
Решение парадокса видимой нейтральности многих молекулярных преобразований при очевидном приспособительном характере эволюции организмов в целом может быть связано с определенной степенью независимости эволюционных изменений разных уровней биологической организации. В индивидуальном развитии организма (онтогенезе) структуры высших уровней организации - сложные многокомпонентные органы - формируются в результате взаимодействий многочисленных разветвленных цепей морфогенетических процессов, которые запускаются разными структурными генами и контролируются генами-модификаторами. При этом многие элементарные изменения молекулярных структур (мутации) могут никак не проявляться в фенотипе, подавляясь "буферным" воздействием генов-модификаторов. Как мы уже подчеркивали, отбор непосредственно воздействует на изменения фенотипа и лишь опосредованно - на структуры более низких уровней организации; в наибольшей степени это относится к молекулярным структурам. Поэтому возможность нейтральных, неприспособительных изменений для молекулярных структур организма в целом существенно выше, чем для макроструктур фенотипа.
Представления о некоторой степени независимости эволюции на разных уровнях биологической организации получили дальнейшее подтверждение при исследованиях филогенетических взаимоотношений разных видов организмов методами молекулярной биологии (по структурным особенностям молекул ДНК и белков). Оказалось, что степень молекулярных различий между многими видами совершенно не соответствует их различиям по макроморфологическим признакам. Так, у млекопитающих в целом межвидовые макроморфологические различия велики при относительно незначительных молекулярных различиях, тогда как для земноводных и птиц характерны противоположные соотношения этих показателей. Эффектный пример подобного несоответствия представляют соотношения разных категорий признаков, характеризующих роды Человек (Homo) и Шимпанзе (Pan). Эти два рода высших приматов значительно отличаются друг от друга по анатомическим, физиологическим и поведенческим признакам. Однако, как показали М.Кинг и А.Вильсон, уровень молекулярных различий между ними не превышает степень различий между видами-двойниками у дрозофилы! Разные виды лягушек, принадлежащие к одному роду Rana, различаются в биохимическом отношении в 20-30 раз значительнее, чем роды Человек и Шимпанзе.
Возможность такой эволюционной независимости молекулярных и макроструктурных признаков определяется характером соотношений между генотипом и фенотипом: морфологические, физиологические и поведенческие изменения могут быть обусловлены не только изменениями структурных генов, но даже в большей мере - изменениями механизмов их фенотипического проявления. Генетически эти механизмы воплощены в генах-регуляторах и модификаторах, которые могут контролировать целые блоки структурных генов. Вероятно, у высших приматов, и вообще у млекопитающих, эволюция основывалась в большей мере На преобразованиях генов-регуляторов и модификаторов, чем структурных генов, тогда как у птиц и амфибий соотношения были обратными.
Таким образом, молекулярная эволюция может происходить быстрее, чем эволюция макроструктур, как бы "забуфериваясь" на промежуточном уровне морфогенеза действием генов-модификаторов, и наоборот - относительно незначительные изменения генетического кода, затронувшие гены-регуляторы и модификаторы, занимающие ключевые позиции, могут привести к существенным макроструктурным изменениям при сохранении относительно консервативной структуры макромолекул.
Мутации в генофондах популяций
Следует избегать крайне упрощенного представления, что эволюционный процесс сводится к появлению новых мутаций. Подлинные эволюционные изменения видов гораздо сложнее. Прежде всего, новая мутация должна войти в целостную систему генотипа, не разрушив сложнейших корреляций - взаимосвязей между разными частями развивающегося организма, обусловливающих нормальное протекание онтогенеза. Иногда говорят об отборе новых мутаций на жизнеспособность на разных уровнях - уровне генотипа, уровне клетки, уровне морфогенеза органов и уровне целостного организма. Термин "отбор" в этом аспекте, строго говоря, применять не следует, ибо отбор как таковой действует только в так называемых стохастических (вероятностных) системах, характеризующихся нежесткими связями между многочисленными взаимозаменяемыми элементами. С другой стороны, большинство организменных систем являются жестко интегрированными, или корреляционными. В таких системах изменение отдельных элементов обычно отражается на всей системе в целом. Поэтому правильнее говорить не об "отборе мутаций на уровне генотипа" как вероятностном процессе, а просто о выявлении жизнеспособности мутаций в процессе онтогенеза.
Наибольшие шансы на выживание имеют мутации малого масштаба, существенно не нарушающие интеграцию целостного организма и не производящие значительных изменений в фенотипе. Крупные мутации чаще всего детальны. Эти соотношения зафиксированы в так называемой "теореме Р.Фишера": вероятность вклада отдельной мутации в эволюцию обратно пропорциональная степени ее воздействия на фенотип. В результате этого сколько-нибудь существенные эволюционные преобразования организмов не достигаются посредством одной мутации. Всякого рода сложные структуры и функции организма контролируются не одним, а многими генами, и их эволюционная перестройка происходит посредством серии малых мутаций в таких полигенных системах.
Нужно подчеркнуть, что жизнеспособность мутантных особей в большой степени зависит от условий внешней среды. Например, мутация плодовых мушек Drosophila funebris "eversae" в культуре (в лабораторных условиях) повышает жизнеспособность, несмотря на ее фенотипические проявления - аномальное положение и форму крыльев, которые делают полет невозможным. В природе утрата способности к полету резко снижает жизнеспособность мутантных мух. С другой стороны, нетрудно представить себе ситуацию, при которой полет был бы невыгоден для насекомого. Вспомним, например, наблюдения Ч.Дарвина, отметившего, что на океанических островах, подверженных действию сильных ветров, многие виды насекомых утратили способность к полету, поскольку полет в этих условиях связан с опасностью заноса насекомого силой ветра далеко в океан. Можно предположить, что в таких условиях достоинства мутации, подобной "everase", проявятся в полной мере.
Громадное большинство мутаций не повышает, а понижает жизнеспособность мутантных организмов в сравнении с нормой или в лучшем случае являются нейтральными. Если данная мутация хотя бы не понижает жизнеспособность значительно, она входит в генофонд вида, сохраняясь у определенного процента особей, - это обогащает генофонд вида новым аллелем, но еще отнюдь не является эволюционным изменением. Появление в генофонде нового аллеля означает лишь возникновение нового варианта какого-то фенотипического признака, который обычно рецессивен и лишь время от времени будет проявляться в гомозиготном состоянии. При этом сам по себе этот новый признак лишен еще всякого приспособительного значения.
Микроэволюция представляет собой направленное изменение генофондов популяций с изменением частот встречаемости в последних различных аллелей. Такие изменения генофондов происходят в результате действия различных факторов, которые будут рассмотрены в главе 3. Важнейшим из них является естественный отбор. Именно естественный отбор из отдельных случайных мутаций формирует приспособления вида к изменяющимся условиям внешней среды, являясь основной творческой силой эволюционного процесса. В этом современные представления вполне соответствуют взглядам Дарвина.
Для выживания вида в изменяющихся условиях необходимо прежде всего, чтобы организмы имели возможность к ним приспосабливаться, т.е. обладали до некоторой степени лабильной организацией, которая могла бы несколько перестраиваться в ходе онтогенеза в соответствии с особенностями внешних условий. Это достигается фенотипической (модификационной) изменчивостью на основе нормы реакции генотипов отдельных особей. Однако норма реакции всякого генотипа имеет пределы своих приспособительных возможностей. Для выживания вида при более значительных изменениях внешней среды необходима перестройка нормы реакции, которая возможна только благодаря изменениям генотипа - мутациям. Мутационная изменчивость имеет случайный (по отношению к изменениям внешних условий) характер, поэтому, чтобы вид был в какой-то мере подготовлен к изменениям внешних условий, необходимо наличие некоторого запаса изменчивости (так сказать, запаса готовых вариантов). Этот запас изменчивости накапливается в генофонде вида за счет мутаций. Различные аллели разных генов образуют всевозможные комбинации в отдельных генотипах. Перекомбинирование генов осуществляется при кроссинговере и независимом расхождении разных пар хромосом во время мейоза, а также в половом процессе. Так возникает комбинативная изменчивость вида. Разные варианты фенотипов, возникших на основе разных комбинаций генов, подвергаются действию естественного отбора. Можно сказать, что генофонд всякого вида обладает определенным резервом комбинационной изменчивости, постоянно пополняемым новыми мутациями и являющимся необходимой предпосылкой для выживания вида в изменяющейся внешней среде.
Вид как биологическая макросистема
Подавляющее большинство организмов в той или иной форме используют половой процесс как важнейший способ обмена генетической информацией между разными особями. Половой процесс не обязательно связан с размножением (например, конъюгация инфузорий или меромиксис бактерий), но всегда объединяет различных особей в единую систему - биологический вид. (Меромиксис - половой процесс у бактерий, при котором происходит конъюгация двух бактериальных клеток и часть хромосомы одной из них переходит в другую). Благодаря половому процессу и мейозу в пределах вида постоянно возникают новые комбинации наследственных признаков, новые генотипы, которые все вместе составляют генофонд вида как целого. Биологический вид, состоящий из многочисленных особей, генетически родственных, но различающихся по своим комбинациям наследственных признаков, представляет собой целостную биологическую макросистему.
Поскольку вид является стохастической (вероятностной) системой, характеризующейся нежесткими связями составляющих его компонентов - особей, его устойчивость к изменениям внешней среды определяется в каждом конкретном случае наиболее удачными комбинациями признаков в генотипах отдельных особей. Этим вид отличается от жестких (корреляционных) систем, подобных отдельным организмам, устойчивость которых определяется наименее устойчивыми в данных условиях компонентами. Так, к гибели данного организма ведет летальная мутация в каком-то одном локусе, т.е. изменение одного гена, тогда как для вида в целом гибель отдельных особей, оказавшихся хуже приспособленными в данных условиях или погибших от случайных причин, является относительно малосущественным фактором. Высокая устойчивость биологического вида к изменениям внешней среды определила его роль как основной формы организации живой материи.
Вид, объединяемый в единое целое с помощью полового процесса, неизмеримо устойчивее к изменениям среды, чем клон - потомство данной особи, полученное посредством бесполого размножения. К организмам, размножающимся исключительно бесполым способом (деление, почкование и т. п.), понятие биологического вида, в сущности, неприменимо. Систематики для удобства описаний таких форм иногда используют термин "агамный вид". Аналогично формы организмов, размножающиеся исключительно путем партеногенеза, называют партеновидами.
В предыдущей главе мы уже рассматривали значение разных форм изменчивости для выживания вида, подчеркнув особую роль резерва комбинативной изменчивости, пополняемого новыми мутациями. Благодаря действию естественного отбора генофонд вида представляет собой не случайный набор аллелей, а определенным образом сбалансированную систему, в которой относительное количество тех или иных аллелей определяется приспособительной ценностью контролируемых ими фенотипических особенностей в данных условиях.
На основе генетической общности регулярно скрещивающихся друг с другом особей формируется их морфологическое, физиологическое и поведенческое сходство, на которое прежде всего обращали внимание систематики, выделяя разные виды организмов именно по критериям морфофизиологических (а в большинстве случаев только морфологических) сходств и различий.
Хотя виды организмов представляют собой реальные биологические системы, дать четкое и исчерпывающее определение понятию "биологический вид" оказалось нелегко. Причины этого коренятся в самой природе этой формы организации живой материи.
Долгое время вид определяли как совокупность морфологически и физиологически сходных особей, способных скрещиваться друг с другом, давая плодовитое потомство. Для практических целей такое определение было удобно, поскольку, казалось, давало вполне отчетливые критерии для идентификации разных видов. Однако по мере накопления новых данных систематики встречались с существенными трудностями в реальном применении указанных критериев.
Прежде всего в таком определении совершенно игнорируется сложная внутренняя структура вида. Уже во времена Дарвина было известно, что многие виды организмов включают устойчивые формы, обитающие в разных частях видового ареала (области географического распространения), - подвиды. Подвиды стабильны и достаточно отчетливо различаются по морфофизиологическим критериям, но в зонах контакта скрещиваются друг с другом, давая плодовитое потомство с промежуточным состоянием признаков (так называемые "зоны интерградации" между подвидами). Ч.Дарвин, изучая изменчивость организмов, пришел к выводу, что индивидуальные различия особей по степени своего выражения плавно переходят в различия подвидов, а различия подвидов столь же постепенно переходят в видовые различия. На основании этого Дарвин заключил, что между видами, подвидами и разновидностями нет принципиальных различий. Этот вывод Дарвина, если понимать его буквально, неверен, поскольку он игнорирует особую биологическую роль вида как формы организации живой материи и как целостной эволюционной единицы. При оценке таксономического статуса многих близких форм организмов возникают сложности - считать ли их подвидами одного вида или самостоятельными, хотя и родственными друг другу, видами.
Популяционная структура вида
Вид представляет собой в действительности гораздо более сложную систему, чем просто совокупность скрещивающихся сходных друг с другом особей. Он распадается на более мелкие естественные группировки особей - популяции, представляющие население отдельных относительно небольших участков в пределах всей зоны распространения (ареала) данного вида. Внутри каждой популяции имеет место наибольшая степень панмиксии; скрешива-ние же особей, происходящих из разных популяций, имеет место относительно более редко, и обмен генетической информацией между разными популяциями более ограничен. Это обусловливает определенную самостоятельность генетических процессов, происходящих в разных популяциях одного вида. В результате каждая популяция характеризуется своим специфическим генофондом с присушим только данной популяции соотношением частот встречаемости разных аллелей и с соответствующими особенностями спектра изменчивости. Эти генетические различия популяций могут иметь как случайный, так и неслучайный характер. Последнее свойственно относительно крупным популяциям (порядка 500 особей и более), длительное время существующим в данном географическом районе.
Природные условия в разных частях ареала вида обычно более или менее различны. В результате отбор имеет разное направление для популяций одного вида, населяющих разные районы. Следствием этого является возникновение относительно устойчивых различий генофондов разных популяций. Особенности популяци-онных генофондов благодаря действию отбора приобретают приспособительный характер: подбор аллелей в конкретном генофонде, обусловливающий специфическую картину комбинативной и модификационной изменчивости в данной популяции, становится оптимальным для условий обитания этой популяции.
Частота встречаемости разных аллелей в популяции определяется частотой прямых и обратных мутаций, давлением отбора, обменом наследственной информацией с другими популяциями в результате эмиграции и иммиграции особей. При относительной устойчивости условий в достаточно большой популяции все указанные процессы приходят к состоянию относительного равновесия, конкретный характер которого определяется, с одной стороны, спецификой условий, а с другой - генетической системой данного вида. В итоге такие достаточно крупные и устойчивые популяции приобретают сбалансированный и оптимизированный отбором генофонд, особенности которого имеют приспособительный характер и обусловливают специфические черты данной популяции (экологические, поведенческие, а во многих случаях и достаточно определенные морфофизиологические показатели).
Различия популяций, населяющих удаленные или относительно изолированные друг от друга области, становятся более отчетливыми в связи с уменьшением обмена генетической информацией между ними. Результатом достаточно продолжительной изоляции является формирование подвидов, под которыми понимаются популяции данного вида, населяющие разные участки видового ареала (т. е. имеющие аллопатрическое распространение) и характеризующиеся устойчивым комплексом морфологических, физиологических и экологических признаков, закрепленных наследственно. Однако подвиды полностью сохраняют скрещиваемость друг с другом, и если контакт между ними вновь расширяется, возникает зона интерградации, в которой в результате гибридизации особи имеют промежуточное состояние признаков. Наличие в пределах вида нескольких устойчиво различающихся друг от друга подвидо-вых форм обозначается термином политипичность вида.
Если ареалы отдельных подвидов достаточно велики, подвиды распадаются на популяции меньшего масштаба - экологи различают несколько уровней таких территориальных (аллопатрических) группировок. Таким образам, внутри вида существует сложная иерархическая система территориальных популяций, которая является приспособлением к оптимальному использованию всего разнообразия условий в разных районах видового ареала.
Поскольку популяции обладают специфическим генофондом, находящимся под контролем естественного отбора, очевидно, что эти естественные группировки особей должны играть важнейшую роль в эволюционных преобразованиях вида. Все процессы, ведущие к каким бы то ни было изменениям вида - к его разделению на дочерние виды (видообразование) или к направленному изменению всего вида в целом (филетическая эволюция), начинаются на уровне видовых популяций. Эти процессы преобразований по-пуляционных генофондов принято называть микроэволюцией. По определению Н.В.Тимофеева-Ресовского, Н.Н.Воронцова и А. В.Яблокова, популяции представляют собой элементарные структурные единицы эволюционного процесса, а векторизованные (направленные) изменения генофондов популяций -- элементарные эволюционные явления.
Вид в пространстве
С учетом внутривидовой популяционной организации Э. Майр определил биологический вид как группу фактически или потенциально скрещивающихся естественных популяций, которая физиологически изолирована от других подобных групп (т.е. от других видов). Следует отмстить, что и это определение не является исчерпывающим, оставляя неопределенность в двух отношениях.
Прежде всего, если две популяции в рассматриваемый момент времени разделены каким-либо географическим барьером, непроходимым для данного вида организмов (горный хребет, морской пролив, пустыня, для водных форм -- суша и т.п.), очевидно, они не обмениваются генетической информацией, хотя еще могут сохранять потенциальную способность к скрещиванию (реально не используемую). Следует ли относить такие популяции к одному виду?
Эта неопределенность совершенно неизбежна, так как она является отражением действительной ситуации, нередко наблюдаемой в природе. Многие виды организмов имеют разорванный, прерывистый ареал, распадаясь на группы изолированных друг от друга популяций. Можно назвать хотя бы такие классические примеры разорванного ареала, как распространение голубой сороки (Cyanopica cyana), встречающейся на Пиренейском полуострове и на Дальнем Востоке, или ареал вьюна (Misgurnus fossilis), обитающего в пресных водоемах значительной части Европы и Юго-Восточной Азии, но отсутствующего на территории между 70 и 90° восточной долготы. Дальнейшая судьба ныне существующих популяций любого вида организмов неизвестна - пойдет ли их обособление дальше, или же барьеры, разделяющие популяции, нарушатся, и восстановится обмен генами между разными популяциями. В первом случае изолированные популяции могут дать начало новым самостоятельным видам, во втором они вновь объединяются в один вид. Разумеется, последнее вероятнее при значительно меньшей степени разрыва ареала, чем в двух приведенных примерах, если изолированные в данный момент популяции географически не столь удалены друг от друга. Так или иначе судьба отдельных популяций зависит в первую очередь от дальнейших изменений среды, которые, как правило, непредсказуемы. Слияние популяций, обособленных в течение более или менее длительного времени, с другими популяциями вида придает всякому эволюционному стволу своеобразный "сетчатый" характер (рис. 10).

В некоторых случаях, особенно у видов с достаточно большим ареалом, может возникнуть еще более любопытная ситуация, которую хорошо иллюстрирует следующий пример. Большая синица (Parus major) широко распространена в Евразии - от Ирландии до Японии и Индонезии (рис. 11). В пределах этого огромного ареала орнитологи выделяют свыше 30 подвидов большой синицы. Все подвиды, соседствующие друг с другом, легко скрещиваются с образованием зон интерградации. Расселение большой синицы на восток осуществлялось, по-видимому, по двум направлениям: северному (леса умеренного пояса, к северу от зоны пустынь и гор Центральной Азии) и южному (Персия, тропические леса Индии и Индокитая). На Амуре встретились две формы большой синицы, проникшие сюда по северному и южному путям расселения: подвид Parus major major (распространенный от Западной Европы до Дальнего Востока) и подвид P. major minor. Эти две формы не скрещиваются друг с другом, т.е. ведут себя как самостоятельные виды. Однако P. m. minor легко скрещивается с более Южными подвидами большой синицы, а те, в свою очередь, - с подвидами, обитающими в Индии и Персии; последние формы легко скрещиваются с европейскими P. m. major. Как же приложить определение биологического вида к этой причудливой картине родственных форм? С одной стороны, P. m. major и Р. m. Minor по их нескрещиваемости друг с другом можно оценить как два разных вида; с другой стороны, они связаны через цепь легко скрещивающихся южных популяций, по которым в принципе возможна передача генетической информации от одного конца цепи до другого. Подобные ситуации встречаются не столь уж редко, как можно было бы подумать. Б. Ренш предложил обозначать такую цепь родственных аллопатрических форм термином "круг рас", а если некоторые формы в этом "круге" заслуживают выделения в качестве самостоятельных видов - "кольцевой вид". Позднее Э.Майр назвал такие аллопатрические родственные формы, которые на основании имеющихся данных трудно определить как подвиды или самостоятельные виды, "полувидами", а группу родственных полувидов (т.е. кольцевой вид Б.Ренша) - "надвидом".

Вид во времени
Вторая неопределенность в концепции биологического вида выявляется при попытке приложить определение Майра к популяциям, разделенным не географически, а во времени, т.е. к популяциям, связанным друг с другом родством не по горизонтали, а по вертикали, как предки с потомками. За истекший промежуток времени вид может существенно измениться. Так как изменения генофонда постепенны, невозможно провести отчетливую границу между предковым видом и любым из видов-потомков, если известен весь эволюционный ствол. Эта проблема неизбежно встает перед палеонтологами при изучении групп организмов, Достаточно полно представленных в палеонтологической летописи. Выход можно отчасти найти, используя ряд дополнительных терминов. Непрерывный ряд последовательных во времени популяций, каждая из которых является потомком предшествующей и предком последующей, называют филетической линией. Случайные отрезки этой последней, обычно известные палеонтологам, представляют собой палеонтологические виды (см. рис. 10). Наконец, биологический вид - это отдельная филетическая линия, рассматриваемая в данный момент времени, включая все живущие одновременно популяции, соответствующие определению Э. Майра. Биологический вид является, так сказать, "срезом" филетической линии плоскостью времени, или филетической линией, рассматриваемой в течение кванта времени (А.Кейн).
Д. Симпсон предложил для наименования отдельной филетической линии особый термин "эволюционный вид", дополнив определение филетической линии указанием на ее эволюционную независимость от других линий, собственную эволюционную роль и определенную тенденцию эволюционных изменений. Эта концепция подчеркивает эволюционную целостность вида и может быть использована в анализе филетических линий, переплетающихся друг с другом или расходящихся как независимые ветви.
Приспособленность и приспособляемость вида
Как уже отмечалось, внутривидовая популяционная организация является приспособлением к оптимальному использованию всего разнообразия условий в пределах видового ареала. Приспособленность всякой популяции к конкретным условиям ее обитания определяется специфическим подбором аллелей в ее генофонде и в результате этого - соответствующей нормой реакции особей, обусловливающей определенные фенотипические особенности. Различия популяций выражаются прежде всего в характере поведенческих реакций особей, совокупность которых определяет такие важные экологические характеристики популяции, как особенности питания, плотность населения, возрастная и половая структура, сезонные реакции, динамика численности и др.
Действие отбора придает указанным особенностям популяций черты целесообразности. Благодаря отбору одновременно с приспособленностью к существующим условиям популяции приобретают также определенную степень приспособляемости, т.е. способность выдержать разного рода изменения условий. Приспособляемость популяций определяется, с одной стороны, широтой нормы реакции составляющих популяцию генотипов (т.е. возможностями их модификационной изменчивости), с другой же стороны - генетической гетерогенностью, разнообразием аллелей в пределах популяционного генофонда (т.е. запасом комбинативной изменчивости). Важность этого мобилизационного резерва изменчивости для выживания популяций и эволюционную необходимость высокой степени генетической гетерогенности природных популяций показал С.С.Четвериков. При изменениях условий, выходящих за рамки нормы реакции генотипов, которые были оптимально приспособлены к прежним условиям, популяция сможет сохраниться только при наличии в ее генофонде таких генотипов, которые окажутся жизнеспособными и в новых условиях. Следовательно, отбор благоприятствует поддержанию генетической гетерогенности популяций. Запас комбинативной изменчивости включает наряду с оптимальными в данных условиях аллелями также относительно слабо приспособленные аллели. Последние снижают общую приспособленность популяции и составляют так называемый генетический груз, который, по образному выражению Д.Холдейна, является той ценой, которую вынуждена платить популяция за право эволюционировать и, добавим, чтобы выжить и сохраниться во времени. Благодаря этому приспособляемость популяции, которой благоприятствует генетическая гетерогенность и, как следствие, генетический груз, до известной степени противостоит приспособленности популяции к существующим условиям, основанной на оптимальных аллелях. Таким образом, в природных популяциях должно существовать некоторое равновесие между этими двумя противоположными тенденциями.
По определению Т. Мёллера, генетический груз представляет собой ту часть популяционного генофонда, которая определяет появление менее приспособленных особей, подвергающихся избирательной выбраковке (элиминации) под воздействием естественного отбора. Общий генетический груз складывается из нескольких составляющих: 1) мутационный груз -- неудачные мутации, существенно снижающие приспособленность; 2) сегрегационный груз - выщепляющиеся в каждом поколении слабо приспособленные гомозиготные особи по таким аллелям, которые в гетерозиготном состоянии обладают высокой приспособленностью (так называемое "сверхдоминирование", см. ниже); 3) субституционный груз - результат изменений приспособленности фенотипи-Ческих признаков при изменениях внешних условий или появлении новых, более удачных фенотипических вариантов.
Эволюционные преобразования вида под воздействием отбора Неизбежно сопровождаются гибелью хуже приспособленных особей. Общее число отбракованных отбором особей при замещении одного аллеля другим Д.Холдейн назвал "платой за отбор". По Расчетам Холдейна, плата за отбор при одновременном эволюционном замещении нескольких независимых аллелей (разных генов) оказывается чрезвычайно высокой: она экспоненциально возрастает с увеличением числа независимых генов, подвергающихся отбору. Из этого был сделан парадоксальный вывод о невозможности быстрых эволюционных преобразований, основанных на одновременном замещении многих независимых аллелей - которые, однако, реально происходили в филогенезе многих групп организмов. Этот парадокс решается, если учесть, что отбор действует, как уже подчеркивалось, не на отдельные гены, а на фенотипы организмов, формирующиеся в онтогенезе на основе взаимодействий целостных генетических систем ("супергенов") в целостном генотипе. В результате отбору подвергаются не отдельные гены, а полигенные комплексы (супергены), и эволюционные преобразования затрагивают сопряженные комплексы фенотипических признаков. Это и определяет возможность высокой скорости эволюции без гибельных последствий для вида в результате слишком высокой платы за отбор (подробнее см. кн.: Грант В. Эволюция организмов. - М, 1980).
Полиморфизм вида
Существующие в данный исторический момент условия среды могут благоприятствовать либо какому-то одному фенотипу, либо сразу нескольким разным фенотипам. В первом случае в природных популяциях преобладает один доминантный фенотип, называемый "диким типом", который скрывает в гетерозиготном состоянии многочисленные рецессивные аллели (таков, например, хорошо известный фенотип окраски домовой мыши). Во втором же случае в одной популяции постоянно встречаются несколько устойчивых, отчетливо различающихся друг от друга форм. Такое состояние называется полиморфизмом популяции (вида).
Очевидно, полиморфизм позволяет популяции лучше использовать разнообразие условий, существующих в пределах ее ареала аналогично тому, как дифференциация аллопатрических популяций позволяет виду лучше использовать разнообразие условий в пределах видового ареала. Полиморфизм повышает одновременно и приспособленность, и приспособляемость популяции, поскольку он всегда основывается на генетической гетерогенности, на существовании в популяции нескольких устойчивых генных комплексов.
Полиморфизм может и не иметь отчетливого приспособительного характера, основываясь на адаптивной равноценности рач-ных вариантов (см. в гл. 1 о полиморфизме белковых комплексов). Но здесь нас интересует в первую очередь адаптивный, сбалансированный отбором полиморфизм. Такой полиморфизм в популяции возникает в следующих основных случаях: 1) отбор действует против фенотипа, обусловленного гомозиготным состоянием определенного аллеля, но благоприятствует гетерозиготному фенотипу, обладающему повышенной жизнеспособностью ("сверхдоминирование" - гетерозиготы превосходят по жизнеспособности оба вида гомозигот); 2) на различных стадиях онтогенеза отбор благоприятствует фенотипическим особенностям, обусловленным разными аллелями; 3) отбор благоприятствует сохранению данного аллеля у особей одного пола, но действует против него у особей другого пола; 4) в разные сезоны года более приспособленными оказываются фенотипы, обусловленные разными аллелями; 5) фенотипы, обусловленные разными аллелями, используют различные условия в пределах одного ареала; 6) отбор благоприятствует сохранению данного фенотипа, когда этот последний редок, и направлен против него, когда он встречается часто.
Явления возрастного полиморфизма и полового диморфизма (вторая и третья формы полиморфизма в этом перечне) хорошо известны. Остальные варианты рассмотрим подробнее. Интересным примером "сверхдоминирования" является полиморфизм некоторых восточноафриканских популяций человека по типам гемоглобина. В этих популяциях у взрослых людей кроме обычного типа гемоглобина А наблюдается также особая форма гемоглобина S, которая отличается от типа А тем, что в одной из его молекулярных цепей место глютаминовой аминокислоты занимает валин. Нормальный аллель НbА встречается с частотой около 80%, мутантный аллель Hbs - около 20%. У гетерозигот НbА Hbs имеются обе формы гемоглобина, а у гомозигот HbsHbs почти весь гемоглобин крови представлен формой S. В этом последнем случае у людей развивается тяжелое заболевание - серповидно-клеточная анемия (эритроциты сморщиваются, принимая форму серпа), часто заканчивающаяся смертью. При таком исходе следовало бы ожидать гораздо меньшей частоты встречаемости мутантного аллеля, чем реально наблюдаемые 20%. Оказалось, что полиморфизм популяций по указанным аллелям связан с повышенной устойчивостью гетерозигот HbAHbs к малярии: количество паразитов в крови гетерозиготных индивидов много ниже, и продолжительность болезни короче, чем у гомозигот НbAНbА. Поскольку малярия является опасным заболеванием, вызывающим значительную смертность среди детей в Восточной Африке, очевидно, что распространение мутантного аллеля Hbs в человеческих популяциях в определенной концентрации будет выгодно, несмотря на опасность смертности гомозигот HbsHbs от анемии. В результате аллель Hbs имеет повышенную концентрацию именно в тех районах, где наиболее распространена малярия.
Наземные улитки Cepaea nemoralis в зоне смешанных лесов Умеренного пояса характеризуются сбалансированным полиморфизмом окраски раковины: выделяют бурую, розовую и желтую формы. Ранней весной наименее заметными на преобладающем фоне окружающей среды оказываются раковины бурой и розовой окраски, летом - желтые раковины. В то же время желтая форма менее заметна среди луговой растительности, тогда как бурая и розовая - в лесу. Очевидно, полиморфизм окраски у улиток соответствует 4-й и 5-й формам полиморфизма в приведенном выше перечне.
Наконец, последняя из указанных форм полиморфизма наблюдается, например, при мимикрии (подражательном сходстве) какого-либо вида с несъедобным для хищников видом-моделью. Такое сходство благоприятно для вида-подражателя, только если численность мимикрирующего фенотипа в его популяциях будет существенно ниже численности несъедобного вида-модели (в противном случае хищник не сможет эффективно "обучаться" не трогать особей с данным фенотипом). В ситуации, представленной на рис. 5, среди самок бабочки Papilio dardanus, разные цветовые устойчивые формы (морфы) имитируют окраску разных несъедобных бабочек-данаид, тогда как самцы P.dardanus вообще не мимикрируют.
Симпатрические внутривидовые группировки
Полиморфизм популяций является основой для возникновения в некоторых случаях устойчивых и обособленных внутривидовых группировок особей ("биологических популяций"), обитающих в одном и том же районе, т.е. обладающих симпатрическим распространением, в отличие от аллопатрических территориальных популяций. К ним относятся, в частности, возрастные и половые группы некоторых видов. Биологические популяции обособляются друг от друга, обитая в пределах одного ареала, либо в результате специфических взаимоотношений с внешней средой, либо благодаря особенностям внутривидовой организации. Примеры подобной внутривидовой дифференциации представляют обособленные друг от друга биологически личиночные и взрослые популяции земноводных или насекомых с полным превращением, или же такие связанные с особенностями размножения группировки, как "гаремы" и стада самцов-холостяков, характерные для целого ряда видов млекопитающих (кашалоты, олени, зебры, куланы и др.).
Особой формой биологических популяций являются так называемые биологические расы. Этим термином обозначают такие устойчивые группировки особей одного вида, которые имеют симпатрическое распространение, но различаются друг от друга определенными экологическими особенностями (например, характером питания, сроками и местами размножения и т.п.). Благодаря этим различиям разные биологические расы одного вида держатся раздельно друг от друга, хотя и имеют общий ареал. В результате обмен генетической информацией между разными биологическими расами одного вида может быть снижен, как и между разными территориальными (аллопатрическими) популяциями.
Примером подобных биологических рас могут служить так называемые сезонные ("яровые" и "озимые") расы ряда видов лососевых и осетровых рыб, описанные Л. С. Бергом. Речь идет о проходных рыбах, которые обитают в море, но для икрометания заходят в реки. Сезонные расы описаны Бергом у кеты (Oncorhyn-chus keta), чавычи (О. tschawytscha), нерки (О. nerka), семги (Salmo salar), каспийского лосося (S.trutta caspius), севрюги (Acipenser stellatus) и других видов. Рыбы "озимой" расы входят в реки осенью, зимой держатся в их низовьях, а рано весной поднимаются вверх по течению для нереста. Рыбы "яровой" расы проводят зиму в море и входят в реки позднее - когда нерест рыб "озимой" расы уже заканчивается. Благодаря этому обе расы одного вида, обитая в одном географическом районе и выметывая икру в одних и тех же реках, в значительной степени изолированы друг от друга во время сезона размножения и обмен генетической информацией между ними затруднен.
Другой пример представляют биологические расы обыкновенной кукушки (Cuculus canorus), описанные А.С.Мальчевским. Как известно, кукушка откладывает яйца в гнезда различных видов воробьиных птиц (это явление иногда называют "гнездовым паразитизмом"). Представители разных биологических рас обыкновенной кукушки проявляют предпочтение к "своему" виду воробьиных, в гнезда которого они откладывают яйца. При этом окраска яиц у кукушек разных рас соответствует окраске яиц того вида птиц, на котором "паразитирует" данная раса кукушек. Следовательно, в этом случае разные биологические расы проявляют уже Достаточно отчетливые наследственные различия морфологического признака (окраска яиц).
Очевидно, биологические внутривидовые группировки, так же как и территориальные популяции, позволяют виду более полно и совершенно использовать ресурсы в пределах занимаемого им ареала.
Подводя итоги, мы можем с полным основанием констатировать, что биологический вид представляет собой сложную систему, обладающую высокой устойчивостью к изменениям внешней среды и приспособленную к оптимальному использованию существующего разнообразия условий. Высокая степень приспособленности и приспособляемости вида обеспечивается несколькими путями, включающими мутационную, комбинативную и моди-фикационную изменчивость и формирующиеся на базе комбинативной изменчивости политипичность и полиморфизм. Вид представляет собой систему с нежесткими связями взаимозаменяемых компонентов (так сказать, с "параллельным включением" звеньев), лишенную какого-либо интегрирующего, контролирующего всю систему центра. Эта сложная динамическая система возникает, поддерживается и совершенствуется под контролем естественного отбора.
Естественный отбор
Как уже было отмечено, мутационная изменчивость необходима, но сама по себе еще недостаточна для осуществления приспособительной и прогрессивной эволюции. Приспособленность к конкретным условиям существования - важнейшая предпосылка выживания и нормальной жизнедеятельности организмов - формируется только благодаря действию естественного отбора. Естественный отбор является творческой силой эволюции, направляющей (векторизующей) эволюционный процесс и интегрирующей отдельные изменения в адаптации. В этом современные представления вполне соответствуют взглядам Дарвина. Однако следует отметить, что отбор не требует перенаселения как обязательного условия, обеспечивающего его эффективность. Борьба за существование, понимаемая в том широком и метафорическом смысле, который вкладывал в него сам Дарвин, означает всю совокупность взаимоотношений организма и внешней среды и, следовательно, должна иметь место и без перенаселения. Естественный отбор поэтому должен действовать и в ситуациях, далеких от перенаселения.
Генетическая сущность отбора заключается в неслучайном (избирательном) выживании изменчивых генотипов и в избирательном участии разных генотипов в передаче генов последующим поколениям. Следовательно, естественный отбор имеет два существенно различающихся аспекта: отбор на выживание, определяющий дифференцированное выживание разных особей, и репродуктивный отбор (не путать с половым отбором), обеспечивающий различную степень участия разных особей в размножении. Эти два аспекта отбора могут не совпадать - например, стерильный гибрид может обладать повышенной жизнеспособностью.
Различные факторы внешней среды, воздействующие на данный вид организмов и требующие определенной приспособительной реакции организмов, представляют собой отдельные силы отбора (или селективные силы). Общее воздействие отбора на данный вид организмов, иногда называемое давлением отбора или селективным давлением, складывается как результат интеграции отдельных селективных сил. Взаимодействие последних друг с другом может оказывать существенное влияние на общий эффект отбора.
Если разные селективные силы имеют несовпадающие направления, общий итог их взаимодействия получает характер эволюционного компромисса, подразумевающего формирование под контролем отбора такого фенотипа, который обеспечивает скорее некоторое оптимальное соотношение разных адаптации, чем достижение максимального совершенства одной из них.
Необходимо отметить, что до сих пор определенное распространение (особенно среди критиков дарвинизма) имеют упрощенные и, по существу, неверные представления о естественном отборе, которые ведут к серьезным ошибкам в понимании механизмов эволюционного процесса. Прежде всего неверно представление о естественном отборе как изолированном действии отдельных селективных сил на отдельные признаки организма, т. е. как отборе отдельных признаков (и тем более - отдельных аллелей). Темпы эволюции, рассчитанные на основе этих представлений, оказываются гораздо более низкими, чем наблюдаемые реально поданным палеонтологии темпы эволюции многих филетических линий, а плата за отбор - непомерно высокой. Напомним, что отбору непосредственно подвергаются не отдельные гены, а определенные фенотипы, формирующиеся в онтогенезе на основе соответствующих генотипов при модифицирующем влиянии внешних условий. Следовательно, отбираются целые генотипы, характеризующиеся такой нормой реакции, которая наиболее благоприятна в данных условиях среды по всей совокупности признаков. Благодаря блочно-иерархическому принципу организации генома отбор может действовать на целостные генетические комплексы (супергены), что на несколько порядков ускоряет эволюционный процесс.
Поскольку всякий генотип характеризуется специфическим спектром мутационных изменений (т. е. обладает способностью с некоторой частотой давать определенные мутации), естественный отбор, благоприятствуя конкретным генотипам, тем самым благоприятствует и соответствующему направлению и частоте возникновения наследственных вариаций. Таким образом, если отбор благоприятствует каким-то фенотипическим признакам, носителем которых является данный генотип, то отбор тем самым влияет и на характер дальнейших эволюционных изменений.
Преобладающие направления изменчивости различных генотипов могут в разной степени соответствовать направлению отбора. В этом случае при наличии двух генотипов, которые являются носителями какого-то нового полезного признака, но различаются своими спектрами изменчивости, отбор, очевидно, будет в большей мере благоприятствовать тому генотипу, возможности изменчивости которого будут ближе к направлению действия отбора. При совпадении преобладающего направления изменчивости и направления отбора приспособительная эволюция пойдет гораздо быстрее, чем при отсутствии такого совпадения. Быстрота возникновения и усовершенствования приспособлений дает очевидное преимущество в борьбе за существование. Таким же образом отбор может воздействовать и на частоту определенных мутаций (через посредство соответствующих генов-мутаторов), поскольку этот показатель существенно влияет на распространенность в популяции благоприятных вариантов.
Следовательно, естественный отбор, выбраковывая одни и сохраняя другие генотипы, тем самым оказывает влияние на направление и частоту возникающих наследственных изменений, что также способствует значительному ускорению эволюционного процесса.
Вторая группа ошибочных представлений о естественном отборе связана с непониманием его творческой роли. Эти представления коренятся в упрощенной концепции отбора, которая на первый взгляд кажется очевидной: отбор понимается как своего рода "сито", устраняющее наименее приспособленные варианты наследственных изменений (мутаций) и сохраняющее более приспособленные. Именно такое понимание отбора характерно, например, для сторонников "нейтральной эволюции" (М. Кимура, С. Оно и др.), подчеркивающих "консервативную природу" отбора и видящих движущую силу эволюции в мутационном процессе (см. гл. 1). Этот очевидный результат действия отбора, несомненно, существует, но отнюдь не исчерпывает его эффект во всей полноте. При концентрации внимания на "консервативной роли" отбора странным образом забывают, что отбор не только выбраковывает неудачные варианты, но и накапливает удачные, формируя из них адаптации любой степени сложности, и преобразует эти адаптации в соответствии с изменениями внешних условий.
Отбор оказывает на организм многостороннее интегрирующее влияние, способствуя оптимизации взаимодействий разных его систем. Приспособительное, формообразовательное и интегрирующее воздействие отбора на организм и обозначается как творческая роль естественного отбора.
Интенсивность действия отбора
Естественный отбор можно представить как вектор, поскольку его, с одной стороны, характеризует величина, которую можно выразить количественно, с другой стороны - направление.
Количественная характеристика действия отбора носит название его интенсивности или эффективности: она представляет собой степень воздействия (давления) отбора на популяцию. Очевидно, давление отбора в разных условиях может быть большим или меньшим. Для количественного выражения интенсивности отбора используют показатель, называемый коэффициентом отбора.
Пусть некоторая популяция включает два типа особей - А и В.
Концентрации особей А и В в популяции соотносятся как СА/СВ=u1.
Если один из этих вариантов имеет преимущество перед другим, измеряемое коэффициентом интенсивности отбора S, то в следующем поколении соотношение концентраций особей А и В в популяции изменится в соответствии с этим коэффициентом:
u2= u1(1-S)
S=(u1-u2)/u1=1-u2/u1
где u2 – соотношение концентрации особей в следующем поколении.
Если u1>u2, то S>0 и особи В имеют какое-то преимущество перед особями А, которое "отмечается" отбором, и концентрация особей В в популяции возрастает от поколения к поколению. Если же u1=u2, то S = 0, следовательно, особи А и В равноценны и соотношение их концентраций в данной популяции не меняется.
Коэффициент интенсивности отбора позволяет сравнивать давление отбора на разные фенотипы. Для полиморфной популяции устанавливают коэффициент отбора всех вариантов по отношению к наиболее приспособленному (т.е. наиболее жизнеспособному и плодовитому в данных условиях) фенотипу. Очевидно, коэффициент отбора представляет собой величину относительную и не может характеризовать действие отбора только на данный фенотип без сравнения с другими вариантами.
Формы отбора
Если интенсивность давления отбора является его количественной характеристикой, то направление естественного отбора определяет качественную специфику его воздействия на популяцию. Выделяют три основных направления действия отбора, о которых обычно говорят как о типах (или формах) естественного отбора.
Стабилизирующий отбор. Если условия внешней среды длительное время остаются стабильными, приспособленность вида к этим условиям постепенно совершенствуется, пока не достигнет определенного оптимума. Совершенство всякого приспособления, разумеется, относительно, обычно возможна и более высокая степень приспособленности к данным условиям. Однако эволюционные возможности всякого конкретного генофонда, приспосабливаемого отбором к данным условиям среды хотя и велики, но ограничены спектром возможных мутационных изменений. Поэтому раньше или позже при постоянстве условий среды и при отсутствии конкуренции с экологически близкими формами генофонд данного вида оптимизируется по отношению к существующим условиям. С этого момента на популяцию начинает действовать стабилизирующий отбор, благоприятствующий сохранению оптимального генофонда, на базе которого формируются оптимально приспособленные к данным условиям фенотипы, которые становятся преобладающими в популяции. Процессы онтогенетического формообразования (морфогенезы), основывающиеся на разных генотипах, направляются стабилизирующим отбором в определенные русла, ведущие к формированию оптимального фенотипа (посредством распространения в популяции соответствующих генов-модификаторов). К.Уоддингтон назвал такое состояние канализованностью процессов морфогенеза.
В результате дикий тип, т.е. преобладающий в природной популяции оптимальный фенотип (или несколько фенотипов в полиморфной популяции), обычно объединяет генетически разнородных особей. Запас комбинативной изменчивости популяции благодаря канализованности процессов морфогенеза оказывается как бы скрытым за фенотипическим однообразием особей. Такое соотношение комбинативной и модификационной изменчивости оптимально при стабильных условиях среды, поскольку запас комбинативной изменчивости необходим для вида как "запас прочности", повышающий приспособляемость к изменениям среды обитания, а подгонка разных генотипов к одному или немногим оптимальным фенотипам повышает приспособленность к существующим в данный момент условиям. Состояние оптимальной стабильности популяции, которое обеспечивается ее специфическим генофондом и канализованностью процессов морфогенеза под контролем стабилизирующего отбора, носит название популяционного гомеостаза.
Стабилизирующий отбор направлен против крайних вариантов изменчивости, наблюдаемых в популяции, и благоприятствует консервативной части последней (рис. 12). В результате средняя величина (среднее состояние) признака остается из поколения в поколение неизменной. Стабилизирующий отбор ведет к большей фенотипической однородности популяции и к стабильности спектра ее изменчивости.

Очевидно, это направление отбора прямо противоположно схеме дивергентной эволюции, разработанной Ч.Дарвином. Под контролем стабилизирующего отбора, действующего длительное время, вид может внешне оставаться неизменным. Однако эта неизменность вида, в сущности, кажущаяся - она касается лини, постоянства оптимальных фенотипов. И.И.Шмальгаузен, разработавший концепцию стабилизирующего отбора, подчеркнул, что в таких условиях генофонд вида продолжает изменяться за счет накопления мутаций, фенотипическое проявление которых взаимно нейтрализуется (важную роль в этом играют гены-модификаторы), а также посредством накопления в гетерозиготном состоянии рецессивных мутаций. Таким образом, генофонд вида обогащается новыми аллелями, которые, оставаясь скрытыми, увеличивают мобилизационный резерв изменчивости, повышающий устойчивость вида к возможным изменениям внешних условий. Эти выводы подкрепляются результатами исследований наследственной изменчивости у разных видов, обитающих в стабильных внешних условиях - например у ряда морских животных (ракообразные, двустворчатые моллюски, иглокожие и другие группы). Оказалось, что генетическая гетерогенность популяций у видов, обитающих в стабильных условиях, не отличается от таковой у видов, обитающих в изменчивой среде.
При действии стабилизирующего отбора может происходить даже дивергенция видов, принимающая, однако, своеобразный характер. Механизм такого видообразования мы рассмотрим в следующей главе.
Направленный отбор наблюдается в том случае, если условия среды благоприятствуют какому-либо одному определенному направлению изменений признака, тогда как все другие варианты изменений подвергаются в той или иной степени негативному давлению отбора (рис. 13). В этом случае в популяции от поколения к поколению происходит сдвиг средней величины признака в определенном направлении. Результатом действия направленного отбора является постепенное изменение популяции в целом, происходящее посредством преобразования популяционного генофонда. В отличие от стабилизирующего отбора изменение частоты встречаемости различных аллелей и закрепление новых мутаций в генофонде при действии направленного отбора быстро отражаются на фенотипическом облике популяции. Это происходит тем быстрее, чем больше интенсивность отбора, если только последняя не превосходит приспособительных возможностей данной популяции - в последнем случае популяция вымирает.

При направленном отборе особенно ярко проявляется воздействие отбора на темпы и преобладающее направление наследственной изменчивости (длящаяся изменчивость, по Ч.Дарвину): получают преимущества такие генотипы, которые обладают соответствующими возможностями дальнейших мутационных изменений, частота определенных мутаций может быть повышена с помощью генов-мутаторов; отбираются гены-модификаторы, способствующие доминантному фенотипическому проявлению благоприятных мутаций.
Осуществляемые под контролем направленного отбора эволюционные преобразования данной популяции затрагивают ее генофонд как целое; обособления каких-либо дочерних популяций (т.е. дивергенции) не происходит. Такую форму эволюционных преобразований вида Д.Симпсон назвал филетической эволюцией.
Разрывающий, или дизруптивный, отбор действует в том случае, когда условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака (рис. 14). В результате происходит возрастание изменчивости с последующим сдвигом среднего состояния признака в разных направлениях. Разрывающий отбор ведет либо к возникновению устойчивого адаптивного полиморфизма популяции, либо к ее дроблению, т. е. к разделению на несколько новых популяций такого же ранга. Последний вариант эволюционных изменений всего ближе к дарвиновской концепции дивергентной эволюции.
Тот или другой результат определяется конкретными условиями действия разрывающего отбора: при наличии какой-либо функциональной взаимозависимости между разными благоприятствуемыми генотипами в единой панмиктической популяции возникает полиморфизм, тогда как под влиянием различных вариантов условий в разных частях ареала при снижении скрещивания между населяющими их субпопуляциями происходит дивергенция последних. Д. Симпсон назвал последнюю форму эволюционных изменений видообразованием, поскольку она ведет к возникновению от одного родительского вида двух или нескольких новых - в противоположность филетической эволюции, преобразующей вид как целое, без разделения на дочерние виды.
При действии дизруптивного отбора у молодых дивергирую-щих видов соотношение фенотипической (модификационной) и комбинативной изменчивости становится противоположным тому состоянию, которое возникает под контролем стабилизирующего отбора: системы канализованности индивидуального развития разрушаются, в популяции происходит значительное усиление комбинативной и модификационной изменчивости, что приводит к возрастанию фенотипического разнообразия. Такая картина изменчивости была описана В.В.Кузнецовым в пределах комплекса видов сиговых рыб Coregonus muksun. Быстрое фе-нотипическое выявление разных вариантов комбинативной изменчивости повышает эффективность действия естественного отбора.
И разрывающий, и направленный отбор приводят к векторизованным изменениям генофонда и фенотипического облика популяций; сходно также их воздействие на онтогенез (разрушение систем его канализованности). Отдельные направления разрывающего отбора, действующие на разные аллопатрические популяции одного вида, можно рассматривать применительно к каждой данной популяции как направленный отбор. Поэтому иногда эти две формы отбора объединяют как движущий отбор, противопоставляемый стабилизирующему.
По отношению к разным адаптациям, различным признакам фенотипа у организмов одного вида отбор может иметь разное направление: для одних он оказывается движущим, для других -стабилизирующим. Когда давление движущего отбора по отношению к данной адаптации ослабевает, стабилизирующий отбор способствует закреплению достигнутых движущим отбором результатов, благоприятствуя оптимизации путей морфогенеза у новых адаптивных признаков для достижения максимальной надежности их воспроизведения в фенотипе (с повышением пенетрантно-сти и экспрессивности соответствующих аллелей посредством воздействия генов-модификаторов, а также механизма генокопиро-вания, см. гл. 1).
Одним из ярких примеров действия движущего отбора является развитие так называемого "индустриального меланизма", описанного более чем у 70 видов бабочек, распространенных в Англии и ряде других стран Европы, а также в США. Индустриальный меланизм представляет собой значительное повышение частоты встречаемости меланистических (имеющих темную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Особенно хорошо изучено это явление у березовой пяденицы Biston betularia (рис. 15) Англии.

До середины XIX в. в популяциях этого вида преобладали особи, имевшие светлую серовато-белую окраску с темными пятнышками, хорошо маскирующую бабочек на покрытых лишайниками стволах деревьев. В 1848 г. в Манчестере был отловлен первый экземпляр бабочки, имевший меланистическую окраску (морфа carbonaria). С тех пор частота встречаемости меланистических особей в популяциях этой бабочки, обитающих в промышленных районах Англии, постепенно возрастала, пока к началу XX в. количество меланистов не достигло в отдельных популяциях 95% всех особей. Популяции с преобладанием светлоокрашенных бабочек обитают ныне лишь на юго-западе Англии, в Северной Шотландии и в Ирландии. Варианты окраски у березовой пяденицы контролируются аллелями одного гена; аллель, обусловливающий темную окраску, имеет доминантное фенотипичес-кое проявление.
Возможно, меланистические особи в небольшой концентрации встречались в популяциях бабочек и до того момента, когда на них впервые обратили внимание ученые. Однако более широкому распространению аллеля, обусловливающего темную окраску насекомых, в те времена не благоприятствовали условия: ба-бочки-меланисты были гораздо лучше заметны на светлом фоне лишайников, покрывавших кору деревьев, чем "нормальные" светлоокрашенные особи. С развитием промышленности загрязнение воздуха вокруг промышленных центров привело к гибели лишайников на стволах деревьев, которые потемнели от копоти. В этих условиях меланистические формы бабочек оказались в лучших условиях, чем особи со светлой окраской, поскольку первые менее заметны на темных стволах деревьев, чем вторые. Это и определило быстрое увеличение концентрации бабочек-меланистов в популяциях промышленных районов. Благодаря меньшей "задымленности" воздуха в Ирландии, Северной Шотландии и Юго-Западной Англии там сохранилось прежнее преобладание светлой формы В. betularia.
Г. Кеттлуэлл в 50-60-х гг. XX в. экспериментально проверил эти соображения, выпустив определенное количество бабочек темной и светлой окраски в загрязненном лесу вблизи Бирмингема и затем повторив этот опыт в незагрязненном лесу в Дорсете. По данным Кеттлуэлла, птицы, питающиеся бабочками и, так сказать, непосредственно осуществляющие отбор последних, в Бирмингеме уничтожили значительно больше светлых бабочек, чем темных, тогда как в Дорсете наблюдалось обратное соотношение. Явление индустриального меланизма предоставляет ученым замечательные возможности для наблюдений за действием отбора в естественных популяциях. Этот эксперимент, "поставленный" природой при участии человека, уже показал, что изменения генофонда популяций могут происходить под контролем отбора чрезвычайно быстро - в течение нескольких десятилетий. Новые наблюдения показывают, что различия меланистических и светлых особей не сводятся только к разнице в окраске. Кеттлуэлл показал, что и те, и другие бабочки выбирают для отдыха субстраты предпочтительно неконтрастирующей окраски, т.е. ме-ланисты предпочитают темный, а "нормальные" особи - более светлый фон. Имеются также данные о различной жизнеспособности гусениц разных форм бабочек. Возможно, за сто с лишним лет, прошедшие со времени обнаружения первых бабочек-меланистов, не только перестроился генофонд отдельных популяций березовой пяденицы, но и произошли определенные преобразования внутри некоторых генных комплексов с участием генов-модификаторов, которые привели к повышению жизнеспособности и изменениям поведения у темных особей. Эти преобразования могли коснуться и степени доминантности фенотипического проявления того и другого аллелей в пользу повышения доминантности аллеля меланизма. Можно ожидать, что дальнейшее изучение индустриального меланизма и подобных ему явлений позволит проверить и многие другие положения теории отбора.
В обычном понимании отбор представляет собой преимущественное выживание и оставление потомства наиболее приспособленными особями. Можно сказать, что в этом аспекте отбор выступает как своего рода "эгоистический" фактор: видовые адаптации совершенствуются как адаптации отдельных особей. Однако, как мы уже подчеркивали в предшествующей главе, популяции и вид в целом являются целостными системами. Поэтому отбор должен также благоприятствовать закреплению и распространению признаков, которые сами по себе для отдельных особей не имеют приспособительной ценности или даже снижают их приспособленность, но обеспечивают существенные преимущества данной группировке особей как целому. Примеры действия такого "альтруистического" аспекта естественного отбора особенно многочисленны в поведении высших животных. Так возникли "сигналы тревоги", характерные для многих видов птиц: обнаружившая врага птица не затаивается в укрытии (что было бы наиболее благоприятно для ее собственной безопасности), а издает громкий крик, выдающий ее местонахождение хищнику, но предупреждающий других особей своего вида (часто сигнал тревоги воспринимается и другими видами). К этой же форме поведения принадлежит коллективная защита детенышей в стаях и стадах некоторых видов млекопитающих (слоны, копытные), защита и помощь другим особям своего вида (например, поддерживание на плаву раненых животных некоторыми китообразными).
При развитии подобных альтруистических особенностей поведения отбор оперирует не отдельными особями, а целыми их совокупностями (минимальной из них является так называемый дем - группа особей, непосредственно связанных в процессе размножения), благоприятствуя сохранению тех из них, в генофонде которых появляются "альтруистические" аллели. Этот аспект действия естественного отбора получил название "групповой отбор".
Концепция полового отбора была разработана Ч.Дарвином для объяснения развития у самцов некоторых видов животных украшающих признаков, нередко гипертрофированных, причудливых и ярких и поэтому отрицательно сказывающихся на защищенности этих особей и требующих для своего развития значительных энергетических затрат. Очевидно, предпочтение самками самцов с подобными признаками нуждается в объяснении. Первая попытка такого объяснения была сделана Р.Фишером, предположившим, что половой отбор, благоприятствующий развитию украшающих признаков, может действовать на основе предпочтения некоторыми самками таких признаков у самцов. В этом случае самцы, обладающие подобными украшениями, получают лучшие шансы оставить потомство, в котором самцы унаследуют от своих отцов эти признаки, а самки - от своих матерей соответствующие сексуальные предпочтения. Этот механизм (именуемый "фишеровскими процессами") обусловливает возникновение полового отбора, благоприятствующего дальнейшему развитию украшающих признаков у самцов. При этом первоначальное предпочтение самками определенных внешних признаков самцов может быть обусловлено особенностями физиологии их органов чувств (гипотеза первоначального сенсорного предпочтения).
А. Захави предложил интересное объяснение приспособительного значения украшающих признаков: степень их развития в целом должна соответствовать жизнеспособности самцов, поскольку такие "избыточные" структуры без ущерба для организма могут развивать лишь наиболее сильные индивиды с высоким уровнем обмена веществ и энергетических процессов, который контролируется генетически ("принцип гандикапа"). Тем самым степень развития у самцов таких украшений может служить для самок признаком, позволяющим выбрать наиболее жизнеспособных самцов - носителей "оптимальных" генов. Вероятно, развитие украшающих признаков под контролем полового отбора может происходить при совместном действии "фишеровских процессов" и "принципа гандикапа".
Факторы, влияющие на действие отбора
Действие того или иного типа (формы) отбора на каждый конкретный вид организмов определяется особенностями генетической системы данного вида, спецификой его взаимоотношений с внешней средой и характером изменений внешних условий. Очевидно, каждому из трех указанных выше основных направлений отбора благоприятствуют совершенно разные условия: постоянство условий существования - стабилизирующему отбору, изменение условий в одном направлении на всем ареале вида - направленному отбору, разнообразие условий существования (или изменение условий в разных направлениях) в пределах видового ареала - дизруптивному отбору. Вообще в филогенезе всякой группы организмов в течение ее длительной истории направление отбора многократно изменяется, хотя в некоторых случаях долгое время может преобладать какое-либо одно постоянное направление. Длительно действующий направленный отбор приводит к быстрым эволюционным преобразованиям значительного масштаба. Напротив, стабилизирующий отбор в некоторых случаях как бы "консервирует" фенотипический облик отдельных видов организмов в течение длительного времени.
В некоторых местообитаниях абиотические условия относительно мало изменялись или даже оставались в целом стабильными в течение многих миллионов лет (таковы, например, определенные зоны морского дна). В таких условиях некоторые виды организмов могут сохранять свой фенотипический облик практически неизменным в течение значительных периодов времени. Это так называемые персистирующие формы ("живые ископаемые"). Широко известными примерами таких организмов являются равноногие рачки-щитни Triops (рис. 16), организация которых не подвергалась заметным изменениям с триасового периода (более 200 млн лет); двустворчатые моллюски Leda, Nucula, Modiolus, существующие с каменноугольного периода (около 300 млн лет); плеченогие (брахиоподы) Lingula (рис. 17), не претерпевшие изменений с девонского периода(около 380 млн лет). Однако не следует рассматривать персистирующие виды как совершенно те же самые формы организмов,которые жили на Земле сотни миллионов лет назад: как мы уже отмечали, и под контролем стабилизирующего отбора генофонды видов продолжают изменяться, и с течением времени в них накапливается все больше мутаций структурных генов и соответствующи хмутаций генов-модификаторов, препятствующих изменениям фенотипа. Таким образом, в генетическом отношении современные персистирующие виды должны значительно отличаться от своих фЕнотипичЕски сходных предков.


Со времени Ч.Дарвина существенно изменились представления о факторах, благоприятствующих действию естественного отбора. Среди факторов, названных Дарвином (см. гл. 2, ч. I), универсальное значение имеет только длительность времени. Высокая изменчивость благоприятна в наибольшей степени для движущего отбора, стабилизирующий отбор действует против большого размаха изменчивости. Обширность ареала вида благоприятствует дизруптивному отбору.
Географическая изоляция территориальных (аллопатрических) популяций вида ныне рассматривается как один из важнейших факторов, способствующих действию разрывающего отбора. Некоторые ученые считают этот фактор непременным условием процесса видообразования. Под географической изоляцией понимается отделение данной популяции от других популяций родительского вида каким-либо географическим барьером, непроходимым для особей данного вида. Таким барьером может быть морской пролив, горный хребет, русло реки или даже просто район с непригодными для жизни данного вида условиями температуры, влажности, с отсутствием необходимой пищи и т. п.
Численность популяций и дрейф генов
Среди факторов, влияющих на эффективность отбора, особое место занимает численность популяций. С одной стороны, нельзя забывать, что действие отбора вообще имеет вероятностный характер (случайные причины могут привести к гибели или воспрепятствовать участию в размножении отдельных особей, являющихся носителями благоприятствуемых отбором признаков), поэтому для действия отбора необходима достаточно большая численность популяций. Однако, с другой стороны, в очень крупных панмиктических популяциях эффективность отбора может быть в определенной степени снижена благодаря относительной редкости выявления рецессивных мутаций в гомозиготном состоянии; обширный генофонд больших популяций более инертен, чем таковой популяций меньшего масштаба. Поэтому для действия движущего отбора наиболее благоприятны популяции средних размеров, но достаточно большие, чтобы отбор мог оказывать свое влияние вопреки действию случайных факторов.
Численность популяций в природе подвержена значительным колебаниям, зависящим от различных изменений внешней среды (таких, как засуха, морозная или бесснежная зима, урожай или неурожай кормовых растений, численность популяций других видов животных и т. п.). В урожайные годы численность популяций данного вида может существенно повыситься, вслед за чем при ухудшении условий наступает ее спад, тем более резкий, чем больше контраст условий в последовательные годы. Особенно ярко эти "популяционные волны" проявляются у видов с коротким жизненным циклом и быстрой сменой поколений (мелкие грызуны, ящерицы, лягушки, насекомые, однолетние растения, микроорганизмы и т.п.), но колебания численности популяций присущи всем без исключения видам организмов.
С.С.Четвериков подчеркнул большое эволюционное значение колебаний численности природных популяций, которые он назвал "волнами жизни". Поскольку для действия отбора важна численность популяций, очевидно, что ее изменения должны сопровождаться изменениями интенсивности, а в некоторых случаях - и направления отбора. Кроме того, колебания численности популяций неизбежно приводят к изменениям концентраций различных аллелей в популяционном генофонде, которые могут иметь случайный характер (исчезновение редких аллелей при снижении численности популяций или, наоборот, их значительное распространение в генофонде и т. п.). Такие изменения генофонда сами по себе представляют материал для действия отбора (отбор оптимального генофонда при последующей стабилизации условий).
В небольших популяциях, подвергшихся изоляции от других Популяций своего вида, влияние случайных факторов может выйти на первый план по отношению к действию отбора. Эта интересная ситуация в 1931 г. была описана Н.П.Дубининым и Д.Д.Ромашовым под названием "генетико-автоматические процессы" и, независимо от них, в том же году С. Райтом, который обозначил ее термином "дрейф генов". Если небольшая популяция (менее 50 особей) оказывается изолированной, прежде всего проявляется эффект, названный "принципом основателя". Генофонд малочисленной популяции с самого начала беднее, чем генофонд родительского вида, и подбор аллелей в генофонде "популяции основателей" случаен из-за случайного подбора особей. Такой генофонд не сбалансирован, поскольку он возник на основе случайности, а не под контролем естественного отбора. Так как отбор действует на вероятностном уровне и требует для своей эффективности достаточной численности особей, дальнейшая судьба генофонда маленькой популяции определяется в первую очередь действием различного рода случайных факторов.
Общей тенденцией будет выпадение из генофонда малой популяции наиболее редких аллелей, сопровождающееся их замещением аллелями, которые с самого начала были более многочисленными. Это ведет к дальнейшему обеднению генофонда малой популяции и в результате к его гомозиготизации. Важно подчеркнуть, что указанные изменения генофонда не имеют приспособительного характера: утраченные аллели могут обладать равной и даже большей приспособительной ценностью, чем аллели, ставшие в популяции преобладающими: ведь потеря первых определяется не действием отбора, а случайной гибелью особей. Такая случайная гибель особей от самых различных причин (хищники, заболевания, факторы абиотической среды) происходит всегда и в любой популяции, но при большом количестве особей она отступает на задний план - в среднем успешнее выживают и оставляют потомство лучше приспособленные организмы. В малой же популяции фактор случайности становится основным. Естественно, что в этом случае лучшие шансы для сохранения имеют первоначально более многочисленные аллели вне зависимости от их приспособительной ценности.
Таким образом, действие случайных факторов обедняет и значительно изменяет генофонд малой изолированной популяции по сравнению с его исходным состоянием. Это явление и называется собственно генетико-автоматическими процессами или дрейфом генов. В результате подобных процессов может сложиться жизнеспособная популяция со своеобразным генофондом (случайно, поскольку отбор в данном случае не играл ведущей роли!). Спектр изменчивости этой популяции существенно отличается от такового родительского вида. С постепенным увеличением численности особей вновь восстановится эффективность естественного отбора, который будет действовать уже на новый генофонд, обладающий специфическим спектром изменчивости. Совокупность всех этих процессов может привести к обособлению нового вида.
Соотношения роли отбора и дрейфа генов при разной численности популяций можно выразить по С.Райту следующим образом:
если численность популяции N<1/(2S) (где S- коэффициент интенсивности отбора, см. выше), преобладает воздействие дрейфа генов; если N>1/(4S) преобладает отбор; в диапазоне N от 1/(4S) до 1/(2S) совместно действуют оба фактора. Так, при умеренной интенсивности отбора (S=0,01) изменения соотношений любой пары аллелей регулируются дрейфом генов при /V<50; при слабой интенсивности отбора (S= 0,001) дрейф генов оказывается определяющим фактором при N< 500 и т.д.
В соответствии с этим генетико-автоматические процессы могут существенно влиять на генофонд популяции основателей в первое время после ее закрепления в новом районе, пока она еще достаточно малочисленна; затем на первый план неизбежно выходит действие естественного отбора. Если условия в новом районе останутся благоприятными, численность популяции быстро увеличится и эффект случайности - дрейф генов - будет все более затушевываться влиянием отбора, как бы ни было слабо его давление. Если же условия среды станут неблагоприятными и численность популяции вследствие этого останется небольшой - это явится непосредственным результатом жесткого давления отбора и дрейф опять-таки не сможет быть определяющим фактором эволюционных перестроек генофонда в течение длительного времени.
Сравнивая эволюционное значение рассмотренных нами факторов, мы можем заключить, что наличие мутаций и естественного отбора необходимо и достаточно, чтобы обеспечить приспособительную и дивергентную эволюцию организмов. Поэтому мутационный процесс и естественный отбор можно обозначить как необходимые факторы эволюционного процесса. При этом отбор является единственным известным ныне фактором эволюции, который объединяет движущее, направляющее и интегрирующее воздействия на организмы, формируя их приспособления и влияя на саму мутационную изменчивость. Прочие рассмотренные нами элементарные эволюционные факторы (колебания численности, обмен генетической информацией между разными популяциями, географическая изоляция, дрейф генов) являются дополнительными по отношению к мутационному процессу и естественному отбору. Разумеется, для полного понимания эволюционного процесса необходимо учитывать сложные взаимодействия всех указанных элементарных эволюционных факторов.
Аллопатрическое видообразование
Как мы уже упоминали, всякая территориальная популяция характеризуется собственным генофондом, частота встречаемости разных аллелей в котором оптимизирована отбором применительно к условиям обитания данной популяции. При обособлении популяции обмен наследственной информацией ("поток генов") между нею и остальными популяциями прекращается. Генофонд изолированной популяции становится самостоятельным. Особенности местных условий определяют специфическое направление отбора, действующего на такую популяцию (это направление является одним из направлений дизруптивного отбора, действующего на вид в целом). Постепенно в генофонде изолированной популяции происходит накопление новых мутаций. Поскольку генофонд данной популяции специфичен, спектр его изменчивости должен несколько отличаться от такового других популяций вида. Специфика местных условий и соответствующее направление отбора усугубляют особенности генофонда изолированной популяции. Поэтому мутации, накапливающиеся в ее генофонде, будут в той или иной степени отличаться от мутаций, которые за это же время закрепятся в других популяциях родительского вида.
Если географическая изоляция достаточно продолжительна, а направления отбора достаточно различны, то изолированная популяция приобретает отчетливые различия с родительским видом, снижающие возможность скрещивания, вплоть до возникновения репродуктивной изоляции (т.е. нескрещиваемости в природных условиях) между ними. Репродуктивная изоляция может основываться на самых разных биологических особенностях близких форм организмов. Выделяют следующие основные формы репродуктивной изоляции: 1) этологическая (различия поведения, особенно брачного); 2) экологическая (различия предпочитаемых местообитаний); 3) временная, или сезонная (различия сроков размножения); 4) морфологическая (различия размеров тела и особенностей строения, важных для опознания половых партнеров, их взаимной стимуляции и спаривания); 5) генетическая (различия генома, приводящие к генетической несовместимости половых клеток). Все эти и другие неупомянутые здесь формы изоляции возникают независимо друг от друга и могут сочетаться в любых комбинациях или действовать поодиночке. Наиболее совершенна генетическая изоляция, исключающая всякую возможность обмена генетической информацией между двумя популяциями.
После возникновения той или иной формы репродуктивной изоляции обособленная популяция становится самостоятельным видом. Если теперь географический барьер, разделявший популяции, нарушится и вновь восстановится контакт между ними -эти два вида либо смогут сосуществовать в одном ареале, либо между ними возникнут конкурентные отношения и один из них будет вытеснять другой, если их биология осталась близкой. Но, так или иначе, это будут уже две обособленные филетические линии, каждая из которых имеет свою эволюционную судьбу.
Такая форма видообразования, основанная на территориальной изоляции разных популяций, получила название аллопатрической. Ч.Дарвин также указывал, что географическая изоляция способствует видообразованию, но придавал этому фактору меньшее значение, чем современная теория эволюции.
Если две изолированные друг от друга популяции одного вида находятся в сходных условиях, длительное время остающихся постоянными, на них будет действовать одинаково направленный стабилизирующий отбор. В таких популяциях формируются практически одинаковые оптимальные фенотипы. Однако под прикрытием этих оптимальных фенотипов в генофонде популяций, как мы видели выше (гл. 3), происходит постепенное накопление новых мутаций. Если обмен генетической информацией между двумя такими популяциями затруднен в течение достаточно длительного времени, в их генофондах могут накапливаться разные мутации. В результате популяции, оставаясь внешне сходными, будут все более различаться в генетическом отношении. С накоплением этих различий возможности их гибридизации постепенно снижаются, вплоть до полной невозможности скрещивания (возникновение генетической несовместимости) особей из разных популяций. На этой стадии две обособленные популяции превращаются в два самостоятельных вида, которые, однако, сохраняют фенотипическое сходство. Такие виды носят название видов-двойников. Сходство видов-двойников может быть столь полным, что даже опытные зоологи-систематики нередко с большим трудом различают подобные формы, единственным надежным критерием для разделения которых является их нескрещиваемость друг с другом.
Примером видов-двойников могут служить хорошо изученные формы плодовых мушек Drosophila pseudoobscura и D. persimilis. За 35 лет исследований в природных популяциях было обнаружено лишь 4 достоверно установленных случая гибридизации между этими фенотипически чрезвычайно сходными видами. Обмену генетической информацией между D. pseudoobscura и D. persimilis препятствуют различия поведения (этологическая изоляция), стерильность гибридных самцов, пониженная жизнеспособность и стерильность потомства от возвратных скрещиваний гибридных самок. Виды-двойники обнаружены в большинстве групп животных и растений.
Изолированные популяции могут возникать двумя путями: либо при появлении каких-либо географических барьеров, разделяющих прежний ареал вида, либо при активном расселении вида на новые территории за пределы занимаемого им ареала. В этом случае относительно небольшие популяции, укоренившиеся в новых районах, могут с самого момента своего возникновения стать территориально обособленными (периферические изоляты). Этот последний случай особенно интересен тем, что периферические изоляты могут сразу подвергнуться действию условий, существенно отличающихся от таковых в пределах ареала вида. Э.Майр высказал предположение, что в периферических изолятах могут в некоторых случаях происходить относительно очень быстрые и резкие изменения генофонда, приводящие к эволюционным изменениям значительного масштаба. Этот процесс получил название "генетическая революция". Основную причину генетической революции Э.Майр видит в том, что генофонд периферического изолята (или популяции основателей) с самого начала не сбалансирован и не оптимизирован отбором. Поэтому при закреплении популяции основателей в новом для вида районе ее генофонд должен подвергнуться существенной перестройке под давлением отбора, до установления новой формы генетического равновесия. При малочисленности популяций основателей преобразования их генофондов ускоряются явлениями дрейфа генов.
Г.Льюис подкрепил гипотезу Э.Майра о возможности генетических революций анализом последствий действия на такие популяции особенно интенсивного отбора, названного им "катастрофическим". Популяции, обитающие в краевой зоне ареала вида, часто подвергаются действию необычных для данного вида экстремальных условий. При крайне неблагоприятных условиях на популяцию действует катастрофический отбор, элиминирующий большинство составляющих ее особей. При этом выживают немногие индивиды, обладающие наиболее высокой фенотипической устойчивостью к неблагоприятным факторам (на основе соответствующих генетических уклонений). После возвращения условий к норме численность популяции восстанавливается, но ее генофонд после действия катастрофического отбора должен существенно отличаться от исходного.
Проблема симпатрического видообразования
Некоторые ученые допускают возможность дивергенции видов из одной и той же территориальной популяции, без ее разделения на аллопатрические группировки (т.е. без географической изоляции). Симпатрическое видообразование должно основываться на высокой степени популяционного полиморфизма, причем устойчивые морфы, имеющиеся в одной популяции, не должны быть взаимозависимы. При наличии такого полиморфизма в исходной популяции действие достаточно интенсивного разрывающего отбора, благоприятствующего наиболее уклонившимся морфам и элиминирующего все промежуточные состояния, может привести к снижению панмиксии и затем к обособлению устойчивых биологических рас - симпатрических внутривидовых группировок, различающихся определенными экологическими особенностями. Дальнейшая экологическая специализация биологических рас под давлением интенсивного разрывающего отбора может привести к их репродуктивной изоляции друг от друга. Д.Смит (1966) сформулировал ряд условий, необходимых для успеха симпатрического видообразования под действием дизруптив-ного отбора: 1) обитание вида более чем в одной экологической нише (т.е. использование различными симпатрическими группировками разных местообитаний, разных кормов и способов их добывания и т. п.); 2) отдельное регулирование численности популяций, занимающих каждую нишу; 3) наличие отмечаемых отбором преимуществ каждой популяции в своей нише; 4) возникновение репродуктивной изоляции между указанными симпатрическими популяциями.
Основной трудностью, стоящей перед концепцией симпатрического видообразования, является неизбежность скрещивания в Исходной полиморфной популяции на первых этапах дивергенции биологических рас, которое должно препятствовать обособлению подобных группировок. Крайний скепсис в отношении симпатрического видообразования проявляет Э. Майр, по мнению Которого интенсивность разрывающего отбора, которая необходима для симпатрического видообразования, должна значительно превышать реально наблюдаемую в природе.
Однако в пользу его реальности говорит формирование у некоторых видов устойчивых симпатрических биологических рас. Примером таких группировок могут служить питающиеся разными кормовыми растениями ивовая и березовая расы жуков-листоедов Lochmaea capreae, между которыми, по данным С.Г.Креславского, существует высокий уровень репродуктивной изоляции (97 %).
Существует еще несколько особых форм симпатрического видообразования, обеспечивающих чрезвычайно быстрое обособление дочерней формы от родительского вида: полиплоидизация, гибридогенез (возникновение в результате гибридизации аллополиплоидных форм, см. гл. 1) и так называемый стасигенез (или стасипатрическое видообразование). Последним термином М.Уайт обозначил обособление новых видов в результате крупномасштабных хромосомных мутаций (инверсий, транслокаций). Во всех этих случаях новый вид сразу оказывается генетически изолированным от родительского, обитая на той же самой территории, и сохраняет свою обособленность, если является жизнеспособным в данных условиях и способен к бесполому или партеногенетиче-скому размножению (в связи с этим такие формы видообразования более обычны у растений, чем у животных). Позднее, после накопления достаточного количества особей с однотипным геномом, может восстановиться половой процесс.
Таким образом, формы видообразования достаточно разнообразны, поскольку в рамки двух его основных категорий - аллопатрического и симпатрического - включен, с некоторой долей условности, ряд особых моделей видообразования, существенно отличающихся своими механизмами (так, симпатрическое видообразование включает гибридогенез, стасигенез, полиплоидизацию и дивергенцию биологических рас).
Темпы видообразования
Подобно формам видообразования столь же разнообразны и его темпы, варьирующие от относительно медленного и постепенного преобразования генофондов географически обособленных популяций при "классической" аллопатрической модели до чрезвычайно быстрого (в процессе смены всего нескольких поколений) обособления генофонда популяции основателей при "генетической революции" и действии катастрофического отбора, или столь же быстрого формирования нового вида при полиплои-дизации и стасигенезе.
В 1972 г. Н.Элдридж и С.Гулд выдвинули получившую широкую известность гипотезу так называемого "прерывистого равновесия", или "пунктуализма", которую они считают альтернативной классическим представлениям дарвинизма о постепенном ("градуалистском") видообразовании. Согласно этой гипотезе история любой филетической линии состоит из чередования длительных периодов эволюционной стабильности - до 99% времени существования вида - - и относительно кратких периодов видообразования: каждый акт видообразования охватывает около 10000-100000 лет. В качестве основного аргумента в пользу своей гипотезы Гулд и Элдридж использовали широко распространенное явление прерывистости филетических линий в палеонтологической летописи, обычно объясняемое фрагментарностью палеонтологических данных и приводящее к описанию дискретных во времени палеонтологических видов.
Сторонники пунктуализма видят основные механизмы ускоренного видообразования в процессах генетических революций, происходящих в периферических изолятах; в сериях макромутаций (особенно в мутациях генов-регуляторов и хромосомных перестройках) и в так называемом "видовом отборе". Этот термин используется для обозначения отбора по признакам, инвариантным (единообразным) для вида в целом. В этом случае виды рассматривают как аналоги особей, причем сторонники пунктуализма полагают, что "видовой отбор" должен приводить к быстрому замещению хуже приспособленных по данному признаку видов лучше приспособленными. Однако необходимо помнить, что отбор - это вероятностный процесс, эффективный лишь при достаточно больших выборках. Число конкурирующих видов обычно значительно меньше минимального числа особей в популяции, начиная с которого отбор становится более эффективным, чем дрейф генов. Отбор по инвариантным видовым признакам, конечно, может происходить, но он базируется не на видах как целостных единицах, а на большом числе составляющих их особей. Поэтому такой отбор ничем принципиально не отличается от обычного естественного отбора и не требует выделения в особую категорию.
Проверка многочисленных палеонтологических данных о характере видообразования в различных филетических линиях, проделанная целым рядом ученых, показала, что в палеонтологических рядах форм можно обнаружить и "градуалистское", и "пунктуалистское" видообразование. Вообще же, "разрешающая способность" палеонтологической летописи, как правило, совершенно недостаточна для того, чтобы по палеонтологическим данным можно было с достаточной достоверностью судить о характере процессов видообразования, поскольку "разрывы" между предками и потомками могут быть связаны как с высокими темпами видообразования, так и с неполнотой палеонтологической летописи.
Ускоренное видообразование, утверждаемое сторонниками пунктуализма, вообще говоря, вполне возможно, и механизмы его известны (это "генетические революции", стасигенез, поли-плоидизация, гибридогенез). Однако неверно главное положение гипотезы прерывистого равновесия, что это - единственная (или резко преобладающая) форма эволюции, чему противоречат многочисленные данные по современным и ископаемым формам.
Хотя сама концепция пунктуализма в целом не выдержала критики, дискуссии с ее сторонниками в 70-80-е гг. способствовали развитию представлений о многообразии форм и темпов видообразования.
Взаимоотношения близких видов
После того как между родственными популяциями сформировались какие-либо механизмы репродуктивной изоляции, эти популяции можно рассматривать как самостоятельные виды. При возникновении вторичного контакта и перекрывания ареалов таких форм (неосимпатрия), между ними могут сложиться различные взаимоотношения в зависимости от степени их экологической дивергенции и генетической изоляции.
Гибридизация. Если экологическая дивергенция уже столь велика, что близкие виды не вступают в остроконкурентные отношения, то степень генетической изоляции не является для них определяющим фактором: даже если виды могут скрещиваться, давая плодовитое потомство, полного воссоединения их генофондов обычно не происходит. Пример подобной ситуации был описан у ящериц-игуанид Sceloporus woodi и S. undulatus, ареалы которых перекрываются во Флориде. Хотя этологические механизмы, препятствующие спариванию представителей этих двух видов, отсутствуют и их гибриды плодовиты, зона гибридизации остается очень узкой, не превышая нескольких миль в ширину. Это связано с предпочтениями разных биотопов, использованием разных способов охоты и разных наборов кормов у двух указанных видов ящериц.
Подобная ситуация может, вероятно, сохраняться довольно долго, но при изменениях условий внешней среды в зоне контакта таких видов экологическая изоляция может оказаться малоэффективной, что приведет к усилению гибридизации родственных форм. Подобные ситуации уже многократно описаны в самых различных группах животных. Например, по данным Д.Джонса, в районе Блумингтона (Индиана, США) происходит регулярная гибридизация двух видов жаб Bufo americana и В. woodhousei fowleri.
К настоящему времени в результате гибридизации произошло морфологическое сближение обитающих здесь популяций обоих видов. Предполагают, что причиной гибридизации, до 1941 г. не отмечавшейся и, очевидно, усилившейся вторично, являются изменения внешней среды деятельностью человека (расчистка лесов, загрязнение водоемов).
Интересный пример усиления гибридизации между родственными видами описан в 1973 г. Л.С.Рябовым: в Воронежской области за последние 13 лет отмечено свыше 100 особей-гибридов волка и домашней собаки, причем наблюдается тенденция к увеличению числа этих гибридов и расширению зоны гибридизации. Известно, что обычные отношения волка и собаки в целом антагонистичны - волки нередко нападают на собак, используя их как добычу. Изменения взаимоотношений этих двух видов, по мнению Л. С. Рябова, связаны с сокращением численности волков и нарушением нормального соотношения числа самцов и самок в их популяциях.
В любом случае, если гибридизация двух видов происходит регулярно хотя бы в небольшой зоне и гибриды жизнеспособны и плодовиты, это должно приводить благодаря возвратному скрещиванию к проникновению некоторого количества генов одного вида в генофонд другого. Это явление получило название интрогрессивной гибридизации (или интрогрессии). Интрогрессивная гибридизация может способствовать усилению наследственной изменчивости родственных видов, обогащая их генофонды новыми аллелями. Э. Майр полагает, что при этом возможен и другой эффект -- проникновение путем интрогрессии новых аллелей в уже сложившиеся, сбалансированные и оптимизированные отбором генофонды популяций родственных видов может нарушить сбалансированность этих генофондов, способствуя снижению приспособленности отдельных особей и популяции в целом. В этом случае отбор должен действовать в пользу усиления механизмов репродуктивной изоляции, препятствующих гибридизации. Происходит завершение процесса дивергенции видов, начавшейся в условиях географической изоляции. Такой вариант видообразования, начавшегося как аллопатрическое и завершающегося в условиях неосимпатрии, иногда называют "парапатрическим".
При более отдаленном родстве скрещивающихся видов их генотипы уже настолько различны, что гомология хромосом во многом нарушена. У таких гибридов мейоз затруднен из-за невозможности нормальной конъюгации хромосом в профазе первого Деления, и гаметогенез протекает с существенными нарушения-Ми, так что жизнеспособными оказываются относительно немногие гаметы, в частности получившие полные (двойные) наборы Хромосом без какой-либо редукции их числа. При копуляции таких гамет во втором гибридном поколении возникают так называемые амфидиплоыды - организмы, имеющие диплоидные наборы хромосом от каждого из двух родительских видов. Цитологически амфидиплоид ведет себя как нормальная диплоидная особь, поскольку каждая хромосома приобрела себе гомолога. Поэтому ам-фидиплоиды второго гибридного поколения могут дать начало новому виду, отличающемуся от обоих родительских - если, конечно, эти амфидишюидные организмы окажутся жизнеспособными и смогут выдержать конкуренцию с обоими родительскими видами. Роль такого способа видообразования, по-видимому, сравнительно невелика, хотя в селекции этот способ используется для выведения новых форм сельскохозяйственных растений (см. с. 57).
Если же происходит обратное скрещивание гибридов первого поколения, полученных от подобной отдаленной гибридизации, при копуляции их диплоидных гамет с гаплоидными гаметами родительских видов получаются триплоидные особи, которые также могут дать начало новым формам организмов при условии их способности к вегетативному размножению (у растений), партеногенезу или гиногенезу (у животных). Таков был путь возникновения партеновидов у некоторых групп позвоночных. В частности, подобные партеновиды описаны в двух семействах ящериц - Lacertidae и Teiidae.
Иногда при отдаленной гибридизации возникают своеобразные комплексы взаимосвязанных форм, подобные комплексу бисексуальных диплоидных видов саламандр Ambystoma laterale и A. jeffersonianum с триплоидными самками, возникшими в результате их гибридизации ("A. platyneum") и размножающимися путем гиногенеза (с активацией триплоидных яиц к дроблению гаплоидными спермиями родительских видов). Еще один характерный пример подобных взаимоотношений представляет комплекс видов европейских зеленых лягушек: озерной (Rana ridibunda), прудовой (R. lessonae) и съедобной (R. esculenta). Как показал Л. Бергер, последняя форма возникла в результате гибридизации двух первых видов. Геном R. esculenta включает гаплоидные геномы R. ridibunda и R. lessonae; гаметы съедобной лягушки получают либо оба этих генома, либо лишь гаплоидный геном R. ridibunda. Популяции съедобной лягушки поддерживаются за счет скрещивания этой формы с прудовой лягушкой, а также за счет новых скрещиваний прудовых и озерных лягушек.
Очевидно, такие формы организмов, которые воспроизводятся только путем гиногенеза (Ambystoma platyneum) или гибридо-генеза (Rana esculenta), не являются настоящими видами; для их обозначения введен особый термин - "клептоны".
Конкуренция. При возникновении неосимпатрии близкородственных видов столь же часто возникает и ситуация, противоположная рассмотренной нами выше, - а именно, когда генетическая изоляция двух видов достаточно совершенна и гибридизация -не играет заметной роли, но экологическая дивергенция незначительна. В этом случае два вида имеют весьма сходные экологические потребности (местообитания, корма, убежища, места размножения и т. п.). В результате между близкородственными видами возникают остроконкурентные отношения, причем один вид нередко вытесняет другой. Этот аспект взаимоотношений между близкими видами был рассмотрен Ч.Дарвином в схеме дивергентной эволюции. В соответствии с выводами Дарвина отбор в данном случае направлен на увеличение экологических различий между родственными видами.
Собрано много примеров остроконкурентных, антагонистических взаимоотношений между близкими видами. Так, лягушка-бык (Rana catesbeiana), обитающая в восточной части Северной Америки и завезенная в Калифорнию в 1914-1920 гг., полностью вытеснила из большинства районов Калифорнии два туземных вида лягушек - R. aurora и R. boylii, ныне сохранившихся лишь в таких местообитаниях, которые не заняты лягушкой-быком. На мелких островках в Адриатическом море у побережья Далмации встречаются два вида ящериц - Lacerta melisellensis и L. sicula, причем распространение этих форм, как правило, имеет аллопатричес-кий характер. По-видимому, L. sicula начала колонизовать острова позднее, чем L. melisellensis, и, по данным М.Радовановича, постепенно вытесняет последний вид. Остроконкурентные отношения были описаны у двух видов саламандр-плетодонтид Plethodon cinereus и P. nettingi в восточной части Северной Америки: первый вид вытесняет второй из всех благоприятных местообитаний на более сухие скальные осыпи, будучи более крупным и выигрывая пищевую конкуренцию.
Однако в некоторых случаях экологическая конкуренция близких видов может и не приводить к обязательному вытеснению (конкурентному исключению) одного вида другим. Если лимитирующие ресурсы (т. е. те факторы внешней среды, которые служат основным объектом конкуренции, - пища, оптимальные местообитания, места размножения и т.п.) имеются в районе симпат-рии конкурирующих видов в достаточном количестве, то популяции этих видов могут сосуществовать в одном местообитании в определенном равновесии без особого обострения конкуренции между ними. Это, в частности, может быть обусловлено действием внутрипопуляционных механизмов регуляции численности вида-доминанта, которые ограничивают последнюю более низким уровнем, чем это допускается наличными ресурсами среды. В этом случае неиспользованные ресурсы позволяют обитать на этой же территории также популяциям более слабого в конкурентном отношении вида. Такая ситуация была описана на примере совместного обитания экологически близких видов лягушек Rana blythi и R. ibanorum в одних и тех же местообитаниях в Сараваке (на острове Борнео). Конкуренция между симпатрическими популяциями этих видов лягушек ограничивает их максимальную численность. Однако рост плотности популяций каждого вида не идет дальше определенного предела даже при низкой численности популяции вида-конкурента. Это связано с тем, что возрастание плотности популяции ограничивает собственный рост в большей степени, чем рост популяции вида-конкурента.
Конкурирующие виды могут сосуществовать в одном местообитании и в том случае, когда каждый из них имеет определенные преимущества в разные сезоны года (подобно разным морфам при видовом полиморфизме, см. гл. 2). Так, по данным А.Дондта, большая синица (Parus major) и синица-лазоревка (P. caeruleus) конкурируют за пищу и ночные убежища. В сезон размножения лазоревка выигрывает пищевую конкуренцию, потребляя более мелких гусениц и личинок и лишая большую синицу значительной части пищевых ресурсов. С другой стороны, в остальное время года большая синица имеет преимущество в конкуренции за убежища для ночевки. В результате между симпатрическими популяциями обоих видов устанавливается определенное равновесие.
Межвидовые отношения в биоценозах и коэволюция
Для понимания процессов видообразования и дальнейшей эволюционной судьбы видов организмов необходимо принимать во внимание не только взаимоотношения близких недавно дивергировавших форм, но и разнообразные взаимосвязи, складывающиеся между филогенетически далекими видами в природных сообществах - биогеоценозах.
По определению В. А. Ковды, биогеоценоз (или экосистема) представляет собой исторически сложившуюся совокупность растений, животных, почвы, рельефа, климата и вод в данном географическом районе. Нас интересуют здесь живые компоненты биогеоценоза, т.е. входящие в его состав виды организмов, сообщество которых называют биоценозом.
(Экосистема - термин, часто используемый как синоним биогеоценоза, имеет более широкое значение, поскольку применяется также для обозначения подсистем последнего и для искусственных комплексов организмов, например, аквариум с его обитателями).
Элементарными функциональными единицами биоценоза являются популяции разных видов организмов, группирующиеся в так называемые консорции - совокупности популяций, объединяемых трофическими (пищевыми) связями с определенными видами автотрофных растений. Органические вещества, синтезируемые растениями, и аккумулированная в этих веществах энергия передаются в консорциях по трофической цепи, включающей растения (продуценты), растительноядных животных (первичные потребители), хищников (вторичные потребители) и различных сапрофагов (третичные потребители -- трупоеды, копрофаги и т.п.). Конкретная величина потока энергии и органических веществ, передающегося по трофической цепи, определяет численность популяций каждого вида организмов, входящего в состав данной консорции. Биомасса первичных, вторичных и третичных потребителей вместе составляет около 1% биомассы продуцентов (растений).
Под влиянием естественного отбора биоценозы приобретают характер устойчивых саморегулирующихся систем с обратной связью, регуляция которых - ценотический гомеостаз - осуществляется посредством вчутрипопуляционных механизмов (отдельно для популяций каждого вида) и на уровне целого биоценоза путем так называемой канализации трофических цепей, под которой понимается повышение их устойчивости за счет развития резервных "обходных путей" для передачи потоков вещества и энергии при нарушении каких-либо звеньев основной цепи. Наличие механизмов саморегуляции придает биоценозу определенную целостность. Изменения популяции одного вида в той или иной степени сказываются на популяциях других видов, входящих в тот же биоценоз. Следовательно, эволюционные изменения каждого вида затрагивают и другие виды, связанные с первым в данном биоценозе и, благодаря связям разных биоценозов друг с другом, могут в какой-то степени отражаться и на популяциях некоторых видов, входящих в другие биоценозы. Кроме того, разные популяции одного вида обычно входят в состав разных биоценозов. Эволюция видов, входящих в один биоценоз, является согласованной (или "когерентной", по В. А. Красилову). Эволюция видов организмов тесно связана с эволюцией соответствующих биоценозов как целостных биологических макросистем. При этом биоценоз может быть более устойчивой системой, чем входящие в его состав отдельные виды. Механизмы ценотического гомеостаза позволяют биоценозам сохранять стабильность, несмотря на выпадение из их состава отдельных видов или их замещение новыми.
Положение и функциональная роль данного вида в биогеоценозе характеризуются в экологии понятием "экологическая ниша". Экологическая ниша определяется, с одной стороны, составом и й данного сообщества (в этом аспекте экологическая ниша представляет собой совокупность различных ресурсов, обеспечивающих возможность существования определенного вида в данном биогеоценозе), с другой - особенностями биологии данного вида. В соответствии с этим каждый вид организмов занимает в биогеоценозе определенную экологическую нишу, но не обязательно все ниши в ценозе заняты в данный момент его исторического развития.
Судьба новых видов, внедряющихся в уже сложившееся сообщество, и их воздействие на виды, входившие в его состав прежде, определяются наличием или отсутствием в биоценозе свободных (или хотя бы используемых не полностью) экологических ниш. Нередко интродукция нового вида приводит к угнетению и даже вымиранию тех или иных местных видов. Например, хорошо известно угнетающее воздействие на многие местные виды в биоценозах островов Тихого океана животных, завезенных на эти острова человеком, - черных и серых крыс, одичавших свиней, коз, собак и кошек. С другой стороны, известны случаи, когда новые виды широко распространяются и приобретают значительную численность в новых местообитаниях, не вызывая существенных нарушений в соответствующих биоценозах. Эффектный пример такого рода представляет широкое расселение египетской цапли (Bubulcus ibis) в биоценозах Южной, а затем Северной Америки. Предполагают, что "популяция основателей" египетской цапли была занесена из Старого Света в Америку в 80-е гг. XIX в. ураганом. Египетская цапля не конкурирует ни с какими американскими видами цаплевых, так как кормится главным образом насекомыми и другими беспозвоночными, собираемыми среди травы в наземных местообитаниях, тогда как большинство других видов цапель добывает разнообразную животную пищу (в основном позвоночных) на мелководье. До появления египетской цапли в Америке соответствующая экологическая ниша в местных биоценозах оставалась незанятой.
Проблемы, связанные с эволюционной ролью биоценозов, по сути дела, лишь намечены в современной науке. Раздел экологии, изучающий биоценозы и их эволюцию (биоценология), переживает бурное развитие. Установлены, в частности, некоторые общие тенденции эволюции биоценозов и определенные циклические изменения, происходящие в процессе этой эволюции (сукцессия биоценозов). "Молодые" биоценозы характеризуются относительно малым количеством составляющих их видов, биопродукция органических веществ в них намного превышает деструкцию, т.е. разрушение органических веществ. В молодых биоценозах преобладают силы движущего отбора, обусловливающие относительно быстрые эволюционные изменения. Постепенно
число видов в биоценозе возрастает, усложняются взаимосвязи различных популяций и увеличивается количество трофических цепей. Соответственно возрастают возможности саморегуляции биоценоза. В зрелых биоценозах, находящихся в состоянии климакса, биопродукция равна деструкции органического вещества, т.е. общая продукция фотосинтеза равна суммарному дыханию всей экосистемы. (Климакс - устойчивое состояние биоценоза, являющееся завершающим этапом сукцессии). В таком зрелом, хорошо сбалансированном биоценозе существует устойчивое равновесие между популяциями различных видов, "притертых" естественным отбором друг к другу. Это благоприятствует действию стабилизирующего отбора, который оказывает тормозящее воздействие на эволюцию.
Вместе с тем Л. Ван Вален и ряд других ученых полагают, что биотические компоненты природных экосистем продолжают эволюционировать и при остающихся стабильными абиотических условиях, поскольку достигнутая степень приспособленности организмов обычно относительна и для сохранения любым данным видом его положения в сообществе необходима непрерывная адаптивная эволюция, в процессе которой и поддерживается колеблющееся равновесие между популяциями разных видов. Эта концепция получила название "гипотезы Черной (или Красной, в англоязычной литературе) королевы" - по соответствующему персонажу сказки Л.Кэррола "Алиса в Зазеркалье", утверждавшему, что "нужно бежать изо всех сил, чтобы оставаться на том же месте".
Внутри биоценозов имеются пары и группы видов, связанных друг с другом более тесно, чем с остальными членами сообщества. Прежде всего это виды организмов, принадлежащие к одной консорции. Однако взаимоотношения разных видов очень разнообразны и отнюдь не сводятся к трофическим связям (например, использование организмами одного вида нор, гнезд или тела особей другого вида как среды обитания; мимикрия, "гнездовой паразитизм", подобный таковому у кукушек, и т.п.). Эволюционные взаимодействия видов организмов, не обменивающихся друг с Другом генетической информацией, но тесно взаимосвязанных биологически, называют коэволюцией. В сущности, эволюция любого вида организмов протекает как коэволюция с теми или другими видами - партнерами по биоценозу (растения и Растительноядные животные, хищники и их жертвы, паразиты и их хозяева, различные формы симбиоза-мутуализма и др.). Многие черты конкретных филогенетических преобразований различных видов организмов можно понять, только учитывая их коэволюционные взаимосвязи. Один из наиболее известных и простых примеров этого - развитие цветка у опыляемых насекомыми покрытосеменных растений.
Поскольку развитие взаимных приспособлений благоприятно для обоих взаимодействующих видов, отбор способствует коэво-люционным изменениям. При кажущейся антагонистичности членов таких коэволюционных пар, как хищник - жертва или паразит - хозяин, коэволюция всегда обусловлена совместимостью партнеров в биоценозе и ведет к развитию таких взаимоотношений между ними, при которых виды-партнеры становятся взаимно необходимыми. Многочисленные работы экологов убедительно показали, что хищники и паразиты, регулирующие численность популяций своих жертв и выбраковывающие среди них в первую очередь больных, ослабленных, генетически неполноценных особей, являются необходимым фактором, обеспечивающим оптимальное состояние популяций видов-жертв.
Коэволюционные межвидовые взаимодействия могут продолжаться длительное время, приводя к глубоким изменениям взаимодействующих видов.
Разрушение биоценоза благодаря каким-либо внешним воздействиям (изменения условий внешней среды, превосходящие возможности саморегуляции сообщества) приводит к распаду сложившихся систем межвидовых взаимодействий и открывает простор для действия интенсивного движущего отбора, который может принять характер катастрофического. При этом утрачивается регулирующее воздействие биоценоза на эволюцию входящих в его состав видов, которая становится несогласованной ("некогерентной", по В. А. Красилову). Темпы видообразования в отдельных филетических линиях могут при этом значительно возрастать.
Некоторые исследователи (В.В.Жерихин, А.С.Раутиан и др.) полагают, что биоценотические кризисы могут возникать и без существенных изменений абиотических факторов внешней среды, развиваясь на основе специфических взаимодействий входящих в сообщество видов организмов, которые эволюционируют и в относительно стабильных условиях в соответствии с "принципом Черной королевы", по Ван Валену. Кризис сообщества может возникнуть в результате прогрессирующей и далеко зашедшей (при длительно остающихся стабильными внешних условиях) специализации большинства видов данного биотического комплекса, снижающей устойчивость сообщества. Относительно кратковременные неблагоприятные изменения внешних условий, которые могут выдержать сбалансированные биоценозы, стимулируют начало кризисных процессов в сообществах, состоящих в основном из специализированных видов при дефиците видов-универсалов, которые могли бы занять в трофических цепях места, освободившиеся при вымирании специализированных форм.
Термин "приспособление" (или адаптация) применяется в современной науке в разных значениях. В широком смысле адоптациями являются любые особенности строения, физиологии, поведения и онтогенеза данного вида, которые обеспечивают (вместе с другими особенностями) возможность специфического образа жизни в определенных условиях внешней среды или определенный способ использования конкретных ресурсов внешней среды. Так, для хищных птиц приспособлениями к хищничеству являются характерная форма клюва и когтей, определенный набор пищеварительных ферментов, необходимых для эффективного переваривания животной пищи, и соответствующие формы поведения, используемые при охоте на различных животных (птиц, млекопитающих, рептилий и др.). Иногда адаптацией называют также процесс развития приспособленности организмов.
Единственный фактор, придающий эволюционным изменениям организмов приспособительный характер, - естественный отбор - непосредственно действует в видовых популяциях, т.е. на уровне микроэволюции. Однако его кумулятивное (накапливающее) действие в полной мере проявляется лишь в процессе смены многих поколений на больших отрезках филетических линий, т.е. в масштабе макроэволюции. Именно на этом уровне из част-Ньк адаптации (незначительных изменений фенотипа, приспосабливающих организмы данного вида к особенностям условий его обитания) формируются адаптации общего значения - сложите морфофункциональные комплексы, обеспечивающие жизнедеятельность организмов в широком диапазоне условий. Частные адаптации специфичны для отдельных видов, тогда как адаптации общего значения характеризуют крупные таксоны (отряды, классы и т.п.) и присущи всем видам, входящим в их состав. Например, летательный аппарат птиц, включающий скелет и мускулатуру передних конечностей, а также перья, образующие несущую поверхность крыла, представляют адаптацию общего значения, тогда как особенности строения этих структур у отдельных видов являются частными адаптациями.
Сложные приспособления и проблема их развития
Каждый вид организмов обладает теми или иными приспособлениями, позволяющими ему существовать в определенных условиях среды. Наличие приспособлений придает строению и жизнедеятельности организмов черты целесообразности, которая всегда обращала на себя внимание естествоиспытателей. Невольно поражают воображение сложность и совершенство таких адаптации как, например, взаимные приспособления цветковых растений и опыляющих их насекомых. Сюда относятся характерные особенности строения и окраски венчика цветов и соответствующие особенности строения челюстного аппарата и конечностей насекомых-опылителей, а также черты их поведения. Поразительны явления мимикрии, или подражательного сходства, когда один из видов имитирует особенности внешнего строения и окраски другого, "маскируясь" под несъедобный или ядовитый для хищника объект: "подражание" листу в форме и окраске крыльев или имитация стебля растений у насекомых-палочников (Phasmoptera) (рис. 18), "подражание" неядовитых видов ядовитым или хорошо защищенным (например, уже упоминавшееся выше сходство самок бабочки-парусника с несъедобными бабочками-данаидами, см. рис. 5) и т.п.
Теория Ч.Дарвина впервые дала научное решение проблемы происхождения адаптации как неизбежного результата естественного отбора.

На заре мутационизма многие ученые склонялись к мысли, что новые приспособления возникают сразу готовыми - как случайный результат мутаций крупного масштаба. Подобные воззрения позднее развивал Р. Гольдшмидт, считавший, что крупные "системные мутации" резко изменяют конституцию организма, приводя к появлению "многообещающих уродов" (hopeful monsters), которые могут случайно оказаться приспособленными к новым условиям существования. Эта концепция маловероятна как с генетической, так и с экологической точки зрения. Как уже отмечалось, сколько-нибудь значительные мутации обычно летальны, поскольку они приводят к нарушениям нормальной интеграции организма на том или ином этапе онтогенеза. Маловероятно возникновение посредством крупной мутации такого нового фенотипа, который оказался бы не только жизнеспособным, но и плодовитым, а к тому же априорно приспособленным к каким-либо конкретным условиям существования. В связи с этим Э.Майр справедливо заметил, что организмы, которые возникнут в результате крупных мутаций, скорее окажутся "безнадежными уродами" (hopeless monsters), чем "многообещающими", по Р.Гольдшмидту.
Но наиболее важным возражением против существенной роли крупных мутаций в возникновении новых типов организации является невозможность объяснить этим путем формирование сложных комплексных адаптаций, образованных разнообразными компонентами, развитие которых кодируется разными генетическими системами и обеспечивается независимыми цепями морфо-генетических процессов в онтогенезе. Именно таковы многие адаптации общего значения, составляющие основной план организации высших таксонов. Например, слуховой анализатор у высших наземных позвоночных (амниот) включает структуры, имеющие разное эволюционное происхождение и формирующиеся в онтогенезе разным путем из различных зачатков (рис. 19). Так, внутреннее ухо развивается из слуховой плакоды (утолщение зародышевой эктодермы); полость среднего уха - из зачатка рудиментарной жаберной щели (брызгальца); слуховые косточки среднего уха (звукопроводящий аппарат) - из элементов подъязычной и челюстной висцеральных дуг, возникающих, в свою очередь, из материала так называемой ганглионарной пластинки эктодермального происхождения; скелет слуховой капсулы - из скелетогенной мезодермы головы; наконец, слуховой нерв и мозговые центры слухового анализатора формируются из невральной эктодермы. Совершенно ясно, что невозможно возникновение всей этой сложной, но целостной системы как путем случайного подбора различных мутаций (например, при дрейфе генов), так и посредством одной крупной "системной" мутации. Несомненно, естественный отбор играет решающую роль в накоплении мутаций и интеграции их фенотипических проявлений, обусловливая развитие приспособлений организмов.

Однако в этой проблеме имеется ряд сложных вопросов, и некоторые из них отчетливо представлял уже Ч.Дарвин. Первая трудность связана с тем, как отбор может контролировать начальные этапы развития нового приспособления, когда новая структура (или функция) еще столь слабо выражена, что ее небольшие изменения не могут принести организму каких-либо заметных выгод. Пытаясь разрешить эту проблему, Ч.Дарвин в некоторых случаях даже пытался привлечь "унаследование последствий употребления и неупотребления органов" (например, для объяснения развития асимметричного расположения глаз у камбал или развития цепкого хвоста у южноамериканских обезьян). Очевидно, такое объяснение было шагом назад по отношению к теории естественного отбора и не может быть признано удовлетворительным.
Особенно трудно объяснить действие отбора на первых этапах развития таких сложных органов, которые вообще могут эффективно функционировать только при достаточно высокой степени своего развития. Таковы, например, уже упоминавшиеся нами сложные адаптации - летательный аппарат птиц, звукопроводящая система среднего уха наземных позвоночных, плавательный пузырь костистых рыб, грудная клетка амниот в роли легочного воздушного насоса и т.п. Трудно понять, какую приспособительную ценность для организма могут иметь первые зачатки подобных сложных органов и как на них будет действовать отбор, поскольку естественный отбор не может "предвидеть" будущего развития данного признака и его будущего значения для организма.
Развитие приспособлений на основе преадаптаций
Путь к решению этой проблемы помогает найти концепция морфофункциональных преадаптаций. Сам термин "преадаптация" принадлежит французскому ученому Л. Кено. Кено понимал под преадаптациями такие особенности организмов, которые возникают как случайные вариации, первоначально безразличные для организма (т.е. не приносящие ему ни вреда, ни заметной пользы), нo после изменения условий существования или образа жизни Данного вида оказываются готовыми приспособлениями, полезными в новых условиях. Очевидно, эти представления Кено о развитии приспособлений из случайных преадаптаций без участия естественного отбора примыкают к воззрениям ранних генетиков-мутационистов (см. с. 44). Однако в концепции Кено было все же рациональное зерно, заключавшееся в предположении, что Сложные приспособления к каким-либо конкретным условиям Могут первоначально развиваться в совершенно иной среде обитания и выполнять другую роль, и лишь позднее - в новых условиях - принять на себя новые, прежде им не свойственные функции. Кено справедливо указывал, что освоение новой среды обитания любым видом организмов возможно только при наличии у него таких особенностей строения, физиологии и поведения, которые дают возможность выжить в новых условиях, но возникнуть эти особенности могли только до начала освоения нового образа жизни, т. е. в прежней среде обитания. Такие особенности и являются преадаптивными.
Кено обратил внимание на то, что, в сущности, уже Дарвин подошел к идее преадаптации, анализируя в "Происхождении видов" развитие некоторых приспособлений. Так, швы между костями черепа млекопитающих облегчают прохождение головы детеныша при родах, однако наличие таких сочленений между костями черепа связано совсем не с живорождением, и возникли эти соединения костей в черепе древних позвоночных задолго до появления живорождения. "Лазающие пальмы" Малайского архипелага имеют приспособления в виде крючков, позволяющие им удерживаться на ветках и стволах деревьев, взбираясь по ним вверх. Но точно такие же крючки существуют и у многих нелазающих родственных видов растений, которым они служат как защитные приспособления.
За время своего существования концепция преадаптации претерпела значительные изменения, и ныне ее положения существенно отличаются от взглядов Л. Кено. В первую очередь это относится к отказу от наивных представлений, что преадаптации являются уже готовыми приспособлениями, возникающими во всей их сложности чисто случайно, без участия естественного отбора.
Концепция преадаптации охватывает самые различные процессы и явления, связанные с развитием у организмов новых приспособлений и принятием органами новых функций. Эти процессы и явления могут относиться к особенностям строения, физиологии, индивидуального развития, поведения и образа жизни организмов. В наиболее общем смысле преадаптациями называют такие свойства организмов, которые потенциально имеют приспособительную ценность для еще не осуществленных форм взаимодействий организма и среды или для новых способов функционирования отдельных органов. Так же как и в отношении термина "адаптация", преадаптацией называют и состояние организма (или органа), и процесс развития преадаптивного состояния. Процесс усовершенствования уже существующих приспособлений к уже освоенной среде обитания (или - более узко - процесс совершенствования прежних способов функционирования отдельных органов) называют постадаптацией.
Современная концепция преадаптации включает два основных аспекта - генетико-экологический и морфо-функциональный. Первый из них стоит ближе ко взглядам Л. Кено.
При генетико-экологическом подходе в качестве преадаптивных рассматриваются такие нейтральные и "слабовредные" особенности фенотипа, которые поддерживаются мутационным процессом, эффектом плейотропии, коррелятивными взаимосвязями в организме и тому подобными механизмами и оказываются адаптивными при изменении условий внешней среды. В сущности, ге-нетико-экологическая концепция преадаптации сводится к представлениям о возможности использования видом первоначально неприспособительных признаков (возникших на основе случайных мутаций) для развития адаптации при изменениях направления отбора. Сами по себе эти представления не вызывают возражений, но, в сущности, этот подход является малопродуктивным, так как конкретный анализ развития преадаптивных признаков в этом аспекте отсутствует. Кроме того, генетико-экологическая концепция преадаптации выпускает из поля зрения творческую роль естественного отбора, за что она нередко подвергалась критике. (Отметим, что взгляды сторонников генетико-экологической концепции преадаптации или даже взгляды Л.Кено и других ранних мутационистов часто критикуются в качестве "теории преадаптации". которая к этим взглядам отнюдь не сводится).
Морфофункциональные преадаптации и смена функций
В отличие от генетико-экологической концепции, рассматривающей возникновение преадаптивного состояния на основе случайного подбора мутаций, морфофункциональная концепция, разработанная Д. Симпсоном и В. Боком, подчеркивает развитие преадаптации на базе прежних приспособлений, под непосредственным и непрекращающимся контролем естественного отбора. Усовершенствование приспособлений к существующим условиям может оказаться преадаптацией к новым условиям среды.
Морфофункциональная концепция преадаптации позволяет разрешить ту проблему, о которой мы упоминали в начале этой главы, - развитие сложных приспособлений, способных эффективно функционировать лишь будучи вполне сформированными. Такие приспособления (подобные челюстному аппарату, среднему уху, плавательному пузырю и др.) формируются в эволюции из существовавших прежде органов, имевших иное приспособительное значение. Соответствующие органы сначала приобретали новую функцию (дополнительную к их прежней основной Функции), а затем, если новая функция оказывалась более важной, происходила смена функций органа. Принцип смены функций органов в эволюции был сформулирован А.Дорном в 1875г. Такой путь филогенетических изменений возможен лишь в тощ случае, если имеется орган, подготовленный своей предшествующей эволюцией к принятию новой, прежде не свойственной ему функции, т.е. орган, находящийся в преадаптивном состоянии. Морфофункциональная концепция преадаптации анализирует эволюционный механизм смены функций органов.
В этом аспекте преадаптивное состояние организма (или отдельного органа) возникает не как непосредственный результат случайных мутаций, а на основе предшествующей приспособительной эволюции. Естественный отбор постепенно совершенствует уже существующие приспособления организмов, благоприятствуя тем вариантам, которые обеспечивают более эффективное и экономичное функционирование различных органов. В некоторых случаях при этом может возникнуть как случайный, побочный эффект приспособительной эволюции преадаптивное состояние органа, когда последний получает возможность принять новую функцию в дополнение к его прежней функции. При этом орган получает и новое приспособительное значение (новую адаптивную роль). Если новая адаптивная роль достаточно важна для организма, дальнейшая эволюция данного органа будет направлена отбором по пути совершенствования этого нового приспособления (рис. 20).
Эволюционный момент, когда орган принимает новую функцию и вследствие этого получает новую адаптивную роль, В.Бок назвал преадаптационным порогом. Прежние функция и адаптивная роль органа могут сохраниться и после принятия им новых. Тогда дальнейшие эволюционные изменения органа определяются компромиссом между двумя направлениями отбора, контролирующими старую и новую адаптации.
Важной особенностью механизма морфофункциональной преадаптации является его пороговый характер: после достижения преадаптационного порога при высокой приспособительной ценности новой адаптивной роли направление эволюционных перестроек данного органа может резко измениться. Одновременно открываются новые возможности в использовании ресурсов внешней среды, что создает предпосылки для интенсивного видообразования, приводящего к возникновению разнообразных вариантов нового конструктивного типа.
Поскольку орган подготовлен к принятию новой адаптивной роли преобразованиями, направленными на усовершенствование прежнего приспособления, т.е. постадаптациями по прежней функции, можно говорить, что преадаптация органа к новой адаптивной роли развивается на основе постадаптаций к прежней.
Потребность в новом приспособлении может существовать у организмов задолго до того, как возникает подходящая преадаптация. Нужно подчеркнуть, что такая потребность далеко не всегда реализуется в эволюции. Поскольку преадаптация возникает как случайный побочный эффект приспособительных эволюционных изменений органа - очевидно, что гораздо чаще должно иметь место как раз отсутствие требуемых преадаптации, чем их развитие. Например, отсутствие предпосылок для вселения вида в новую среду обитания в целом гораздо более обычно, чем наличие требуемых для этого преадаптации.
Целый ряд поразительных примеров смены функций, иллюстрирующих разные формы морфофункциональных преадаптаций, демонстрируют эволюционные преобразования висцерального скелета позвоночных животных. У предков позвоночных (протокраниот, по А.Н.Северцову) висцеральный скелет, вероятно, состоял из серии цельных (нерасчлененных на отделы) хрящевых колец - жаберных дуг, охватывавших передний отдел пищеварительной трубки (рис. 21) и препятствовавших спадению ее стенок. При активизации образа жизни для этих животных стала необходимой интенсификация дыхания. Последняя была достигнута посредством усиления тока воды через жаберную область с развитием здесь нагнетательно-всасывающего насоса. У бесчелюстных позвоночных (предков современного класса круглоротых - Cyclostomata) жаберные дуги потеряли свою первоначальную независимость, соединившись в единую жаберную решетку продольными хрящевыми балками. При вентиляции жабер жаберная решетка, образованная гибкими хрящами, сжимается (при сокращении специальных мышц) и затем расправляется (за счет эластичности хрящевых жаберных дуг) как целое. Этот способ вентиляции жаберной области был, вероятно, связан с развитием энтодермальных жабер, расположенных внутри от жаберных дуг.

Совершенно иным путем происходила интенсификация обмена воды в жаберной области у предков челюстноротых позвоночных, у которых жабры имеют эктодермальное происхождение и располагаются снаружи от висцерального скелета (см. рис. 21). У этих животных сохранилась самостоятельность отдельных жаберных дуг, и каждая из них расчленилась на несколько подвижных отделов. Сокращение жаберных мышц складывает каждую жаберную дугу как гармошку, энергично сжимая жаберную область. Этот путь эволюции оказался преадаптивным для возникновения челюстей. Передние висцеральные дуги при наличии сильных мышц, сжимающих "гармошку", получили возможность способствовать удерживанию добычи. Подчеркнем, что эта новая адаптивная роль была приобретена на основе преадаптации, связанной с усовершенствованием прежней (дыхательной) роли висцеральных дуг. Новая адаптивная роль (захват и удерживание добычи) оказалась столь важной, что прежняя роль (участие в дыхательных движениях) у передней жаберной дуги (челюстной) отступила на второй план, а затем была утрачена (рудиментарная жабра челюстной дуги сохраняется у всех рыб в виде так называемой ложножабры, теряющей непосредственную связь с челюстной дугой и находящейся у костных рыб на внутренней поверхности жаберной крышки).
Так жаберная дуга превратилась в челюсти - орган схватывания и удерживания добычи. Наличие челюстей стало одним из важнейших преимуществ, способствовавших победе челюстноротых позвоночных над бесчелюстными в борьбе за существование. Среди современных позвоночных челюстноротые составляют примерно 99,8% от общего числа видов, а круглоротые - лишь 0,2 %. Возможно, челюсти не могли развиться у предков круглоротых из-за объединения жаберных дуг в единую жаберную решетку. В этом случае мы наблюдаем как раз такую ситуацию, когда в эволюции большой группы животных (бесчелюстные позвоночные) не была реализована биологическая потребность в развитии челюстей - важного органа добывания пищи, защиты и нападения, так как не возникло необходимой преадаптации.
После прохождения преадаптационного порога в эволюционных изменениях челюстной дуги у челюстноротых позвоночных начался период постадаптаций, усовершенствовавших приспособления к схватыванию и удерживанию добычи. Эти постадаптации привели к возникновению задней опоры челюстной дуги в области челюстного сустава на следующую сзади висцеральную дугу (подъязычную), верхний элемент которой (гиомандибулярный) стал подвеском челюстей. Однако подъязычная дуга сохранила и свою прежнюю роль - участие в вентиляции жаберной области, и у хрящевых рыб несет даже жаберные лепестки. Очевидно, здесь мы имеем пример эволюционного компромисса между двумя направлениями отбора, одно из которых требует сохранения и усовершенствования прежней дыхательной функции подъязычной Дуги, другое - развития новой опорной функции. Оба эти направления отбора контролировали дальнейшие эволюционные преобразования подъязычной дуги у рыб, с одной стороны, обусловив развитие механизма жаберной крышки у костных рыб, позволившего еше более интенсифицировать вентиляцию жаберной области, с другой -- вызвав появление различных механизмов Подвески челюстей к черепу, связанных с разными способами Добывания пищи (амфистилия, гиостилия и др.).
Рассмотренное в этом примере преадаптивное состояние органа возникло в результате усовершенствования его прежней функции; такую форму преадаптации можно назвать собственно (в узком смысле) морфофункциональной. В других случаях преадаптивное состояние может развиваться в процессе редукции органа после утраты его прежней адаптивной роли - редукционная преадаптация. Наконец, третью категорию составляют онтогенетические преадаптации, возникающие на основе эмбрионального или личиночного состояния некоторых органов, сохраняющегося во взрослом организме, в результате задержки онтогенеза на соответствующих стадиях.
Явление редукционной преадаптации можно проиллюстрировать процессом эволюционного развития звукопередающей системы среднего уха у наземных позвоночных-тетрапод. У предков земноводных возник аутостилический череп, в котором гиомандибулярный элемент подъязычной дуги утратил прежнюю роль подвеска челюстей; кроме того, редукция жаберного дыхания сопровождалась прекращением деятельности вентиляционного механизма жаберной крышки, которая также редуцировалась. Потеряв обе свои прежние функции, гиомандибулярный элемент также подвергся некоторой редукции. Этот элемент располагается в черепе сбоку от слуховой капсулы, его отростки упираются в стенку последней и в накладные кости височной области черепа, под которыми здесь располагается полость - рудимент жаберной щели между челюстной и подъязычной дугами (брызгальце). В воздушной среде эта полость оказывается заполненной воздухом; ее наружная стенка после редукции жаберной крышки становится относительно тонкой и, вероятно, могла вибрировать в ответ на колебания окружающего воздуха, как барабанная перепонка. Гиомандибуляре, упиравшееся в эту перепонку и в стенку слуховой капсулы, передавало трансформированные колебания воздуха ко внутреннему уху. Так гиомандибуляре функционально превратилось в первую слуховую косточку среднего уха - слуховой столбик или стремечко. В дальнейшем постадаптации усовершенствовали звукопередаюшие свойства этого элемента. Вероятно, подобная же редукционная преадаптация произошла и у предков млекопитающих после возникновения вторичного челюстного сустава при последовавшей затем редукции костей первичного челюстного сустава - квадратной и сочленовной, превратившихся в наковальню и молоточек среднего уха.
Примером эволюционного использования онтогенетических преадаптации может служить сохранение эмбрионального хрящевого скелета во взрослом состоянии, характерное для некоторых групп позвоночных (хрящевые рыбы, круглоротые, отчасти бесхвостые и хвостатые земноводные). В приспособительном отношении это было, вероятно, связано с необходимостью облегчения скелета (см. ч. III) при активизации образа жизни или при выходе на сушу.
Для освоения новой среды обитания, существенно отличающейся от прежней, организмы должны обладать целым комплексом особенностей строения, физиологии и поведения, позволяющих им существовать в этой новой среде. Такое состояние называется комплексной или конституциональной преадаптацией. Например, древнейшие земноводные, начавшие освоение суши Как среды обитания, обладали таким строением конечностей, органов дыхания и кровообращения и специфическими особенностями образа жизни и поведения, вся совокупность которых была необходима для выхода на сушу из водоемов. Все эти особенности, составившие комплексную преадаптацию, развились как специфические приспособления предков земноводных - кистеперых рыб - к жизни в пресных водоемах тропического пояса, в которых частым явлением было значительное уменьшение растворенного в воде кислорода (см. ч. III). В комплексной преадаптации, в сущности, объединяются морфофункциональный и генетико-экологический аспекты преадаптадии. Как указал Г.Оше, комплекс уже сложившихся морфофункциональных преадаптации для своей реализации должен быть завершен некоторыми деталями, которые могут контролироваться немногими ключевыми мутациями. Например, эти мутации могут вызвать такие изменения физиологии или поведения, которые позволят данному виду начать освоение новой среды обитания. Это и преобразует комплекс преадаптации в новое приспособление.
В связи с этим упомянем еще один интересный пример преадаптации, замечательный тем, что он реализовался не в далеком прошлом, как упоминавшиеся выше, а в наше время. Такие особенности строения попугаев, как крупный острый клюв с изогнутым вниз надклювьем (рис. 22), крепкие острые когти, мощная мускулатура челюстного аппарата, шеи и конечностей, выглядят хорошо приспособленными для хищного способа питания. Однако все эти признаки, несомненно, сформировались как приспособления к лазанию По деревьям: при лазании клюв также используется как хватательный орган, своего Рода "пятая конечность", тем более что надклювье подвижно сочленяется с черепом, и клюв как целое может активно подгибаться вниз или отгибаться вверх (что нетрудно заметить, наблюдая за лазанием крупных попугаев по решетке клетки). Подавляющее большинство попугаев - мирные птицы, питаЮщИеся растительной пищей, главным образом различными плодами и семенами.

Однако известен один вид попугаев, который действительно использовал указанные признаки при освоении хищного образа жизни. Попугай кеа (Nestor notabilis) (см. рис. 22), обитающий в горных районах южного острова Новой Зеландии, питается обычно различной растительной пищей (фрукты, нектар цветов), а также мелкими беспозвоночными животными (черви, насекомые) которых добывает из-под камней. Однако начиная с 1868 г. были отмечены случаи нападения кеа на овец, пригоняемых на горные пастбища. Хищнические наклонности проявляют отдельные особи попугаев: они расклевывают на спине овец огромные раны (до 10 см), от которых жертва быстро погибает. Мясо погибших овец поедают не только птицы-убийцы, но и другие кеа. Поскольку овцеводство появилось в Новой Зеландии после прихода туда европейцев (до этого на острове не было других млекопитающих, кроме крыс и летучих мышей), очевидно, что переход попугаев кеа к хищничеству произошел, можно сказать, на наших глазах. При этом строение клюва и когтей попугаев не претерпело никаких изменений - их использование для новой роли было преадаптировано прежними приспособлениями, облегчавшими попугаям лазание по деревьям и питание плодами растений. Попугаи, нападающие на овец, ничем не отличаются от других кеа. Возможно такое изменение поведения обусловлено возникновением соответствующей мутации, позволившей использовать по-новому существовавшие прежде структуры.
Необходимо подчеркнуть, что путь преадаптации - широко распространенный, но не единственно возможный способ развития новых приспособлений. Некоторые структуры, особенности физиологии и поведения могут с самого начала формироваться как "постадаптации" (здесь термин чисто условный, поскольку отсутствует период преадаптации). Условием для этого является такая приспособительная ценность уже первых зачатков новой структуры, которая достаточна для возникновения положительного давления естественного отбора, благоприятствующего новому признаку. Примером сложного органа, возникшего таким путем, является глаз, как это убедительно показал Ч.Дарвин.
Относительность приспособленности и инадаптивная эволюция
При высокой степени совершенства многих адаптации еще Ч.Дарвин подчеркивал относительность приспособленности и воз можность в большинстве случаев появления лучше приспособленных организмов, чем уже существующие. При отсутствии конкуренции приспособления совершенствуются отбором не максимально возможного, а до некоторого удовлетворительного уровня, после чего интенсивность отбора существенно падает. Достигаемая в каждом конкретном случае степень совершенства любого приспособления определяется, с одной стороны, особенностями организации данного вида, с другой же - интенсивностью отбора, в свою очередь зависящей от степени соответствия видовых приспособлений существующим внешним условиям. Кроме того, развитие разных адаптации у данного вида организмов может в какой-то мере противоречить друг другу, ставя различного рода взаимные ограничения. В подобных ситуациях отбор благоприятствует формированию некоторого оптимального соотношения между разными адаптациями, а не достижению максимального совершенства той или другой из них (принцип адаптивного компромисса, по К.Гансу).
Относительность приспособленности организмов ярко проявляется в многочисленных случаях неудачного решения тех или иных эволюционных проблем в филогенезе различных групп организмов. Такой "неудачный выбор" направления эволюционных преобразований, который может раньше или позже привести к эволюционному тупику, к назреванию неразрешимых или трудноразрешимых внутренних противоречий, был назван В.О.Ковалевским в 70-е гг. XIX в. инадаптивной эволюцией.
В.О.Ковалевский проанализировал это явление на примере филогенетических преобразований конечностей некоторых парнокопытных млекопитающих. В эволюции различных групп копытных в связи с приспособлением к быстрому бегу по относительно твердой почве происходила редукция боковых пальцев. Эта редукция у разных филетических линий ископаемых парнокопытных осуществлялась по-разному. У некоторых форм (Dicotyles, Gelocus, Sus) редукция пальцев происходила вместе с соответствующими перестройками в запястье и предплюсне, так что вес животного передавался на усиливающиеся средние пальцы (третий и четвертый), а конечность в целом сохраняла прочность и устойчивость (адаптивный путь изменений). У других же парнокопытных (Anoplotherium, Anthracotherium, Entelo-don, Hyopotamus, Xiphodon) редукция боковых пальцев опережала перестройку запястья и предплюсны, сохранявших более примитивное строение (рис. 23). В этом случае возникала механически несовершенная конструкция конечности, которая не обеспечивала равномерного распределения нагрузок в скелете при дальнейшей редукции боковых пальцев (инадаптивный путь). Преобразования конечностей по инадаптивному пути происходили в целом быстрее, чем более сложные гармонические перестройки по адаптивному пути. В результате этого инадаптивный путь изменений способствовал более раннему расцвету соответствующих филетических линий, но в дальнейшем при конкуренции с представителями более медленно, но гармонично развивавшихся линий инадаптивные формы вымерли.
При анализе подобных ситуаций неизбежно возникает вопрос: как объяснить развитие инадаптивных изменений? Не противоречит ли "инадаптивная эволюция" представлениям о развитии приспособлений под контролем естественного отбора? Однако противоречия здесь нет. Дело в том, что на первых этапах эволюционных преобразований многих органов (в данном случае - в процессе редукции боковых лучей конечностей у парнокопытных) и адаптивные, и инадаптивные варианты изменений могут быть примерно в равной степени полезны для вида. Естественный отбор благоприятствует любому изменению, повышающему приспособленность организмов, вне зависимости от того, как пойдет развитие этого признака в дальнейшем. Строго говоря, термин В.О.Ковалевского "инадаптивная", т.е. неприспособительная, эволюция неудачен, так как отбираются только приспособительные (адаптивные) изменения организмов. Однако в ходе дальнейшей эволюции обнаруживается преимущество одного пути эволюционных преобразований перед другими. Механическое несовершенство конечностей "инадаптивных" форм копытных выявилось лишь при достаточно далеко зашедшей редукции боковых пальцев, а в начале этого процесса указанные формы, вероятно, ничем не уступали представителям "адаптивных линий".
Таким образом, можно назвать инадаптивным такой путь филогенетических изменений, при котором формирующееся приспособление заключает в себе предпосылки для возникновения тех или иных внутренних противоречий, которые нарастают мере развития приспособления, приводя в конечном итоге к эволюционному тупику. Как показал В.О.Ковалевский, на начальных стадиях развития инадаптивные изменения могут даже и определенные преимущества перед более медленно протекающими гармоническими преобразованиями - именно благодаря быстроте развития инадаптивных вариантов. Гармонические преобразования требуют большего числа эволюционных "проб и ошибок" и соответственно большего времени для своего осуществления. Вероятно, можно сказать, что быстрота эволюционных преобразований нередко оказывается обратно пропорциональной степени их совершенства.
Инадаптивный путь эволюции не обязательно ведет к вымиранию - возникшие противоречия могут быть различными способами компенсированы. Рассмотрим, например, эволюционные преобразования сердца у рептилий завропсидного ствола (черепахи, клювоголовые, ящерицы, змеи, крокодилы). У предков завропсидных пресмыкающихся в желудочке сердца возникла горизонтальная перегородка, разделившая его на спинную и брюшную камеры и способствовавшая распределению артериальной крови в дуги аорты, а венозной - в легочную артерию (рис. 24). Однако полное разделение желудочка этой перегородкой, которое совершенно прекратило бы смешивание артериальной и венозной крови, невозможно - при этом легочная артерия, начинающаяся от брюшной камеры желудочка, оказалась бы отрезанной от обоих предсердий, открывающихся в спинную камеру. В результате у большинства пресмыкающихся перегородка в желудочке сердца остается неполной и сохраняется смешивание артериальной и венозной крови в сердце, что препятствует интенсификации метаболизма до уровня, необходимого для достижения гомойотермии (независимости температуры тела от температуры внешней среды). У предков крокодилов и птиц первичная горизонтальная перегородка в желудочке сердца редуцировалась и вместо нее возникла новая - вертикальная перегородка, полностью разделившая желудочек на левую (артериальную) и правую венозную) камеры. После редукции у предков птиц левой дуги начинавшейся от правой камеры желудочка, полностью прекратилось смешивание артериальной и венозной крови, что стало одной из важных предпосылок для развития у птиц гомойотермии. У предков млекопитающих в желудочке сердца сразу возникла вертикальная перегородка, которая, пока она была неполной, вероятно, не давала этим животным каких-либо преимуществ в разделении артериальной и венозной крови по сравнению с "типичными" пресмыкающимися - но этот путь эволюционных преобразований сердца позволил в дальнейшем разделить желудочек полностью без сложных дополнительных перестроек. Можно полагать, что предки завропсидных рептилий пошли по "инадаптивному" пути в разделении желудочка сердца по сравнению с более удачным ("адаптивным") решением этой проблемы у терапсидных рептилий - предков млекопитающих.
Инадаптивные изменения неизбежно должны быть обычным явлением в филогенезе различных групп организмов, ибо единственный фактор эволюции, обусловливающий развитие приспособлений, - естественный отбор - действует, так сказать, "в настоящем времени" и не может предвидеть эволюционных последствий преобразований, благоприятных в данный исторический момент и при данных условиях, но могущих при их дальнейшем развитии привести к эволюционному тупику и вымиранию.
Гиперадаптивность
На фоне обшей относительности совершенства приспособлений, наглядно проявляющейся в явлениях инадаптивной эволюции, особенно контрастно выглядит так называемая гиперадаптивность или конструктивная избыточность многих биологических систем, представляющая собой более или менее значительное (в некоторых случаях многократное) превышение их адаптивных возможностей по отношению к обычным условиям их функционирования. Это относится, например, к прочности костей на разрыв и сжатие (у млекопитающих она близка к прочности стали), к силе сокращения мышц (для одной мышцы лягушки она может примерно в 5 раз превышать вес всего тела) и т.п. Поразительную гиперадаптацию к действию низких температур проявляют в состоянии анабиоза споры ряда бактерий, а также некоторые микроскопические многоклеточные животные - коловратки (Rotatoria - класс типа нитчатых червей) и тихоходки (Tardigrada -- группа, близкая к кольчатым червям и членистоногим), которые могут выдержать температуру жидкого гелия (-270°С), близкую к абсолютному нулю.
Проявлением гиперадаптивности является также наличие в составе некоторых органов такого количества структурных элементов, которое во много раз превышает необходимое для их нормального функционирования при обычных условиях. Например, в женском яичнике закладывается от 40 до 200 тыс. яйцевых фолликулов, из которых овулирует в течение всей жизни женщины не более 500.
Важность и необходимость гиперадаптивности для выживания вида в изменчивой среде очевидна. В истории любого вида организмов неизбежно встречаются периоды необычных (экстремальных) условий, ставящих под угрозу существование целых популяций. Проявления гиперадаптивности представляют собой "запас прочности" систем организма, позволяющий им выдерживать повышенные функциональные нагрузки в таких экстремальных условиях. Конструктивная избыточность многократно повышает надежность биологических систем и является необходимым условием для выживания организмов, когда давление отбора достигает предельных значений.
Однако механизм возникновения гиперадаптивности является одной из сложных проблем адаптациогенеза. Естественный отбор, действуя лишь "в настоящем времени", может благоприятствовать только таким изменениям организмов, которые обеспечивают их носителям при данных условиях какие-либо преимущества в выживании или оставлении потомства. В то же время подавляющее большинство особей данного вида в течение всей жизни не подвергается воздействию экстремальных условий, при которых обладатели гиперадаптивного состояния получили бы соответствующие преимущества. Следовательно, гиперадаптивные признаки сами по себе имеют чрезвычайно мало шансов быть "замеченными" естественным отбором, и еще менее вероятны накопление и интеграция соответствующих изменений в ряду последовательных поколений. И совсем уже невероятным выглядит прямое закрепление отбором таких гиперадаптивных особенностей, которые обеспечивают сверхустойчивость организма (вплоть до условий, вообще не наблюдающихся ныне в биосфере Земли, например температур, близких к абсолютному нулю).
Чтобы разобраться в этой проблеме, прежде всего отметим, что понятие гиперадаптивности объединяет широкий круг очень разнородных явлений (в этом легко убедиться на приведенных выше примерах). Вероятно, возникновение разных форм конструктивной избыточности определяется различными причинами.
Вообще для нормального функционирования любого органа При самых обычных (средних) условиях полезно, чтобы диапазон его функциональных возможностей был существенно шире обычно используемого. Это позволяет избежать сбоев, вероятных в крайних областях функционального диапазона: краевые зоны неустойчивого функционирования отодвигаются за пределы обычно используемой "рабочей части" этого диапазона. Другими словами, некоторая потенциальная гиперадаптивность полезна уже для нормального функционирования органа в обычных условиях. Поэтому естественный отбор в процессе совершенствования любой адаптации благоприятствует расширению функциональных возможностей органа в гиперадаптивную область.
Дальнейшему усилению гиперадаптивности способствует действие катастрофического отбора (см. с. 108) в экстремальных условиях, раньше или позже неизбежно возникающих в истории любой филетической линии. При действии катастрофического отбора вид может выжить только в том случае, если в его популяциях имеются особи, обладающие соответствующей конструктивной избыточностью по сравнению с "нормой". При крайне неблагоприятных условиях катастрофический отбор элиминирует практически всю прежнюю популяцию, за исключением особей-носителей гиперадаптивного состояния отбираемых признаков. После возврата условий к прежней норме выжившие особи формируют новую популяцию, потомки которой будут сохранять конструктивную избыточность просто как своего рода нейтральные признаки (поскольку в нормальных условиях высокая степень гиперадаптивности отбором не контролируется) до следующей экстремальной ситуации. Очевидно, в длительной истории филетической линии важность гиперадаптаций для выживания вида не вызывает сомнений. Конструктивная избыточность, обеспечивающая запас прочности систем организма и дающая виду шансы на выживание в экстремальных условиях, является результатом действия катастрофического отбора в прошлой истории вида.
Кроме того, как показал П.В.Пучков, многие проявления конструктивной избыточности возникают как побочный результат контролируемого отбором развития обычных адаптации организмов к специфическим условиям их обитания. Так, гиперадаптивная устойчивость яиц жаброногих раков к действию низких температур (до -180°С), вероятно, возникла как побочный результат развития приспособлений, позволяющих этим организмам выживать при длительном пересыхании временных водоемов (благодаря особому химизму яиц и структуре их оболочек яйца жаброногих раков выдерживают высыхание до 9 лет), а также проходить без повреждений сквозь кишечный тракт проглотившего яйцо хищника.
Этот путь развития гиперадаптивных особенностей, очевидно, соответствует рассмотренному выше механизму морфофункиио-нальной преадаптации, обеспечивающему возможность расширения функций органов и приобретения ими новой адаптивной роли. Механизм преадаптации позволяет объяснить наиболее значительные проявления конструктивной избыточности организмов.
Анализ развития приспособлений организмов подводит нас к новому кругу проблем, относящихся уже к уровню макроэволюции. Собственно говоря, такие аспекты адаптациогенеза, как морфофункциональные преадаптации, инадаптивная эволюция, гиперадаптивность, мы по необходимости уже рассматривали в широкой исторической перспективе, т.е. именно в масштабе макроэволюции. С этими вопросами тесно связаны специфические проблемы макроэволюции: механизмы типогенеза (формирование основного плана строения, или типа организации высших таксонов) и типостаза (последующая длительная устойчивость этого общего плана организации), направленность эволюционного процесса и др. Чтобы разобраться в этих сложных проблемах, целесообразно предварительно рассмотреть реалии макрофилогенеза: его исторические масштабы, процессы формирования и историческую судьбу основных групп организмов, причины их расцвета и вымирания, соотношения этих процессов с изменениями среды обитания в геологической истории. В сущности, эволюция организмов представляет собой форму существования живой материи в изменяющейся с течением времени внешней среде, и все современные проявления жизни на любом уровне организации живой материи могут быть поняты лишь с учетом их эволюционной истории. Эти проблемы и рассматриваются в данной части книги.
Однако прежде чем мы приступим к рассмотрению последовательных этапов эволюции органического мира, нам необходимо отчетливо представлять временные масштабы этих процессов, т. е. масштабы геологического времени. Длительное и кропотливое изучение абсолютного и относительного возраста горных пород в разных регионах земного шара, потребовавшее работы нескольких Поколений геологов и палеонтологов, позволило наметить основные вехи геологической истории. История Земли разделена на крупные временные промежутки, называемые геологическими эрами; эры (за исключением наиболее древних) разделены на геологические периоды, а те, в свою очередь, - на эпохи (табл. 2). Границы Между этими подразделениями соответствуют разного рода изменениям геологического и биологического (палеонтологического) Характера: усиление вулканизма и горообразовательные процессы; поднятия или опускания значительных участков континентальной коры, приводившие к соответствующим вторжениям или отступаниям моря (морские трансгрессии и регрессии); существенные изменения фауны и флоры и т.п. Поскольку подобные события происходили в истории Земли нерегулярно, продолжительность различных эпох, периодов и эр различна.
Отложения древнейших археозойской и протерозойской эр содержат чрезвычайно мало ископаемых остатков организмов; по этому признаку археозой и протерозой нередко объединяют под названием "криптозой" (этап скрытой жизни), противопоставляя трем последующим эрам - палеозою, мезозою и кайнозою, объединяемым как "фанерозой" (этап явной, наблюдаемой жизни).
Таблица 2. Основные подразделения геологической истории Земли, их абсолютный возраст и продолжительность (в млн. лет):
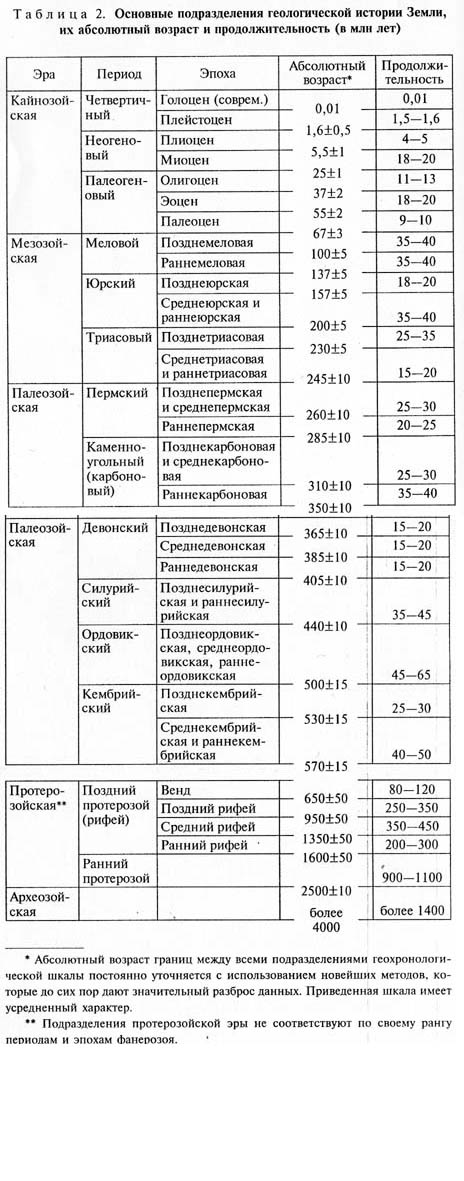
В последние десятилетия в позднепротерозойских (вендских) отложениях были сделаны довольно многочисленные находки ископаемых остатков разнообразных многоклеточных животных ("Эдиакарская фауна", см. с. 157). Поэтому некоторые ученые считают целесообразным присоединить венд к фанерозою.
Возраст Земли определяется различными учеными по-разному; можно указать приближенную цифру 5 млрд лет. Древнейшие известные на Земле горные породы - граниты Кольского полуострова - имеют возраст (установленный радиоактивным методом) не менее 4 млрд лет.
Развитие жизни в первой половине криптозоя
Предполагаемые древнейшие остатки организмов были описаны из археозойских пород с возрастом 3,1-3,8 млрд лет из различных районов земного шара (Гренландия, Южная Африка, Западная Австралия). Из этих находок лучше известны ископаемые остатки, обнаруженные в кремнистых сланцах системы Свазиленд (Восточный Трансвааль, Южная Африка). В этих осадочных породах встречаются различные микроскопические структуры, которые рассматриваются как ископаемые остатки одноклеточных сине-зеленых водорослей (Archaeospheroides barbertonensis) и бактерий (Eobacterium isolatum). Первые имеют сфероидную форму диаметром 17-20 мкм, вторые - палочковидную длиной 0,5-0,7 мкм и толщиной 0,2-0,3 мкм. Отмечены также нитчатые формы, напоминающие нитчатые синезеленые водоросли. Геохимическими методами было показано наличие в осадочных породах, вмещающих эти ископаемые остатки, ряда органических веществ, которые, судя по содержанию в них разных изотопов углерода, могут иметь биогенное происхождение, некоторые из них могут быть производными хлорофилла. Это указывает на возможность существования фотосинтезирующих организмов уже 3,2 млрд лет назад.
Необходимо оговориться, что принадлежность археозойских ископаемых остатков к древнейшим организмам не может пока считаться вполне доказанной. Некоторые ученые полагают, что это - псевдоископаемые, образовавшиеся абиогенным путем'. Структурно археозойские ископаемые остатки очень просты, поэтому пока нет оснований утверждать, что они не могли возник-луть абиогенно.
Однако в осадочных породах Западной Австралии, имеющих возраст 3,4-3,5 млрд лет, вместе с остатками микроорганизмов, подобных таковым из южноафриканских местонахождений, были обнаружены древнейшие строматолиты - следы жизнедеятельности водорослей (вероятно, синезеленых). Строматолиты найдены также в несколько более молодых отложениях Южной Африки в Зимбабве возрастом 2,9-3,2 млрд лет. Все эти находки убедительно свидетельствуют, что около 3,3 млрд лет назад на Земле уже существовали водоросли. Очевидно, появление жизни на Земле должно было произойти раньше - может быть, 4-4,25 млрд лет назад.
Протоорганизмы были, вероятно, гетеротрофными формами, которые питались готовыми высокомолекулярными органическими соединениями, содержавшимися в первобытном океане ("первичном бульоне") и имевшими абиогенное происхождение. В археозое жизнь существовала в условиях бескислородной восстановительной атмосферы, вероятно, в водоемах на глубине порядка 10-50 м. Десятиметровый слой воды защищал протоорга-низмы от губительного действия жесткого ультрафиолетового излучения Солнца.
В промежутке времени между 3,3 и 3,5 млрд лет какие-то формы древнейших организмов - прокариот, еще не имевших ядра, приобрели способность к фотосинтезу. В связи с этим произошла дивергенция прокариот на два главных ствола их эволюции:
1) бактерии (Bacteriae), в большинстве сохранившие в той или иной форме гетеротрофный способ питания и совершенствовавшие его применительно к новым условиям существования, и
2) синезеленые водоросли (Cyanophyta), развившие автотрофное питание посредством фотосинтеза. Бактерии от использования органических веществ абиогенного происхождения по мере уменьшения их содержания в окружающей среде и распространения фотосинтезирующих организмов все в большей степени переходили к питанию органическими веществами, синтезированными водорослями.
В течение последующих 1,5-2 млрд лет - на протяжении большей части криптозоя - происходила эволюция прокариот, в ходе которой возникли новые варианты их строения, метаболизма, питания, но основные их черты, характеризующие прокариотический уровень организации, оставались неизменными. Вероятно, около 3,3 млрд лет назад появились нитчатые формы синезс-леных водорослей. Нитчатый тип строения представляет собой цепочку клеток, возникшую путем их последовательного деления в одном направлении. Клетки в цепочке связаны друг с другом лишь механически -- разрыв цепочки никак не сказывается на жизнедеятельности и жизнеспособности составляющих ее клеток. Поэтому нитчатые формы нельзя приравнивать к многоклеточному состоянию, характеризующемуся дифференциацией и интеграцией целостного организма, различные части которого выполняют разные функции и подчинены целому.
В отложениях первой половины протерозоя (возрастом 2,5- 1,7 млрд лет) ископаемых остатков организмов несколько больше, а главное - их природа не вызывает таких сомнений, как для более древних микроископаемых. Наиболее известны ископаемые остатки флоры, обнаруженные в Канаде в кремнистых сланцах формации Ганфлинт на северном берегу озера Верхнего. Возраст этих отложений составляет около 1,9 млрд лет. В строматолитовых кремнистых породах здесь встречаются ископаемые остатки довольно разнообразных микроорганизмов (рис. 25). Обычны нитчатые формы длиной до нескольких сот микрометров (микронов) и толщиной 0,6-16 мкм различного строения: простые и ветвистые, нечленистые и разделенные по длине на сегменты (клетки?), причем последние могут быть разной формы - округлые и удлиненные. Строение некоторых нитевидных организмов (Gunflintia), по мнению Е. Баргхоорна, напоминает таковое современных синезеленых водорослей, другие же сходны с железобактериями. Имеются и одноклеточные микроорганизмы диаметром 1-16 мкм, также различного строения: от простых округлых, снабженных толстой оболочкой Huroniospora, до сложных причудливой формы Eoastrion и Kakabekia. В этих породах обнаружены и молекулярные ископаемые - органические вещества, подобные найденным в более древней формации Свазиленд, но более разнообразные.


В осадочных породах, образовавшихся в промежутке времени между 2 и 1 млрд лет назад, часто встречаются строматолиты (рис. 26), что говорит о широком распространении и активной фотосинтезирующей деятельности синезеленых водорослей в этот период.
Жизнь в среднем протерозое и происхождение эукариот
Следующий важнейший рубеж в эволюции жизни документируется рядом находок ископаемых остатков в отложениях, имеющих возраст 0,9-1,3 млрд лет. Среди них наиболее известна ископаемая флора, обнаруженная в углистых кремнистых сланцах формации Биттер-Спрингс (возраст 0,9 млрд лет) в Центральной Австралии. Помимо одноклеточных и нитчатых форм синезеленых водорослей (по мнению Д. Шопфа, они уже относятся к современным семействам этих растений) здесь найдены прекрасной сохранности остатки одноклеточных организмов размером 8-12 мкм, в которых удалось различить внутриклеточную структуру, похожую на ядро (рис. 27). Обнаружены также стадии деления одного из видов этих одноклеточных организмов (Glenobotrydion aenigmati-cus), напоминающие стадии митоза - способа деления эукарио-тических (т.е. имеющих ядро) клеток. Сходные ископаемые остатки содержащих ядро клеток и стадий митоза были найдены также в Калифорнии в отложениях, имеющих возраст 1,3 млрд лет, а также на реках Мироедиха (Туруханский район) и Мая (Юго-Восточная Якутия) в отложениях возрастом 0,9-1 млрд лет.

Если интерпретация всех этих ископаемых остатков правильна1, это означает, что около 1,6-1,3 млрд лет назад эволюция организмов прошла важнейший рубеж - был достигнут уровень организации эукариот.
Ранний протерозой с полным основанием называют "веком синезеленых водорослей", получивших самое широкое распространение; это было время их расцвета. Благодаря их фотосинтетической деятельности постепенно повышалось содержание кислорода в атмосфере. Первоначально свободный кислород быстро использовался как окислитель в различных химических реакциях, в частности в процессах отложения осадочных железных руд при участии ферробактерий (основная масса железорудных толщ на всей Земле сформировалась в промежутке 2,2-1,9 млрд лет назад). После завершения этих процессов, радикально изменивших геохимический облик поверхностных слоев земной коры, кислород стал накапливаться в атмосфере во все более заметных количествах. Большие масштабы в это время приобрела и строматолитообразующая деятельность нитчатых синезеленых водорослей, которая привела к образованию огромных толщ карбонатных пород.
Возникновение эукариот - это следующий важнейший рубеж в до-кембрийской эволюции организмов после возникновения фотосинтеза. Эукариоты обладают значительно более сложной и совершенной организацией клетки, чем прокариоты. Протоплазма эукариотической клетки сложно дифференцирована: в ней обособлены ядро и другие различные органоиды (митохондрии, пластиды, комплекс Гольджи, центриоли и др.), отделенные от цитоплазмы полупроницаемыми мембранами и выполняющие различные функции, снаружи клетка окружена клеточной мембраной. Наружная и внутренние клеточные мембраны образуют единый мембранный комплекс. Мембраны имеют сложное молекулярное строение (липидная "пленка", пронизанная белковыми "каналами"), обусловливающее избирательное проникновение веществ. У прокариот вместо клеточной мембраны клетка обволакивается единственной гигантской молекулой мукопептида (вещество, состоящее из аминокислот, углеводов и липидов). В ядре - интегрирующем центре клетки эукариот - локализован хромосомный аппарат, в котором сосредоточена основная часть
кода наследственной информации. У прокариот нет такого интегрирующего центра и подобной упорядоченности наследственной информации; элементы аппарата наследственности отчасти включены в единственную так называемую "хромосому" бактериальной клетки, отчасти находятся в цитоплазме. Упорядоченность аппарата наследственности и сложность организации клетки у эукариот требуют такой же упорядоченности в передаче наследственной информации при клеточном делении. В связи с этим у эукариот развился весьма совершенный механизм клеточного деления - митоз, обеспечивающий точное и эквивалентное распределение наследственной информации в обе дочерние клетки. У прокариот митоза не наблюдается.
Одним из важнейших эволюционных достижений эукариот является возникновение механизма цитоплазматической подвижности, связанное с формированием аппарата микротрубочек (и его производных), в котором локализованы сократительные белки: миозин, актин, тубулин (у прокариот актин и тубулин встречаются редко, а миозин неизвестен). Источником энергии для функционирования сократительных белков является аденозинтрифосфат (АТФ) - основной аккумулятор и переносчик химической энергии в клетке. Возникновение митоза стало возможным именно благодаря развитию этого аппарата, с ним же связано и совершенствование механизмов активной подвижности организма во внешней среде. (Среди прокариот подвижностью обладают некоторые жгутиконосныс бактерии. Однако жгутики бактерий не способны изгибаться. Они образованы белком флагеллином, не обладающим АТФ-азной активностью. Источником энергии для движения бактерий является градиент ионов водорода на бактериальной мембране).
Эукариотный уровень организации обеспечивает более эффективное выполнение всех клеточных функций и открывает перспективы для дальнейшей прогрессивной эволюции.
Имеются две основные точки зрения на проблему происхождения эукариотической клетки и соответственно эукариот как одной из высших таксономических групп. Первая из них - так называемая аутогенная концепция - предполагает постепенную дифференциацию прокариотической клетки, в ходе которой развился мембранный комплекс. Сначала могла возникнуть наружная клеточная мембрана, затем ее локальные впячивания внутрь клетки образовали внутриклеточные мембраны. На основе дифференциации мембранного комплекса структурно оформились клеточные органоиды. Согласно аутогенной концепции эукариоты возникли от одной из групп прокариот (от какой именно - указать сейчас невозможно).
Вторая точка зрения (симбиогенная концепция) получила широкую известность после работ Л.Маргулис (Саган), в 1967-1971 гг. обосновавшей гипотезу о возникновении эукариотической клетки из симбиоза разных прокариот: крупной клетки - хозяина - и более мелких организмов, поселившихся в цитоплазме первого. Последние дали начало различным органоидам. Аргументы в пользу этой точки зрения сводятся к наличию в некоторых органоидах эукариотической клетки (митохондрии, пластиды) собственной ДНК, некоторому сходству нуклеотидных последовательностей этих органоидов с таковыми ряда прокариот, а также су-шествованию сложных симбиотических комплексов среди современных организмов (известно, например, около тысячи видов-эндосимбионтов, обитающих в организме одноклеточных простейших животных). Согласно симбиогенной концепции предками эукариот должны быть сразу несколько групп прокариот.
Симбиогенная концепция получила широкое распространение в течение двух последних десятилетий. Однако все же аутогенная (классическая) точка зрения представляется в настоящий момент лучше обоснованной и согласующейся с общими тенденциями прогрессивной эволюции организмов (см. ч. IV). ДНК органоидов могла возникнуть путем сегрегации (обособления) фрагментов первоначально единого генома.
Для эукариот характерен половой процесс, сущностью которого является обмен наследственной информацией между организмами, принадлежащими к одному биологическому виду. Как мы видели выше, половой процесс объединяет наследственную информацию, присущую разным индивидам, в единый видовой генофонд и, с другой стороны, обеспечивает перекомбинацию наследственной информации в каждом следующем поколении. Половой процесс известен и у некоторых прокариот (бактерии), но у них он неупорядочен и не обеспечивает эквивалентного обмена разных особей наследственной информацией.
Вероятно, практически одновременно с развитием у эукариот полового процесса у них должен был сформироваться и мейоз, необходимый для редукции числа геномов, удваивающегося при половом процессе. Мейоз также способствует возрастанию комбинативной изменчивости вида. Результатом ее резкого усиления у эукариот было значительное убыстрение их эволюции. Вместе с тем организация клеточного ядра и аппарата клеточного деления 1у эукариот обеспечивает гораздо более совершенную защиту генома от горизонтального переноса генов, чем это характерно для прокариот. Горизонтальный перенос генов для эукариот теряет существенное значение как фактор эволюции.
Уже около 0,9 млрд лет назад произошло разделение эволюционного ствола древнейших эукариот на ряд ветвей, давших начало разным типам водорослей: зеленым (Chlorophyta), бурым (Phaeophyta), красным (Rhodophyta) и др., а также грибам (Fungi). Д.Шопф полагает, что описанные выше ископаемые остатки различных эукариот из местонахождений возраста 0,9-1,3 млрд лет принадлежат ко всем этим группам низших растений.
Вероятно, в это время существовали уже и одноклеточные животные -- простейшие (Protozoa), либо возникшие от общего Предкового ствола всех эукариот, либо обособившиеся от ранних Представителей одной из групп растений. От какой именно группы - с уверенностью сказать сейчас невозможно: разные исследователи связывали происхождение животных с каждой из названных групп растений. По общей совокупности данных, к предкам Protozoa наиболее близкими кажутся некоторые одноклеточные зеленые водоросли. Не случайно таких жгутиконосных одноклеточных, как эвгленовые, вольвоксовые и др., способных и к фотосинтезу, и к гетеротрофному питанию, ботаники традиционно рассматривают в составе типа зеленых водорослей, а зоологи - в составе типа простейших животных.
Обращает на себя внимание отсутствие ископаемых остатков животных в отложениях большей части криптозоя. Остатки некоторых одноклеточных простейших (радиолярии и, возможно, фо-раминиферы) и ряда групп многоклеточных животных появляются лишь в вендских отложениях. Это может быть связано и с более поздним возникновением животных, и с худшими возможностями фоссилизации (т.е. перехода в ископаемое состояние) их клеток, лишенных в отличие от растительных прочной целлюлозной оболочки. Скелета же протерозойские животные не имели.
Происхождение многоклеточных организмов
Вероятно, 700-900 млн лет назад на Земле появились первые многоклеточные животные и растения. У растений возникновение многоклеточного уровня организации, вероятно, произошло на основе дифференциации лентообразных колоний, образовавшихся путем бокового срастания прикрепленных нитчатых форм или благодаря делению клеток последних в двух взаимно перпендикулярных направлениях (в одной плоскости). У колоний, прикрепленных одним концом к субстрату, различные участки находились в разных условиях по отношению к падающему свету, субстрату и водной среде. В связи с этим естественный отбор благоприятствовал возникновению некоторой дифференциации частей колонии. Первым шагом было возникновение полярности колонии; на одном ее конце выделялись клетки, служившие для прикрепления к субстрату (для них характерно ослабление фотосинтеза, потеря способности к делению), на другом же конце - верхушечные клетки, интенсивно делившиеся и образовавшие своего рода "точку роста" колонии. Естественный отбор благоприятствовал приобретению клетками колонии способности делиться в разных направлениях; это приводило к ветвлению, что увеличивало поверхность колонии. Деление клеток вдоль трех взаимно перпендикулярных осей или переплетение отдельных нитей вело к возникновению многослойного "объемного" тела. В процессе его дальнейшей дифференциации сформировались многоклеточные органы, выполнявшие разные функции (фиксация на субстрате, фотосинтез, размножение). Одновременно между разными клетками растения складывалась определенная взаимозависимость, что, собственно говоря, и знаменует достижение многоклеточного уровня организации.
У животных активный образ жизни требовал более совершенной и сложной дифференциации организма, чем у растений. Сложность организации многоклеточных животных (Metazoa) и разнообразие ее конкретных форм стимулировали разработку различных гипотез о происхождении Metazoa.
Первая из них берет начало в работах Э.Геккеля, который в разработке своей теории гастреи основывался на сформулированном им биогенетическом законе, согласно которому онтогенез данного вида организмов представляет собой сжатое и сокращенное повторение (рекапитуляцию) хода филогенеза его предков (подробнее см. в ч. IV). В соответствии с этим Э.Геккель полагал, что филогенез древнейших Metazoa в определенной степени повторяется в онтогенезе современных низших многоклеточных животных (рис. 28). Согласно Геккелю, предками Metazoa были колониальные простейшие, обладавшие сферическими колониями с однослойной стенкой, подобными бластуле - одной из ранних стадий эмбрионального развития современных многоклеточных животных. Геккель назвал эту гипотетическую предковую форму "бластеей". При направленном плавании сферическая колония - бластея - ориентировалась одним полюсом вперед, как это наблюдается и у современных колониальных простейших, например у Volvox. Согласно Геккелю, на переднем полюсе колонии возникло впячивание ее стенки внутрь, подобно тому, как это происходит при инвагинационной гаструляции в онтогенезе некоторых современных Metazoa. В результате образовался многоклеточный организм - "гастрея", стенка тела которого состоит из двух слоев, экто- и энтодермы. Энтодерма окружает внутреннюю полость - первичный кишечник, открытый наружу единственным отверстием - первичным ртом. Организация гастреи соответствует принципиальному плану строения кишечнополостных (тип Coelenterata), которых Геккель и рассматривал как наиболее примитивных многоклеточных животных.


И.И.Мечников обратил внимание на то, что у примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша - бластулы), что характерно для более высокоорганизованных групп, а посредством миграции некоторых клеток из однослойной стенки тела внутрь (рис. 29). Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. Такой способ гаструляции гораздо проще, чем инвагинация, так как не требует сложного направленного и координированного смещения целого пласта клеток, и, вероятно, примитивнее инвагинации. В связи с этим Мечников модифицировал гипотезу Геккеля следующим образом. В сфероидной колонии простейших - жгутиконосцев клетки ее однослойной стенки, захватывавшие (фагоцитировавшие) пищу, мигрировали для ее переваривания внутрь, в полость колонии (подобно миграции клеток будущей энтодермы в Процессе гаструляции кишечнополостных). Эти клетки образовали рыхлое внутреннее скопление -- фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток - кинобласт - осуществлял функции защиты и движения организма. Для захвата новых пищевых частиц клеткам фагоци-тобласта, по мысли Мечникова, не было необходимости возвращаться в поверхностный слой: располагаясь непосредственно под кинобластом, клетки фагоцитобласта захватывали пищевые частицы псевдоподиями, выдвигаемыми наружу в промежутках между клетками фагоцитобласта. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. В дальнейшем как приспособление к повышению активности питания у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. По мнению некоторых ученых, это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц. Гипотеза Мечникова, как и гипотеза Геккеля, рассматривает в качестве наиболее примитивных многоклеточных животных кишечнополостных и губок.
Важные сведения для понимания ранних этапов эволюции Metazoa были получены при изучении крайне примитивного многоклеточного животного трихоплакса (Trichoplax adhaerens), обнаруженного в Красном море Ф. Шульце еще в 1883 г., но детально исследованного лишь в 1970-е гг. нашего века К. Греллом и А. В. Ивановым. Трихоплакс (рис. 30) имеет уплощенное тело, лишенное полярности. Поверхность тела, обращенная вверх, покрыта плоским, а нижняя - цилиндрическим мерцательным эпителием. Внутри, между эпителиальными слоями, соответствующими кинобласту, находится полость с жидким содержимым, в Котором располагаются веретеновидные и звездчатые клетки. Эти Последние можно рассматривать как фагоцитобласт. Размножайся трихоплакс бесполым способом - делением и почкованием. А.В. Иванов указал, что трихоплакс представляет собой как бы Живую модель фагоцителлы, и предложил выделить эту форму в °собый тип животных Phagocytellozoa. По-видимому, трихоплакс Подкрепляет позиции гипотезы фагоцителлы И.И.Мечникова. Однако по современным представлениям непосредственными потомками фагоцителлозой среди многоклеточных были не кишечнополостные, а примитивные червеобразные животные, близкие по уровню организации к плоским ресничным червям - турбелляриям.

Первые ископаемые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650-570 млн лет назад) существовали уже разнообразные животные, вероятно принадлежавшие к различным типам. Немногочисленные отпечатки мягкотелых вендских животных известны из разных районов всех континентах земного шара, кроме еще малоисследованной Антарктиды. Ряд интересных находок был сделан в позднепротерозойских отложениях на территории России -- на Кольском полуострове, Архангельской области, на реке Мая и на Оленекском поднятии в Якутии и т.д.
Наиболее известна богатая позднепротерозойская фауна, обнаруженная в Центральной Австралии в районе Эдиакары к северу от г.Аделаида. Исследовавший эту фауну М.Глесснер считает, что она включает несколько десятков видов очень разнообразных многоклеточных животных, относящихся к разным типам (рис. 31). Большинство форм принадлежат, вероятно, к кишечнополостным. Это медузоподобные организмы, вероятно "парившие" в толще воды (Ediacara flindersi, Beltanella gilesi, Medusinites asteroides и др.), и прикрепленные к морскому дну полипоидные формы, одиночные или колониальные, напоминающие современных кораллов альционарий, или морские перья (Rangea longa, Arborea arboкa, Pteridinium simplex и др.). Замечательно, что все они, как и Другие животные эдиакарской фауны, лишены твердого скелета.
Кроме кишечнополостных в составе эдиакарской фауны найдены остатки червеобразных животных, причисляемых к плоским и кольчатым червям (Spriggina flounderi и разные виды Dickinsonia). Некоторые виды организмов интерпретируют как возможных пред-Ков членистоногих (Praecambridium sigillum, напоминающий по Характеру сегментации тела трилобитов и хелицеровых) и иглокожих (Tribrachidium heraldicum с телом дисковидной формы, на плоской поверхности которого выступают три валика, и Arkaria adami с пятилучевой звездообразной впадиной на ротовой стороне тела и с подобием амбулакральных желобков). Наконец, имеется целый ряд ископаемых организмов неизвестной таксономической принадлежности.
Многие вендские организмы были обнаружены также в вендских отложениях разных районов России: медузоподобные эдиакария и медузинитес - на полуострове Рыбачьем, птеридиниум - на севере Якутии, напоминающая сприггину вендия - в районе Яренска Архангельской области и т.д. Местонахождения вендской фауны, по богатству не уступающие эдиакарскому, были найдены на реке Сюзьма на Онежском полуострове и на Зимнем берегу Белого моря. Здесь были обнаружены ископаемые остатки свыше 30 видов бесскелетных многоклеточных животных, размеры которых варьировали от 3 мм до 30 см. Среди них вероятные представители кишечнополостных, плоских и кольчатых червей, членистоногих, иглокожих, а также ряд форм, принадлежащих к каким-то неизвестным группам. Вообще родство вендских организмов с современными группами, большинство из которых достоверно известно начиная с кембрия, остается проблематичным - различия очень велики, и некоторые исследователи считают, что известные ныне вендские организмы не связаны прямым родством с более поздними кембрийскими, а представляют слепые эволюционные ветви.
М.А.Федонкин, исследовавший Беломорскую фауну вендских животных, полагает, что некоторые из этих организмов обладают признаками нескольких разных типов животных и могут представлять исходные формы, занимающие промежуточное положение. Федонкин обратил также внимание на сходство ряда вендских организмов с личиночными стадиями некоторых современных животных, хотя вендские организмы имеют значительно более крупные размеры, чем соответствующие личинки. При всем разнообразии планов строения тела у организмов вендской фауны ("вендобионтов") их объединяют некоторые общие черты организации: отсутствие скелета, конечностей, вероятно, также дыхательных и пищеварительных органов. Многие вендобионты вели неподвижный прикрепленный образ жизни. Некоторые исследователи полагают, что вендские организмы питались осмотически через поверхность тела или же с помощью живших в их теле фото- или хемосинтезируюших симбионтов - одноклеточных водорослей и бактерий.
Хотя среди вендских животных преобладают мягкотелые бесскелетные формы, вероятно, в те времена существовали уже и немногие виды, обладавшие раковиной. Такова, например Cloudina, имевшая простую трубчатую раковину, состоявшую из органического вещества и кальцита. Клаудина была обнаружена в карбонатных породах, которые переслаиваются с отложениями, содержащими остатки эдиакарской фауны мягкотелых животных.
Все эти данные указывают на широкое распространение фаун мягкотелых животных в вендское время. Накопление материалов по вендским ископаемым организмам позволило некоторым исследователям поставить вопрос о расширении рамок фанерозоя, с включением в его состав "эдиакария" - периода, охватывающего промежуток времени от 670 до 550 млн лет назад (в приведенной на с. 143 схеме геохронологии этот период соответствует венду в составе протерозоя).
Поскольку вендская фауна столь разнообразна и включает довольно высокоорганизованных животных, очевидно, что до ее возникновения эволюция Metazoa продолжалась уже очень долго. Вероятно, многоклеточные животные появились значительно раньше - где-то в промежутке 700-900 млн лет назад. (Недавно появились первые данные об ископаемых остатках многоклеточных животных, найденных в Канаде и в Китае в породах, имеющих такой геологический возраст и отделенных от пород с остатками здиакарских организмов слоями ледниковых отложений).
Таким образом, в позднем протерозое (600-650 млн лет назад) уже существовали такие группы многоклеточных животных, как губки, кишечнополостные, плоские и кольчатые черви и даже, возможно, предки членистоногих. Судя по достигнутому уровню организации, можно предполагать, что к этому времени обособились также эволюционные стволы нитчатых червей (тип Nemathelminthes), предков моллюсков и предков вторичноротых животных - олигомерных червей.
Докембрийский филогенез Metazoa можно гипотетически представить следующим образом (рис. 32). От колониальных жгутиковых (по мнению ряда авторов - от гетеротрофных форм, принадлежавших к отряду Protomonadida) путем дифференциации и интеграции колонии, с миграцией внутрь колонии клеток фагоцитобласта на заднем полюсе тела, возникли первые многоклеточные животные, организация которых соответствовала фа-гоцителле (по И. И. Мечникову). Мало изменившимися потомками этих древнейших многоклеточных являются современные Phagocytellozoa (Trichoplax adhaerens). Примитивные многоклеточные были свободноплавающими (за счет работы мерцательного эпителия - кинобласта) животными, питавшимися различными микроорганизмами - простейшими и одноклеточными водорослями.

При дальнейшем развитии приспособлений к активному питанию происходила постепенная эпителизация фагоцитобласта, т.е. преобразование рыхлого скопления клеток в организованный клеточный пласт - эпителий кишечника. Эпителизация фагоиито-бласта, вероятно, началась с развития на заднем по движению полюсе тела постоянного ротового отверстия. Как отметил К. В. Беклемишев, на этой стадии филогенеза организм стал питаться как целое, а не как совокупность отдельных самостоятельно фагоцитирующих клеток. Вероятно, к этому времени сформировалась и интегрирующая организм нервная система в виде эпителиального нервного сплетения. Активное плавание требовало способности ориентироваться в пространстве и координировать работу всех органов. Для осуществления этих функций на аборальном (противоположном ротовому отверстию) полюсе тела возник нейро-рецепторный комплекс, включавший нервный ганглий, осязательные щетинки и статоцист (орган равновесия). Подобный аборальный орган имеется у современных гребневиков (тип Ctenophora), а также у свободноплавающих личинок очень многих групп животных: плоских и кольчатых червей, моллюсков, членистоногих, полухордовых, иглокожих и др. Эту гипотетическую стадию филогенеза древних Metazoa можно назвать "стомо-фагоцителлой" (подчеркивая эпителизацию лишь ротового отдела фагоцитобласта).
Возможно, на этой стадии филогенеза произошла первая крупная дивергенция филогенетического ствола древних многоклеточных, связанная с тем, что некоторые группы этих животных перешли к освоению морского дна, другие же продолжали совершенствовать приспособления к активной жизни в толще воды.
Современные низшие плоские черви - бескишечные турбеллярии (Acoela) в целом сохранили тот уровень организации, который, вероятно, был характерен для древнейших многоклеточных, впервые перешедших к освоению подвижного образа жизни на дне водоемов. От вендских представителей этих турбеллярий могли возникнуть филогенетические стволы, ведущие к другим группам плоских червей, к нитчатым червям и к предкам кольчатых червей (протоаннелидам). От протоаннелид обособились, с одной стороны, предки моллюсков, с другой - предки членистоногих. У всех этих групп произошла дальнейшая дифференциация фагоцитобласта. У низших червей эпителизировалась лишь его центральная часть, что привело у плоских червей к формированию разветвленного кишечника с единым отверстием - "ртом", ведущим во внешнюю среду, а у нитчатых червей - к образованию сквозного кишечника с ротовым и анальным отверстиями. ;У высших групп (кольчатые черви, моллюски и членистоногие) эпителизировался весь фагоцитобласт: не только его центральная часть (энтодермальный кишечник), но и периферическая (мезодерма и ее производные). Последнее привело к развитию вторичной полости тела - целома, стенки которой образованы мезодер-мальным целомическим эпителием. Более примитивные представители кольчатых червей, моллюсков и членистоногих обладают характерной личиночной стадией - трохофорой. В связи с этим указанные группы иногда объединяют под названием Trochozoa.
У тех потомков стомофагоцителлы, которые продолжали совершенствовать адаптации к жизни в толще воды, также произошла эпителизация центрального и отчасти периферического фагоцитобласта: возникла гастральная полость (первичный кишечник) и ее периферические ветви (гастроваскулярные каналы). К этому уровню организации среди современных животных ближе всего стоят гребневики, вероятно сохранившие примитивный образ жизни в толще воды. От их позднепротерозойских предков, которых можно Назвать "проктенофорами", с переходом к прикрепленной жизни На морском дне возникли стрекающие кишечнополостные (тип Coelenterata, или Cnidaria).
Другие филогенетические линии, ответвившиеся от проктенофор, также осваивали морское дно, но с развитием приспособлений к активному передвижению по субстрату, подобно турбелляриям и их потомкам, но на другом исходном уровне организации. У этих форм в результате завершения эпителизации периферического фагоцитобласта также образовалась вторичная полость тела - целом, но возникла она совершенно другим способом, чем у Trochozoa. В онтогенезе животных, происходящих от проктенофор, вторичная полость тела обособляется от первичного кишечника, как его боковые карманообразные выпячивания (первоначально было, вероятно, три пары таких выпячиваний), которые затем отшнуровываются от стенок кишки (рис. 33). Такой способ развития целома получил название энтероцельного - в отличие от схизоиельного способа, характерного для Trochozoa, у которых целом возникает в результате появления полостей внутри скоплений мезодермальных клеток, без всякой связи с первичным кишечником. О. и Р. Гертвиги и И.И.Мечников обосновали гипотезу, согласно которой энтероцельный целом возник в эволюции из гастроваскулярных каналов проктенофорных предков (энтероцельная теория происхождения целома). Энтероцельный целом характерен для типов погонофор (Pogonophora), шетинкочелюстных (Chaeto-gnatha), плеченогих (Brachiopoda), мшанок (Bryozoa) и ряда других, в том числе группы так называемых вторичноротых животных (Deuterostomia), объединяющей типы хордовых (Chordata), иглокожих (Echinodermata) и полухордовых (Hemichordata). У вторичноротых животных имеется много общего, в частности особое положение дефинитивного (присущего взрослым организмам) рта, возникающего на полюсе тела, противоположном первичному эмбриональному рту - бластопору. На месте же последнего развивается анальное отверстие. Вторичноротые, несомненно, имеют общее происхождение; в качестве их предков указывают гипотетическую группу олигомерных червей, тело которых было разделено на три отдела, имелся вторичный рот и энтероцельный целом. Среди современных вторичноротых к уровню организации олигомерных червей ближе всего стоят свободноживущие полухордовые, представителем которых является желудевый червь (Balanoglossus).
Особое положение среди многоклеточных животных занимают губки (тип Porifera, или Spongia). Эта группа характеризуется очень Примитивным общим уровнем организации: губки, по существу, не имеют эпителизованного фагоцитобласта, упорядоченного внутреннего строения, настоящего кишечника, нервной системы, рецепторов и т. д. Губки отличаются от всех остальных Metazoa чрезвычайно своеобразным онтогенезом, в ходе которого происходит инверсия зародышевых листков (экто- и энтодерма, так сказать, меняются местами). Новейшие данные молекулярных исследований показали, что губки имеют общее происхождение со всеми многоклеточными. Вероятно, они представляют собой очень раннюю боковую ветвь, обособившуюся на уровне фагопителлы. Древнейшие ископаемые остатки губок известны из вендских (эдиа-карских) отложений в Австралии.
"Взрывная эволюция" в начале кембрия
Граница между протерозойской и палеозойской эрами отмечается резким изменением в составе и богатстве ископаемой фауны. После в основном "немых" толщ верхнего протерозоя, среди которых рассеяны относительно редкие местонахождения эдиакар-ской фауны, в осадочных породах кембрия (первого периода палеозойской эры), начиная с самых нижних горизонтов, внезапно возникает огромное разнообразие и обилие остатков ископаемых организмов. Среди них помимо низших многоклеточных - губок и кишечнополостных - встречаются также представители высокоразвитых типов животных - плеченогих, моллюсков, членистоногих и др. К концу кембрия появляются почти все известные типы многоклеточных животных. Этот взрыв формообразования на границе протерозоя и палеозоя - одно из самых загадочных событий в истории жизни на Земле. Благодаря этому начало кембрийского периода является столь заметной вехой, что нередко все предшествующее время в геологической истории, т. е. весь криптозой, именуют "докембрием".
Находки эдиакарской фауны и другие данные по докембрийс-ким организмам говорят о том, что эволюция многоклеточных Животных началась задолго до рубежа нижнего кембрия и привела к формированию разнообразных групп. Есть основания считать, что в позднем протерозое уже сложились многие типы Metazoa. Крайняя редкость захоронений ископаемых представителей этих Фупп в отложениях позднего протерозоя, может быть, объясняйся отсутствием у большинства доксмбрийских животных твердого скелета. В раннем кембрии такой скелет появился у самых Разных групп животных. При этом скелетные ткани и анатомия скелета были совершенно различны у разных групп: от гибких членистых хитиновых панцирей членистоногих до монолитных известковых раковин моллюсков и плеченогих. Было высказано предположение, что причиной появления большого количества ископаемых остатков организмов в раннекембрийских отложениях было изменение гидрохимического режима водоемов, благоприятствующее скелетообразованию по чисто химическим причинам, в частности, указывают на усиленное отложение фосфоритов в морских осадках раннего кембрия. Но такое предположение не увязывается с указанным разнообразием форм и химического состава скелета у кембрийских животных. Как могло благоприятствовать повышение содержания фосфатов в воде образованию, например, кремниевой раковины, или хитина, представляющего собой комплекс азотсодержащих полисахаридов? Таким образом, едва ли правильно пытаться свести "нижнекембрийскую революцию" только к скелетизации организмов. Замечательная ископаемая фауна, включающая разнообразных животных, как наделенных скелетом, так и "мягкотелых", была обнаружена в начале 1980-х гг. в средне-кембрийских сланцах Бёрджес в Британской Колумбии в Канаде. В состав этой фауны входит около 120 родов, среди которых представители губок, кишечнополостных, кольчатых червей-полихет, моллюсков, членистоногих, иглокожих, полухордовых, хордовых, плеченогих и других групп, в том числе загадочных организмов неизвестной таксономической принадлежности. Среди них выделяются крупные хищники Anomalocaris, достигавшие 2 м длины и обладавшие странным обликом: удлиненное обтекаемое тело, большие стебельчатые глаза, пара членистых околоротовых придатков, служивших, вероятно, для захвата добычи; челюстной аппарат из множества подвижных пластинок, брюшной плавник, расчлененный на ряд последовательных лопастей. В 1994 г. остатки ископаемой фауны, близкой по составу к фауне Бёрджес, были обнаружены в нижнекембрийских отложениях Южного Китая.
Для объяснения резких изменений состава и общего облика фауны и флоры в некоторые моменты истории Земли, в том числе на рубеже протерозоя и палеозоя, некоторые ученые (в частности, О. Шиндевольф) пытаются привлечь "взрыв мутаций", вызванный какими-то катастрофическими изменениями внешних условий. Подавляющее большинство мутаций снижают жизнеспособность мутантных организмов - этим пытаются объяснить массовое вымирание видов прежней фауны и флоры. В то же время общее усиление мутационного процесса, по мнению Шиндевольфа, приводит к возникновению и быстрому распространению новых форм. При этом сразу возникают новые типы организации - скачкообразно, посредством крупных мутаций. Возможную причину такого "мутационного взрыва" О.Шиндевольф видел в повышении уровня жесткой космической радиации в результате вспышки Сверхновой звезды на достаточно близком расстоянии от Солнца.
К сожалению, эта концепция не может объяснить ни одного конкретного случая изменений фауны и флоры. Почему вымерли одни группы организмов и преуспели другие, прежде сосуществовавшие с первыми? В сущности, гипотеза "мутационного взрыва" просто подменяет анализ конкретных ситуаций постулированием универсальной причины для объяснения любого крупномасштабного эволюционного изменения. Как мы уже неоднократно подчеркивали, основой эволюционного процесса являются малые мутации, и филогенетические преобразования большого масштаба складываются из них под контролем естественного отбора. Крупные же мутации обычно детальны. Наконец, расчеты показали, что вспышки Сверхновых звезд не могут вызвать такого повышения радиации на поверхности Земли, которое имело бы предполагавшийся О.Шиндевольфом мутагенный эффект.
Вообще всегда предпочтительнее искать объяснения событиям, происходящим на Земле, не прибегая к таким гипотезам, которые постулируют универсальное влияние каких-то космических факторов, никак не опосредованное конкретными земными условиями.
В этом отношении от других концепций выгодно отличается гипотеза, выдвинутая в 60-е гг. Л.Беркнером и Л.Маршаллом, объясняющая многие важнейшие события в ходе докембрийской и послекембрийской эволюции, а также на рубеже криптозоя и фанерозоя, опираясь на закономерные изменения условий в среде обитания древних организмов. Эта гипотеза связывает развитие жизни на Земле с изменениями содержания кислорода в земной атмосфере.
В современной атмосфере Земли содержится около 21% кислорода (это соответствует его парциальному давлению 159 мм ртутного столба). Свободный кислород атмосферы необходим подавляющему большинству современных организмов как окислитель в процессах дыхания. Водные организмы обычно дышат кислородом, растворенным в воде, но этот последний образует с атмосферным кислородом единую систему: избыток кислорода, выделяющегося в воде в результате фотосинтеза водных растений, Поступает в атмосферу, а кислород атмосферы растворяется в поверхностном слое воды, по тем или другим причинам обедненной растворенным кислородом.
Однако кислородная атмосфера, столь богатая этим элементом, среди всех планет Солнечной системы присуща только Земле. Это не случайно: высокая химическая активность кислорода приводит к тому, что в условиях планет кислород в свободном состоянии долго существовать не может: участвуя в различных химических реакциях, он оказывается связанным в виде окислов и других соединений. Обилие кислорода в атмосфере современной Земли - результат фотосинтеза, осуществляемого в течение 3 млрд лет зелеными растениями. В процессах фотосинтеза из углекислого газа и воды с использованием энергии солнечного света синтезируются органические вещества (первичная биопродукция) и выделяется свободный кислород.
СО2 + Н2О --> С(Н2О) + О2 + 502 кДж/моль
Первичная атмосфера Земли, существовавшая во времена зарождения жизни, 3,5-4 млрд лет назад, имела восстановительный характер и состояла, вероятно, из водорода, азота, паров воды, углекислого газа, аммиака, метана, аргона и небольших количеств других газов, в том числе и кислорода. Количество свободного кислорода в первичной атмосфере не могло превышать 0,001 от современного его содержания. Это небольшое количество кислорода выделялось в результате фотодиссоциации воды ультрафиолетовыми лучами; кислород быстро входил в различные химические реакции и вновь оказывался в химически связанном состоянии.
Зарождение жизни произошло в бескислородной среде, и кислород из-за своей высокой окислительной способности первоначально был ядовит для протоорганизмов, у которых отсутствовали соответствующие защитные биохимические системы. Вероятно, протоорганизмы по способу питания являлись гетеротрофами, использовавшими в пищу различные органические соединения абиогенного происхождения, которыми, по мнению большинства ученых, были обогащены водоемы раннего археозоя (состояние "первичного бульона", по А.И.Опарину) и на базе которых и возникла сама жизнь. Для освобождения энергии, необходимой в жизненных процессах, первоначально использовалась анаэробная диссимиляция (брожение):
С6Н12О6 --> 2СН3СН2СН + 2СО2 + 210 кДж/моль
С появлением фотосинтеза (первыми фотосинтезируюшими организмами были синезеленые водоросли) в атмосферу стал выделяться кислород.
В условиях бескислородной атмосферы распространение жизни было гораздо более ограниченным, чем ныне. Дело в том. что организмы не имеют эффективных механизмов защиты от гибельной жесткой ультрафиолетовой части излучения Солнца (с длиной волны менее 250 им). В современной атмосфере жесткая ультрафиолетовая радиация поглощается озоновым экраном - слоем озона (О3), образующегося на высоте около 50 км из кислорода (О2) под воздействием солнечного излучения и распределяющегося в основном в 15-60 км от земной поверхности. Озоновый экран надежно защищает живые организмы, которые могут существовать как в водоемах, так и на поверхности суши и в нижних слоях атмосферы. В бескислородной атмосфере раннего протерозоя озоновый экран отсутствовал, и жизнь могла развиваться только под зашитой слоя воды толщиной около 10 м. Получающие наибольшие количества энергии солнечного излучения поверхностные слои воды были недоступны для организмов. Естественно, что совершенно безжизненны были и материки.
Однако фотосинтез, осуществлявшийся в океане планеты синезелеными, а позднее и различными группами эукариотических водорослей, в течение 2 млрд лет медленно, но неуклонно повышал содержание свободного кислорода в атмосфере. Когда содержание кислорода достигло 0,01 от современного (так называемая точка Пастера, соответствующая парциальному давлению кислорода 1,59 мм ртутного столба), у организмов впервые появиласьвозможность использовать для удовлетворения своих энергетических потребностей аэробную диссимиляцию. Другими словами, после достижения точки Пастера стало возможно дыхание, которое почти в 14 раз энергетически эффективнее, чем брожение:
С6Н12О6 + 6О2 --> 6СО2 + 6Н2О + 2,87 мДж/моль - по сравнению с 210 кДж/моль, освобождающимися при брожении. Это был важнейший переломный момент в развитии жизни.
Среди современных организмов так называемые факультативные аэробы, каковыми являются многие бактерии и некоторые дрожжевые грибы, при уменьшении содержания кислорода ниже точки Пастера используют брожение, при повышении его содержания выше этой точки - дыхание (эффект Пастера).
Переход к аэробной диссимиляции в эволюции древних организмов произошел, разумеется, не сразу - для этого необходимо развитие соответствующих ферментативных систем, - но виды, приобретшие способность к дыханию, получили огромный энергетический выигрыш и в результате возможность резко интенсифицировать метаболизм и все жизненные процессы. (как выяснилось, ферментная система клеточного дыхания возникла путем небольшой модификации ферментной системы фотосинтеза - см. обзор по происхождению эукариот; и впрямь, ведь эти два процесса основаны на почти одной и той же последовательности хим. реакций, только идущей в противоположные стороны; при этом надо учитывать, что практически все биохимические реакции обратимы. Это замечательный пример "смены функций" на молекулярном уровне, показывающий к тому же, что даже крупнейшие эволюционные новообразования могут возникать путем небольших модификаций чего-то уже имеющегося в организме - А.М.) . Это явилось предпосылкой к дальнейшей прогрессивной эволюции и, вероятно, способствовало ускорению эволюционных преобразований.
Но достижение пастеровской точки в развитии атмосферы Земли Знаменовалось не только появлением возможности аэробной Диссимиляции. При содержании кислорода в атмосфере в количестве 0,01 от современного формируется озоновый экран, который может защитить от жесткой ультрафиолетовой радиации уже и верхние слои воды в водоемах (требуется "помощь" лишь примерно 1 м воды). Это, во-первых, позволяет организмам освоить верхние слои водоемов, наиболее богатые солнечной энергией; в результате резко усиливается эффективность фотосинтеза, увеличивается биопродукция и выделение свободного кислорода. Во-вторых, чрезвычайно расширяется арена жизни: условия в водоемах значительно разнообразнее на малых глубинах, чем на больших. Освоение этого разнообразия условий в богатой энергией среде обитания неминуемо должно было привести к резкому повышению разнообразия форм жизни, к подлинному взрыву формообразования.
По расчетам Беркнера и Маршалла, точка Пастера в эволюции атмосферы Земли была пройдена примерно 620 млн. лет назад; по мнению некоторых других ученых, возможно, значительно раньше - в промежутке 700-1000 млн лет назад. Но, во всяком случае, точка Пастера была пройдена в позднем протерозое, незадолго (в геологическом смысле слова) до рубежа нижнего кембрия. В этом Беркнер и Маршалл видят ключ к решению загадки нижнекембрийского взрыва формообразования в эволюции организмов, который последовал за достижением точки Пастера в атмосфере и логически вытекает из последствий этого события (интенсификация метаболизма, освоение множества новых разнообразных местообитаний, усиление фотосинтеза, возрастание биопродукции, убыстрение эволюции).
После достижения содержания кислорода в атмосфере, равного 0,1 от современного, озоновый экран уже в состоянии полностью защитить организмы от действия жесткой ультрафиолетовой радиации. С этого момента организмы могут начать освоение суши как среды обитания. По расчетам Беркнера и Маршалла, это должно было произойти в конце ордовика (около 420 млн лет назад). Действительно, примерно к этому времени относится появление первых наземных организмов (см. гл. 2). (По мнению ряда других ученых, содержание кислорода в атмосфере соответствующее 10% от современного, было достигнуто уже к началу кембрия, примерно 580 млн лет назад).
Современное содержание кислорода в атмосфере было достигнуто в конце пермского периода.
Гипотеза Беркнера и Маршалла привлекательна не только своей логичностью и последовательностью, но и перспективностью дальнейшего развития этих идей. Как мы уже упоминали, на рубеже нижнего кембрия у самых различных групп организмов развивается твердый скелет, облегчающий их фоссилизацию. Формирование скелета может быть также непосредственным следствием повышения содержания кислорода в атмосфере. Как показали Р. и Е. Раффы, при низком содержании кислорода в окружающей среде размеры тела многоклеточных животных не могли быть большими (вследствие низкого уровня метаболизма и энергетики организмов); газообмен с внешней средой, вероятно, осуществлялся диффузно, через поверхность тела; при этом толщина стенок тела не могла превышать нескольких миллиметров. Для таких организмов не возникало необходимости в опорном внутреннем скелете, а защитные наружные скелетные образования препятствовали бы газообмену. К.Тоув пришел к выводу, что у докембрийских животных в условиях малого содержания кислорода в окружающей среде не могло быть также хорошо развитых соединительнотканных образований, формирующих основу для развития скелета. Прочность соединительнотканных структур основывается на содержании в них белка коллагена, в состав которого входит аминокислота оксипролин. Ее образование возможно только при достаточно высоком содержании кислорода в окружающей среде. Следовательно, при низком содержании кислорода синтез коллагена был биохимически затруднен, и организмы не могли иметь прочных соединительнотканных структур, а поэтому и скелета, и сильно развитой мышечной системы (работа которой эффективна лишь при наличии соответствующих опорных образований).
Д. Роудс и Дж. Морзе исследовали распространение различных животных в современных водоемах с пониженным содержанием кислорода в воде (в Калифорнийском заливе и в Черном море). Была обнаружена отчетливая корреляция между содержанием кислорода в воде и характером донной фауны (бентоса). При содержании кислорода менее 0,1 мл на 1 л воды многоклеточные животные в составе бентоса отсутствуют; при 0,3-1 мл/л встречаются небольшие мягкотелые (бесскелетные) животные, зарывающиеся в ил; наконец, в более поверхностных слоях с содержанием кислорода более 1 мл/л обитают самые разнообразные животные, обладающие известковым скелетом. Эти данные представляют своего рода живую иллюстрацию к концепции Беркнера и Маршалла.
Подведем некоторые итоги. Обособление большинства типов животных произошло, вероятно, в позднем протерозое, 550-800 млн лет назад. Примитивные представители всех групп многоклеточных были небольшими лишенными скелета животными. Продолжавшееся накопление кислорода в атмосфере и увеличение мощности озонового экрана к концу протерозоя позволили животным Увеличить размеры тела. Организмы получили возможность широко расселиться на малых глубинах различных водоемов, что способствовало значительному повышению разнообразия форм животных (вендская фауна).
Однако в позднем венде эта древняя фауна многоклеточных Животных подверглась значительному вымиранию, которое сопровождалось новым существенным уменьшением размеров тела организмов, переживших вымирание. Возможно, этому способствовало чрезвычайно мощное оледенение (по некоторым данным, среднегодовая температура Земли понижалась в это время примерно до 5°С). К концу венда ледники отступили, и условия вновь стали благоприятными для широкого расселения организмов.
(Более подробно о возникновении жизни - см. http://warrax.croco.net/51/eskov/cover_eskov.html) .
Арена жизни в палеозое
На протяжении палеозоя море неоднократно наступало на континент, заливая опускавшиеся участки континентальных платформ - морские трансгрессии начала кембрия, первой половины ордовика, когда была затоплена наибольшая за весь фанерозой часть континентов, раннего силура, девона и карбона. Морские трансгрессии сменялись поднятием суши, сопровождавшимся горообразовательными процессами и морскими регрессиями (отступление морей) во второй половине кембрия, в конце ордовика, в самом начале и в конце силура, в середине карбона и в пермском периоде. Мощные горные хребты поднялись в конце силура в Скандинавии, Шотландии, Гренландии, на востоке Северной Америки и в ряде других районов земного шара (Каледонская фаза горной складчатости), в каменноугольном и пермском периодах - в Средней Европе (Арденны, Судеты, Гарц, Шварцвальд и др.), на Урале и в Средней Азии (Алай, Тянь-Шань, Ку-энь-Лунь), на севере Сибири, в Австралии и др. (Герцинская фаза складчатости).
Согласно классической теории, которая берет начало в работах Ч.Ляйеля, происходившие в геологической истории изменения границ суши и моря объясняются вертикальными смешениями участков земной коры. Однако ныне все большее признание получает концепция мобилизма -- горизонтальных перемещении континентальных блоков, или "дрейфа континентов", выдвинутая в начале XX в. А. Вегенером. Современный вариант этой концепции - теория тектоники литосферных плит, или "новая глобальная тектоника" - - основывается на глобальном синтезе результатов геолого-геофизических наблюдений и во многом отличается от взглядов Вегенера.
По современным геофизическим данным, астеносфера - верхний слой верхней мантии Земли, располагающийся на глубинах 60-250 км, - имеет пониженную вязкость. Предполагают, что в мантии существуют конвекционные потоки ее вещества, источником энергии для которых являются, вероятно, радиоактивный распад и гравитационная дифференциация вещества мантии. Движение этих потоков со скоростью несколько сантиметров в год происходит в горизонтальной плоскости в верхнем слое верхней мантии от областей подъема к областям опускания ее вещества. В это движение вовлекаются литосферные плиты, как бы плавающие в состоянии изостатического равновесия на поверхности астеносферы и служащие основанием для континентов. При столкновении континентальных плит их края деформируются с возникновением зон складчатости и проявлений магматизма. При столкновении континентальной и океанической плит последняя как бы подминается первой и растекается под ней в астеносфере. Концепция мобилизма в ее современном виде не свободна от противоречий и, вероятно, еще подвергнется существенным перестройкам. Однако уже и теперь она дает удовлетворительное объяснение целому ряду как геологических, так и биологических проблем. К последним относятся, в частности, характерные особенности географического распространения многих видов животных и растений, а также некоторые исторические изменения фауны и флоры.
Согласно представлениям тектоники плит положение и очертания материков и океанов в палеозое были совершенно иными, чем теперь. Палеогеографические реконструкции, выполненные различными учеными с использованием разных методов, довольно существенно различаются в деталях. Однако большинство авторов согласны, что в кембрии южные материки (Южная Америка, Африка, Австралия, Антарктида, а также Индия) были объединены в суперконтинент, называемый Гондваной (рис. 34). Гондвана располагалась главным образом в Южном полушарии, от полюса (находившегося в современной Северо-Западной Аф-Рике) до экватора. В основном в экваториальной зоне находились меньшие по размерам северные материки: Северо-Американский, Восточно-Европейский, Сибирский и микроконтиненты (Средне-Европейский, Казахстанский и др.). Между ними и Гондваной был океан.
В ордовикском периоде северные континенты начинают постепенно сближаться друг с другом. В силуре и девоне Северо-Амери-канский континент объединился с Восточно-Европейским, с образованием Евроамерики. Пояс Каледонской складчатости в Северо-Западной Европе и Восточной Северной Америке сформировался при столкновении этих континентальных плит. В раннем карбоне сближение континентов продолжалось. Этому соответствовало возникновение поясов Герцинской складчатости. Процесс сближения континентов завершился во второй половине палеозоя (поздний карбон - ранняя пермь) образованием гигантского суперконтинента Пангеи, протянувшегося от Южного полюса, который сместился к этому времени в Антарктиду, далеко в Северное полушарие. Северные материки образовали в составе Пангеи комплекс, получивший название Лавразии.
Перемещения континентальных плит, сопровождавшиеся изменениями рельефа суши и морского дна, обусловливали изменения направлений морских течений и атмосферной циркуляции. Сложные взаимодействия всех этих факторов определяли климатические изменения как местного, так и глобального характера. Согласно Ч.Бруксу, относительное увеличение площади суши в высоких широтах, особенно расположение полюса на суше, вызывает общее понижение температуры поверхности планеты, тогда как увеличение площади суши в низких широтах и расположение полюсов в океане - повышение температуры.
В первой половине палеозойской эры на Земле господствовал в целом теплый климат: средняя температура поверхности Земли была сравнительно высокой, а перепад температур между полюсами и экватором невелик, климатическая зональность была выражена относительно слабо. Теплый период в конце палеозоя, начиная со второй половины карбона, сменился холодным, с более низкой средней температурой, резко выраженной климатической зональностью и значительной разницей температур между полюсами и экватором. Возможно, это связано с расположением в это время центральной части Гондваны в районе Южного полюса. В позднем карбоне и ранней перми мощный ледниковый щит покрывал Антарктиду, Австралию, Индию, южные части Африки и Южной Америки - это было время великого оледенения Гондваны. В верхнепалеозойских отложениях всех этих районов широко распространены тиллиты - ископаемые остатки древних ледниковых морен. В первой половине палеозоя тиллиты имели гораздо более ограниченное распространение; по-видимому, оледенение в конце ордовика и девоне затрагивало лишь приполярные районы (Северо-Западная Африка в ордовике, Аргентина в силуре и девоне). Северный полюс в палеозое находился в океане.
Жизнь в палеозойских морях и пресных водоемах
Вероятно, в кембрийском периоде, как и в докембрии, основные проявления жизни были сосредоточены в морях. Однако организмы уже заселили все разнообразие доступных в море местообитаний, вплоть до прибрежного мелководья, и, возможно, проникли в пресные водоемы.
Водная флора была представлена различными водорослями, основные группы которых возникли еще в протерозое.
Теплые моря кембрийского периода были обильно населены разнообразными животными. Наиболее богата была донная фауна неглубоких морей, прибрежных отмелей, заливов и лагун (во время морских трансгрессий мелководные местообитания получают очень широкое распространение). На морском дне жили разнообразные прикрепленные животные: губки, кишечнополостные (одиночные и колониальные представители различных групп полипов), стебельчатые иглокожие, плеченогие и др. Большинство из них питалось различными микроорганизмами (простейшие, одноклеточные водоросли, бактерии и др.), которых они различными способами отцеживали из воды (фильтрационное питание). Некоторые колониальные организмы, обладавшие известковым
скелетом, возводили на дне моря рифы, подобно современным коралловым полипам. В раннем палеозое это были гидроидные полипы-строматопоры (подкласс Stromatoporoidea), древние коралловые полипы-табуляты (подкласс Tabulata), мшанки (тип Bryozoa) и археоциаты (тип Archaeocyatha). Археоциаты - группа примитивных прикрепленных животных, широко распространенных в раннем кембрии, но полностью вымерших к концу силура. Археоциаты имели известковый скелет в виде своеобразного кубка (рис. 35), стенки которого пронизывали многочисленные поры. Размеры таких кубков варьировали от нескольких миллиметров до 40 сантиметров. Снаружи кубок был одет мягкой оболочкой. Мягкие ткани внутри кубка, видимо, имели губчатое строение, будучи пронизаны системой полостей.
Ряд других групп прикрепленных морских животных, также переживших расцвет в палеозойской эре, сохранились до наших дней в лице немногих видов. Таковы, например, стебельчатые иглокожие (подкласс Pelma-tozoa), представленные в палеозое 7 классами, из которых до современности дожили лишь морские лилии (Crinoidea). Сходная судьба была и у плеченогих - своеобразных прикрепленных животных, снабженных двустворчатой раковиной и внешне напоминающих двустворчатых моллюсков, но резко отличающихся от них внутренним строением. Раковина плеченогих имеет спинную и брюшную створки, а не две боковые, как у моллюсков. Замечательно, что с кембрийского периода до наших дней просуществовал род брахиопод Lingula (см. рис. 17), который, судя по ископаемым остаткам, сохранил свое строение практически без изменений свыше 500 млн лет, став одним из наиболее ярких примеров эволюционного консерватизма.
С морским дном были тесно связаны и многие другие группы животных. Различные черви приспособились к роющей жизни в толще донных осадков. Среди них были примитивные вторичноротые - кишечнодыша-Щие (Enteropneusta), принадлежащие к типу полухордовых. Наиболее древние ископаемые остатки этих животных были обнаружены в среднекембрийских отложениях (фауна Бёр-Джес), но, несомненно, возникла эта группа еще в докембрии.
Среди водорослей и кораллов по морскому дну медленно передвигались свободноживущие иглокожие (представители подтипа Eleutherozoa - морские звезды, офиуры, голотурии и др.) и моллюски, основные группы которых известны, начиная с кембрия. Свободноживущие иглокожие возникли, вероятно, от прикрепленных предков, унаследовав от них внешнюю радиальную симметрию тела. Организация моллюсков сложилась в процессе приспособления древних червеобразных животных к питанию различными донными обрастаниями и остатками погибших организмов, пищей обильной и легкодоступной. Такой способ питания не требовал большой подвижности. А для защиты мягкого тела у моллюсков развилась прочная и тяжелая известковистая раковина, еще более ограничившая их подвижность.
Но одна из групп моллюсков, вопреки общей эволюционной тенденции этого типа к развитию малоподвижных донных форм, уже в кембрии сумела перейти к освоению свободного плавания. Это были первые головоногие -- наутилоидеи, или кораблики (Nautiloidea), дожившие в лице единственного рода Nautilus (рис. 36) до наших дней. Раковина корабликов преобразовалась в гидростатический аппарат, позволяющий этим моллюскам изменять свою плавучесть, поднимаясь или опускаясь в толще воды без мышечных усилий. Раковина разделена поперечными перегородками на камеры, тело животного помещается в последней, самой большой из них. Перегородки пронизаны сифоном - длинным выростом тела, в котором проходят кровеносные сосуды. Камеры раковины заполнены жидкостью, которая может всасываться в кровь и затем выводиться из организма почками; вес раковины уменьшается, и моллюск всплывает. При погружении происходят обратные процессы. Активное плавание головоногих в толще воды происходит посредством выброса струи воды из мантийной полости через специальную воронку - животное движется реактивным способом.

В девоне появились и другие группы головоногих моллюсков, в частности аммониты (Ammonoidea), имевшие, как и кораблики, спирально закрученную раковину. А в нижнем карбоне возникли первые представители высших головоногих, у которых раковина постепенно редуцировалась и оказалась заключенной внутри мягких тканей •тела (подкласс внутреннераковинных, Endocochlia). Эти преобразования были связаны с дальнейшей активизацией образа жизни головоногих. Древнейшими внутрен-нераковинными были белемниты (Belem-nitida), которые еще имели довольно большую удлиненно-коническую раковину (рис. 37), лишь прикрытую мантийной складкой тела1. В ископаемом состоянии наиболее часто сохраняется ростральная часть раковины белемнитов. Аммониты и белемниты играли видную роль в морской фауне верхнего палеозоя и мезозоя.

В толще и на поверхности воды в морях обитали и другие животные. Более примитивные группы использовали пассивное плавание, дрейфуя по течению и удерживаясь на поверхности с помощью особых плавательных пузырей или "поплавков", заполненных газом. Таковы плавающие колониальные кишечнополостные сифонофоры (Siphonophora), живущие в морях и поныне.Впервой половине палеозоя подобный образ жизни вели граптолиты (подтип Graptolithina типа полухордовых, рис. 38). Среди граптолитов были также "псевдопланктонные" виды, не имевшие поплавков и прикреплявшиеся к плавающим водорослям. Были и формы, прикреплявшиеся к морскому дну. Граптолиты, появившиеся в кембрии, полностью вымерли в раннем карбоне (большинство их исчезли еще до конца силура).

Уже в раннем кембрии существовали и столь высокоорганизованные животные, как членистоногие, представленные тремя обособленными подтипами: жабродышащими (Вгап-chiata), хелицеровыми (Chelicerata) и трилобитами (Trilobita). Последний подтип - трилобиты (рис. 39) - чисто палеозойская группа, достигшая расцвета в раннем кембрии, когда они составляли до 60% всей фауны. Трилобиты сохраняли свои позиции до середины ордовика, после чего началось их угасание. Они окончательно вымерли к середине пермского периода. Для трилобитов характерно расчленение защищавшего тело спинного щита двумя продольными бороздами на 3 доли (отсюда и название подтипа). Эти членистоногие были чрезвычайно разнообразны. Их размеры варьировали от 1 до 75 см, хотя большинство не превышало 10 см. Трилобиты были в основном придонными животными. Они ползали по дну на членистых конечностях, состоявших каждая из двух ветвей, но могли, вероятно, и плавать в толще воды (среди трилобитов были даже формы, специализированные к плаванию), и зарываться в песок. По способу питания большинство трилобитов были, по-видимому, микрофагами, отцеживавшими из воды мелкий планктон или поедавшими ил.
Среди водных палеозойских членистоногих были и хищники, из которых особенно выделяются эвриптериды (отряд Eurypterida, принадлежащий к подтипу хелицеровых), родственные современным скорпионам, но достигавшие длины около 2 м (рис. 40). Это были самые крупные членистоногие, когда-либо существовавшие на Земле. Эвриптериды известны, начиная с ордовика. Они достигли наибольшего расцвета в силуре и первой половине девона, но уже с середины девона число их видов начинает сокращаться. В нижней перми эти крупные хелицеровые исчезают. Возможно, судьба эвриптерид определилась тем, что примерно с середины девона в роли крупных водных хищников с ними начинают конкурировать различные группы хищных рыб. Позвоночные (подтип Vertebrata типа хордовых) оказались лучше приспособленными к быстрому плаванию в толще воды. Проблема передвижения у древних позвоночных была решена принципиально иначе, чем у членистоногих, которые плавали посредством гребных движений конечностей. У позвоночных развитие внутреннего осевого скеле-
|та (хорды, а затем позвоночника) позволило использовать практически всю мускулатуру тела для создания силы поступательного движения посредством волнообразных изгибаний туловища и мощных ударов основного движителя - хвостового плавника. После развития челюстей (см. стр. 130-131) позвоночные получили преимущество перед крупными членистоногими и в приспособлениях нападения и защиты: костные или хрящевые челюсти позвоночных, расположенные в толще приводящих их в движение мышц, механически прочнее и надежнее, чем наружные хитиновые челюсти членистоногих, возникшие из членистых конечностей.
Благодаря этим преимуществам позвоночные уже в девоне заняли доминирующее положение среди активно плавающих (нектонных) животных с крупными размерами тела. Членистоногие же оказались лучше приспособленными среди относительно мелких организмов: громадное большинство членистоногих ныне имеют небольшие размеры. Зато в этой сфере членистоногие развили ни с чем не сравнимое разнообразие мелких форм.
Позвоночные (Vertebrata) вместе с бесчерепными (Acrania) и оболочниками (Tunicata) входят в состав типа хордовых (Chordata). Позвоночные появляются в палеонтологической летописи, начиная с нижнего ордовика. Более древние хордовые животные, которые были общими предками позвоночных, бесчерепных и оболочников, до самого последнего времени оставались в ископаемом состоянии неизвестными. В связи с этим о происхождении хордовых выдвигалось много различных гипотез (вплоть до совершенно невероятных предположений о возникновении этих животных от головоногих моллюсков или от членистоногих, общий план строения которых принципиально отличается от такового хордовых). Выше мы уже рассматривали происхождение всех вторичноротых животных, вероятными общими предками которых были олигомерные черви, близкие по своей организации к современным кишечнодышащим полухордовым (Enteropneusta). Если эволюция полухордовых и иглокожих была связана с развитием приспособлений к жизни на морском дне (прикрепленной или роющей в толще донных осадков), древние хордовые, вероятно, перешли к активному плаванию в толще воды. Приспособления к нектонно-му образу жизни и создали предпосылки для дальнейшей прогрессивной эволюции этой группы. При активизации плавания отбор благоприятствовал усовершенствованию нервной системы, органов чувств и локомоторного аппарата. С этим связано развитие метамерии мышц тела и возникновение хорды - уникального внутреннего скелета. Хорда представляет собой продольный опорный тяж, прочность которого основана на несжимаемости жидкого содержимого особых вакуолей (принцип гидроскелета). Кроме того, в хорде имеются сократимые волокна, подобные мышечным; их сокращение повышает жесткость данного участка хорды и способствует быстрому выпрямлению соответствующего сегмента тела.
А.Н.Северцов сопоставил особенности организации разных подтипов хордовых, выбирая общие для всех групп признаки, которые можно истолковать как примитивные, и обрисовал гипотетический облик древнейших хордовых животных, потомков олигомерных червей и предков позвоночных, бесчерепных и оболочников. Этих гипотетических протохордовых А. Н.Северцов назвал примитивными бесчерепными (Acrania primitiva). Примитивные бесчерепные (рис. 41) обладали билатерально симметричным, сжатым с боков телом без обособленной головы и снабженным спинным и хвостовым плавниками. Скелет представлен хордой и соединительнотканными структурами. Пищеварительная система, как у всех низших хордовых, начиналась большой глоткой с эндостилем, наджаберной бороздой и жаберными щелями, пронизывающими ее боковые стенки; сзади глотка продолжалась коротким кишечником с печеночным выростом. Питание осуществлялось путем слизевой фильтрации: отцеживание из воды, прогоняемой мерцательным эпителием через глотку и жаберные щели, взвешенных в ней микроорганизмов и частиц ила, слизевым фильтром на жаберных щелях. Над хордой тянулась продольная нервная трубка - центральная нервная система. Кровеносная система включала продольные сосуды-аорты: брюшную (под глоткой) и спинную (под хордой), соединенные жаберными артериями. Мышцы всего тела имели метамерное строение.
В 1995-1999 гг. китайские палеонтологи Ж.Я.Чен, Г.К.Чжоу, Ч.В.Ли и др. сделали ряд замечательных находок ископаемых остатков древнейших хордовых животных в нижнекембрийских отложениях Юнънаня (Южный Китай). В тонкослоистых глинистых сланцах смогли сохраниться отпечатки структур, образованных мягкими тканями. Строение этих раннекембрийских хордовых. Названных юннанозооном (Yunnanozoon) и хайкоуэллой (Haiko-uella), оказалось очень близким к организации гипотетических Примитивных бесчерепных, обоснованной А. Н.Северцовым. Хай-Коуэлла, которая изучена более полно, была небольшим (25-40 мм) животным с ланцетовидным, сжатым с боков телом, дифференцированным на голову, туловище и короткий хвост. Имелись спинной, хвостовой и брюшной непарные плавники. Вдоль всего тела тянулась хорда. Мышцы тела были разделены на сегменты и сильнее развиты в спинной части туловища. На нижней стороне головы располагался окруженный небольшими щупальцами рот, который вел в объемистую глотку. По ее дну тянулся железистый желобок-эндостиль, а боковые стенки были пронизаны 6-7 парами жаберных щелей и укреплены жаберными дугами, на которых находились жаберные нити. Над хордой тянулся продольный тяж центральной нервной системы, который у хайкоуэллы в голове был дифференцирован как головной мозг с 3 отделами. Возможно, у хайкоуэллы имелись и парные глаза. Позади глотки находилось сердце, от которого кпереди тянулась под глоткой брюшная аорта, связанная жаберными артериями со спинной аортой.
Вероятно, расхождение филогенетических стволов ранних хордовых, которые дали начало подтипам позвоночных, бесчерепных и оболочников, произошло в самом раннем кембрии. Возможно, юннанозоон был близок к предкам бесчерепных, а хайкоуэлла - к предкам позвоночных. Ископаемые остатки древнейших представителей оболочников (Palaeobotridium) обнаружены в верхнекембрийских отложениях Невады в Северной Америке.
С позднего кембрия до позднего триаса существовала загадочная группа древних животных - конодонты, или конодонтофо-риды (Conodontophorida), долгое время известные лишь по многочисленным микроскопическим конусовидным зубам (которые, собственно, и именуются конодонтами), укрепленным небольшими группами или поодиночке в зубных пластинках. В 1983 г. в нижнем карбоне Шотландии были обнаружены ископаемые остатки мягкого тела животного (Clydagnathus), оотовой аппарат которого образован конодонтами. Это существо имело червеобразное уплощенное с боков тело длиной около 5 см, пару больших глаз, хвостовой плавник и метамерные мышцы. Электронно-микроскопические исследования показали, что конодонты образованы тканями, характерными для позвоночных животных, - хрящом, костью и эмалеподобным веществом. Д. Бриггс, Р.Олдридж и ряд других палеонтологов считают, что конодон-тофориды принадлежали к низшим хордовым и близкородственны с позвоночными или даже входят в состав последних.
Древнейшие позвоночные были рыбообразными животными, лишенными челюстей, с телом, защищенным панцирем (рис. 42). Отсюда название этой группы - - панцирные бесчелюстные (Ostracodermi). Их панцирь был образован крупными щитами и мелкими щитками и пластинками, состоящими из 3 слоев примитивных костных тканей (в том числе аспидина и дентина). Ас-пидиновые щитки были снаружи покрыты своеобразной шагренью из мелких зубчиков, образованных дентином и, вероятно, эмалью (подобной зубной). Голову этих животных сверху и сбоку, а иногда и снизу защищал сплошной головной щит с отверстиями для глаз, непарного органа обоняния и так называемого "теменного глаза" - светочувствительного верхнего придатка промежуточного мозга, игравшего у примитивных позвоночных важную роль в регуляции различных функций организма в соответствии с суточным и годовым циклами и с уровнем освещенности внешней среды. Задняя часть тела была покрыта небольшими щитками, не препятствовавшими изгибанию этого отдела и работе хвоста. Настоящих парных конечностей у остракодерм, видимо, еще не было, но по бокам тела позади головы имелась пара покрытых чешуями придатков или плавниковых складок.
Наличие у некоторых остракодерм хорошо окостеневшего черепа позволило выяснить ряд важных особенностей внутреннего строения этих животных. Оказалось, что многие признаки остракодерм Поразительно сходны с соответствующими особенностями современных миног и миксин, принадлежащих к классу круглоротых (Cyclostomata). Помимо отсутствия челюстей у остракодерм и круглоротых сходно строение жаберного аппарата (жаберные мешки, Расположенные внутри решетки скелетных висцеральных дуг), непарного органа обоняния, внутреннего уха (лишь с 2 полукружными каналами), головного мозга и др. Однако круглоротые, в отличие от своих палеозойских родственников, совершенно лишены костной ткани (их скелет представлен лишь хордой и хрящами). В ископаемом состоянии круглоротые почти не известны; лишь недавно в среднекарбоновых отложениях Иллинойса в США были открыты ископаемые остатки миног (Mayomyzon) и мик-син (Myxinikela). Может быть, к предкам круглоротых близок Palaeospondylus - загадочная среднедевонская форма с хорошо сохранившимся позвоночником, черепом, напоминавшим череп круглоротых, и хвостовым плавником, но без всяких признаков костного панциря.
Современные круглоротые являются полупаразитами (миноги) и трупоедами (миксины). Остракодермы, вероятно, питались илом или отцеживали из воды микропланктон с помощью слизевого фильтра на жаберных щелях, как и личинки современных миног. У некоторых остракодерм обнаружены следы структуры, сходной с эндостилем - железистым желобком на дне глотки, клетки которого выделяют слизь для слизевой фильтрации.
Наличие хорошо окостеневшего скелета у раннепалеозойских остракодерм при отсутствии окостенений у круглоротых привело к тому, что мнения исследователей о соотношениях скелетных тканей в эволюции и онтогенезе позвоночных разделились. А.Н.Северцов, Д.В.Обручев и ряд других ученых аргументировали теорию, согласно которой древнейшие позвоночные не имели костного скелета и поэтому не сохранились в ископаемом состоянии. Костный скелет начал формироваться в покровах в виде отдельных "кожных зубов", подобных мелким плакоидным чешуям современных хрящевых рыб - акул и скатов. В дальнейшем происходило срастание этих чешуи друг с другом с образованием все более массивных комплексов. Из верхнесилурийских и девонских отложений известны остатки остракодерм (Thelodus; см. рис. 42), лишенных тяжелого костного панциря, но покрытых плакоидными чешуйками.
Однако все более древние остракодермы (из ордовика и раннего силура) имели хорошо развитый костный скелет, причем плакоидные чешуйки образовывали наружный слой щитков панциря. В связи с этим палеонтологи А.Ш.Ромер, Л.Хальстед и др. считают более вероятным, что элементы костного скелета появились у древнейших позвоночных по крайней мере столь же давно, как и хрящевые структуры. В сущности, хрящ и кость представляют собой лишь крайние состояния скелетных тканей позвоночных, связанные промежуточными формами. Хрящ более пластичен, менее прочен, чем кость, и способен как к поверхностному, так и внутреннему росту. Благодаря этому хрящ является лучшей эмбриональной скелетной тканью, а кость - лучшей основой скелета взрослых позвоночных.
В филогенезе позвоночных происходила активизация образа жизни: переход от придонной жизни к нектонной, от бенто- и планктонофагии к хищничеству. При этом отбор благоприятствовал освобождению тела от тяжкого бремени защитного костного панциря, характерного для ранних остракодерм. Облегчение тела происходило по-разному: либо путем постепенного преобразования крупных щитов в гибкий и легкий чешуйный покров, либо посредством редукции костной основы - с сохранением поверхностных плакоидных чешуи (телодонты, хрящевые рыбы) или с полной утратой и этих последних (круглоротые). Второй путь преобразований (полная или частичная утрата костной ткани) был, очевидно, проще по механизму перестроек онтогенеза (с задержкой развития скелета на эмбриональной стадии) и мог поэтому осуществиться быстрее, чем более сложное постепенное преобразование скелетных структур, требовавшее существенных перестроек морфогенетических механизмов.
В филогенезе позвоночных были испробованы разнообразные варианты перестроек примитивного тяжелого скелета; судьба разных эволюционных ветвей при этом сложилась различно. В конечном итоге наибольший успех выпал на долю тех групп позвоночных, которые избрали более медленный, но гораздо более перспективный путь усовершенствования костного скелета, а не отказались от него полностью. Поэтому утрату костного скелета у таких групп, как круглоротые или хрящевые рыбы, можно с полным основанием рассматривать как пример инадаптивной эволюции, как ее понимал В. О. Ковалевский.
Остракодермы пережили свой расцвет в силуре и первой половине девона. К концу девона панцирные бесчелюстные вымирают, вытесненные более прогрессивными группами позвоночных, принадлежащими к стволу челюстноротых (Gnathostomata). Челюстноротые появились в палеонтологической летописи в позднем силуре (остатки древнейших рыб из группы акантодий - Acanthodii); в девоне существовали уже разнообразные группы всех классов рыб. Их общие предки пока остаются неизвестными. Некоторые палеонтологи допускают возможность происхождения челюстноротых от примитивных остракодерм из группы разнощитковых (Heterostraci), которые не имели таких специфических особенностей бесчелюстных, как непарный орган обоняния, открывающийся на верхней стороне головы, энтодермальные жаберные мешки и др. Расхождение филогенетических стволов Позвоночных, ведущих к бесчелюстным (классы остракодерм и Круглоротых) и челюстноротым (все остальные классы позвоночных) произошло, вероятно, в позднем ордовике или раннем силуре. Это расхождение было связано с различными путями интенсификации жаберного дыхания, рассмотренными нами выше. Как мы видели, преобразования висцерального скелета на пути, избранном челюстноротыми, привели к формированию челюстного аппарата, наличие которого вместе с развитием парных конечностей стало важнейшим преимуществом, обеспечившим победу челюстноротых в конкуренции с бесчелюстными. Современные круглоротые насчитывают всего лишь около 50 видов против примерно 20000 видов различных групп рыб.
Древнейшие представители челюстноротых, обитавшие в морях и пресных водоемах раннего и среднего девона, были, как и остракодермы, одеты более или менее развитым костным панцирем, особенно мощным у панцирных рыб -- плакодерм (класс Placodermi). Панцирь плакодерм (рис. 43) состоял из двух подвижно сочлененных друг с другом частей, одна из которых защищала голову, а другая - переднюю часть туловища. По бокам передней части туловища располагались подвижные придатки, возможно гомологичные передним конечностям других челюстноротых.

У плакодерм не было настоящих зубов, их заменяли острые бивнеобразные выступы челюстных костей (у других групп челюстноротых зубы возникли из плакоидных чешуи кожного панциря, переместившихся по краям рта на челюсти). Среди плакодерм были мелкие придонные формы, вероятно конкурировавшие с остра-кодермами, и крупные хищники, например верхнедевонская ди-нихтис (Dinichthys), у которой лишь одетая панцирем передняя часть тела имела длину около 3,2 м. Плакодермы вымерли в раннем карбоне.
Другой примитивной группой палеозойских челюстноротых были уже упоминавшиеся акантодии (Acanthodii). Их тело покрывали многочисленные костные пластинки и крупные чешуи, были окостенения и во внутреннем скелете. У акантодии еще не установилось обычное для позвоночных число пар конечностей; по бокам тела (рис. 44) тянулся ряд плавников (до 7 пар), возникших, вероятно, в результате дифференциации первоначальных плавниковых складок. В дальнейшем средние плавники, расположенные ближе к центру тяжести тела (и поэтому малоэффективные как рули глубины и стабилизаторы движения) подверглись ре-дукции, так что сохранились лишь передняя и задняя пары. Челюсти у акантодии были связаны с черепом без участия второй висцеральной дуги (подъязычной). Такое состояние также очень примитивно: у всех остальных рыб верхний (гиомандибулярный) элемент подъязычной дуги становится подвеском суставной области челюстной дуги. Акантодии обитали в пресных и морских во-Доемах с позднего силура до середины пермского периода, когда эта древняя группа подверглась вымиранию.
В первой половине девона существовали уже и представители трех подклассов костных рыб (класс Osteichthyes): лучеперых (Actinopterygii), двоякодышащих (Dipnoi) и кистеперых (Crosso-pterygii). Первая группа, эволюция которой была в основном связана с освоением всей толщи воды в пресных водоемах и морях, в палеозое была относительно малочисленна. Оба других подкласса, объединяемые в группу хоановых рыб (Choanichthyes), напротив, пережили расцвет в палеозое. Их организация сформировалась в процессе освоения широко распространенных в палеозое пресных водоемов типа обширных мелководных озер, лагун и эстуариев, хорошо прогреваемых солнцем, обильно заросших водной растительностью и отчасти заболоченных. В воде таких водоемов из-за гниения растений нередко возникал дефицит кислорода. Важнейшим приспособлением к жизни в таких водоемах были легкие как дополнительный орган дыхания, позволявший использовать кислород воздуха при его недостатке в воде. Легкие представляют собой слепые мешковидные выпячивания самой задней части глотки, куда рыба через рот заглатывает воздух, используя для этой цели обычный жаберный насос.
(Вероятно, орган, соответствующий легким, возник очень рано в эволюции костных рыб; его гомологом у актиноптеригий является плавательный пузырь, служащий гидростатическим аппаратом, но используемый примитивными лучеперыми и как легкое. По некоторым данным, этот орган мог возникнуть еще у примитивных челюстноротых - общих предков плакодерм, хрящевых и костных рыб.).
Хоановые рыбы вели в основном скрытный придонный образ жизни, охотясь в гуще растительности. Кистеперые были хищниками, подстерегавшими добычу в укрытиях, а двоякодышащие приспособились к питанию малоподвижными животными с твердым панцирем (моллюски, ракообразные). Плавники хоановых рыб приобрели особое строение, удобное для передвижения у дна водоемов: наружная "рулевая" лопасть, образованная тонкими плавниковыми лучами, редуцировалась, зато был сильно развит их внутренний костный скелет (базальные и радиальные элементы), выдвинутый из тела наружу вместе с мышцами, образуя мясистую лопасть в основании плавника (рис. 45). Такие плавники, называемые саркоптеригиями, позволяли рыбе переползать по дну, скрываясь среди растений.

Для ориентации в подводных зарослях важную роль приобретает обоняние. Чтобы обеспечить постоянный обмен воды в органе обоняния, задние ноздри (у рыб в каждый обонятельный мешок ведут два отверстия - переднее и заднее) перемещаются под верхнюю челюсть, становясь внутренними ноздрями, или хоанами (отсюда название "хоановые рыбы"). Хоаны позволяют вентилировать орган обоняния, используя изменения давления в рото-глоточной полости, которые возникают при движениях жаберных крышек в процессе вентиляции жабер.
Кистеперые рыбы были очень многочисленны в пресных и солоноватых водоемах в девоне и начале карбона, но сокращение их местообитаний при усилении горообразовательных процессов и общем поднятии материков постепенно привело эту группу к упадку. Одна из линий эволюции кистеперых рыб - целаканты (Coelacant-hiformes) - вторично перешли к жизни в море, и два вида этой группы сохранились до нашего времени в водах Мозамбикского пролива (Latimeria chalumnae) и в море Сулавеси (L. menadoensis).
Двоякодышащие в палеозое не были столь широко распространены, как кистеперые. Ныне в пресных, иногда пересыхающих водоемах тропиков еще существуют 6 видов этой группы, относящихся к 3 родам: Neoceratodus в Австралии, Protopterus в Африке и Lepidosiren в Южной Америке. Интересно, что ареал географического распространения современных хоановых рыб соответствует разобщенным частям Гондваны.
Освоение суши
Первые вполне достоверные остатки наземных организмов известны начиная с силура. Заселение безжизненных прежде материков было длительным процессом, развивавшимся в течение нескольких десятков миллионов лет, на протяжении ордовика, силура и девона.
(Имеются некоторые данные и о более древних наземных организмах; так, из отложений среднего ордовика Южного Казахстана были описаны остатки Растения Aedalophyton caradocki, которое интерпретируют как наземное - по наличию кутикулы и устьиц).
Вероятно, начало освоению суши положило появление пленчатых обрастаний прибрежных камней и скал в полосе прибоя и приливно-отливной зоне синезелеными водорослями; в водорослевых обрастаниях поселялись бактерии. В условиях низких континентов раннего палеозоя с очень пологими берегами эта переходная зона между морем и сушей имела значительную протяженность. Сформировавшиеся здесь пленчатые бактериально-водорослевые обрастания ("маты") часто перекрывались неорганическими материалами - - продуктами выветривания горных пород, сносившимися с суши. В результате этих процессов происходило первичное почвообразование. Формирование почвы стало предпосылкой для освоения суши растениями и животными, что, в свою очередь, усилило процессы почвообразования. Этому способствовало накопление органических веществ. Внесение в почву лигнина - химически устойчивого углевода, содержащегося в ксилеме сосудистых растений, замедляло процессы разложения почвенной органики. Почва аэрировалась ходами, которые проделывали в ней почвенные животные (черви и членистоногие).
Древнейшими растениями, имевшими в основном "наземный" облик, но населявшими, вероятно, пограничную зону между водой и сушей и влажные местообитания в низменных прибрежных районах, были псилофиты (Psilophyta, или Rhyniophyta) из группы риниевых (Rhyniales) - невысокие травянистые или кустар-никовообразные растения (Rhynia, Cooksonia), еще не имевшие настоящих корней и листьев; органы бесполого размножения (спорангии) располагались на концах дихотомически разветвленных побегов (рис. 46). В их древесине уже имелись примитивные сосудистые образования - трахеиды. Вероятно, в жизненном цикле этих растений происходила четкая смена поколений - диплоидного спорофита и более просто устроенного гаплоидного гаме-тофита.
Риниевые могли возникнуть от каких-то высших водорослей (может быть, близких к харовым) еще в ордовике. Псилофиты ныне рассматриваются как сборная группа, объединяющая древнейшие наземные растения - риниевые и возникшие от них зостерофиллофиты (Zosterophyllales) и тримерофиты (Trimero-phytales), обладавшие более развитым ветвлением с особыми спорангиеносными ветвями, мелкими листочками и некоторыми Другими прогрессивными особенностями. Псилофиты известны начиная со среднего силура и достигли расцвета в раннем и среднем девоне.
В верхнесилурийских отложениях встречаются также ископаемые остатки спор и гиф, которые интерпретируют как древнейшие известные остатки наземных грибов - аскомицетов. Вероятно, грибы принимали важное участие в процессе почвообразования.
К этому времени (силур - девон) относится и возникновение наиболее древних толщ горючих ископаемых, образующихся при неполном разложении скоплений растительных остатков.
Вероятно, в самом раннем девоне или даже в позднем силуре от самых примитивных риниевых могли возникнуть мохообразные (Bryophyta), а от высших псилофитов произошли различные группы сосудистых растений, приобревшие листья, корневую систему и более совершенные способы размножения: от зостеро-филлофитов - плаунообразные (Lycopsida), а от тримерофитов - членистостебельные, или хвощевые (Sphenopsida) и папоротникообразные (Pteropsida). Представители этих групп растений, лучше приспособленных к жизни на суше, в позднем девоне повсеместно вытеснили псилофитов и сформировали первую настоящую наземную флору, включавшую и древовидные формы. Эту флору называют "археоптерисовой" по широко представленным в ней видам папоротников Archaeopteris. К этому времени относится и появление первых голосеменных (Gymnospermae), называемых птеридоспермами или семенными папоротниками - Pteridospermae, возникших от древних разноспоровых папоротников и еще сохранявших внешний облик древовидных папоротникообразных.
В процессе приспособления к жизни на суше у растений совершенствовались вегетативные органы, в которых дифференцировались различные ткани: покровные (защищавшие растение от избыточного испарения и различных повреждений), механические (опорные, особенно важные в условиях возросшей в воздушной среде силы тяжести) и проводящие (обеспечивавшие транспорт веществ между различными органами). Особенно сложной для растений на суше стала проблема осуществления полового процесса: в воде подвижные мужские гаметы легко достигали яйцеклеток, тогда как в воздушной среде при неподвижности растений встреча гамет невозможна без специальных приспособлений, обеспечивающих их перенос от одного растения к другому. У высших наземных растений такие приспособления сформировались из гаплоидного поколения (гаметофита), которое редуцируется до одной - нескольких клеток (пыльца семенных растений). Диплоидное же поколение (спорофит) становится основным в жизненном цикле. Противоположное соотношение этих поколений в жизненном цикле - с преобладанием гаметофита - имеется лишь у мхов, что, вероятно, решающим образом ограничило адаптивные возможности этой группы в освоении суши: мхи выживают лишь в условиях достаточно высокой влажности и тесно связаны с субстратом.

Во влажном и теплом климате, характерном для первой поло вины каменноугольного периода, широкое распространение получила обильная наземная флора, имевшая характер густых влажных тропических лесов (рис. 47). Среди древовидных растении выделялись представители плаунообразных - лепидодендрон (Lepidodendraceae), достигавшие 40 м в высоту и до 2 м в диаметре у основания, и сигиллярии (Sigillariaceae) тридцатиметровой высоты. Стволы этих деревьев были покрыты характерным рельефным узором, образованным следами отпавших листьев (у взрослых деревьев листья сохранялись лишь в высокой кроне, где ствол дихотомически ветвился). Среди этих гигантов палеозойского леса высились и хвощеобразные пирамидальные каламиты (Calamitales) с членистым стеблем и мутовками мелких жестких листьев. Очень обильны были различные папоротники, как ползучие, так и древовидные, хотя и не достигавшие такой высоты, как плаунооб-разные, и составлявшие второй ярус лесной растительности и подлесок. Голосеменные были представлены разнообразными птеридоспермами, а также новой группой - кордаитами (Cordaitales). Кордаиты были высокими деревьями, стволы которых напоминали знакомые нам хвойные, но высоко расположенные кроны были образованы крупными, до 1 м в длину, ланцетообразными или лентовидными листьями. В древесине всех этих деревьев не обнаружено годичных колец, что говорит об отсутствии четко выраженной сезонности климата.
Наличие густых тропических лесов вдоль морских побережий, изрезанных лагунами, пересеченных дельтами рек и на больших территориях заболоченных, создавало предпосылки для интенсивного углеобразования. Мощные толщи каменных углей, сформировавшиеся в это время в разных районах земного шара, дали название каменноугольному периоду.
По мере заселения суши растениями возникли предпосылки для освоения наземной среды обитания животными. Вероятно, первыми на суше появились какие-то мелкие беспозвоночные, питавшиеся органическими остатками в почве. Возможно, такие животные появились уже к середине силурийского периода. Древнейшие известные наземные членистоногие - мелкие плоскоспинные многоножки (Myriapoda) со сдвоенными сегментами тела (как у современных двупарноногих - кивсяков) - были обнаружены в позднесилурийских и раннедевонских отложениях. М.С.Гиляров показал, что для многих групп беспозвоночных животных освоение суши началось с использования почвы как промежуточной среды обитания, в которой ряд факторов был в какой-то степени близок к показателям водной среды (высокая влажность, плотность субстрата). В почве можно использовать многие приспособления, сложившиеся в воде, при относительно незначительной их перестройке (аппарат передвижения, органы дыхания и др.). Вероятно, этим путем на сушу проникли из водоемов различные группы червей, которые в основном и остались почвенными обитателями, и членистоногих. Жизнь в почве и близких к ней местообитаниях характерна для наиболее примитивных современных групп членистоногих (онихофоры, многоножки, низшие насекомые - аптериготы и др.).
Приспособление животных к жизни на суше требовало, во-первых, перестройки локомоторного аппарата для передвижения в условиях возросшей по сравнению с водой силы тяжести; во-вторых, развития органов дыхания, способных усваивать кислород воздуха и избавлять организм от углекислого газа; в-третьих, формирования приспособлений, защищающих организм от обезвоживания. Последние включают развитие кожных покровов, снижающих потери воды с испарением (эпикутикула у членистоногих), а также совершенствование органов выделения, которые должны выводить из организма экскреты в малотоксичной и концентрированной (обезвоженной) форме. У водных животных чаще всего конечным продуктом азотистого обмена является аммиак, ядовитый и требующий для своего растворения значительного количества воды. У наземных форм выводятся из организма такие соединения, как мочевина, мочевая кислота, гуанин и др.
Замечательно, что очень сходные приспособления к наземной жизни формировались независимо и параллельно у представителей разных эволюционных стволов: в типе членистоногих, например, у нескольких самостоятельных линий среди хелицеровых (у скорпионов, пауков, сольпуг, клещей); во многом сходные адаптации развивались и у трахейных (у многоножек и насекомых). Отметим, что указанные выше приспособления растений к на-земности также формировались независимо и параллельно у пла-унообразных, членистостебельных, папоротникообразных. Параллельная эволюция вообще очень широко распространена в природе. Причины этой закономерности будут рассмотрены в последней части книги.
Из девона известны представители нескольких групп наземных членистоногих: палеозойская группа панцирных пауков (Soluta), клещи (Acaromorpha) и низшие первичнобескрылые группы насекомых (Insecta). Несомненно, разнообразие наземных беспозвоночных в девонском периоде было значительно большим, чем это представляется по дошедшим до нас скудным ископаемым остаткам.
Во второй половине раннекарбоновой эпохи появились наделенные крыльями высшие насекомые - птериготы. Вероятно, предки крылатых насекомых перешли к лазающей жизни на стволах и в Кронах деревьев. Для планирования при падениях и прыжках у Этих животных служили боковые уплощенные выступы тела на нудных сегментах. Из этих неподвижных придатков в процессе приспособительной эволюции естественный отбор сформировал удивительный в своем совершенстве летательный аппарат. Первоначально крылья имелись на всех 3 сегментах груди, хотя передняя пара еще сохраняла примитивное строение в виде неподвижных плоскостей, как это было, например, у палио-диктиоптеры (Palaeodictyoptera) (рис. 48). У большинства известных групп насекомых эта передняя пара крыльев утрачена. К концу карбона уже существовали разнообразные группы летающих насекомых, некоторые из них дожили до современности (поденки, стрекозы, тараканы, прямокрылые, равнокрылые, скорпионницы). В палеозое насекомые были единственными летающими животными. Некоторые виды стрекоз тогда достигали крупных размеров: у верхнекарбоновых - нижнепермских Meganeura длина тела достигала 30 см, размах крыльев до 65 см.

Разнообразие наземных беспозвоночных в карбоне было дополнено появлением растительноядных брюхоногих моллюсков (Gastropoda) из группы легочных (Pulmonata), дышащих воздухом. Вслед за беспозвоночными к освоению наземной среды обитания приступили и позвоночные. Их выход на сушу был возможен лишь после появления соответствующей пищевой базы (достаточно обильной фауны наземных беспозвоночных). Эти крупные животные не могли использовать почву как промежуточную среду между водой и сушей. Поэтому освоение суши для позвоночных было возможно только в условиях влажного и теплого климата, наиболее благоприятного для животных, физиология которых еще во многом подобна физиологии рыб. С другой стороны, комплекс преадаптаций для перехода к наземной жизни должен был сформироваться у предков наземных позвоночных еще в водной среде.
Предками наземных позвоночных (тетрапод) стали кистеперые рыбы, обладавшие легкими и снабженными мощной мускулатурой парными конечностями - саркоптеригиями, которые, вероятно, позволяли этим рыбам не только передвигаться по дну водоемов среди густых зарослей, но и выбираться на мелководье, проползать по мелким протокам из одного водоема в другой. Внутренний скелет этих конечностей был образован одной крупной костью, сочлененной с поясом конечностей (плечевым или тазовым), далее следовал второй отдел из двух параллельных костей, и дистально располагались многочисленные мелкие косточки (рис. 45, в). В этой схеме легко просматривается прототип скелета конечностей наземных позвоночных.
Вероятно, некоторые виды кистеперых рыб перешли к освоению новой кормовой базы - обильной фауны беспозвоночных, обитавших на мелководье, среди прибрежных растений и по берегам водоемов. Это направление приспособительной эволюции вело дальше к освоению суши, с ее бурно развивавшейся флорой и фауной, в составе которой еще не было крупных хищников. Эту экологическую нишу и заняли наземные позвоночные.
Отметим, что приспособления к наземности независимо и параллельно развивались в нескольких линиях эволюции кистеперых рыб. В связи с этим Э.Ярвик выдвинул гипотезу о дифилетическом происхождении наземных позвоночных от двух разных групп кистеперых рыб (Osteolepiformes и Porolepiformes). Однако целый ряд ученых (А.Ромер, И.И.Шмальгаузен, К.Томсон, Э.И.Воробьева) подвергли аргументы Ярвика критике. Ныне большинство исследователей считают более вероятным монофилетическое происхождение тетрапод от остеолепиформных кистеперых, хотя при этом допускается возможность парафилии, т.е. достижения уровня организации земноводных несколькими близкородственными филетическими линиями остеолепиформных рыб, эволюционировавшими параллельно.

Наиболее древние представители земноводных (класс Amphibia) ихтиостеги (Ichthyostegalia, рис. 49) известны из верхнедевонских отложений Гренландии. Ихтиостеги были довольно крупными животными (свыше 80 см длиной), имевшими уже хорошо Развитые пятипалые конечности наземного типа, с помощью которых они, несомненно, могли переползать по суше. Однако значительную часть жизни ихтиостеги, вероятно, проводили в водоемах, будучи, по удачному выражению Ярвика, "четвероногими рыбами". Об этом говорит наличие у них настоящего хвостового плавника, напоминавшего хвост кистеперых рыб, а также сохранение во взрослом состоянии сейсмосенсорных рецепторов ("боковой линии" - органов чувств, которые функционируют только в водной среде, воспринимая инфразвуковые колебания и электромагнитные поля). В черепе ихтиостег сохранялись рудименты костей жаберной крышки (возможно, имелись и жабры, укрепленные на хрящевых жаберных дугах). Тело было покрыто мелкими костными чешуйками.
Вероятно, древнейшие земноводные, подобные ихтиостегам, в водной среде проигрывали конкуренцию многочисленным и разнообразным кистеперым рыбам, поскольку парные конечности наземного типа не могли быть эффективными рулями глубины при плавании. Преимущества пятипалых конечностей проявлялись лишь в наиболее мелких участках водоемов и на суше. Скорее всего, между кистеперыми рыбами и их потомками - земноводными-в девоне существовали конкурентные отношения, способствовавшие расхождению этих групп в разные адаптивные зоны: кистеперые рыбы доминировали в водоемах, а земноводные заняли промежуточные местообитания между водой и сушей (мелкие прибрежные участки водоемов, где свободное плавание затруднено; заболоченные районы; области с избыточной влажностью на суше). Попутно отметим, что этот тип местообитаний, промежуточных и пограничных между водой и сушей, остался и доныне наиболее благоприятной для земноводных адаптивной зоной, где амфибии сохранили преимущества и перед рыбами, и перед более совершенными наземными группами тетрапод. В позднем девоне уже началась дивергентная эволюция древнейших земноводных. Помимо ихтиостег в конце девона существовали представители и ряда других групп амфибий, ископаемые остатки которых были обнаружены в различных районах. Любопытно, что число пальцев у этих животных часто достигало 7-8.
В карбоне начинается расцвет древних амфибий, представленных в верхнем палеозое большим разнообразием форм (рис. 50), которых условно объединяют под названием стегоцефалов, т.е. покрытоголовых. Этот термин, не имеющий ныне таксономического значения, подчеркивает характерную особенность этих животных, голова которых была защищена унаследованным от ры сплошным костным панцирем (так называемый стегальный череп), пронизанным лишь отверстиями ноздрей, глазниц и "теменного глаза".

Наиболее известной из групп стегоцефалов являются лабиринтодонты (Labyrinthodontia), к числу которых принадлежат и ихтиостеги. Их название связано с любопытной особенностью строения зубов, также унаследованной от кистеперых рыб: эмаль и дентин образуют многочисленные складки, так что на поперечном шлифе возникает картина, напоминающая лабиринт.
Лабиринтодонты в верхнем палеозое были одной из наиболее Распространенных и обильных видами групп позвоночных. К ним принадлежали и мелкие, и крупные, более 1,5 м, формы. В карбоне преобладали виды со слабыми конечностями и длинным телом, которые, вероятно, обитали в многочисленных болотах. В пермское время появились крупные крокодилообразные стегоцефалы с большой уплощенной головой (подобные Eryops, см. рис.50, б), а также мелкие виды с лучше развитыми конечностями и укороченным туловищем и хвостом (Cacops, рис. 50, в). Эти последние жили, скорее всего, преимущественно на суше, питаясь различными беспозвоночными, хотя, как и все земноводные, нуждались в повышенной влажности воздуха и близости водоемов для 0тКдадки икры. Как считает ряд ученых, мелкие лабиринтодонты, державшиеся по берегам водоемов и спасавшиеся от наземных врагов прыжками в воду, стали предками бесхвостых земноводных (Anura), возникших, вероятно, в позднепермское время. Древнейший известный представитель бесхвостых амфибий Triadoba-trachus из раннего триаса Мадагаскара уже имел характерные пропорции тела и основные структурные особенности лягушек, хотя еще сохранял небольшой хвост.
Два других отряда современных амфибий в ископаемом состоянии известны из значительно более поздних отложений: хвостатые (Urodela) из средней юры, безногие, или червяги (Apoda). из ранней юры. Проблема их происхождения остается дискуссионной. По многим признакам скелета и мускулатуры головы хвостатые амфибии близки к бесхвостым и, возможно, имеют с ними общее происхождение.
Безногие амфибии стоят несколько особняком, сохраняя во многих отношениях наиболее примитивное строение среди современных земноводных. Некоторые палеонтологи связывают их происхождение с филогенетическим стволом стегоцефалов, объединяемых в подкласс лепоспондильных (Lepospondyli). (Согласно другой точке зрения, все три отряда современных амфибий имели общее происхождение, вероятно от лабиринтодонтов).
Среди лепоспондильных было много форм с сильно удлиненным телом и слабо развитыми конечностями. Таковы, например, лизорофы (Lysorophia). Представители другого отряда лепоспондильных - аистоподы (Aistopoda), как и червяги, совсем утратили конечности. Однако они не могли быть предками червяг, так как обладали рядом специфических особенностей строения. К лепоспондильным принадлежат также нектридии (Nectridia). Одно из их семейств отличалось причудливым строением черепа, задние углы которого непомерно разрастались вбок и назад в виде длинных конических выростов, так что голова в целом сверху выглядела подобной бумерангу. Предполагают, что эти выросты могли играть роль своего рода подводных крыльев, создававших при движении некоторую подъемную силу, способствуя планированию в воде.
От примитивных лабиринтодонтов уже в девоне обособилась группа антракозавров (Anthracosauria), древнейший представитель которых Tulerpeton был обнаружен в верхнедевонских отложениях Тульской области. Первоначально эти стегоцефалы также обитали в прибрежной зоне водоемов. Однако среди антракозавров очень рано наметилась тенденция к освоению наземных местообитаний. Как и в большинстве случаев, эта тенденция независимо и параллельно развивалась в нескольких разных филетических линиях, в каждой из которых постепенно формировались во многом сходные приспособления к жизни на суше. Антракозавров и тех из их потомков, которые сохранили, несмотря на наземный образ жизни, общий уровень организации и характерные черты размножения и онтогенеза земноводных (с откладкой икры в водоемах и водными личинками, имеющими жабры), объединяют в подкласс батрахозавров, или "ящероземноводных" (Batrachosauria). К батрахозаврам принадлежат сеймурии (Seymouria; рис. 51), кот-лассии (Kotlassia) и родственные им формы, существовавшие в пермском периоде. Сеймурии долгое время считали не земноводными, а пресмыкающимися, так близки они по строению черепа, позвоночника и конечностей к настоящим рептилиям. Однако позднее были обнаружены остатки личиночных стадий этих животных, имевших следы каналов сейсмосенсорных органов на костях черепа и, следовательно, живших в воде.
От каких-то примитивных батрахозавров в раннем карбоне возникли настоящие рептилии (Reptilia), к которым, в частности, относят небольшую форму Westlothiana, обнаруженную в нижнекарбоновых отложениях Шотландии. В ее организации сочетаются прогрессивные (рептильные) особенности черепа и примитивное состояние ряда признаков позвоночника и конечностей ("мозаичность"). Р.Кэррол считает, что непосредственные предки пресмыкающихся могли быть близки по основным особенностям к позднекарбоновым гефиростегам (Gcphyrostegidae, рис. 51), которые, однако существовали слишком поздно, чтобы быть действительными предками рептилий. Возможно, гефиростеги были реликтовой группой, сохранившей ряд архаических особенностей строения, характерных для раннекарбоновых предков пресмыкающихся.
Несомненные рептилии известны начиная со среднего карбона. Е.Олсон указал на близкое совпадение времени появления пресмыкающихся и разнообразных групп высших насекомых-птеригот. Отметив, что все наиболее древние рептилии были относительно мелкими насекомоядными животными, Олсон пришел к выводу, что формирование пресмыкающихся как группы было тесно связано с использованием новой богатой пищевой базы, возникшей на суше с появлением насекомых.
Вероятно, уже в карбоне у пресмыкающихся сформировался целый комплекс приспособлений, позволивших им и их потомкам стать в полной мере наземными животными. Важнейшим из этих приспособлений, которое сделало возможными многие другие существенные изменения организации пресмыкающихся, было развитие эффективного механизма вентиляции легких посредством движений грудной клетки. Он сделал ненужным кожное дыхание, которое у земноводных является необходимым дополнением легочного, так как из-за несовершенства вентиляции легких, где скапливается избыток углекислого газа, земноводные нуждаются в выведении последнего через кожу. Поэтому в коже рептилий стало возможным усиление процессов ороговения эпидермиса, защитившего организм от постоянной потери влаги через покровы (неизбежного при кожном дыхании у земноводных) и риска гибели в сухом воздухе от обезвоживания. После этого пресмыкающиеся получили возможность заселить практически любые местообитания на суше, вплоть до безводных пустынь. Этому способствовало также развитие метанефрической (тазовой) почки и механизмов обратного всасывания воды из мочи, позволявших выводить из организма максимально концентрированные экскреты. После ликвидации кожного дыхания у рептилий стало возможным разделение в сердце артериальной (поступающей из легких) и венозной (идущей от всех остальных органов) крови, что также невозможно для земноводных, так как у них от кожи кровь поступает в правое предсердие, а от легких - в левое, и разделение этих потоков крови в желудочке сердца затруднило бы использование кожного дыхания. Все эти важнейшие достижения рептилий наряду с целым рядом усовершенствований скелета (увеличение подвижности головы с развитием особой формы передних шейных позвонков, некоторые преобразования в скелете конечностей и др.) создали важнейшие предпосылки для полного овладения наземной средой обитания.
Одновременно у древнейших рептилий сформировался второй комплекс приспособлений, избавивших от связи с водоемами все процессы, связанные с размножением. Это было появление внутреннего осеменения вместо характерного для земноводных наружного; формирование крупных яиц, обильно снабженных запасами питательных веществ (желтком) и способных обеспечить длительное развитие зародыша; образование прочных яйцевых оболочек, защищающих яйцо на суше от обезвоживания и механических повреждений; и наконец, возникновение особых зародышевых оболочек - амниона, серозы и аллантоиса, обеспечивающих благоприятную среду для развития зародыша, газообмен и накопление экскретов (из-за невозможности их выведения из яйца во внешнюю среду). По признаку наличия этих зародышевых оболочек высшие классы наземных позвоночных объединяются в группу амниот, противопоставляемую низшим позвоночным - анамниям.
Древнейшие и наиболее примитивные пресмыкающиеся принадлежали к подклассу котилозавров (Cotylosauria). Название "котилозавр" подчеркивает характерную особенность черепа этих животных, защищающего голову сплошным костным панцирем, как у стегоцефалов, но более высокого и узкого и несколько напоминающего у некоторых котилозавров при взгляде сверху чашу или кубок, откуда и название (греч. kotylos - чаша, кубок). Центральной группой котилозавров являются капториноморфы (Captorhinomorpha, рис. 52), древнейшие представители которых известны из среднекарбоновых отложений. Это были небольшие, до 50 см длиной, насекомоядные животные.


Уже во второй половине каменноугольного периода возникло большое разнообразие форм пресмыкающихся, среди которых появились помимо насекомоядных растительноядные животные (диадектиды) и крупные хищники, питавшиеся позвоночными (некоторые более поздние капториноморфы и другие группы). Удивительно, что некоторые филетические линии рептилий уже в карбоне вернулись в водоемы, став полуводными или даже полностью водными животными. Особенно интересны мезозавры (Mesosauria, рис. 53), конечности которых преобразовались в ласты, а длинные узкие челюсти (удобные для схватывания добычи в воде) были усажены многочисленными тонкими и острыми зубами. Остатки мезозавров были найдены в Южной Африке и Бразилии. Это хорошо соответствует представлениям о существовании в верхнем палеозое единого материка Гондваны.
Жизнь в позднем палеозое
Как мы уже упоминали, начиная с верхнего карбона в Южном полушарии усилились процессы оледенения. На свободной от ледников территории Гондваны установился умеренный прохладный климат с выраженной сезонностью. Об этом говорит, в частности, наличие годичных колец в древесине гондванской флоры этого времени, получившей название глоссоптериевой по широко распространенным в ней видам семенного папоротника Glossopte-ris - небольшим растениям с крупными цельными листьями.
В состав глоссоптериевой флоры кроме различных семенных папоротников (птеридоспермов) входили также представители других групп голосеменных растений: кордаитов, гинкговых и хвойных. Глоссоптериевая флора была характерна для обширных территорий, соответствующих современным Индии, Афганистану, Южной Африке, Южной Америке, Австралии, Новой Зеландии и Антарктиде.
На северных континентах, входивших в состав Лавразии и располагавшихся в раннепермское время в значительной степени в экваториальном поясе, сохранялась растительность, близкая к тропической флоре карбона, но уже с меньшим богатством видов лепидодендронов и сигиллярий. К середине пермского периода во многих районах Лавразии климат становится все более засушливым, что приводит к изменению растительности: исчезают папоротники, каламиты, древовидные плаунообразные и другие влаголюбивые растения тропических лесов. Лишь на востоке Лавразии (современные Китай и Корея) климат и флора оставались в известной мере близкими к таковым в каменноугольное время.
Животный мир также претерпел значительные изменения на протяжении пермского периода, ставшие особенно драматичными во второй его половине. В наибольшей степени это коснулось морских животных, среди которых многие группы подверглись значительному угнетению, сопровождавшемуся уменьшением численности и разнообразия видов, вплоть до полного вымирания целых классов. Примерно в середине пермского периода вымирают последние трилобиты, эвриптериды, бластоидеи (один из классов прикрепленных иглокожих), палеозойские группы морских лилий. К концу перми исчезают тетракораллы, значительно сокращается число видов фораминифер среди простейших, морских ежей и офиур среди иглокожих, аммоноидей и наутилоидей среди моллюсков, остракод среди ракообразных, губок, мшанок и плеченогих. Из позвоночных вымирают акантондии, многие палеозойские группы хрящевых рыб. В пресных внутриконтинентальных водоемах значительно снижается численность хоановых рыб. К концу палеозоя вымирают лепоспондильные стегоцефалы. Изменения в наземной фауне были не столь значительны.
Пермское вымирание по своим масштабам принадлежит к категории так называемых "великих вымираний" и сравнимо, например, с широко известным великим вымиранием в конце мелового периода. Причины массовых вымираний, приводивших к радикальным изменениям общего облика фауны и флоры, во многом остаются загадочными. Вымирание вообще сопровождает эволюцию организмов как ее неизбежный побочный результат: в борьбе за существование выживают лишь те виды, которые оказались наиболее приспособленными при данных условиях. Вымирание отдельных видов происходило на любом отрезке эволюционной истории мира организмов либо в результате конкуренции с биологически близкими видами, либо по причине неспособности данного вида выдержать те или иные изменения абиотических факторов внешней среды. В обоих случаях возможности модифи-кационной изменчивости вымирающего вида и резерв его комби-нативной изменчивости оказываются недостаточными, чтобы обеспечить его выживание в изменяющейся внешней среде.
В принципе те же общие причины определяют фатальную судьбу для каждого вымирающего вида и в эпохи массовых вымираний, но тогда одновременно обрывается множество филетических линий. Это может происходить и достаточно постепенно (численность вымирающих видов падает от поколения к поколению на протяжении больших промежутков времени), но в огромной исторической перспективе геологического времени эти процессы нередко выглядят как относительно быстрые и резкие изменения фауны и флоры.
Очевидно, массовые вымирания обусловлены какими-то причинами, имевшими общее значение для эволюционной судьбы сразу многих филетических линий, принадлежащих к разным группам организмов. Конечно, при этом совсем не обязательно, чтобы изменения какого-то одного или нескольких важных факторов прямо влияли на все подвергающиеся вымиранию виды организмов. Нарушения структуры биоценозов, выходящие за пределы возможностей естественной саморегуляции этих биологических макросистем, должны прямо или косвенно сказываться на многих видах организмов. Вероятно, существенной чертой эпох великих вымираний было именно разрушение многих биоценозов (биоценотические кризисы, по определению Б.Б. Родендорфа и В.В.Жерихина). Биоценотические кризисы могут быть вызваны самыми различными причинами, как абиотическими - изменения климата, рельефа и т.п., так и биотическими - появление и массовое расселение новых групп организмов, причем часто имеет место целый комплекс взаимодействующих факторов, что очень затрудняет анализ конкретных случаев, особенно в далеком прошлом.
О причинах пермского вымирания было выдвинуто несколько гипотез. Большинство из них в конечном итоге опирается на теорию дрейфа континентов. Согласно гипотезе, предложенной Д.Валентайном и Э.Мурсом, образование Пангеи обусловило значительное сокращение общей площади мелководных бассейнов в области континентального шельфа, которая является наиболее благоприятной средой для жизни многих групп морских организмов. Это должно было привести к существенному нарушению многих сложившихся в середине палеозоя морских биоценозов и, как следствие, к вымиранию многих видов.
Другая гипотеза, выдвинутая Р.Боуэном, обращает внимание на потепление климата и завершение великого оледенения Гондваны во второй половине пермского периода, что могло быть связано с относительно небольшим смещением входивших в ее состав континентальных плит, в результате которого Южный полюс оказался в океане. Потепление климата привело к бурному таянию ледников и поступлению в океан огромного объема талых ввод, которое должно было вызвать понижение солености морской воды. Пермское вымирание отразилось в первую очередь именно на тех группах морских организмов, которые наиболее чувствительны к изменениям солености: кораллы, морские лилии, мшанки, брахиоподы, головоногие моллюски и др. Новая стабилизация температурного и солевого режима океана произошла уже в триасовом периоде, и разнообразие форм морских беспозвоночных к концу триаса вновь стало возрастать.
Для наземных животных изменения физических условий в позднем палеозое не имели столь катастрофических последствий, как для представителей морской фауны. Пермский период стал временем расцвета древних пресмыкающихся, быстро осваивавших все новые местообитания, вытесняя хуже приспособленных конкурентов из числа стегоцефалов и батрахозавров, с запозданием пытавшихся закрепиться на суше.
Котилозавры разделились на несколько основных эволюционных стволов, первоначальная дифференциация которых была связана, вероятно, с освоением разных способов питания и различных местообитаний. Самыми крупными котилозаврами были растительноядные парейазавры (рис. 54), известные из средне- и верхнепермских отложений Европы и Африки. Эти животные, Достигавшие 3 м длины, имели широкое массивное тело и крупный тяжелый череп с поразительно маленькой полостью, вмещавшей головной мозг. Наружная поверхность толстых черепных Костей была покрыта своеобразным бугорчатым рельефом из приросших к ним остеодерм (вторичных кожных окостенений, часто встречающихся у рептилий), по бокам черепа выступали толстые костные пластины-"щеки" (отсюда название этих животных: Pareiasauria - "щекастые ящеры"). Парейазавры были, вероятно, неповоротливы и медлительны, от нападений хищников их защищал панцирь из толстых костных бляшек-остеодерм, рядами сидевших в коже. Судя по строению зубов (довольно слабых, с плоской листовидной коронкой), пищей парейазавров служили какие-то сочные и мягкие растения. Возможно, парейазавры обитали по берегам водоемов и болот, питаясь водной растительностью, и были палеозойскими аналогами современных бегемотов.


Самые крупные хищники пермского времени принадлежали к подклассу зверообразных, или синапсидных рептилий (Theromorpha, или Synapsida). У этих пресмыкающихся в височной области черепа для его облегчения и предоставления необходимого пространства для сокращения мощных челюстных мышц возникла одна пара широких отверстий, или височных окон, пронизавших наружный черепной панцирь и ограниченных по бокам скуловыми дугами (рис. 55). Такой тип черепа, называемый синап-сидным, дал одно из названий подкласса.
Самыми древними зверообразными рептилиями были пеликозавры (Pelycosauria), появившиеся уже в позднем карбоне. Их расцвет приходится на раннепермское время, когда многие их представители достигали крупных размеров, до 2 м. Некоторые пеликозавры (диметродон - Dimetrodon, эдафозавр - Edaphosaurus и др.) имели странное приспособление: гипертрофированные остистые отростки спинных позвонков поднимались над позвоночником почти на 1 м, а у эдафозавра (рис. 56) на этих отростках имелись еще и короткие боковые выросты. Вероятно, между ними была натянута складка кожи, образуя огромный продольный гребень ("спинной парус"). Может быть, эта складка играла роль в терморегуляции, увеличивая поверхность тела, что могло в одних ситуациях усиливать теплоотдачу, в других же - содействовать повышению температура тела при обогревании в солнечных лучах (повышение температуры тела путем инсоляции, называемое гелиотермией, присуще всем современным рептилиям).

Большинство пеликозавров вымерло уже в середине пермского периода, вероятно не выдержав конкуренции с представителями возникшей от пеликозавров более прогрессивной группы зверообразных рептилий - терапсид (Therapsida). В позднепермской эпохе терапсиды стали доминирующей группой пресмыкающихся. Этому способствовал целый ряд присущих им прогрессивных особенностей, в частности усовершенствование наземного передвижения. У большинства рептилий сохранилось примитивное положение конечностей, при котором плечо и бедро располагаются в целом горизонтально, локоть и колено направлены вбок, и тело животного удерживается на весу между конечностями усилиями мышц (отсюда характерное для рептилий "пресмыкание", когда брюхо при движении лишь незначительно приподнимается над землей). У высших терапсид локтевой сустав повернулся назад, коленный - вперед, бедро и плечо (и конечности в целом) оказались расположенными под телом, высоко приподнятым над субстратом; для поддержки тела при этом не требуется столь значительной работы мышц, как у типичных пресмыкающихся.
У многих терапсид произошла дифференциация зубной системы на передние острые "резцы", крупные кинжалообразные "клыки" и заклыковые щечные зубы, которые у высших представителей этой группы (териодонтов, или "зверозубых", - Theriodontia) приобрели усложненную коронку с тремя бугорками, что позволяло более эффективно пережевывать пищу. Пережевывание пиши в ротовой полости требовало задержки дыхания, в связи с чем у терапсид развивается вторичное нёбо, разделившее примитивную ротовую полость на два этажа: верхний (носовой ход) и нижний (собственно ротовая полость).

Терапсиды были очень разнообразны: среди них были и хищники различных размеров, вплоть до трехметровых иностранцевий (Inostrancevia, рис. 57) с клыками более 10 см длиной (своего рода "саблезубых тигров" пермского периода), и растительноядные животные, иногда имевшие причудливое строение. Для некоторых растительноядных дейноцефалов (Deinocephalia, их название буквально означает "страшноголовые") характерны большие костные бугры или шишки на темени и щечной области черепа, придававшие голове этих животных устрашающий вид (скорее всего, это были защитные приспособления). Были и хищные дейноцефалы, например приземистый и длинный титанофонеус - Titanophoneus, большие лапы которого, по мнению Ю.А.Орлова, были снабжены плавательными перепонками, а сам этот хищник вел полуводный образ жизни.
В позднепермское время были широко распространены дици-нодонты (Dicynodontia), утратившие все зубы, кроме пары огромных верхних клыков, сохранявшихся у некоторых видов лишь у самцов. Беззубые челюсти были покрыты роговым клювом. Нелегко представить себе способ питания этих странных животных, среди которых были и сравнительно мелкие, и очень крупные, размеров носорога, виды. Предполагали, что дицинодонты питались падалью или поедали питательную сердцевину стволов саговниковых растений, раскалывая или разрывая их своими клыками, или выкапывали корневища хвощеобразных растений, в изобилии произраставших по берегам водоемов.
Местонахождения с ископаемыми остатками целых комплексов видов позднепермских рептилий (котилозавров, дицинодонтов, дейноцефалов, териодонтов) с довольно сходным общим обликом фауны известны в Южной Африке и Восточной Европе. Замечательные местонахождения позднепермских тетрапод были обнаружены на территории России. На берегу Северной Двины, в районе г.Котласа, В.П.Амалицким в 1897г. были найдены остатки крупных парейазавров, дицинодонтов, хищных териодонтов (среди них были огромные иностранцевии), батрахозавров и лабиринтодонтов. Еще одна интересная позднепермская фауна была раскопана в 1957-1960 гг. в Очерском районе Пермской области. В состав Очерской фауны, исследованной П.К.Чудиновым, входили хищные и травоядные дейноцефалы (среди последних огромные рогатые эстемменозухи, Estemmenosuchus), хищные териодонты, терапсиды, близкие к предкам дицинодонтов, лабиринтодонты. Костеносные породы этих местонахождений сформировались из глинистых и песчаных осадков речных дельт1.
В некоторых филогенетических стволах рептилий продолжалось совершенствование приспособлений к насекомоядности, которая была, вероятно, самой примитивной формой питания пресмыкающихся. Это направление эволюционных преобразований привело к появлению небольших и проворных ящерицеобразных животных, причем сходный тип строения сформировался параллельно в нескольких независимых филетических линиях. Одна из них сохранила общий уровень организации котилозавров, со сплошным покровом накладных костей в височной области черепа. Это были проколофоны (Procolophonia). Остатки примитивных проколо-фонов, никтифрурета (Nyctiphruretus, рис.58) и никтеролетера (Nycteroleter) - легких большеглазых животных с удлиненным телом и хвостом общей длиной до 30-40 см были найдены в верхнепермских отложениях на берегах р. Мезени.

В других группах насекомоядных рептилий произошли некоторые прогрессивные изменения организации, в частности усовершенствование строения черепа. Эти животные, давшие начало двум центральным подклассам рептилий: лепидозаврам (Lepidosauria) и архозаврам (Archosauria) - приобрели удлиненные челюсти, удобные для схватывания мелкой подвижной добычи, и две пары височных окон, расположенных друг над другом и разделенных тонким костным мостиком - верхней височной дугой. Так возник диапсидный тип черепа (см. рис. 55, в), легкая и ажурная конструкция которого позволила усовершенствовать так называемый кинетизм - подвижность верхней челюсти и некоторых связанных с ней костей, унаследованную наземными позвоночными от кистеперых рыб (рис. 59).

Примитивные лепидозавры (миллерозавры - Millerosauria), у которых лишь началось развитие височных окон и усиление кинетизма черепа, известны из верхнепермских отложений Южной Африки. В породах примерно того же возраста сохранились остатки первых рептилий, имевших настоящий диапсидный череп - эозухий (Eosuchia), наиболее известным представителем которых была янгина (Youngina). Отдаленным потомком эозухий является дожившая до нашего времени новозеландская гаттерия (Sphenodon), которая принадлежит к отряду клювоголовых (Rhynchocephalia), появившихся в раннем триасе. От эозухий или от миллерозавров возникли настоящие ящерицы (Lacertilia), у которых в связи с усилением подвижности верхней челюсти (см. рис. 59) редуцировалась нижняя височная дуга и череп приобрел легкую ажурную конструкцию. Ящерицы достоверно известны начиная со среднего триаса.
Вероятно, эозухий были также предками высших рептилий, принадлежащих к подклассу архозавров, которые оставили самые яркие страницы в палеонтологической истории мезозоя. Д.Уотсон высказал предположение, что обособление предков архозавров от ранних насекомоядных лепидозавров было связано с переходом первых к питанию позвоночными. Этим объясняется более Массивная конструкция черепа архозавров и развитие у них более крупных зубов, сидящих в отдельных ячейках (текодонтность). Для архозавров характерно также более выпрямленное положение конечностей, чем у лепидозавров.
Наиболее ранним известным архозавром является позднепермский проторозавр (Protorosaurus), пропорциями тела и размерами (1-2 м) напоминавший современных ящериц-варанов.
В то же время происходили существенные изменения рельефа Земли и положения континентов, которые привели к формированию в общих чертах существующих ныне континентов и океанов. В начале триаса сохранялись относительно высокие материки. Затем началась морская трансгрессия с образованием мелких краевых морей Тихого океана, прерывавшаяся в начале и в конце юрского периода Древне- и Позднекиммерийскими фазами горообразования. В меловом периоде новая морская трансгрессия привела к затоплению значительных территорий Европы и Северной Африки краевыми теплыми морями древнего океана Тетис.
По представлениям теории тектоники плит, общей тенденцией в мезозое был распад суперконтинента Пангеи и расхождения континентальных плит (рис. 60). В триасе этот процесс только начался. Лавразия сместилась несколько к северу. При этом происходило раскрытие океана Тетис, отделявшего Лавразию от Гондваны. Затем Лавразия раскололась на Северную Америку и Евразию, между которыми, начиная с раннеюрского времени, началось формирование северной части Атлантического океана. Вероятно, уже к концу юрского периода начался раскол Гондваны и, соответственно, возникновение Индийского океана и южной части Атлантики, отделивших Африку от Австралии и Антарктиды на востоке и юге и от Южной Америки на западе. Соединение северной и южной частей Атлантики произошло уже в позднемеловое время.
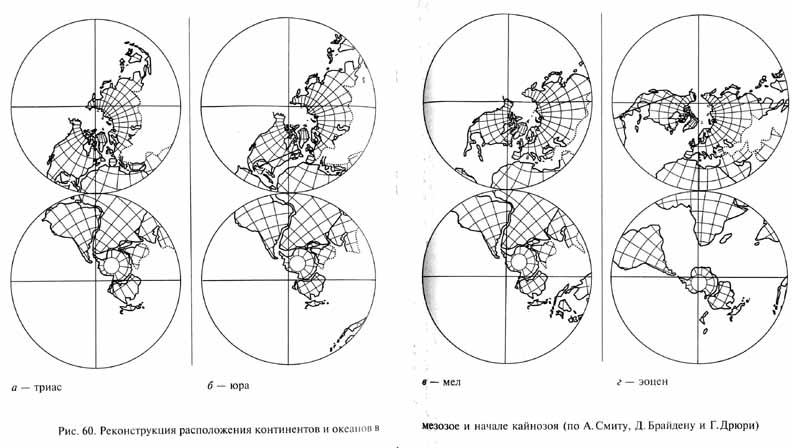
В меловом периоде расхождение континентов, которые в конце палеозоя входили в состав единой Пангеи, значительно прогрессировало. Продолжали расширяться Атлантический и Индийский океаны. При движении американских плит к западу на их западных окраинах возникли мощные зоны горной складчатости: в юрском периоде - Сьерра-Невада, в меловом - Анды и Скалистые горы (Позднекиммерийская и Ларамийская фазы горообразования). Индостан и Мадагаскар обособились от Африки, Индостанская плита начала смещение к северо-востоку. Сложные взаимные смещения Африки и Европы привели к постепенному закрытию океана Тетис. Из материков, входивших в состав Гондваны, в конце мезозоя остались соединенными лишь Антарктида и Австралия. Зато материки, составлявшие Лавразию и разделившиеся в начале мезозоя, к концу мела вновь соединились, правда противоположными концами: Азия и Северная Америка сошлись в области Чукотки и Аляски. Так возникла Берингия, сухопутный мост, связавший Америку и Европу на месте современного Берингова пролива.
Обновление флоры и фауны в триасе
Пермское оледенение и последовавшее после его завершения глобальное потепление климата привели к значительному изменению общего облика флоры. Вымерли древовидные плаунооб-разные, столь характерные для лесов второй половины палеозоя. Исчезли также семенные папоротники (птеридоспермы) и кордаиты. Их место занимают разнообразные представители других групп голосеменных растений: саговниковых (Cycadophyta), гинк-говых (Ginkgoales) и хвойных (Coniferales) (см. рис. 68, изображающий ландшафт юрского периода, где представлены указанные голосеменные, появившиеся уже в триасе). Особенно характерны для наземной флоры первой половины мезозоя были саговниковые: древовидные пальмообразные, с высокими колоннообразными стволами, увенчанными пучком перистых листьев (Williamsonia), или приземистые, с бочкообразным или клубневидным коротким и толстым стволом (Cycadeoidea). Саговниковые, хвойные и гинкговые формировали общий облик лесов в первой половине мезозойской эры.

Рубеж между пермским и триасовым периодами был отмечен также существенными изменениями в фауне наземных позвоночных. В пермское время преобладающими группами пресмыкающихся были зверообразные рептилии и котилозавры, тогда как группы с диапсидным черепом (лепидозавры и архозавры), объединяемые под названием "завропсиды", оставались сравнительно Немногочисленными. Это соотношение резко изменилось к началу мезозоя. Из числа котилозавров в триас перешли лишь проколофоны (вымирающие к концу этого периода), а из зверообразных - немногие группы дицинодонтов и высших териодонтов. Зато обилие и разнообразие диапсидных рептилий неуклонно возрас-Тает, и во второй половине триаса они становятся доминирую-Щими. При этом нужно отметить, что высшие териодонты обладали рядом прогрессивных признаков, отсутствовавших у завропсид (например, были способны к пережевыванию пищи, значительно повышающему степень ее усвоения организмом; вероятно, они имели теплоизолирующий волосяной покров и т.д.). Победа в борьбе за существование была достигнута завропсидами, так сказать, вопреки прогрессивным чертам организации териодонтов и, очевидно, основывалась на каких-то важных преимуществах первых.
П.Робинсон предложила гипотезу, связывающую изменения фауны рептилий в триасе с особенностями физиологии завропсид и зверообразных пресмыкающихся, о которых с известной долей вероятности можно судить по соответствующим характеристикам современных потомков тех и других. Современные завропсиды (в широком смысле этот термин используется для объединения всех современных групп этого класса и птиц) характеризуются наличием целого комплекса приспособлений к жизни в условиях жаркого засушливого климата. Завропсиды могут выдержать значительно более высокие температуры тела, чем потомки зверообразных рептилий - млекопитающие (многие ящерицы - до 44 °С, птицы - до 43 °С, млекопитающие - лишь до 39 °С; для Млекопитающих смертельно повышение температуры тела до 43 °С, Для птиц - до 46-47 °С). При этом рептилии используют энергию солнечного облучения для повышения температуры своего тела До оптимального уровня (гелиотермия); в связи с этим для них сохраняет важное значение "теменной глаз", регулирующий различные функции организма в зависимости от интенсивности падающего света.
Конечным продуктом белкового обмена у завропсид является мочевая кислота, тогда как у млекопитающих - мочевина. Мочевая кислота может образовывать перенасыщенные растворы, поэтому для ее выведения из организма требуется примерно в 10 раз меньше воды, чем для выведения мочевины. Очевидно, у завропсид выделительная система гораздо лучше экономит воду для организма, чем у млекопитающих.
Логично предположить, что организация завропсид складывалась под контролем отбора, приспосабливавшего этих рептилий к условиям засушливого и жаркого климата, тогда как зверообразные рептилии сформировались в местообитаниях с более влажным и прохладным климатом. Такие местообитания, вероятно, были широко распространены на континентах, входивших в состав Гондваны, в эпоху великого палеозойского оледенения. К концу пермского периода и в триасе во многих регионах климат становится все более засушливым (или, по крайней мере, характеризовался значительными сезонными засухами). П.Робинсон связывает это с изменениями морских течений и преобладающих направлений циркуляции атмосферных потоков в результате дрейфа континентов, входивших в состав Лавразии и Гондваны. Засушливость климата должна была дать важные преимущества в борьбе за существование тем группам наземных животных, которые обладали более высокой устойчивостью к действию высоких температур, были способны лучше экономить воду и с помощью специальных форм поведения могли использовать энергию солнечного облучения для достижения оптимальной температуры своего тела. Поскольку по всем этим показателям завропсиды превосходили зверообразных рептилий, последние были оттеснены в еще сохранившиеся более прохладные и влажные местообитания. В неблагоприятных для них условиях, при жесткой конкуренции со стороны завропсид, численность зверообразных рептилий значительно упала, и большинство их групп вымерло.
В немногих сохранившихся в триасе филогенетических стволах высших териодонтов продолжались изменения, наметившиеся еще среди пермских терапсид. Общее направление этих эволюционных изменений обозначают иногда термином "маммализация", т. е. развитие комплекса признаков, характерных для высших амниот -млекопитающих. Здесь мы вновь встречаемся с параллельной эволюцией: сходные черты строения независимо возникали в разных филетических линиях териодонтов. Среди этих признаков было постепенное увеличение размеров полушарий переднего мозга, приобретение мягких губ (что делало возможным сосание) и волосяного покрова. По мнению Л.П.Татаринова, зачаточные волоски у териодонтов имели осязательную функцию, располагаясь вблизи рта (как специализированные осязательные волосы - вибриссы у млекопитающих). Однако в среднетриасовых отложениях были обнаружены ископаемые следы цинодонта (Cynodontia - "собакозубые", одна из групп высших териодонтов) с отпечатками волос. Поэтому имеются основания считать, что у териодонтов волосяной покров был развит уже на всем теле, как у млекопитающих, и, вероятно, имел ту же основную функцию, как и у последних, т. е. служил для теплоизоляции (это понятно, если принять гипотезу о возникновении и развитии зверообразных рептилий в прохладном климате).
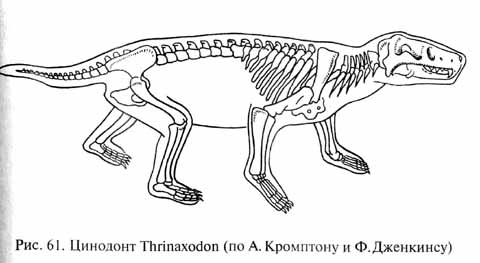

У цинодонтов (рис. 61) продолжалось совершенствование механизма пережевывания пищи с развитием окклюзии (смыкания коронок верхне- и нижнечелюстных зубов в щечной области) и продольных и поперечных движений нижней челюсти. Для обеспечения этих движений произошли сложные перестройки челюстных мышц, сыгравшие важную роль в дальнейшей эволюции челюстного аппарата. Механически наиболее благоприятное расположение челюстных мышц было достигнуто при разрастании зубной кости нижней челюсти назад и вверх с развитием высокого венечного отростка, к которому прикреплялась височная мышца (рис. 62). При этом задние кости нижней челюсти подверглись редукции. Гипертрофия задней части зубной кости Привела к возникновению ее контакта (при движениях нижней Челюсти) с чешуйчатой костью черепа. Между двумя этими костями возникла суставоподобная связь, которая оказалась механически более эффективной опорой и осью вращения для нижней челюсти, чем первичный челюстной сустав (между квадратной Костью верхней и сочленовной костью нижней челюсти). Преобразования нижней челюсти оказались преадаптивными для формирования новой опоры нижней челюсти - вторичного челюстного сустава.
Вторичный челюстной сустав, присущий среди всех позвоночных только млекопитающим, стал поэтому важнейшим диагностическим признаком последних. Редуцированные кости первичного челюстного сустава, освободившиеся от своей прежней функции, вошли в состав цепи косточек среднего уха в качестве двух наружных ее элементов - наковаленки и молоточка, что стало еще одной характерной особенностью млекопитающих, обеспечившей более тонкую слуховую чувствительность в области высоких звуковых частот.

Наиболее древние ископаемые остатки млекопитающих или зверей (Mammalia) известны из верхнетриасовых отложений. Это были небольшие хищные животные (рис. 63), питавшиеся различными мелкими позвоночными и беспозвоночными. Вероятно, они предпочитали прохладные и влажные местообитания и вели преимущественно ночной образ жизни. Скорее всего, у них уже существовало выкармливание детенышей молоком, поскольку оно присуще всем современным группам млекопитающих, а расхождение филогенетических стволов, ведущих, с одной стороны, к однопроходным (Monotremata), с другой - к сумчатым (Marsupialia) и плацентарным (Placentalia), произошло, возможно, уже в позднем триасе. Млечные железы возникли у древних млекопитающих из видоизмененных потовых желез. Возможно, первоначальной функцией млечных желез было не столько выкармливание, сколько "выпаивание" детенышей, т. е. снабжение их необходимой влагой и солями. Дефицит влаги был особенно опасен для новорожденных детенышей млекопитающих в условиях засушливого климата. Судя по способу развития однопроходных, наиболее примитивных среди современных млекопитающих, древнейшие звери еще были яйцекладущими животными, которые высиживали свою кладку или вынашивали яйца в специальной сумке на животе.
В конце триаса млекопитающие уже были довольно разнообразны и принадлежали к нескольким обособленным группам: три-конодонтам (Triconodontia), симметродонтам (Symmetrodontia) и харамиидам (Haramiyidae), различавшимся по строению зубов и, вероятно, по способам питания и пережевывания пищи.
Несмотря на ряд прогрессивных морфофизиологических особенностей (некоторое увеличение головного мозга, особенно его больших полушарий; забота о потомстве и выкармливание его молоком; пережевывание пищи; волосяной покров, расположение конечностей под туловищем, обеспечивающее более совершенную локомоцию), млекопитающие в мезозое не достигли больших успехов в конкуренции с рептилиями. Они оставались относительно малочисленными мелкими животными и, вероятно, вели ночной или сумеречный образ жизни, характерный для большинства зверей и ныне. Возможно, именно несовершенство систем терморегуляции и выделения, приспособленных к условиям относительно низких температур и высокой влажности, снижали для мезозойских млекопитающих шансы на успех в конкуренции с завропсидами. Хотя мезозойские млекопитающие уже обладали теплоизолирующим шерстным покровом и, вероятно, могли усилить теплоотдачу за счет потоотделения, у них не было еще совершенной интеграции разных терморегуляционных механизмов в единую систему. Об этом можно судить по состоянию современных однопроходных, у которых еще нет настоящей гомойотермии, и температура тела колеблется в пределах 26-34 °С.
В триасе начался подлинный расцвет рептилий. Особенно многочисленными становятся лепидозавры и архозавры. На ранних этапах их эволюционной истории эти два филогенетических ствола оставались еще очень близкими друг к другу, поэтому нередко трудно с достаточной уверенностью дифференцировать примитивных лепидозавров и архозавров. Таковы, например, ринхозавры (Rhynchosauria), которых одни авторы сближают с клювоголовы-ми среди лепидозавров, а другие относят к архозаврам. Ринхозавры получили широкое распространение в триасовое время. Это были крупные животные (1,5-5 м) с широким черепом, мошной жевательной мускулатурой и многорядными зубами на верхнечелюстных и зубных костях. Ринхозавры были растительноядными и, вероятно, конкурировали с дицинодонтами.
Настоящие клювоголовые (ринхоцефалы), родственные современной гаттерии, известны начиная с позднего триаса. В среднем триасе уже существовали ящерицы (группа эолацертилий), которые вскоре становятся очень разнообразными. Например, средне-триасовый танистрофей (Tanystropheus) обладал гипертрофированной шеей, длина которой превышала длину туловища (рис. 64, б)', общие размеры животного достигали 6 м. Возможно, эти рептилии, которых некоторые ученые также считают не ящерицами, а архозаврами, вели водный образ жизни, используя свою феноменальную шею при ловле рыб и головоногих моллюсков, как более поздние плезиозавры. Среди позднетриасовых эолацертилий имелись формы (Kuehneosaurus, Icarosaurus), приспособившиеся к планирующему полету: на удлиненных ребрах, выступавших по бокам туловища, была натянута летательная перепонка (рис. 64, а).

Возможно, вымирание последних котилозавров - проколофонов - в конце триаса было связано с их неудачей в конкуренции с экологически близкими, но более совершенными ящерицами.
Многочисленны и разнообразны в триасе были и представители архозавров. Корневой группой этого мощного филогенетического ствола являются псевдозухии, или текодонты (Pseudosi или Thecodontia), - преимущественно небольшие животные, примитивные представители которых внешне несколько напоминали ящериц. Но у псевдозухии череп сохранял типичное диапсидное строение, а зубы сидели в ячейках (текодонтный тип), а не прикреплялись к краю челюсти, как у ящериц. Для псевдозухии характерна тенденция к удлинению задних конечностей, связанная с переходом к быстрому бегу на двух ногах (рис. 65). Нужно отметить, что и ящерицы при максимальной скорости бегут лишь на двух задних конечностях, приподняв переднюю часть тела и длинный хвост над субстратом. Такой способ убыстрения бега (связанный с сохранением примитивной постановки конечностей, с горизонтальным положением плеча и бедра) был, видимо, характерен и для псевдозухии, но у последних получил более значительное развитие. Среди их потомков появились настоящие двуногие формы. Большинство псевдозухии были хищниками мелких и средних размеров (самые крупные достигали 3,5 м в длину). Некоторые из них стали быстро бегающими наземными животными, другие приспособились к лазанию по деревьям, были также полуводные формы - фитозавры (Phytosauria), имевшие внешнее сходство с крокодилами. В позднем триасе от псевдозухии возникли и настоящие крокодилы (Crocodylia), оказавшиеся лучше приспособленными к роли амфибиотических хищников и быстро вытеснившие фитозавров.

И фитозавры, и крокодилы конкурировали в водоемах с последними стегоцефалами - лабиринтодонтами, представленными в триасе очень крупными формами с огромным уплощенным черепом, достигавшим у Mastodonsauras 125 см длины. Однако и эти гиганты не выдержали конкуренции с водными рептилиями. Разнообразие и численность стегоцефалов в первой половине мезозоя неуклонно сокращались, и последние стегоцефалы (плагиозавры) вымерли в раннемеловое время.
В триасе появились также черепахи (Chelonia). Костно-роговой двуслойный панцирь черепах оказался замечательной защитной адаптацией, позволившей этим рептилиям сохранить многие примитивные черты организации, соответствующие уровню котилозавров. Происхождение черепах до сих пор остается дискуссионным. Древнейший известный их представитель, позднетриасовый Proganochelys, уже имел основные черты строения черепах. Разные исследователи видели предков черепах в различных группах котилозавров (от капториноморфов до парейазавров). М.Ф.Ивахненко, основываясь на некоторых особенностях строения черепа, высказал гипотезу о происхождении черепах от особого филогенетического ствола, названного Е.Олсоном парарептилиями и эволюционировавшего от уровня стегоцефалов независимо от других пресмыкающихся. Однако эта гипотеза требует допущения, Что все основные особенности амниот, рассмотренные нами выше (скелет, кожные покровы, дыхательная, кровеносная и выделительная системы, а также амниотический онтогенез), развивались независимо и параллельно у настоящих рептилий и парарептилий. Это представляется менее вероятным, чем параллельное (и неоднократное) формирование сходных особенностей черепа в разных линиях эволюции рептилий. Решение проблемы происхождения черепах, очевидно, ждет новых палеонтологических данных.
Жизнь в мезозойских морях
Теплые моря, заливы и лагуны в мезозое были богаты жизнью. После пермского вымирания разнообразие морских организмов в триасе вновь возрастает. По-видимому, в мезозое возник новый тип водорослей - диатомовые (Diatomeae). Во всяком случае, их ископаемые остатки достоверно известны, начиная лишь с юрских отложений.
В донной фауне опять обильны мшанки, многочисленны морские ежи и морские звезды. Постепенно возрастает численность и разнообразие брюхоногих моллюсков. Среди двустворчатых моллюсков выделяется мезозойская группа рудистов (Rudistae), прикреплявшихся к субстрату одной из створок своей асимметричной раковины, достигавшей у некоторых видов 1,5 м. В триасе начинается бурный расцвет аммонитов и белемнитов. Мезозойские головоногие были очень разнообразны и по форме раковины, и по размерам, варьировавшим у аммонитов от нескольких миллиметров до 3 м. Белемниты, подобно современным кальмарам, стремительно проносились в толще воды, охотясь за мелкими рыбами, а аммониты медленно плавали, регулируя глубину погружения с помощью раковины - гидростатического аппарата.
Постепенно обновилась ихтиофауна. В юрское время сформировались современные группы пластинчатожаберных хрящевых рыб (Elasmobranchia): акулы (Selachoidea) и скаты (Batoidea). Среди костных рыб появились новые группы лучеперых: костные ганои-ды (Holostei) и костистые рыбы (Teleostei). Первые пережили свой расцвет в юре, вторые возникли в юрское время, а начиная с мела стали доминирующей группой (к костистым принадлежит 95% всех современных видов рыб). Продолжали существовать в мезозойских морях кистеперые - целаканты.
Хрящевые рыбы (Chondrichthyes) полностью утратили костный скелет, характерный для всех древнейших палеозойских рыб. Это было, вероятно, связано с необходимостью освободиться от тяжелого наружного костного панциря при активизации образа жизни. Костные рыбы (Osteichthyes) также решали проблему облегчения костного панциря, но не путем его утраты вместе с внутренним костным скелетом, а посредством постепенных и более длительных преобразований, которые привели к формирований гибкого, легкого и прочного чешуйного покрова. Можно сказать, что в отношении преобразований наружного и внутреннего скелета хрящевые рыбы в сравнении с костными пошли по инадаптивному пути. В конкуренции с костными рыбами хрящевые рыбы компенсировали несовершенство своего скелета увеличением размеров тела и рядом прогрессивных изменений организации: совершенствование головного мозга, внутреннее осеменение, откладка крупных яиц, защищенных прочными оболочками, снабженных большим количеством желтка и обеспечивающих длительное развитие зародыша.
Всякая группа организмов, сумевшая в ходе эволюционных преобразований достичь нового уровня организации, под давлением естественного отбора начинает экологическую экспансию, осваивая все доступные местообитания, способы питания и т.п. При этом закономерно происходит адаптивная радиация - расхождение признаков у потомков общей предковой группы в процессе приспособительной эволюции, ведущее к возникновению разнообразных видов. Разнообразие развивающихся при этом приспособлений и возникающих жизненных форм зависит от совершенства и пластичности организации данной группы. Несомненно, одной из самых поразительных в этом отношении групп являются рептилии, которые в мезозое освоили практически все возможные типы местообитаний и дали фантастическое разнообразие жизненных форм, не превзойденное ни одним другим классом позвоночных животных.
Триас ознаменовался успешным освоением рептилиями не только пресных водоемов (танистрофеи, фитозавры и крокодилы), но и морей. Морские рептилии получили в мезозое широкое распространение и были чрезвычайно разнообразны: одни из них жили на мелководье в прибрежной зоне и, вероятно, сохраняли некоторую связь с сушей, другие же стали обитателями открытого океана и никогда не выходили на берег. Среди первых наиболее известны завроптеригии (Sauroptcrygia). Это были животные довольно странного облика (рис. 66): массивное широкое туловище, имевшее, однако, обтекаемую форму, впереди продолжал ось длинной и тонкой шеей (у некоторых форм она была длиннее туловища), а сзади - длинным сжатым с боков веслообразным хвостом, несущим сверху небольшой хвостовой плавник. Передние и задние конечности у более специализированных к жизни в море плезиозавров (Plesiosauria) имели строение широких и мощных ластов, которые были основными движителями в воде. Хвостовой Плавник был невелик и мог помогать при плавании с малой скоростью, а в основном он, вероятно, служил рулем. Ласты могли служить опорой при выходе плезиозавров на прибрежные отмели Для откладки яиц. как это делают современные морские черепахи. Однако даже такое кратковременное возвращение на сушу для Некоторых плезиозавров было очень затруднено их огромными размерами: ряд юрских и меловых видов достигал 12-16 м (плиозавры, элазмозавры).

Причудливый облик плезиозавров и упоминавшихся выше та-нистрофеев имеет современную аналогию, хотя и в миниатюре: у морских змей-ластохвостов (Hydrophis) также имеется утолщенное туловище, плоский хвост и длинная тонкая шея (в 4-5 раз более тонкая, чем туловище). Такие пропорции тела помогают водному хищнику использовать массивное и широкое туловище как своего рода опору в воде на плаву, по отношению к которой голова на длинной и гибкой шее имеет значительную подвижность при ловле добычи и борьбе с нею.
Пищей завроптеригиям служили различные рыбы и головоногие (аммониты и белемниты). Большинство плезиозавров, обладавшие небольшой головой на тонкой и длинной шее, питались относительно некрупной добычей. Зато появившиеся в позднеюрское время крупные плиозавры (Pliosauroidea) имели огромный череп (до 3 м у 12-метровых ящеров) с острыми зубами до 10 см длиной. Эти морские хищники могли, вероятно, питаться крупными глубоководными головоногими подобно современным кашалотам и даже нападать на более мелких плезиозавров и других морских рептилий.
С завроптеригиями некоторые палеонтологи сближают также плакодонтов (Placodontia). Эта любопытная группа морских пресмыкающихся существовала в триасе. Некоторые плакодонты напоминали современных морских черепах и даже имели панцирь из остеодерм, другие - современных морских млекопитающих - сирен (дюгони и ламантины). Зубы плакодонтов имели форму широких массивных пластин и, вероятно, служили для раздавливания раковин моллюсков и панцирей ракообразных.

Наиболее совершенными пловцами среди морских рептилий мезозоя, бороздившими как прибрежные моря, так и открытый океан, были ихтиозавры (Ichthyopterygia). Их название означает "рыбоящеры", что подчеркивает внешнее сходство этих животных с рыбами. Тело ихтиозавров приобрело идеально приспособленную для быстрого плавания рыбообразную форму (рис. 67). Размеры ихтиозавров чаще всего составляли 3-5 м, но некоторые формы (например, Shonisaurus) достигали длины 18 м. Череп ихтиозавров напоминал череп дельфинов большими глазницами, длинной узкой мордой, у основания которой сверху располагались ноздри, и длинными тонкими челюстями, усаженными многочисленными тонкими и острыми зубами. Большинство ихтиозавров плавали за счет работы мощного хвостового плавника, расположенного, как у рыб, вертикально, спинной плавник служил стабилизатором движения. Однако скелет плавников у ихтиозавров был образован не костными или хрящевыми элементами, как у рыб, а плотной соединительной тканью, как у китообразных среди млекопитающих. Парные конечности, преобразованные в ласты, играли роль рулей глубины. У некоторых ихтиозавров, подобных Eurhinosaurus, парные конечности были сильно развиты и использовались как веслообразные движители при плавании. В ластах ихтиозавров, как и у плезиозавров, было значительно увеличено число фаланг пальцев, а у некоторых видов увеличено было и число пальцев (до 10), так что скелет конечности включал до 200 небольших костей, расположенных правильными рядами, обеспечивая прочность и гибкость ластов.
Развитие вторичного, т.е. не унаследованного от общих предков, сходства строения у разных видов организмов в результате формирования приспособлений к сходному образу жизни (т.е. сходному способу использования одних и тех же ресурсов внешней среды) получило название конвергенции. Одним из классических примеров конвергенции является развитие сходных признаков у ихтиозавров, дельфинов и рыб.
Ихтиозавры, вероятно, уже не могли выходить на сушу даже для откладки яиц. Возможно, эти рептилии были живородящими. Живорождение (даже с образованием примитивной плаценты) не так уж редко встречается среди пресмыкающихся (оно характерно для целого ряда видов современных ящериц и змей). О живорождении у ихтиозавров свидетельствуют некоторые палеонтологические данные: были обнаружены скелеты крупных особей (самок?), внутри которых находились скелетики маленьких ихтиозавров, иногда свернутые кольцом (как зародыши в яйце), иногда располагавшиеся так, что хвостовая часть скелета детеныша должна была выступать наружу из отверстия клоаки самки. Можно предполагать, что эти находки связаны с гибелью самки вместе с детенышем в момент родов. Судя по этим данным, у ихтиозавров при родах, как и у современных китообразных, детеныши выходили хвостом вперед, и сразу же рефлекторно начинал работать хвостовой плавник.
Любопытно, что вместе с некоторыми скелетами ихтиозавров были обнаружены остатки фоссилизованной кожи, причем сохранились даже пигментные гранулы. Возможно ихтиозавры имели темно-коричневую окраску. Было обнаружено также фоссилизованное содержимое желудков ихтиозавров, в нем оказались остатки головоногих моллюсков и рыб. Вероятно, в мезозойских морях ихтиозавры соответствовали в экологическом отношении современным дельфинам. Приспособление к сходному образу жизни в одной и той же среде обитания привело к значительному конвергентному сходству тех и других.
Происхождение специализированных водных рептилий - завроптеригий и ихтиозавров - до сих пор остается дискуссионным, поскольку, как и в большинстве случаев, ранние этапы эволюции этих групп не представлены в палеонтологической летописи. Некоторые палеонтологи склоняются к мнению об их независимом происхождении непосредственно от какой-либо группы котилозавров (капториноморфов или проколофонов). Высказывалось также предположение об общем происхождении завроптеригий и ихтиозавров от котилозавров или от примитивных синапсидных рептилий (пеликозавров), а в последние годы получила распространение гипотеза об их возникновении от примитивных диапсидных рептилий, близких к эозухиям (в этом случае принимают, что нижнее височное окно в их черепе вторично закрылось благодаря разрастанию костей).
Завроптеригии в лице своих наиболее примитивных представителей - нотозавров (Nothosauria), имевших еще не настоящие ласты, а лишь укороченные конечности с плавательными перепонками между пальцами, появились в раннем триасе. Ихтиозавры известны начиная с нижнего триаса. Расцвет обеих групп приходится на юрский период и первую половину мела.
Ископаемые остатки завроптеригий и ихтиозавров нередко встречаются в юрских осадочных породах на Русской равнине. Целые скелеты плиозавров были обнаружены при разработках горючих сланцев в Поволжье.
Помимо завроптеригий и ихтиозавров к жизни в море перешли и многие другие мезозойские рептилии. Можно сказать, что все подклассы пресмыкающихся имели в море своих представителей. В позднеюрское время появились первые морские черепахи. Архозавры были представлены в море специализированными группами крокодилов, конкурировавшими с ихтиозаврами. Морские крокодилы были очень многочисленны во второй половине мезозоя. Некоторые из них (метриоринхи - Metriorhynchidae), как и ихтиозавры, приобрели рыбообразное тело, вертикальный хвостовой плавник и ластообразные конечности.
Лепидозавры также делали неоднократные попытки "овладеть океаном". Среди морских ящериц наиболее замечательны позднемеловые мозазавры (Mosasauridae), достигавшие 10 м длины и ставшие столь же совершенными пловцами, как ихтиозавры и метриоринхи. Мозазавры по времени своего появления и существования были самыми поздними из этих трех конвергентных групп. Высказывались предположения, что конкуренция со стороны мозазавров могла сыграть роль в вымирании ихтиозавров, большинство видов которых исчезло уже в середине мела.
Век динозавров
К концу триаса произошло постепенное выравнивание климатических условий на значительной части суши, о чем говорит, в частности, очень однообразный и в целом сходный характер флоры на больших пространствах континентов, входивших прежде в состав Гондваны и Лавразии. Растительность юрского периода (рис. 68) характеризовалась преобладанием различных групп голосеменных (цикадовые, беннеттиты, хвойные, гинкговые) и древовидных папоротников; широко распространены были также крупные хвощи, среди которых, например, Equisetites arenaceus достигал 10 м высоты и 25 см в диаметре. Мезозойские хвойные были значительно разнообразнее и шире распространены, чем в наше время. В эту эру появились все современные семейства хвойных и большинство современных родов. Через Канаду, Сибирь и северную Часть Европы простирался пояс хвойных и листопадных лесов, произраставших в условиях умеренного климата. Южнее располагался еще более обширный пояс вечнозеленой субтропической растительности с обилием влажных и заболоченных местностей. Тропический экваториальный пояс был значительно шире современного.
В условиях ровного и очень теплого климата в средней части мезозоя завропсидные рептилии достигли максимального расцвета. Это было время удивительных животных, получивших широкую известность под названием "динозавры" ("страшные ящеры"). Название это относится к нескольким независимым группам рептилий из подкласса архозавров и в современном понимании не имеет определенного таксономического статуса (подобно термину "стегоцефалы", см. выше). Обычно динозавров распределяют в два отряда: ящеротазовых (Saurischia) и птицетазовых (Ornithischia), отличавшихся друг от друга рядом признаков. В частности, у них было разное строение тазового пояса: у птицетазовых лобковая кость имела особый отросток, тянувшийся назад под седалищной костью, тогда как у ящеротазовых такого отростка не было (рис. 69).

Динозавры одним или несколькими независимыми корнями (вопрос этот еще остается дискуссионным) произошли от псевдозухий. Древнейшие остатки ящеротазовых известны из средне-триасовых, а птицетазовых - из верхнетриасовых отложений Восточной и Южной Африки и Южной Америки. Примитивные динозавры унаследовали от псевдозухий тенденцию передвигаться на двух задних конечностях - бипедальность. В.Б.Суханов показал, что при сохранении примитивного положения конечностей (см. с.210) бипедальная локомоция позволяет увеличить скорость. Кроме того, при двуногом передвижении передняя часть тела высоко поднимается над почвой, что увеличивает обзор и уменьшает опасность перегрева при длительном пребывании животного на сухих открытых участках, где почва раскаляется под лучами яркого солнца. Бипедальность была широко распространена среди динозавров, хотя некоторые их группы по различным причинам вновь вернулись к опоре на четыре конечности.
Как и у высших звероподобных рептилий, у динозавров конечности располагались под телом в вертикальной плоскости, что обеспечивало большую экономичность передвижения по сравнению с группами, сохранившими позу пресмыкания. Вероятно, двуногие ящеры передвигались подобно современным страусоподобным птицам, используя бег или ходьбу с чередованием опоры на левую и правую ноги. Длинный мускулистый хвост уравновешивал высоко поднятую тяжелую голову и переднюю часть тела. Возможно, хвост служил также дополнительной опорой при стоянии в выпрямленной позе. Передние конечности у бипедальных динозавров либо использовались как хватательные, либо подвергались редукции.
В юрском и меловом периодах динозавры доминировали среди наземных позвоночных, заселив всевозможные местообитания и дав большое разнообразие форм, которые поражают воображение гротескной причудливостью облика, а иногда и огромными размерами.

Безусловный рекорд в последнем отношении принадлежит ящеротазовым динозаврам из группы завропод (Sauropoda), среди которых были самые крупные из когда-либо существовавших на Земле четвероногих животных: например, диплодок (Diplodocus, рис. 70) достигал 25-30 м длины при высоте в области середины спины около 4 м, а вес этого гиганта составлял около 30 тонн. Несколько более короткий (24 м) брахиозавр (Brachiosaurus) имел более высокие передние конечности и массивное тело, вес которого по расчетам мог достигать 45-50 тонн. Из всех животных Земли на протяжении всей истории жизни больше и тяжелее завропод лишь некоторые современные китообразные, но киты - чисто водные животные, тогда как завроподы оставались по своему общему облику наземными четвероногими. Правда, и завроподы значительную часть жизни, скорее всего, проводили в воде или по берегам водоемов. Об этом говорят, помимо гигантских размеров и веса (при которых очень трудно свободно передвигаться по суше), многие особенности строения этих удивительных животных, например смещение наружных носовых отверстий по верхней стороне черепа назад, так что ноздри располагались рядом с глазницами. Когда животное находилось в воде, длинная шея позволяла поднимать голову над поверхностью для дыхания и обзора (брахиозавр мог поднять голову выше крыши трехэтажного дома), а также добывать корм со дна водоемов. Судя по строению зубов (мелких, тонких, иногда ложкообразных), завроподы были растительноядными животными, питавшимися, вероятно, какими-то сочными, мягкими и обильными водными растениями.
Колоннообразные конечности завропод опирались на огромную стопу, пальцы которой были вооружены большими когтями. Вероятно, последние помогали животным прочно удерживаться на месте при ударах волн. Возможно, завроподы могли и плавать с помощью сжатого с боков мощного хвоста. Завроподы обитали по берегам морей и озер, где они могли бродить в воде многочисленных лагун, речных дельт и эстуариев, в приливо-отливной зоне (занимавшей на низменных материках мезозоя гораздо большие площади, чем ныне), среди мангровых зарослей.
Другой группой яшеротазовых были тероподы (Theropoda) -хищные двуногие ящеры, занявшие в мезозое место синапсидных хищных рептилий пермского периода. Среди теропод более примитивны были целюрозавры (Coelurosauria) - мелкие подвижные формы (размерами от нескольких десятков сантиметров до нескольких метров) с длинными передними конечностями, помогавшими при ловле добычи. Питались они, вероятно, разнообразной пищей, включавшей крупных беспозвоночных и мелких позвоночных (ящериц, лягушек, млекопитающих и т.п.), а также, возможно, раскапывали кладки яиц более крупных рептилий. Некоторые целюрозавры, возможно, специализировались на этом последнем способе питания. В частности, такое предположение было высказано о позднемеловой группе орнитомимид (Ornithomimidae), легких и быстрых страусоподобных целюрозавров с длинными хватательными передними конечностями и беззубыми челюстями, одетыми, вероятно, роговым клювом.
Второй группой теропод были карнозавры (Carnosauria), к которым принадлежали самые крупные хищники, когда-либо существовавшие на суше: мегалозавры (Megalosauridae) и дейнодонты (Deinodontidae). Некоторые из этих чудовищ (например, тираннозавр - Tyrannosaurus rex, "царственный ящер-тиран", рис. 71) достигали длины 12-14 м. Полутораметровый череп, вооруженный острыми кинжалообразными изогнутыми зубами с пильчатыми краями (до 10-15 см длиной), располагался на высоте 4-6 м. По расчетам, вес тираннозавров достигал 7 тонн. Передние конечности у этих гигантских хищников редуцировались до маленьких придатков с 2-3 пальцами. Зато череп обладал подвижной верхней челюстью: черепной кинетизм, унаследованный от далеких предков, у теропод усилился и усовершенствовался, как и у лепидозавров, но несколько иным путем. Вероятно, подвижность верхней челюсти в какой-то мере компенсировала редукцию передних конечностей. Подвижность верхней челюсти позволяет лучше удерживать добычу; кинетический череп в этом отношении имеет такое же преимущество перед акинетическим, как рука с гибкими пальцами перед клешней.

Вероятно, основным способом охоты у крупных карнозавров было преследование добычи (едва ли такие гиганты могли подстерегать добычу в засаде) с последующим страшным ударом тяжелого черепа, вооруженного кинжалообразными зубами, сверху, с высоты огромного роста хищника в шею или спину жертвы. Не случайно у многих растительноядных динозавров имелись специальные приспособления для защиты шеи и спины от ударов сверху.
Среди карнозавров были не только гигантские, но и более мелкие хищники размером 1-2 м. При сохранении одного общего плана строения карнозавры были достаточно разнообразны. Упомянем, например, мелового спинозавра (Spinosaurus), у которого остистые отростки туловищных позвонков были гипертрофированы, достигая длины 1,8 м. Очевидно, вдоль спины этого крупного двуногого хищника тянулся высокий гребень, подобный спинному парусу некоторых пеликозавров, и также, вероятно, игравший роль в терморегуляции.
Все птицетазовые динозавры были растительноядными животными. Среди них имелись и двуногие, и четвероногие ящеры. Первые (подотряд орнитопод - Ornithopoda) варьировали в размерах от 1 до 15м. Вес взрослых особей крупных видов составлял до 5-7 тонн. Это были наземные или полуводные формы с довольно разнообразными пропорциями тела и внешним обликом. Представители наземных групп: пситтакозавры (Psittacosauridae - "ящеры-попугаи") и игуанодоны (Iguanodontidae) имели на пальцах задних конечностей небольшие копытца. Вероятно, они обитали bo влажных тропических лесах, питаясь сочной зеленью и плода-Ми. Пситтакозавры получили свое название за внешнее сходство Их черепа (с высоким сжатым с боков роговым клювом) с чере-Пом попугаев; игуанодоны же обладали крупными зубами, коронки которых с небольшими зубчиками на режущем крае напоминали зубы современных растительноядных ящериц-игуан. Большие пальцы на передних конечностях игуанодонов были вместо копыт снабжены крепкими острыми когтями, вероятно, использовавшимися при обороне от хищников. Размеры игуанодонов достигали 10 м.

У позднемеловых орнитопод - гадрозавров (Had-rosauridae, рис. 72), называемых также "утконосыми динозаврами" (за форму передней части челюстей, лишенной зубов и одетой роговым "утиным" клювом), между пальцами передних и задних конечностей имелись плавательные перепонки, сохранившиеся у мумифицированных экземпляров, найденных в Северной Америке. Гадрозавры жили, подобно завроподам, по берегам различных водоемов и в мангровой зоне, но в более закрытых лесных и, возможно, болотистых ландшафтах. Зубы этих животных, расположенные в задней части челюстей, были приспособлены к перетиранию жесткой растительной пищи: позади клюва располагались несколько рядов зубов, тесно прилежащих друг к другу и составлявших "зубные батареи", которые включали до 500 зубов в одной половине челюсти (т. е. общее число зубов было около 2000). У многих гадрозавров череп был снабжен гребнем или шлемом, образованным разросшимися предчелюстными, носовыми и лобными костями. Внутри шлема находилась полость, связанная с носовым ходом. Назначение этого приспособления остается пока непонятным.

Другие группы птицетазовых динозавров вернулись к четвероногому передвижению. Пожалуй, наиболее странный облик среди них имели стегозавры (Stegosauria, рис. 73). Это были крупные (6-9 м в длину) животные с маленькой низко посаженной головой, позади которой высокой крутой дугой горбилась спина. Задняя часть туловища была приподнята на задних конечностях, которые были вдвое длиннее передних. Более длинные задние конечности вообще характерны для подавляющего большинства динозавров, что, вероятно, связано с двуногостью примитивных динозавров. Но самой причудливой особенностью стегозавров было защитное приспособление в виде двух рядов огромных костных пластин и шипов, вертикально укрепленных в толстой коже животного над позвоночником и тянувшихся от затылка до кончика хвоста. Вероятно, стегозавры были медлительными сухопутными животными. Они просуществовали лишь до раннего мела, когда их сменили другие крупные растительноядные динозавры, вероятно лучше защищенные от нападений гигантских меловых карнозавров.

В течение всего мела существовали анкилозавры (Ankylosauria, рис. 74), которых называют иногда "ящерами-танками". Анкилозавры имели длину 3,5-5 м при среднем весе 3,7 тонны. В противоположность стегозаврам они имели широкое приземистое тело, поддерживаемое короткими толстыми ногами. У анкилозавров чрезвычайно сильное развитие получили остеодермы - вторичные кожные окостенения, прираставшие к поверхности костей черепа (придавая ему облик, напоминавший череп древних парейазавров) и образовавшие костный панцирь на шее, туловище и хвосте из поперечных рядов пластинок или шипов.
В позднемеловое время существовала еще одна группа растительноядных четвероногих птицетазовых динозавров - рогатые динозавры (Ceratopsia, рис. 75), которых можно назвать своего рода ящерами-носорогами. Цератопсы достигали длины 6 м при высоте до 2,5 м и среднем весе около 4,3 тонны. Защитные приспособления цератопсов включали прочный костный "воротник", образованный разрастаниями теменных и чешуйчатых костей назад и закрывавший шею сверху и сбоку. Помимо этого органа пассивной защиты многие цератопсы имели приспособления для, так сказать, "активной обороны": на морде и над глазами возвышались три крепких и острых рога (отсюда название одного из родов этих ящеров: Triceratops - "трехрогий"), иногда на скулах сидела еще пара дополнительных рогов. Вероятно, взрослые цератопсы могли обороняться даже от нападений крупных хищников.

Среди биологических особенностей динозавров чаще всего обращают внимание на широкое распространение среди этих рептилий гигантизма. Действительно, многие динозавры были очень крупными, а некоторые - гигантскими животными. Однако необходимо отметить, что отнюдь не все динозавры были гигантами, среди них были и сравнительно небольшие ящеры, хотя и эти последние были в среднем крупнее, чем большинство лепидозавров. Крупные размеры тела обеспечивают животным определенные физиологические преимущества. С увеличением размеров поверхность тела возрастает приблизительно в квадрате, а объем и масса - в кубе по отношению к линейным размерам. В результате отношение поверхности тела к его массе у более крупных животных будет при одинаковых пропорциях тела существенно меньше, чем у мелких. Поэтому крупные размеры оказываются в энергетическом отношении выгоднее: теплоемкость тела велика, а теплообмен с внешней средой через поверхность тела относительно мал; следовательно, уменьшается зависимость температуры тела от внешних температур, уменьшается риск перегрева и переохлаждения организма. По расчетам ученых, у крупных динозавров для повышения температуры тела всего на 1 градус потребовалось бы 86 часов непрерывного пребывания на солнцепеке. Соответственно при большой массе и относительно малой поверхности тела замедляется и понижение температуры тела при пребывании животного в среде с низкой температурой. В условиях преобладания ровных теплых климатов в течение большей части мезозоя крупные динозавры, вероятно, обладали практически постоянной оптимальной температурой тела при общем характере метаболизма и систем терморегуляции, вполне сходными с таковыми у современных рептилий. Такая температура тела постепенно формировалась в онтогенезе: молодые животные имели небольшие размеры, и их температура должна была изменяться, как и у современных рептилий. Вероятно, они также использовали гелиотермию для достижения оптимальной температуры.
(Р.Баккер высказал предположение, что динозавры были настоящими гомойотермными животными, т.е. имели постоянную температуру тела, основанную на высоком уровне метаболизма и интегрированных системах терморегуляции, подобно птицам и млекопитающим. Однако эта гипотеза основана лишь косвенных данных, легко поддающихся противоположной интерпретации).
Физиологически динозавры были, скорее всего, в целом сходны с современными архозаврами - крокодилами. Вполне вероятно наличие у динозавров четырехкамерного сердца и двух дуг аорты, начинающихся от разных желудочков и имеющих между собой связь (анастомоз), что позволяло, как у современных рептилий, регулировать потоки крови, направляемые в отходящие от сердца сосуды, в зависимости от физиологической потребности. В частности, при обогревании в солнечных лучах (или при нырянии у водных рептилий) уменьшается приток крови из правого желудочка в легочные артерии и соответственно возрастает в левую дугу аорты.
Как и у современных рептилий, головной мозг динозавров имел сравнительно небольшие размеры. Особенно это бросается в глаза у гигантских ящеров, полость мозговой коробки которых поражает своими ничтожно малыми размерами по отношению к размерам черепа и всего тела. Зато спинной мозг был сильно утолщен в крестцовой области, причем нередко это утолщение было много больше по объему, чем головной мозг. Вероятно, здесь находились нервные центры, ответственные за работу задних конечностей, хвоста и различные безусловно-рефлекторные реакции. Особенно велико было крестцовое утолщение спинного мозга у завропод. Например, у диплодока его поперечный диаметр примерно в 20 раз превышал таковой головного мозга, что, собственно, и отмечено в названии этого ящера: Diplodocus - "двуум".
Как и большинство современных рептилий, динозавры были яйцекладущими животными. Известны многочисленные находки ископаемых яиц, которые считают принадлежащими динозаврам. В верхнемеловых отложениях Прованса во Франции, Монтаны в США и ряде других районов были обнаружены хорошо сохранившиеся кладки яиц, предположительно отложенные различными орнитоподами. Кладки включают по 4-8 яиц, объем которых составляет 0,4-3,3 литра. Внутри известковой скорлупы некоторых ископаемых яиц были обнаружены скелеты эмбрионов. Не исключено, что отдельные виды динозавров могли быть живородящими. Весьма вероятно, что динозавры охраняли свои гнезда и некоторое время ухаживали за детенышами, как это делают современные крокодилы.
Ископаемые остатки динозавров известны на всех континентах; недавно они были обнаружены в районах, располагавшихся в конце мезозоя за Полярным кругом (до 85° северной и южной широты). Климат всей Земли во второй половине мезозоя был значительно теплее современного, и в этих высоких широтах произрастали хвойные и лиственные леса. Однако среднегодовые температуры здесь составляли лишь от 5 до 8 °С и зимой, вероятно, падали ниже нуля. В связи с этим интересной проблемой является образ жизни динозавров, обитавших за Полярным кругом. Полярной ночью крупные пресмыкающиеся вряд ли могли найти для себя достаточно пищи. Трудно представить также, как они могли сохранить оптимальную температуру тела в течение длительного периода низких температур внешней среды, поскольку крупные размеры тела исключали для них возможность зимовки в убежищах. Возможно, полярные динозавры ежегодно совершали сезонные миграции за границы Полярного круга.
Летающие ящеры и птицы
Завропсидные рептилии в мезозое овладели не только сушей и морем, но и воздухом. По крайней мере две группы этих пресмыкающихся приобрели способность к полету.
Первой из них были птерозавры (Pterosauria) - летающие ящеры (рис. 76, 77), появившиеся в конце триасового периода. Крылья птерозавров, как у летучих мышей, были образованы летательными перепонками, натянутыми между передними и задними конечностями и телом. Но если у летучих мышей крыло Поддерживается четырьмя удлиненными пальцами передней ко-Нечности, то у птерозавров - лишь одним гипертрофированным Четвертым пальцем. Три других пальца кисти у них были свободны, имели когти и располагались на сгибе крыла. Вероятно, птерозавры могли использовать их при лазании, цепляний и ходьбе, птерозавры были способны к разнообразным формам полета (машущего и планирующего), как это было показано аэродинамическими расчетами и экспериментами с моделями, имитирующими летательный аппарат этих животных. Судя по многочисленным ископаемым следам, оставленным птерозаврами при ходьбе по земле, они передвигались по субстрату, опираясь на все четыре конечности: сложенные крылья были широко расставлены в стороны и опирались на пальцы.
Многие особенности строения птерозавров предвосхитили связанные с полетом приспособления птиц и летучих мышей, возникшие независимо у каждой из этих трех групп летающих позвоночных. Так, скелет птерозавров характеризовался высокой прочностью и легкостью, многие кости были пневматизированы, многие кости черепа слились друг с другом, сильно развитая грудина несла продольный киль для прикрепления мощных летательных мышц. Хотя у примитивных птерозавров сохранялись зубы, высшие представители этой группы утратили их и приобрели роговой клюв. Головной мозг летающих ящеров в целом ряде отношений напоминал мозг птиц: полушария переднего мозга значительно увеличены, тогда как обонятельные доли редуцированы, очень велик мозжечок, крупные зрительные доли среднего мозга оттеснены вниз и отчасти прикрыты большими полушариями и мозжечком.
Тело птерозавров было покрыто не чешуей, как у большинства рептилий, а тонкими волосками, сохранившимися, как и летательные перепонки, на отпечатках в тонкозернистых породах, вмещающих ископаемые остатки. Волосяной покров является эффективным теплоизолятором. Его наличие говорит в пользу достижения птерозаврами настоящей гомойотермии - постоянной температуры тела, регулируемой физиологическими механизмами.
Более примитивной группой летающих ящеров являются юрские рамфоринхи (Rhamphorhynchoidea), обладавшие длинным хвостом и хорошо развитыми зубами (см. рис. 76). В позднеюрское время появляются птеродактили (Pterodactyloidea), у которых хвост редуцировался, как и у высших птиц (см. рис. 77). Вероятно, птеродактили обладали более совершенным полетом, чем рамфоринхи. Разные виды летающих ящеров сильно различались по размерам и массе тела (от 4 г до 75 кг), строению челюстей, форме крыльев и, очевидно, по характеру полета и образу жизни. Среди птеродактилей встречались виды размером с воробья, а самые крупные птерозавры были самыми большими летающими животными, когда-либо существовавшими на Земле. Долгое время рекорд размеров принадлежал позднемеловому птеранодону (Pteranodon) - огромному ящеру, размах крыльев которого составлял 7,5-8 м, а их площадь около 6 м2; масса тела достигала 18-25 кг. Череп птеранодона имел странный торчащий далеко назад затылочный гребень (может быть, уравновешивавший тяжелый клюв), и общая длина черепа от конца гребня до переднего конца беззубых челюстей достигала 2 м.
Однако в конце 70-х г. были обнаружены ископаемые остатки еще более крупных по-зднемеловых птерозавров (Quetzalcoatlus, Titanopteryx, Azhdarcho и др.) размах крыльев которых достигал 15-16 м, а масса тела - 75 кг. По одной из гипотез, эти гигантские птерозавры питались трупами динозавров (т.е. были аналогами современных крупных птиц-падалыциков: грифов, сипов, стервятников и др.), по другой - добывали пищу, планируя над водой и опуская вниз голову на длинной шее.
Среди более мелких видов птеродактилей и рамфоринхов были насекомоядные и рыбоядные формы. Возможно, некоторые птерозавры могли плавать, подобно современным водоплавающим птицам. У некоторых птеродактилей (Belonochasma, Ctenochasma) в челюстях сидели тесно посаженные крайне многочисленные, до 1000, очень тонкие и длинные щетинообразные зубы, которые могли использоваться как цедильный аппарат при питании планктоном.
Мелкие птеродактили и длиннохвостые рамфоринхи при полете, вероятно, часто взмахивали крыльями, тогда как гигантские формы величественно парили на большой высоте, используя поддержку восходящих потоков воздуха и помогая полету редкими взмахами огромных крыльев.
Птерозавры были единственными летающими позвоночными на протяжении большей части юрского периода. В поздней юре у них появились конкуренты, дальнейшая эволюция которых привела к формированию более совершенного летательного аппарата и к достижению более высокого общего уровня организации. Это были птицы (Aves).
Впрочем, примитивные позднеюрские птицы еще во многих отношениях уступали летающим ящерам и не могли спорить с ними за первенство в воздухе. Остатки древнейших птиц, широко известных археоптериксов (Archaeopteryx lithographica), были найдены в тонкозернистых сланцах Золенгофена в Германии; ныне известно уже 7 экземпляров археоптерикса.

В организации археоптерикса (рис. 78) причудливо объединены, с одной стороны, птичьи, а с другой - рептильные признаки. Его тело было покрыто настоящими перьями, причем крупные маховые перья с асимметричными опахалами, как у современных птиц, Формировали несущую поверхность крыльев. Мозговая коробка была Увеличена по сравнению с типичными рептилиями, и головной Мозг рядом особенностей напоминал птичий, хотя и не в большей степени, чем у птерозавров. С другой стороны, общий облик черепа был вполне обычен для архозавров; челюсти несли хорошо развитые текодонтные зубы. Некоторые птичьи особенности имелись также в поясах конечностей археоптерикса (например, характерная "вилочка", образованная ключицами). Позвоночник сохранял примитивное строение. Он состоял из амфицельных (двояковогнутых) позвонков и продолжался сзади в длинный хвост, к которому перья прикреплялись по бокам, а не веером на конце копчика, как у современных птиц. Кости не были пневматизиро-ваны, а грудина невелика и лишена киля - следовательно, у археоптерикса не могло быть мощных летательных мышц. Скелет передних конечностей был во многом сходен с таковым рептилий, имелись три длинных свободных пальца с когтями. Таким образом, в организации археоптерикса, как в мозаике, соединены признаки, присущие рептилиям и птицам. Де Вир назвал подобные виды "мозаичными формами".
Вряд ли археоптериксы хорошо летали. Скорее всего, они лишь перепархивали на небольшие расстояния среди кустов и ветвей деревьев. Современные им птерозавры, несомненно, обладали значительно более совершенным полетом.
Птицы, как и летающие ящеры, возникли от мелких архозавров, приспособившихся к лазанию по деревьям. Более древние наземные предки птиц, несомненно, использовали двуногое передвижение, о чем можно судить по строению задних конечностей археоптериксов и современных птиц, сходному с типом строения конечностей бипедальных ящеротазовых динозавров-теропод. При лазании по деревьям и прыжках с ветки на ветку полезны любые приспособления, позволяющие планировать, удлиняя прыжки. У птерозавров таким приспособлением стали летательные перепонки, а у птиц - крупные маховые перья. Перья возникли в результате преобразования роговых чешуи, характерных Для всех рептилий. Первоначально основной функцией перьевого покрова была теплоизоляция. Усовершенствование этой первичной функции, сопровождавшееся разрастанием перьев и усложнением их структуры, явилось преадаптацией для использования перьев в формировании несущей поверхности крыльев, сначала Для планирования, а затем и для машущего полета.
Птерозавры, по всей вероятности, произошли от одной из групп среднетриасовых псевдозухий. Аналогичное происхождение, но от Другой группы псевдозухий. ряд ученых приписывает и птицам. А.Вокер, в частности, полагает, что предки птиц были близки к Предкам крокодилов: и тех и других, согласно Вокеру, следует Искать среди псевдозухий-сфенозухид (Sphenosuchidae), которые, возможно, были древесными лазающими формами.
Согласно другой гипотезе, птицы произошли не непосредственно от псевдозухий, а от их потомков - одной из групп юрских динозавров. Д.Остром указал на значительное сходство многих особенностей скелета археоптерикса и целюрозавров (примитивных ящеротазовых теропод). Сходные черты прослеживаются практически во всех отделах скелета этих животных. В пользу этой гипотезы говорит и значительный промежуток времени, отделяющий археоптерикса от самых поздних известных псевдозухий (около 20 млн лет), тогда как средне- и позднеюрские целюрозавры по времени своего существования оказываются значительно ближе к археоптериксу.
Однако в 1991 г. Ш.Чаттерджи описал ископаемую форму, названную им Protoavis ("протоптица"), остатки которой были найдены в верхнетриасовых отложениях Техаса в США. По мнению Чаттерджи, протоавис обладал многими важными признаками птиц (строение ряда костей черепа, позвонков, костей конечностей и их поясов, пропорции и крупные размеры головного мозга) и был способен к настоящему машущему полету. Е.Н.Курочкин также считает протоависа близким к птицам, но еще не способным летать. Если эти данные подтвердятся, происхождение птиц придется отодвинуть по крайней мере в первую половину юрского периода.
Что же касается археоптерикса, то его филогенетические взаимосвязи с другими группами птиц остаются дискуссионными. Ряд исследователей полагает, что археоптерикс, выделяемый в особый подкласс ящерохвостых птиц (Saurarae), не является предком современных птиц (подкласса птицехвостых - Ornithurae), a представляет раннюю боковую ветвь филогенетического ствола, ведущего к последним. Е.Н.Курочкин сближает с археоптериксовыми группу ископаемых меловых птиц - энанциорнисов (Enantiornithes), уже обладавших летательным аппаратом, характерным для высших птиц, но сохранявших ряд общих с археоптериксами примитивных признаков скелета.
В раннемеловое время появляются также наиболее древние представители настоящих птицехвостых (или веерохвостых) птиц. Судя по ископаемым остаткам раннемеловых птиц Ambiorthus и Ibero-mesornis, эти животные по всем основным признакам уже были настоящими птицами и хорошо летали. Возможно, приспособления к полету развивались независимо и параллельно в филогенетических стволах энанциорнисов и веерохвостых птиц.
К концу мелового периода разнообразие птиц значительно возросло. Среди позднемеловых птиц известны формы, близкие к некоторым современным группам (ржанкообразные, журавлеоб-разные, предки дятлообразных и др.), но существовали еще виды, сохранявшие архаическое строение. Таковы были энанциорнисы, а среди птицехвостых птиц - гесперорнисы (Hesperornithes) ихтиорнисы (Ichthyornithes) (рис. 79). Головной мозг у этих групп был еще относительно невелик, в челюстях сидели мелкие острые зубы, у некоторых видов позвонки оставались амфицельными, а редукция хвостового отдела еще не достигла состояния, типичного для высших птиц. Многие позднемеловые птицы были околоводными и водоплавающими формами, причем гесперор-нисы вторично утратили способность к полету. Крылья у геспе-рорнисов были редуцированы, и плавали эти птицы с помощью работы мощных задних конечностей. По своему внешнему облику и, вероятно, в экологическом отношении гесперорнисы напоминали современных гагар.
С усовершенствованием полета, приобретением гомойотермии и усложнением головного мозга птицы становились все более серьезными конкурентами летающих ящеров. Возможно, конкуренция со стороны птиц была косвенной причиной эволюции птерозавров в направлении все большего возрастания размеров, вплоть до гигантских летающих ящеров позднего мела. Во второй половине мелового периода происходила широкая адаптивная радиация птиц, в процессе которой возникли основные современные их группы (большинство из них известны начиная с палеогена). В верхнемеловых отложениях Аргентины были обнаружены ископаемые остатки нелетающей птицы Patagopteryx, которая, возможно, родственна предкам страусов. Таким образом, уже на ранних этапах своей эволюции довольно многие птицы утрачивали способность к полету, что, вероятно, было связано с отсутствием пресса хищников в их местообитаниях. Часто это происходило, например, на островах, где не было наземных хищников.
Изменения наземных биоценозов во второй половине мезозоя
Как мы уже отмечали в начале этой главы, физико-географические условия в течение большей части мезозоя оставались в целом стабильными. На континентах господствовал ровный, теплый и влажный климат. На этом фоне весьма удивительным выглядит экологический кризис сообщества наземных позвоночных, который, вероятно, имел место в ранней и средней юре. Н.Н.Каландадзе и А.С.Раутиан показали, что этому времени соответствует чрезвычайное обеднение фауны наземных тетрапод, выразившееся в резком падении числа их семейств и родов по сравнению с триасом. Это относится к большинству групп амфибий и рептилий. В позднеюрское время этот кризис был преодолен, и разнообразие форм наземных позвоночных вновь резко возрастает. Отсутствие видимых внешних (абиотических) причин юрского кризиса заставило предположить, что его причины коренились в специфике развития самого сообщества организмов, механизм устойчивости которого нарушились в результате значительной специализации большинства входивших в его состав видов и дефицита видов-универсалов.
Во второй половине мезозоя на фоне привлекающей широкое внимание эволюции архозавров происходили и многие другие -может быть, внешне менее эффектные, но не менее важные события.
В начале мела от ящериц возникли змеи (Ophidia). Среди ящериц нередко появлялись формы с сильно удлиненным телом и редуцированными конечностями (многие из них существуют и ныне). Такое строение приобретали виды, приспосабливавшиеся к жизни в густых зарослях, узких пространствах среди камней, к рытью в лесной почве или в песке. Вероятно, предки змей также обитали в подобных укрытиях и отчасти были роющими животными (роющий образ жизни сохраняют и некоторые современные группы змей). Однако представители основного ствола эволюции змей вновь вернулись на поверхность почвы, что было связано с переходом к питанию все более крупной добычей (современные специализированные роющие змеи Scolecophidia являются микрофагами, питающимися муравьями, термитами и т.п.). Однако в отличие от других хищников, питающихся крупной добычей, змеи не разрывают ее на части, а проглатывают целиком. Освоение такого способа питания у змей сопровождалось чрезвычайным усилением кинетизма черепа, в котором самостоятельную подвижность получает каждая половина (левая и правая) и верхней, и нижней челюстей, и даже внутри этих челюстных комплексов многие кости подвижны друг относительно друга. Это позволяет .змее проглатывать целиком добычу, поперечный диаметр которой нередко в несколько раз превышает диаметр головы хищника: челюсти постепенно охватывают тело жертвы, как бы "переступая" по ней, и челюстной аппарат широко растягивается. Древнейшая известная змея - Dinilysia patagonica из верхнемеловых отложений Патагонии - уже обладала основными признаками настоящих змей, хотя некоторые особенности ее черепа сохраняли сходство с признаками ящериц. В позднемеловую эпоху Уже существовали крупные змеи, родственные современным удавам.
Млекопитающие в течение "века динозавров" оставались сравнительно малозаметными членами мезозойских биоценозов. Это были небольшие зверьки, которые вели скрытный образ жизни, Избегая открытых пространств. Вероятно, большинство из них имели сумеречную или ночную активность. Однако и среди млекопитающих происходила адаптивная радиация. Помимо разнообразных насекомоядных и мелких хищников: триконодонтов (Triconodontia), докодонтов (Docodontia), симметродонтов (Sym-metrodontia), пантотериев (Pantotheria) - уже во второй половине юрского периода появились и растительноядные звери, принадлежавшие к отряду многобугорчатых (Multituberculata). Возможно, происхождение многобугорчатых связано с позднетриасовыми харамиидами (Haramiyidae). Название "многобугорчатые" связано со строением зубов этих животных, в некоторых отношениях удивительно сходных с зубами грызунов и приспособленных к перетиранию растительной пищи. Настоящие грызуны возникли уже в кайнозое.
Вероятно, в раннемеловую эпоху произошло расхождение эволюционных стволов сумчатых (Marsupialia) и плацентарных (Р1а-centalia) млекопитающих. Центром эволюции сумчатых была Америка, а плацентарные сформировались в Евразии. Позднее сумчатые через Антарктиду, еще до окончательного расхождения этих частей Гондваны, проникли в Австралию, которая и стала их второй родиной после широкого расселения плацентарных. Плацентарные обладали более высоким развитием головного мозга и более совершенным онтогенезом с длительным развитием зародыша в матке матери и рождением хорошо сформированного детеныша. Вероятно, эти прогрессивные особенности определили успех плацентарных в конкуренции с сумчатыми. Изолированное положение Австралии воспрепятствовало проникновению туда плацентарных и сделало ее уникальным "заповедником" более примитивных групп - сумчатых и однопроходных (Monotremata). Последние стоят особняком среди современных млекопитающих: в ископаемом состоянии они известны лишь из кайнозойских (миоценовых) отложений, а судя по особенностям строения скелета, однопроходные ближе к многобугорчатым, докодонтам и триконодонтам, чем к сумчатым и плацентарным. Последние две группы обнаруживают родственные связи с двумя другими отрядами архаических млекопитающих - симметродонтами и пантотериями. Вероятно, пантотерии были предками эволюционного ствола, в раннем мелу разделившегося на стволы плацентарных и сумчатых. Таким образом, все группы ископаемых и современных млекопитающих распределяются в два основных комплекса (которым разные исследователи придают различный таксономический ранг)-прототерии, или первозвери (Prototheria), включающие однопроходных, докодонтов, триконодонтов и многобугорчатых, и терии, настоящие звери (Theria), - плацентарные, сумчатые, пантотерии и симметродонты. Два этих главных филогенетических ствола млекопитающих обособились друг от друга очень рано, в позднем триасе. Не исключено, что это произошло на уровне организации высших териодонтов, и основные диагностические признаки млекопитающих были приобретены представителями Терий и прототерий независимо друг от друга и параллельно.
(В последние годы получила распространение альтернативная гипотеза, выводящая однопроходных от териевых млекопитающих (возможно, от юрских панэтериев). С однопроходными сближают раннемелового Steropodon, обладающего признаками примитивных териевых.).
Может быть, наиболее значительным по своим последствиям событием, происшедшим примерно в середине "века динозавров" и значительно изменившим облик позднемезозойских биоценозов, было появление покрытосеменных или цветковых растений (Angiospermae). Первые достоверные остатки представителей этой группы высших растений встречаются в нижнемеловых отложениях с абсолютным возрастом около 120 млн лет. Древнейшие покрытосеменные в основном были мелколистными кустарниками или небольшими деревьями. Однако недавно в нижнемеловых отложениях Австралии были также обнаружены остатки травянистых покрытосеменных растений. В начале позднего мела покрытосеменные приобретают самое широкое распространение и огромное разнообразие размеров и форм. Среди них становятся многочисленными крупнолистные растения, в том числе представители современных семейств магнолиевых, лавровых, платановых, кленовых и др.
О происхождении покрытосеменных до сих пор нет единого мнения. Основываясь на наличии некоторых признаков "покрытосемянности" у ряда групп домеловых растений, некоторые ученые предполагают, что покрытосеменные возникли гораздо раньше раннего мела, может быть, даже в конце палеозоя. Однако отсутствие достоверных ископаемых остатков цветковых в отложениях более древних, чем нижнемеловые, говорит в пользу более позднего происхождения этой группы - в поздней юре или в самом начале раннего мела.
Характерные особенности покрытосеменных, в частности образование завязи, защищающей семяпочки (макроспорангии) посредством срастания краев плодолистиков (макроспорофиллов), постепенно развивались у нескольких групп голосеменных растений, которых на этом основании называют иногда "проангио-спермами". При этом, как показал Б.А.Красилов, происходило параллельное и независимое развитие признаков покрытосеменных в разных филетических линиях проангиоспермов (кейтоние-вых - Caytoniales, чекановскиевых - Czekanowskiales, диропалоста - Dirhopalostachyaceae), существовавших в конце юры - начале мела. Как и в других подобных случаях, высказывались предположения о независимом полифилетическом происхождении разных групп покрытосеменных, например двудольных, однодольных и казуариновых, от различных голосеменных.
Так или иначе становление организации ангиоспермов происходило постепенно, и разные их признаки (покрытосемянность развитие цветка, крайняя редукция полового гаплоидного поколения - гаметофита, появление в древесине настоящих сосудов) развивались независимо друг от друга, и в разных филетических линиях опережающее развитие получали различные признаки, что обычно наблюдается при параллельной эволюции. Эти соотношения были зафиксированы правилом независимой эволюции отдельных признаков Г.Осборна и "перекрестом специализаций" по О.Абелю. Поэтому пока трудно указать исходную для покрытосеменных филетическую линию среди проангиоспермов.
Широкое распространение покрытосеменных к середине мелового периода и приобретение ими ведущей роли среди флоры в большинстве наземных биоценозов произошло за относительно небольшие в геологических масштабах сроки порядка 15-20 млн лет, но не носило "взрывного" характера. Неоднократно отмечалось, что это изменение в составе наземной флоры не обнаруживает корреляции с какими бы то ни было заметными в геологической летописи преобразованиями абиотических условий. Расселение и адаптивная радиация покрытосеменных в первой половине мела имели характер постепенного, хотя и относительно быстрого, процесса, обычно наблюдаемого после приобретения новой группой организмов важных приспособлений, обеспечивающих преимущества в борьбе за существование. Среди этих приспособлений свою роль сыграли и покрытосемянность, и связанная с развитием цветка энтомофилия (опыление с помощью насекомых), и высокая устойчивость вегетативных органов к солнечному свету, подмеченная М.И.Голенкиным.
Столь существенное изменение флоры в середине мела неминуемо должно было сказаться на всем облике биосферы, и в первую очередь на биоценозах суши.
Великое вымирание
Последние века мезозойской эры были временем драматических событий, сущность которых пока еще не вполне ясна. Возможно, эти события были в какой-то мере подготовлены только что рассмотренными нами изменениями флоры. Вслед за "победным шествием" покрытосеменных в течение позднего мела вымирают их предшественники - беннеттиты и проангиоспермы, сильно сокращается распространение и разнообразие папоротников саговников. Общий облик флоры позднего мела уже всецело определяется ангиоспермами; из голосеменных сохранили свои позиции лишь хвойные.
Изменения флоры прежде всего сказались на насекомых. На протяжении позднего мела энтомофауна постепенно обновилась: исчез целый ряд архаических семейств и появились группы, существующие и поныне. (См. об этом ст. В.В.Жерихина в кн.: Развитие и смена беспозвоночных на рубеже мезозоя и кайнозоя. Мшанки, членистоногие, иглокожие. - М., 1980.).
Однако в широколиственных и хвойных лесах и на открытых равнинах позднего мела по-прежнему доминировали различные динозавры, в воздухе реяли гигантские летающие ящеры, в морях были обильны разнообразные морские рептилии (плезиозавры и мозазавры, а в позднем мелу, по новым данным, и последние ихтиозавры, морские черепахи), в пресных водоемах - многочисленные крокодилы. В это время существовали самые крупные известные крокодилы - дейнозухи, Deinosuchus, длина черепа которых достигала 2 м, а общая длина - около 16 м. Во второй половине мела, в течение более 45 млн лет после повсеместного распространения покрытосеменных, общий облик фауны оставался в целом прежним, типичным для века динозавров.
Но в конце мелового периода за относительно краткие (в геологических масштабах) сроки произошло вымирание многих групп позвоночных и беспозвоночных животных, наземных, водных и летающих. Вымирают и гигантские формы, и животные мелких размеров, и растительноядные, и хищные. К началу кайнозоя в большинстве регионов вымерли динозавры, летающие ящеры, плезиозавры, мозазавры, последние ихтиозавры, 8 из 10 позднемеловых семейств крокодилов, архаические группы орнитуровых и все энанциорнисовые птицы. Среди беспозвоночных вымирание постигло широко распространенных в юре и мелу двустворчатых моллюсков - рудистов, аммонитов, белемнитов и многих наутилоидных головоногих, вымерли многие виды морских лилий. Значительным было вымирание морского фито- и зоопланктона.
Следует подчеркнуть, что это великое вымирание не сопровождалось одновременным повышением численности и разнообразия видов каких-то других групп. Как и в пермском периоде, Произошло значительное общее обеднение фауны. Только в кайнозое (так сказать, на освободившемся месте) начинается экспансия слабее затронутых вымиранием групп (млекопитающие, птицы, наземные чешуйчатые рептилии, бесхвостые земновод-НЬ1е). С другой стороны, как и во время пермского вымирания, и на рубеже мезозоя и кайнозоя некоторые группы животных как бы "остались в стороне" от происходивших событий: их разнообразие и численность не претерпели существенных изменений. Среди позвоночных это различные группы рыб, хвостатые земноводные, черепахи.
Как и в пермском периоде, великое вымирание в конце мела не носило характера "мировой катастрофы": физико-географические условия на рубеже мела и палеогена не претерпели каких-либо внезапных и резких изменений. С достаточной достоверностью можно говорить лишь о некотором похолодании климата к концу мелового периода, происходившем постепенно и отразившемся на растительных сообществах: в районах, где удается проследить всю последовательность отложений на рубеже мела и палеогена, обнаруживается постепенное замещение теплолюбивых видов растений видами, приспособленными к более прохладному климату (например, в Северной Америке произошла замена субтропических лесов лесами умеренного пояса). Однако в тропическом поясе существенных изменений растительности и, вероятно, климата не произошло.
Процесс вымирания был кратким лишь в геологическом смысле: он продолжался в течение миллионов лет, когда постепенно угасали вымирающие филетические линии. Остается неясным, в какой мере эти процессы происходили одновременно на разных континентах и в разных океанах и морях. Например, по данным Р.Слоуна, на западе Северной Америки динозавры (трицератопсы, тероподы и др.) существовали еще несколько миллионов лет в начале палеогена, после их вымирания в других местах. Аналогичные данные имеются также для Индии и некоторых других регионов. Но так или иначе итог был один на всем земном шаре, что, собственно, и придает этому вымиранию, как и другим массовым вымираниям, загадочный характер.
В гипотезах о причинах вымирания в конце мела нет недостатка; эта волнующая проблема всегда привлекала внимание исследователей. Достаточно подробный обзор многочисленных гипотез потребовал бы отдельной книги и далеко выходит за рамки возможностей данной работы. Мы вынуждены здесь ограничиться рассмотрением лишь некоторых основных концепций.
Поскольку подвергшиеся вымиранию группы организмов в конечном итоге исчезли повсеместно, многие ученые предполагали, что причины таких явлений должны были иметь характер всемирных катастроф. Первая из катастрофических гипотез была выдвинута еще Ж.Кювье, который считал причиной великог вымирания в конце мела вулканическую деятельность, связан ную с Альпийской фазой горообразования. Несомненно, усиление вулканизма оказывает влияние на органический мир не толь ко непосредственно (излияние лав, покрывающих большие территории, которые становятся на длительное время непригодными для жизни, и другие губительные для организмов факторы s вулканических извержений), но также косвенно. Происходят значительные изменения ландшафта; в атмосферу выбрасываются огромные количества вулканической пыли и углекислого газа, снижающие прозрачность воздуха; все это сказывается на климате. Однако в фанерозое проявления вулканизма всегда имели локальный характер, и непосредственный эффект вулканической деятельности мог сказываться лишь на относительно небольшой части земной поверхности. С другой стороны, горообразовательные процессы, сопровождавшиеся вулканизмом, происходили в разных областях земного шара и в юрское время, и в меловом периоде задолго до эпохи великого вымирания, не приводя к катастрофическим последствиям для динозавров и их современников. Поэтому вулканизм сам по себе не мог быть причиной великого вымирания, хотя и играл, вероятно, существенную роль в изменениях климата.
В последнее десятилетие оживленную дискуссию вызвала гипотеза Л. и У. Альварецов, согласно которой причиной катастрофы, вызвавшей великое вымирание на рубеже мела и палеогена, было столкновение с Землей одного или нескольких астероидов. В более новом варианте этой гипотезы, получившей название "импактной" (от англ. impact - удар, толчок), предполагается столкновение Земли не с астероидом, а с гигантской кометой или с несколькими кометами. В качестве доказательств этой космической катастрофы указывают на повышенное (примерно в 30 раз) содержание иридия (которому приписывают астероидное или кометное происхождение) в слое глинистых отложений на границе мела и палеогена, наличие в пограничных отложениях застывших капель расплава, ударно-метаморфизированных кристаллов кварца, а также большого количества частиц угольной сажи, которые, как предполагают, образовались при ураганных пожарах, возникших после космической катастрофы.
При падении на Землю крупных астероидов должны были возникнуть гигантские кратеры (обычно диаметр кратера примерно в 10 раз превышает диаметр упавшего метеорита). Кратеров "нужного размера", образовавшихся в конце мела, на Земле до сих пор Не обнаружено; самый крупный известный ныне кратер Чиксхулуб, расположенный на севере Юкатана в Мексике и образовавщийся около 65 млн лет назад, имеет диаметр около 180 км. сторонники импактной гипотезы допускают, что астероид упал в океан.
Механизм воздействия такого космического удара на биосферу Земли понимается различными учеными по-разному. Некоторые исследователи полагают, что массовое вымирание было вызвано резким повышением температуры воздуха и океана (при возможном отравлении вод цианистыми соединениями) и возникновением ураганных пожаров на суше. Другие ученые (среди них авторы импактной гипотезы Альварецы) считают более вероятным развитие событий по так называемому "сценарию ядерной зимы", разработанному при анализе вероятных последствий термоядерной войны. Засорение атмосферы метеоритной пылью и частицами сажи от ураганных пожаров должно было привести к значительному снижению прозрачности воздуха, в результате которого должна была значительно понизиться температура нижних слоев атмосферы, океана и почвы и резко сократиться фотосинтез. Это могло привести к разрушению биоценозов и массовому вымиранию растений и животных как на суше, так и в океане.
Некоторые палеоклиматологические данные действительно указывают на понижение среднегодовых температур на 5-6°С к концу мелового периода, проявившееся особенно заметно в приполярных и средних широтах, где субтропическая растительность сменилась лесами, характерными для умеренного климата. Однако в тропическом поясе значительных изменений температурного режима не произошло, а общий ход этих климатических изменений отнюдь не соответствует "сценарию ядерной зимы" и импактной гипотезе, так как эти процессы развивались постепенно в течение нескольких миллионов лет.
Проведенные в дальнейшем широкие исследования отложений на границе мела и палеогена показали, что в некоторых районах слои, содержащие остатки сажи, расположены значительно ниже слоя, обогащенного иридием и, очевидно, возникли значительно раньше последнего. Более того, не подтвердилось и единство "иридиевого слоя" - в разных областях соответствующие отложения имеют разный возраст и не могли возникнуть в результате одной космической катастрофы. Наконец, все аномальные явления, используемые как аргументы в пользу импактной гипотезы, могли возникнуть под воздействием чисто "земных" причин - например в результате вулканической деятельности, которая, как мы уже отмечали, значительно усилилась к концу мела в Индии, Северной Америке и некоторых других районах. Кроме того, и сам процесс вымирания организмов в конце мела был, как уже подчеркивалось, в достаточной степени растянут во времени (постепенное обеднение фауны происходило в течение более 7 млн лет). Этот процесс не был ни внезапным, ни строго одновременным по всей Земле и для всех групп организмов, а само вымирание началось задолго до времени образования иридиевого слоя и было отнюдь не всеобщим, а выборочным, причем некоторые организмов оказались им практически не затронутыми. Нужно принять во внимание, что вообще на Земле известны и большие кратеры, возникшие, вероятно, при падении болидов (например кратер Монтана на атлантическом шельфе у берегов Канады диаметром около 45 км, образовавшийся в конце раннего эоцена, или среднеолигоценовый кратер Попигай на Таймыре - диаметром около 100 км. Однако падение этих крупных небесных тел не привело к заметным изменениям в биосфере и не имело своими последствиями Процессов массового вымирания.
Таким образом, общая совокупность имеющиеся ныне данных говорит в целом против катастрофических гипотез вымирания в конце мела (так же, как и в другие геологические эпохи).
Высказывались предположения о связи вымирания динозавров с изменениями биотических факторов, в качестве которых называли, в частности, конкуренцию со стороны млекопитающих или преобразования флоры связанные с широким распространением покрытосеменных в середине мела. Однако млекопитающие возникли еще в позднем триасе и на протяжении примерно 130 млн лет, прошедших до конца мезозоя, оставались сравнительно малозаметной и малозначительной группой животных.
Существует гипотеза, что важную роль в вымирании динозавров могло сыграть преобладание покрытосеменных в позднемеловых растительных сообществах, поскольку Покрытосеменные в биохимическом отношении существенно отличаются от тех групп растений, которые служили пищей растительноядным животным до середины мела. Однако динозавры сосуществовали с ангиоспермами около 70 млн лет, и фауна динозавров, включавшая многочисленные и разнообразные растительноядные виды, процветала не менее 45 млн лет после широкий экспансии покрытосеменных растений.
Не следует также забывать и о других группах животных (в частности, морских) вымерших в конце мезозоя: очевидно, указанные биотические факторы сами по себе не могут объяснить вымирания плезиозавров мозазавров, русистов, морских лилий и т.д. Поскольку вымирание коснулось одних групп животных и почти или совсем не затронуло другие, ключ к пониманию событий, происходивших на рубеже мезозоя и кайнозоя, видимо, следует искать не только в изменениях внешних факторов, но и особенностях организации и биологии подвергшихся вымиранию животных.
Особая трудность заключается в том, что в конце мела вымерли группы животных значительно отличавшиеся друг от друга экологически и обитавшие в разной среде (наземные, амфибиотические, пресноводные и морские). И пока остается неясным, было ли вызвано вымирание столь различных животных, как всевозможные динозавры, летающие ящеры, аммониты, рудисты и т.д. какой-то одной внешней причиной (хотя бы косвенно) или же одновременным действием разных факторов, не связанных друг с другом причинно.
Поскольку динозавры более всего привлекали внимание, в большинстве гипотез обсуждается в первую очередь вымирание именно этих животных. В поисках "слабого места" в организации динозавров, которое могло бы способствовать их вымиранию при определенных <изменениях внешних условий, многие ученые останавливались на особенностях теплообмена этих рептилий. Как уже говорилось, вероятнее всего, динозавры gоставались физиологически пойкилотермными (холоднокровными) животными, как и все современные пресмыкающиеся. Однако, используя гелиотермию, динозавры (особенно крупные формы) в условиях ровного и теплого климата юры и мела могли поддерживать температуру своего тела на практически постоянном, оптимальном для функций организма уровне. При отсутствии значительных сезонных изменений климата, подобных, например, современным в средних широтах, у динозавров не могло выработаться каких-либо физиологических или поведенческих механизмов для ?успешной зимовки.
В поисках тех изменений внешних условий, которые вызвали вымирание динозавров, Д.Аксельрод и Г.Бейли вновь обратились к процессам горообразования и вулканизма, происходившим в конце мела, последствия которых могли иметь важное значение, хотя и не носили ?характера катастрофы. Мезозой был в целом эрой низкого стояния материков. Альпийская фаза горообразования, постепенно развивавшаяся в юре и мелу, сопровождалась значительным общим поднятием суши к концу мезозоя. Результатом этого, а также снижения прозрачности атмосферы вследствие вулканической деятельности было постепенное понижение среднегодовой температуры в течение 20 млн лет примерно на 5 °С. Но, вероятно, еще более важным фактором было возрастание неравномерности температурных условий в умеренном поясе с развитием все более резко выраженной сезонности климата и значительным возрастанием перепада между максимальными и минимальными температурами. Об этом, в частности, говорят изменения растительности: появление в позднемеловое время в средних широтах листопадной флоры вместо субтропических лесов, а в начале палеогена и листопадная флора была в той или иной степени замещена более холодолюбивыми хвойными лесами. К такому направлению изменений климата динозавры были приспособлены значительно хуже, чем млекопитающие и птицы, у которых уже сформировалась настоящая гомойотермия, а также чем рептилии, которые могли переживать неблагоприятные сезоны года в неактивном состоянии (ящерицы, змеи, черепахи). Последний путь адаптации для динозавров был затруднен в связи крупными размерами (которые были столь выигрышны в энергетическом отношении на протяжении юры и мела), а также спецификой их теплообмена: не будучи гомойотермными, динозавры были приспособлены к практически постоянным оптимальным температурам. Отметим, что, говоря здесь о крупных размерах, мы не имеем в виду гигантские формы, а вообще крупные - более 1 м, а именно таковы были мелкие динозавры. Отметим также, что ныне в умеренном поясе пресмыкающиеся представлены лишь мелкими видами, как правило, менее 1 м, которые могут успешно выдержать зимовку в различных убежищах. Все крупные современные виды рептилий (крокодилы, крупные виды змей, ящериц и черепах) - тропические животные.
С этой гипотезой можно согласовать и наблюдения французских палеонтологов об аномалиях скорлупы яиц, часто встречающихся в ископаемых кладках динозавров из верхнемеловых отложений Прованса. Высказывалось предположение, что эти аномалии были результатом повторных прижизненных приостановок процесса формирования скорлупы во время развития яиц в яйцеводах самок динозавров, которые могли быть вызваны похолоданием.
Достоинствами разбираемой гипотезы являются, во-первых, согласование ряда достаточно разнообразных и достоверных данных, во-вторых, признание постепенности происходивших на Земле изменений и самого процесса вымирания. Однако и эта гипотеза оставляет открытыми ряд серьезных вопросов: почему динозавры и летающие ящеры не сохранились в тропиках, где даже при некотором понижении средней температуры сохранялись в целом теплые и ровные климаты на протяжении всего фанерозоя (и где выжили, например, крокодилы, которые, вероятно, были физиологически близки к динозаврам); почему повсеместно вымерли морские рептилии и ряд других групп морских организмов, ведь в океане, особенно в низких широтах, не могло возникнуть температурной неравномерности, сравнимой с таковой на суше.
Возможно, удовлетворительный ответ на последний вопрос дает гипотеза французского палеонтолога Л.Гинзбурга, также опирающаяся на геологический факт поднятия материков к концу мела, с которым была связана значительная морская регрессия. В ходе этой регрессии уровень моря к концу мелового периода по сравнению с его серединой понизился на 180-200 м. При этом акватория эпиконтинентальных морей (т.е. покрытых морем частей континентальных платформ) уменьшилась примерно в 50 раз. Меловые теплые эпиконтинентальные моря являются наиболее благоприятной для жизни зоной мирового океана, наиболее обильные видами организмов. Вероятно, столь значительное сокращение их акватории не могло не сказаться на самых различных группах морских организмов. Однако при этом остается непонятной выборочность вымирания: почему вымерли морские рептилии, многие группы моллюсков и др., но практически не пострадали, например, костистые рыбы.
(Еще одной проблемой данной теории является относительная стремительность вымирания по сравнению с постепенностью геологических и климатических изменений. Однако это можно объяснить тем, что в конце мела могло произойти "исчерпание запаса прочности" и переход "границы устойчивости" биоценозов, за которым последовал их стремительный распад).
Таким образом, вымирание как наземных, так и морских групп организмов в конце мела могло быть в конечном итоге следствием горообразовательных процессов и поднятия континентов во второй половине мезозоя. Существенное воздействие на климат в направлении общего похолодания мог оказать усиленный вулканизм, которым отмечен конец мелового периода, например вулканическая деятельность в Индии продолжалась не менее 0,5 млн лет. Эти постепенно происходившие преобразования условий медленно, но неуклонно нарастали, приводя к изменениям морских течений и преобладающих направлений атмосферных потоков. Воздействие этих изменений на фауну и флору было чрезвычайно сложным и многообразным, как непосредственным, так и косвенным.
Массовые вымирания разного масштаба происходили в фанерозое неоднократно, крупнейшие из них - в раннем кембрии, позднем ордовике, поздней перми и в конце мелового периода. Было сделано немало попыток уловить какую-то периодичность массовых вымираний, однако интервалы между ними очень варьируют, составляя 20-60 млн лет. Массовые вымирания меньшего масштаба имели место в триасе и в первой половине юры.
Вообще существует плавный и постепенный переход в масштабах вымирания от происходившего во все эпохи фонового до массовых вымираний, причем последние - при всей их эффектности - охватывают лишь около 5% всех явлений вымирания в истории Земли, остальные 95% приходятся на менее заметное фоновое вымирание.
Общая совокупность данных говорит против катастрофических причин массовых вымираний. Вероятно, в большинстве случаев масштабы и специфика процессов вымирания в конкретные эпохи истории биосферы более определялись состоянием экосистем, чем изменениями абиотических факторов. Последние же играют роль стрессового механизма, "проверяющего на прочность" механизмы устойчивости биоценозов, приводя к упадку и вымиранию некоторых видов организмов. Как уже упоминалось, устойчивость биоценозов имеет свои пределы: если нарушения структуры биоценоза выходят за эти пределы, начинается распад всей экосистемы. При этом нарушаются сложившиеся прежде пути передачи органических веществ и энергии в биосфере. Тогда вымиранию подвергаются новые виды, которые сами по себе еще не были затронуты непосредственно изменениями абиотических факторов. Этот процесс будет нарастать лавинообразно, пока тем или иным путем не будет достигнуто новое равновесие между биосинтезом и деструкцией органических веществ, между видами растений, растительноядных животных, хищников и микроорганизмов, т.е. пока не сформируются новые устойчивые и способные к саморегуляции экосистемы - биоценозы.
Можно сказать, что великое вымирание представляет собой рассматриваемый в далекой исторической перспективе внешний результат сложных перестроек биоценозов и биосферы в целом, которые могли начаться задолго до рубежа мезозоя и кайнозоя и, постепенно нарастая, достигли к этому времени своей кульминации.
Тропические и субтропические флоры преобладали в начале палеогена в Северной и Южной Америках, Африке и Южной Азии. Даже в Гренландии и на Шпицбергене была разнообразная растительность, свойственная теплому умеренному климату: секвойи, болотные кипарисы, гинкго, буки, дубы, платаны, клены, магнолии, липы, березы и др. Любопытно развитие у многих высокоширотных растений гигантских листьев до 30-40 см. Климат в этих местах был в палеогене близок к современному климату Южной Франции или Калифорнии.
А на обширных территориях Центральной Азии с начала палеогена намечается тенденция к аридизации (повышениие засушливости) климата, которая, постепенно усиливаясь, прогрессировала на протяжении всего этого периода и неогена. Это привело к усыханию обильных здесь в позднемеловое время внутриконтинентальных пресноводных бассейнов, к постепенному обезлесиванию ландшафта с развитием к концу палеогена (олигоценовая эпоха) открытых пространств типа лесостепей, степей и саванн. Вместо влаголюбивой сочной лесной растительности получают преобладание злаки (Gramineae), жесткие и относительно сухие листья которых хорошо защищены от обезвоживания. Местами в саваннах возвышались отдельные группы деревьев с высоко расположенными кронами. Более густая древесная и кустарниковая растительность сохранялась по долинам рек. Аналогичные открытые сухие ландшафты получили в начале неогена широкое распространение и на других континентах, но наиболее засушливыми в кайнозое оставались районы Центральной Азии.
Во второй половине палеогена началось постепенное глобальное похолодание климата, которое усилилось в неогене. Оно завершилось в плейстоценовой эпохе четвертичного периода развитием нового великого оледенения. Процесс похолодания климата был очень медленным и постепенным. Все более отчетливо проявлялась климатическая зональность. Климатические изменения легко прослеживаются по преобразованиям флоры. Теплолюбивая флора в Евразии и Северной Америке постепенно отступала к югу, сменяясь в умеренной зоне листопадными и хвойными лесами. Изменения климата представляют собой внешний итог сложнейшего взаимодействия множества различных факторов (космических, географических и атмосферных), среди которых важную роль играют направления морских течений и характер атмосферной циркуляции. Эти последние зависят, в свою очередь, от изменений рельефа и положения континентов.
Согласно представлениям теории тектоники плит, в кайнозое продолжалось движение континентов, которое постепенно привело к современной картине их размещения. От Северо-Американского материка отделилась Гренландия. Смещения Африки и Индии почти совсем закрыли мезозойский океан Тетис. На его месте ныне существуют Средиземное, Черное и Каспийское моря, а на востоке - моря Зондского архипелага, однако среди ученых еще нет единства мнений о преемственности этих бассейнов по отношению к Тетису. Индостанская плита, смещаясь к северо-востоку, наконец, соединилась с Азией. Австралия, вероятно, лишь в эоцене обособилась от Антарктиды и стала быстро (в геологическом смысле) продвигаться к северу, приближаясь к экватору (см. рис. 60).
Смыкание континентальных плит сопровождалось развитием горной складчатости в зонах их контакта. В кайнозое сформировался Альпийский складчатый пояс. В палеогене и неогене поднялись горные цепи Атласа, Пиренеев, Альп, Апеннин, Балкан, Карпат, Крыма, Кавказа, Копетдага, Памира и Гималаев, завершилось формирование Анд, Кордильер и ряда других горных систем. Неоген характеризуется высоким положением материков И наибольшей степенью осушения их территорий за весь фанерозой, с этим связано преобладание открытых сухих ландшафтов в неогене.
Северный полюс, находившийся в мезозое в северной части Тихого океана и на близких к океанскому побережью областях Северо-Восточной Азии и Аляски, в кайнозое оказался в центре почти замкнутого бассейна Северного Ледовитого океана, который сам сформировался лишь в кайнозое. Этот бассейн в значительной своей части закрыт для доступа меридиональных морских течений, которые могли бы обеспечить более равномерное распределение температур в мировом океане и на поверхности Земли в целом. Благодаря этому Северный Ледовитый океан при расположении в нем Северного полюса оказался аналогичным континенту в смысле возникновения там огромного "очага холода", "мешка льдов". Возможно, это было важнейшей из причин, вызвавших охлаждение климата в кайнозое, которое завершилось великим оледенением.
Первый обширный ледниковый щит сформировался в Антарктиде в миоценовую эпоху неогена около 20 млн лет назад. До этого в течение палеогена в Антарктиде сохранялся умеренный прохладный климат, и ледники имелись лишь в горах. Развитие ледникового щита Антарктиды вместе с постепенно расширявшейся зоной плавучих льдов в Северном Ледовитом океане значительно увеличили отражение солнечных лучей (альбедо) земной поверхности, что, в свою очередь, стало фактором, способствовавшим дальнейшему охлаждению климата. Климатическое равновесие все более нарушалось в сторону увеличения контрастов климатических зон. Наконец, в плейстоценовой эпохе четвертичного периода (около 3 млн, а по некоторым данным, даже 1-5 млн лет назад) началось великое оледенение, сопровождавшееся развитием гигантских ледниковых щитов в Европе и Северной Америке. Оледенение прерывалось несколько раз резко наступавшими потеплениями (межледниковые эпохи, или интерстадиалы), во время которых ледниковый покров на континентах Европы и Северной Америки стаивал практически полностью. Затем происходило новое понижение температур с последующим оледенением. Период этих изменений составлял в среднем около 100 тыс. лет. Такой характер процесса изменений климата говорит за то, что развитие оледенения в четвертичном периоде носит пороговый характер: при незначительном нарушении неустойчивого температурного баланса, когда средние температуры падают ниже некоторого порогового уровня, начинает быстро прогрессировать оледенение и наоборот. Среди причин, влиявших на колебания климата в ту или другую сторону, указывали на вулканизм (снижение прозрачности атмосферы в результате ее засорения вулканическим пеплом), а также на колебания наклона экватора Земли к плоскости ее орбиты и изменения параметров самой орбиты под действием притяжения других планет.
Максимальное из плейстоценовых оледенений (около 250 тыс. лет назад) покрывало территорию около 45 млн км (т. е. примерно 30% суши), ледниковый шит в Европе достигал широты 48° 30', а в Северной Америке - 37°. Объем льдов на суше в это время составлял около 55 млн км3. Соответственно уровень мирового океана в эпохи оледенений значительно падал (на 85-120 м), и возникали сухопутные связи между континентами и островами, разобщенными ныне проливами. Например, Берингийская суша связывала в эпохи оледенений Аляску и Чукотку, Британские острова соединялись с Европой и т.д. Последнее оледенение завершилось около 10 тыс. лет назад.
Развитие жизни в палеогене
В начале палеоценовой эпохи фауна млекопитающих оставалась, по существу, той же, что и в позднемеловое время. В ней были представлены лишь группы, возникшие еще в мезозое: растительноядные многобугорчатые, внешне напоминавшие грызунов, но, возможно, родственные прототериям - однопроходным, а также архаические представители сумчатых и плацентарных, питавшиеся насекомыми и другой мелкой добычей. Для всех архаических млекопитающих были характерны такие примитивные особенности, как относительно небольшой мозг, простые треугольные зубы (за исключением многобугорчатых), пятипалые конечности, опиравшиеся при передвижении на всю кисть и стопу (плантиградность).
Предки млекопитающих - синапсидные рептилии-териодонты вымерли еще в позднем триасе. Однако в 1992 г. было опубликовано сенсационное сообщение о находке в позднепалеоценовых отложениях Альберты в Канаде фрагмента нижней челюсти, ряд особенностей которой очень сходен с состоянием цинодонтов. Если данные об этом палеоценовом цинодонте, получившем название Chronoperates, подтвердятся, придется заключить, что одна из филетических линий териодонтов просуществовала весь мезозой и перешла в кайнозой, пережив (вместе с некоторыми линиями динозавров) рубеж мела и палеогена. (Однако хроноператес может оказаться не цинодонтом, а представителем одной из очень архаических мезозойских групп млекопитающих - симметродонтов, сохранившим дополнительные кости в нижней челюсти).
К середине палеоценовой эпохи разнообразие млекопитающих значительно возросло и настолько, что можно предположить расхождение их предковых линий еще до конца мела. Но основная адаптивная радиация плацентарных и сумчатых происходила в палеоцене и эоцене, когда сложились все основные отряды кайнозойских млекопитающих. (При обсуждении филогении различных отрядов плацентарных в этой главе не будут рассматриваться приматы, эволюционная история которых разбирается в следующей главе).
От примитивных насекомоядных плацентарных возникли всеядные (использующие как животную, так и растительную пищу), а затем и настоящие растительноядные формы. Растительноядность у некоторых групп плацентарных развилась в палеоцене. Начало этого направления приспособительной эволюции было представлено архаическими копытными - кондиляртрами (Condylarthra), древнейшие представители которых известны из верхнемеловых отложений Южной Америки. Это были сравнительно небольшие животные размером с зайца, более поздние представители достигали длины тела около 180 см. Внешне они больше походили на хищников, чем на копытных, и сохраняли большие клыки. Однако их коренные зубы имели хотя и низкие, но широкие коронки, пригодные для перетирания не особенно жесткой растительной пищи. Ключицы были утрачены, что говорит о приспособлении локомоторного аппарата к быстрым "монотонным" движениям конечностей при беге, а концевые фаланги пятипалых кисти и стопы были защищены небольшими копытцами (по крайней мере у некоторых форм, подобных Phenacodus). Вероятно, при беге конечности этих животных опирались уже не на всю стопу, а лишь на пальцы. Возможно, примитивные кондиляртры были предками других групп копытных.
Среди этих последних уже в позднем палеоцене и эоцене появились более специализированные, крупные и причудливые формы. Характерны в этом отношении диноцераты (Dinocerata - "страшнорогие"), которые были самыми крупными наземными млекопитающими эоценовой эпохи, достигая размеров современных носорогов. Относительно короткие и толстые пятипалые конечности этих массивных животных несли копыта. Череп у некоторых форм (например, уинтатерия - Uintatherium, рис.80) имел рогоподобные костные выросты и кинжалообразные острые клыки. Вероятно, эти крупные животные были хорошо защищены от нападений современных им хищников. Однако диноцераты вымерли уже к концу эоцена. Скорее всего, их вымирание связано с конкуренцией с более прогрессивными группами копытных, которую диноцераты проиграли благодаря общему консерватизму своей организации, в частности сохранению относительно очень небольшого головного мозга.
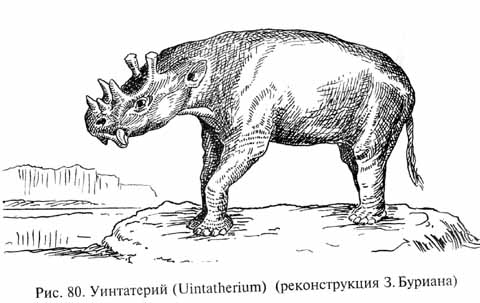
В палеоцене и эоцене появились такие прогрессивные группы растительноядных млекопитающих, как непарнокопытные (Perissodactyla), парнокопытные (Artiodactyla), грызуны (Rodentia), зайцеобразные (Lagomorpha) и ряд других. Их быстрая адаптивная радиация привела к вымиранию архаических групп растительноядных зверей: кондиляртр, диноцерат и родственных последним пантодонтов (Pantodonta) к концу эоцена, многобугорчатых в раннем олигоцене.
Своеобразным убежищем для некоторых примитивных групп млекопитающих стала Южная Америка, которая к концу раннего эоцена обособилась от Северной Америки и оставалась изолированной до плиоцена. В палеогене и неогене там существовал удивительный мир причудливых форм, сохранявших общий примитивный уровень организации; их судьбу мы рассмотрим несколько ниже. Среди высших копытных раньше других начали адаптивную радиацию непарнокопытные, которые уже в эоцене были представлены разнообразными формами. Центром эволюции этого отряда была, по-видимому, Северная Америка, откуда известны наиболее ранние и примитивные представители различных семейств, как доживших до современности (лошади, тапиры, носороги), так и вымерших (титанотерии, халикотерии и др.). Непарнокопытные пережили свой расцвет в палеогене, причем их эволюционная история стала одной из самых ярких страниц в кайнозойской палеонтологической летописи.
Примитивные эоценовые непарнокопытные были небольшими животными, у которых уже наметились изменения конечностей, связанные с их приспособлением к быстрому бегу, в частности, удлинение средних пальцев и элементов пястья и плюсны при одновременной редукции боковых лучей в кисти и стопе. У эоценовых форм было по 4 пальца на передних и по 3 на задних конечностях (такое состояние сохранилось у современных тапиров). Эволюционная тенденция к развитию тонких удлиненных конечностей как приспособление к убыстрению бега наиболее ярко выражена в филогенезе лошадей (Equidae, рис. 81), который является одним из наиболее изученных. Близко к основанию филогенетического ствола лошадиных стоит раннеэоценовый эогиппус (Eohipipus). Это животное размерами и пропорциями тела напоминало небольшую собаку (длина тела составляла около 50 см). Вероятно, эогиппусы жили в лесных зарослях и питались сочной и мягкой растительностью. В филогенезе лошадей происходило постепенное увеличение размеров тела, сопровождавшееся коррелятивным удлинением морды. Эти тенденции проявились уже у позднеэоце-новых (Orohippus, Epihippus) и олигоценовых (Ме8огирри5, Miohip-tous) лошадей. Последние достигали размеров более 1 м и сохранили лишь по 3 пальца на всех конечностях. Они оставались лесными животными. Дальнейшая эволюция лошадей связана с приспособлением к жизни в открытых местообитаниях, широко распространившихся в неогене, и будет рассмотрена нами ниже. Тенденция к увеличению размеров тела еще сильнее была выражена у титанотериев и носорогов, эволюция которых также начиналась с небольших легко построенных эоценовых форм. Но уже в олигоцене эти непарнокопытные были представлены огромными животными, с размерами которых могут сравниться среди наземных млекопитающих лишь неогеновые хоботные.


Титанотерии или бронтотерии (Bronthotheriidae, рис. 82) достигали в холке высоты около 2,5 м при длине тела до 4,5 м. Их длинный и низкий череп с поразительно маленькой мозговой полостью имел на носовых или на лобных костях большие рогоподобные выступы - парные или в виде толстого срединного "рога", раздвоенного у вершины. Титанотерии обитали в лесах, на влажных лугах вблизи водоемов, а некоторые виды вели, вероятно, полуводный образ жизни. Питались они сочной лесной или водной растительностью. Титанотерии вымерли уже в раннем олигоцене. Их заместили представители другой группы непарнокопытных - носороги (Rhinocerotoidea), пять видов которых дожили до нашего времени. В олигоцене же и в неогене это была процветающая и очень разнообразная группа.
В отличие от современных носорогов, несущих на морде один или два рога (представляющих собой, как это ни кажется странным, пучки сросшихся волос), многие древние носороги были безрогими (название этой группе было дано по современным формам). Среди них имелись относительно легкие, так называемые "бегающие носороги" - гиракодонты (Hyracodontidae), полуводные, внешне несколько напоминавшие бегемотов аминодонты (Amynodontidae), но особенно замечательны были гигантские носороги (Indricotheriidae), распространенные в Азии в олигоцене в начале миоцена. Это были крупнейшие наземные млекопитающие, превосходившие даже крупных хоботных и уступавшие размерам и массе тела лишь завроподам: длина их тела достигала 7-8 м при высоте в холке до 5,5 м. Довольно длинная и мощная шея могла поднять огромную голову (череп имел до 1,2 м в длину) на высоту около 8 м над землей (рис. 83). Гигантские носороги обитали в открытых ландшафтах, лесостепях и саваннах, которые были широко распространены в Азии уже в олигоцене. Вероятно, эти животные питались листвой деревьев с высокими кронами. Дальнейшая аридизация азиатских степей в неогене привела к их вымиранию.

Заслуживает упоминания еще одна интересная группа непарнокопытных, просуществовавшая от позднего эоцена до раннего плейстоцена - халикотерии (Chalicotheriidae). По размерам тела и форме головы эти животные несколько напоминали лошадей, но их трех-четырехпалые конечности были снабжены не копытами, а большими изогнутыми когтями, похожими на когти гигантских ленивцев (см. ниже) и муравьедов. Любопытно, что создатель палеонтологии Ж.Кювье в свое время описал когтевую фалангу халикотерия как принадлежащую муравьеду, будучи уверен в существовании жестких и однозначных корреляций между различными частями организма. Среди палеонтологов нет единства мнений По поводу образа жизни этих странных существ. Возможно, халикотерии, подобно гигантским ленивцам, питались молодыми побегами и листьями деревьев, ветви которых они подгибали и удерживали когтистыми лапами. По другому предположению, халикотерии использовали когти для выкапывания из почвы питательных Клубней и корневищ растений.
Адаптивная радиация парнокопытных началась несколько по-3ДНее, чем непарнокопытных, однако в олигоцене уже существовали примитивные представители современных семейств свиных (Suidae), верблюдов (Camelidae), оленьков (Tragulidae) и ряд вымерших групп. Анализируя филогенетические преобразования скелета конечностей у палеогеновых парнокопытных, В.О.Ковалевский разработал концепцию об адаптивных и инадаптивных изменениях органов в эволюции (см. ч. II, гл. 5).
Эволюция парно- и непарнокопытных в палеогене происходила на территориях Северной Америки и Евразии, связанных друг с другом Берингийским сухопутным "мостом".
В Африке сформировалась в палеогене своя оригинальная фауна растительноядных млекопитающих. В ее состав входили разнообразные животные: крупные хоботные (Proboscidea); небольшие даманы, или жиряки (Hyracoidea), напоминавшие своим внешним обликом и образом жизни некоторых грызунов; вымершие еще в палеогене эмбритоподы (Embrithopoda), внешне сходные с носорогами и титанотериями; наконец, перешедшие к полуводному и водному образу жизни десмостилы (Desmostylia), несколько напоминавшие давно вымерших плакодонтов, и сиреновые (Sirenia), у которых развился хвостовой плавник, передние конечности преобразовались в ласты, а задние редуцировались. При всем внешнем несходстве этих групп животных друг с другом ряд характерных общих признаков (увеличение одной пары резцов, строение коренных зубов, редукция ключиц, наличие копыт, положение млечных желез и др.) позволяют предполагать их общее происхождение. Некоторые авторы считают, что их предками могли быть примитивные кондиляртры (однако в Африке, где возникли все эти группы, ископаемых остатков кондиляртр пока не обнаружено).
Древнейшим представителем хоботных является среднеэоце-новый нумидотерий (Numidotherium). Более известен меритерий (Moeritherium) из верхнего эоцена - нижнего олигоцена, который размерами и внешним обликом, вероятно, напоминал тапира и имел зачаточный хобот. Вторые пары резцов в верхней и нижней челюстях были сильно увеличены; их дальнейшая гипертрофия у более поздних хоботных привела к формированию бивней.
В раннем олигоцене появились древнейшие мастодонты - более высокоразвитые хоботные, внешне уже напоминавшие настоящих слонов. Название "мастодонт" означает буквально "сосце-зуб", что связано с характерной формой увеличенных коренных зубов, коронки которых имели сосцевидные бугорки. По паре гипертрофированных резцов-бивней было как в верхней, так и в нижней челюсти. Морда у мастодонтов оставалась удлиненной, поэтому хобот, представляющий собой результат разрастания верхней губы и носа, был относительно коротким. Мастодонты были много крупнее меритерия, некоторые виды уже в олигоцене достигали размеров современных слонов. Расцвет хоботных наступил уже в неогене.
Сирены, в современной фауне представленные лишь двумя родами (ламантином - Trichechus и дюгонем - Dugong), обособились, возможно, еще в палеоцене в связи с приспособлением к постоянной жизни в воде. Сирены возникли в прибрежных водах теплых морей еще существовавшего тогда океана Тетис и были широко распространены уже в эоцене. В отличие от сирен десмостилы, обитавшие в олигоцене и миоцене по берегам Тихого океана, сохранили обе пары конечностей и могли передвигаться по суше, хотя и очень неуклюже, так как их конечности были широко расставлены в стороны. Характерно строение их массивных коренных зубов, образованных 4-8 дентиновыми трубками, покрытыми тонким слоем эмали, - возможно, эти зубы служили для раздавливания раковин моллюсков.
В палеогеновых морях помимо сирен появились и другие млекопитающие, которые заняли освободившиеся экологические ниши морских рептилий. Это были древнейшие китообразные (Cetacea), остатки которых обнаружены в среднеэоценовых отложениях Северной Африки, т. е. также в области эпиконтиненталь-ных морей океана Тетис. Пропорции тела архаических китов (Archaeoceti) значительно отличались от таковых у современных групп: туловище было длинное и относительно тонкое (у наиболее крупной формы, зейглодона - Zeuglodon - до 21 м в длину при диаметре не более 2,5 м). Длинные узкие челюсти большого, до 1,2 м в длину, и низкого черепа были усажены впереди коническими, а сзади сжатыми с боков острыми зубами. Короткие передние конечности располагались близко к голове, а задние были сильно редуцированы (полностью развитые задние конечности сохранялись у эоценовых Ambulocetus и Basilosaurus). По всей вероятности, китообразные возникли от каких-то примитивных наземных хищников, перешедших к рыбоядности и постепенно развивавших приспособления к амфибиотической, а потом и к водной жизни. Примитивные представители современных групп китообразных: зубатых (Odontoceti) и беззубых (Mystacoceti) китов появились соответственно в позднем эоцене и олигоцене. (По мнению некоторых палеонтологов, происхождение китообразных связано не с хищными, а с древнейшими парнокопытными).
Морская ихтиофауна палеогена уже включала все современные крупные группы рыб, хотя видовой состав и отличался от современного. Уже в плиоценовой эпохе неогена существовала крупнейшая из хищных акул Megalodon, длина тела которой достигала 30 м, а диаметр открытой пасти был свыше 2 м.
Вернемся, однако, на сушу, где разворачивались наиболее интересные события кайнозойской эволюции органического мира. До сих пор мы концентрировали внимание на растительноядных млекопитающих, в связи с их важной ролью в биоценозах как первичных потребителей растительной биомассы. От примитивных насекомоядных предков возникли также разнообразные хищники, широко представленные среди палеогеновых млекопитающих.
Древнейшие представители отряда плацентарных хищников (Carnivora) - креодонты (Creodonta) по некоторым чертам своей организации были близки к примитивным копытным - кондиляртрам и, вероятно, имели общее с ними происхождение. Креодонты разделяли все примитивные особенности архаических групп млекопитающих (длинный низкий череп с маленькой мозговой коробкой, пятипалая конечность, плантиградность и др.)- Ископаемые остатки креодонтов известны, начиная с нижних горизонтов палеоцена, поэтому можно полагать, что обособление их эволюционного ствола произошло еще до конца мела. Наиболее ранние креодонты имели небольшие размеры, тонкие конечности и стройное тело, представляя переходный тип от насекомоядных предков. Однако уже в палеоцене появились более крупные хищники, среди которых арктоцион (Arctocyon) размерами и телосложением напоминал медведей. В раннем олигоцене существовали еще более крупные и тяжело построенные креодонты; самый большой из них, эндрюсархус (Andrewsarchus), достигал длины около 4 м при высоте в холке до 2 м, его череп имел длину 0,9 м. И крупные, и мелкие креодонты питались различными архаическими растительноядными млекопитающими и другой доступной им добычей. Поведение креодонтов было, вероятно, относительно простым по сравнению с поведением современных хищников как в отношении приемов охоты, так и в уходе за потомством.
После возникновения новых групп млекопитающих с более высоко развитым головным мозгом креодонты должны были сойти со сцены. Уже из верхнеэоценовых отложений известны ископаемые остатки древнейших представителей высших хищников (Fissipedia), обладавших сравнительно большим головным мозгом и рядом других прогрессивных признаков. В олигоцене последние креодонты вымирают.
В палеогене помимо млекопитающих процветали и многие другие группы наземных позвоночных. Во влажных тропических лесах, в пресных водоемах в изобилии встречались бесхвостые И хвостатые земноводные; в более открытых местообитаниях были многочисленны ящерицы и змеи. Число чешуйчатых рептилии в кайнозое ничуть не уменьшилось, и возникли некоторые новые семейства (например, в палеогене сформировались высшие змеи - Caenophidia, к которым принадлежат полозовые, или ужовые, - Colubridae, и все группы ядовитых змей). На суше, в пресных водоемах и в морях жили разнообразные черепахи. Представители единственной группы архозавров, пережившей великое вымирание в конце мела - крокодилы - процветали в жарком климате палеогена и были широко распространены на всех континентах. Наконец, потомки архозавров, птицы в кайнозое стали самой обильной видами группой наземных позвоночных.
Между птицами и млекопитающими произошел своего рода "раздел сфер влияния": большинство видов зверей, возникших от сумеречных и ночных мезозойских млекопитающих, сохранили и в кайнозое предпочтение к ночной активности, тогда как птицы, потомки гелиотермных рептилий, остались преимущественно дневными формами. Например, в составе современной фауны тропических лесов Габона и Панамы дневной образ жизни ведет 96 % видов птиц, тогда как для 70% видов млекопитающих характерна ночная активность. Разумеется, оба класса высших амниот отчасти проникли и в "чужую сферу влияния": имеются преимущественно дневные группы млекопитающих и ночные птицы. Но это относится к значительному меньшинству видов в обоих классах и потребовало развития у них специальных адаптации, не характерных для данного класса в целом. У млекопитающих, по мнению Ш.-Д.Пьера, с освоением дневной активности связано увеличение размеров тела и развитие специализированной пищеварительной системы, позволяющей использовать относительно грубые, трудно перевариваемые и объемные растительные корма (напомним о крупных и очень крупных животных среди растительноядных млекопитающих). Интересно появление в раннем эоцене крупных нелетающих наземных птиц с редуцированными крыльями. Некоторые из них обладали высоким массивным клювом с берыми краями и были, вероятно, опасными хищниками. Эоценовые диатримы (Diatryma. рис. 84), Достигавшие двухметровой высоты, или более поздние (олигоцен-плейстоцен) и еще более крупные (до 3 м) фороракосы (Phororhacidae) напоминали двуногих хищных динозавров (разумеется, на более высоком уровне организации птиц). Существование таких крупных нелетающих птиц показывает, что после вымирания динозавров птицы, как и млекопитающие, сделали попытку занять освободившиеся в биоценозах экологические ниши крупных хищников. Неудача птиц в этой роли может быть связана с меньшим по сравнению со зверями совершенством размножения и трудностями насиживания яиц и выращивания птенцов для нелетающих видов, численность которых как крупных хищников не могла быть высокой. Во всяком случае, эти своеобразные "пернатые динозавры" вымерли после появления высших групп хищных млекопитающих.

Вероятно, еще в меловое время от остальных птиц обособились нелетающие страусоподобные птицы - группа бескилевых (Ratitae), ископаемые остатки которых известны из палеоцена Бразилии (Diogenornis) и среднего эоцена Германии (Palaeotis). Возможно, бескилевые птицы представляют собой ряд независимых филогенетических стволов, эволюционировавших параллельно друг другу. Утрата способности к полету происходила на разных этапах эволюционной истории птиц у многих групп и была, вероятно, связана с отсутствием опасных хищников в местообитаниях, занимаемых птицами. Не случайно нелетающие птицы часто возникали на островах и изолированных континентах (Австралия, Южная Америка).
Уже с позднеэоценового времени известны пингвины (Impen-nes), ископаемые остатки которых обнаружены в Новой Зеландии и Южной Америке. Вся эволюционная история этой группы протекала в Южном полушарии. Высказывались предположения, что уход пингвинов с суши в океан связан с оледенением Антарктиды, но это неверно: пингвины как группа морских рыбоядных птиц, утративших способность к полету, сформировались задолго до начала оледенения (и среди современных пингвинов многие виды гнездятся в умеренной зоне, а один вид - даже в тропиках). Ближайшими родственниками пингвинов являются хорошо летающие трубконосые птицы (Procellariae), к которым относятся альбатросы и буревестники.
На отрезанных материках
Мы уже упоминали, что Австралия, Южная Америка и Антарктида по крайней мере с эоцена полностью обособились друг от друга и от всех других континентов широкими морскими проливами и морями. К. сожалению, о развитии жизни в Антарктиде перед началом последнего оледенения известно очень мало.
Австралия уже со времени распада Гондваны занимала обособленное положение, будучи связана с другими материками, главным образом, через Антарктиду (см. рис. 60). Поэтому в Австралию ко времени ее изоляции, вероятно, еще не проникли плацентарные млекопитающие. В Австралии сформировалась уникальная фауна сумчатых и однопроходных, отчасти сохранившаяся до наших дней. Однако палентологическая история австралийских млекопитающих известна еще очень плохо. Вероятно, современная фауна этого материка значительно обеднена по сравнению с предшествовавшими эпохами кайнозоя. Относительно хорошо известна поздняя - плейстоценовая - фауна Австралии. Среди плейстоценовых сумчатых здесь можно упомянуть дипротодона (Diprotodon) - самого крупного среди известных сумчатых, достигавшего размеров носорога с черепом около 1 м и мощными бивнеподобными резцами в верхней и нижней челюстях. Очень своеобразен был "сумчатый лев" (Thylacoleo), у которого коренные зубы были редуцированы, зато имелись огромные режущие предкоренные. Некоторые палеонтологи считают
;это животное хищником, другие же полагают, что "сумчатый лев" питался какими-то плодами.
В Южную Америку до ее отделения от Северной Америки успели проникнуть некоторые архаические группы плацентарных млекопитающих, имелись там и сумчатые. На основе этих групп в течение длительной изоляции Южной Америки сложилась удивительная фауна архаических млекопитающих. В Северной Америке, Европе и Азии в течение палеогена и неогена плацентарные млекопитающие в целом сделали большой шаг вперед по пути морфофизиологического прогресса (увеличение и усовершенствование головного мозга, усложнение поведения, прогрессивные изменения локомоторного аппарата, органов пищеварения, зубной системы и т. д.). В Южной Америке все это время продолжали процветать архаические группы, сохранявшие примитивное строение головного мозга, но также на свой лад совершенствовавшие органы пищеварения и локомоции, причем многие их адаптации оказались очень сходными с соответствующими приспособления-Ми млекопитающих Старого Света и Северной Америки.
Очень разнообразны и причудливы были южноамериканские растительноядные млекопитающие. Копытные были представлены группами нотоунгулят (Notoungulata), астрапотериев (Astra-Potheria), литоптерн (Litopterna) и пиротериев (Pyrotheria). Нотоунгуляты известны также из палеоцена Азии (где, вероятно, и возникла эта группа) и раннего эоцена Северной Америки, но исчезли повсеместно уже в эоцене, сохранившись и процветая лишь в Южной Америке. Южноамериканские нотоунгуляты были очень разнообразны; среди них были и проворные мелкие животные, подобные типотериям (Typotheria), с длинными задними конечностями, которые напоминали кроликов пропорциями тела, размерами и строением зубов; и тяжелые коротконогие токсо-донты (Toxodonta), достигавшие 2,75 м длины и похожие на некоторых безрогих носорогов.
Астрапотерии и пиротерии были крупнейшими копытными Южной Америки, достигая размеров крупных носорогов и даже слонов. С последними они были сходны также строением конечностей и наличием хобота, а пиротерии удивительно напоминали настоящих хоботных еще и обликом черепа и зубной системы (вплоть до крупных бивнеподобных резцов в верхней и нижней челюстях).
В эволюции литоптерн возникли формы, конечности которых были поразительно сходны с лошадиными (яркий пример эволюционного параллелизма). В процессе приспособления к быстрому бегу по относительно плотной почве саванн у этих литоптерн усиливался средний палец и редуцировались боковые (рис. 85), вплоть до полной их редукции у миоценового тоатерия (Thoatherium). Тоатерий был в этом отношении даже более специализирован, чем настоящие лошади, сохраняющие рудименты боковых пальцев в виде так называемых грифельных косточек (см. рис. 81). Наиболее поздний представитель литоптерн - плейстоценовая макраухения (Macrauchenia) размерами и пропорциями тела напоминала верблюда, но имела трехпалые ноги, а ноздри на черепе располагались сверху, как у хоботных животных (возможно, макраухения также имела небольшой хобот).

Таким образом, при отсутствии в Южной Америке настоящих хоботных, носорогов, лошадей, зайцеобразных и других групп млекопитающих, эволюционировавших в Евразии и Северной Америке, там конвергентно возникли формы, удивительно напоминавшие внешне своих аналогов.
Зато совершенно специфичны южноамериканские неполнозубые (Edentata). Эта группа, возможно, возникла в Северной Америке, откуда известны остатки древнейших палеоценовых неполнозубых, но центром ее эволюции стала Южная Америка в условиях географической изоляции. И современные неполнозубые (ленивцы, броненосцы, муравьеды) обитают в основном в Южной Америке, за исключением нескольких видов, проникших в южную часть Северной Америки после восстановления ее связи с Южной. Среди вымерших неполнозубых интересны родственные броненосцам глиптодонты (Glyptodontidae, рис. 86), достигавшие до 5 м в длину. Их тело было одето сплошным панцирем из сросшихся мелких костных пластинок - остеодерм. "Шапочка" из остеодерм прикрывала сверху голову. Пальцы коротких мощных лап были защищены копытами.


Но особенно интересной группой неполнозубых были гигантские наземные ленивцы, или мегатерии (Megatheriidae, рис. 87). Эти животные, как и глиптодонты, известны начиная с олигоце-на. Некоторые более поздние представители наземных ленивцев достигали огромных размеров: плейстоценовый мегатерий (Megatherium) был высотой со слона при длине тела около 6 м. Внешний облик этих странных существ был необычен. Огромное тело опиралось на мощные конечности, особенно велики были задние лапы с большими широкими ступнями. Передние конечности несли крупные изогнутые когти и при ходьбе опирались не на ладонную, а на боковой край тыльной стороны, сгибая пальцы внутрь. Вероятно, эти гиганты передвигались медленно и довольно неуклюже. Судя по строению их скелета, мегатерии могли вздымать свое огромное тело вертикально, опираясь на задние лапы; передними конечностями они при этом пригибали ветви и кроны деревьев вниз, цепляясь крепкими когтями. Питались гигантские ленивцы листьями. Конечно, не все мегатерии были столь велики, меньшие виды имели длину тела примерно 2-3 м. Гигантские ленивцы были широко распространены в Южной Америке, а после соединения двух Америк проникли и в Северную.
Плацентарных хищников в Южной Америке не было. Соответствующие экологические ниши занимали в основном разнообразные сумчатые хищники (Borhyaenidae), коротконогие и большеголовые звери, вооруженные острыми клыками и крепкими когтями; самые крупные виды достигали размеров медведей. С сумчатыми хищниками конкурировали упомянутые нами выше полутора - трехметровые хищные нелетающие птицы - фороракосы, а также самые крупные из известных летающих птиц - тераторнисы (Teratornithidae). Среди них аргентавус (Argentavus), известный из верхнемиоценовых отложений Аргентины, достигал в размахе крыльев 7,5 м при вероятной массе тела около 80 кг. Череп аргентавуса (длиной до 55 см) и других тераторнисов был вооружен мощным клювом. Вероятно, эти птицы могли проглатывать целиком добычу размером с зайца.
Существует предположение, что в Южной Америке вернулись к наземной жизни и крокодилы из группы себекозухий (Sebecosuchia), которые стали новой группой сухопутных хищных архозавров. У себекозухий череп был сжат с боков, а не уплощен сверху, как у большинства крокодилов, плоский череп которых с обращенными вверх глазницами и ноздрями характерен для хищников, подстерегающих в воде наземную добычу.
В плиоценовую эпоху неогена вновь восстановилась связь между Южной и Северной Америками, и по сухопутному "мосту" в Южную Америку хлынула волна новых, более современных групп млекопитающих (непарно- и парнокопытные, зайцеобразные, хоботные, хищники-фиссипедии). Это привело к быстрому вымиранию большинства специфических южноамериканских групп растительноядных и хищных млекопитающих. До плейстоцена продержались лишь немногие литоптерны и нотоунгуляты. Зато неполнозубые оказались вполне конкурентоспособными по отношению к высшим группам млекопитающих и даже расселились в Северную Америку. Это относится и к глиптодонтам, и к гигантским ленивцам, обитавшим в плейстоцене на огромных территориях от Аризоны и Флориды до Аргентины. Вселились в Северную Америку и гигантские хищные птицы - тераторнисы. Гигантские ленивцы дожили, несомненно, до времени появления в Америке человека. В некоторых пещерах в Северной и Южной Америке сохранились не только скелеты этих животных, но и остатки их шкур - вместе с каменными и костяными орудиями человека. Возможно, последние мегатерии вымерли всего несколько тысячелетий назад. Не исключено, что преследование со стороны людей послужило одной из причин вымирания этих медлительных и неуклюжих животных. В то же время, забегая несколько вперед, отметим, что в конце плейстоцена в обеих Америках вымерло подавляющее большинство существовавших здесь прежде многочисленных крупных млекопитающих.
Развитие жизни в неогене
Как мы уже упоминали, в течение неогена произошло значительное повышение материков, сопровождавшееся морской регрессией, постепенным иссушением климата и прогрессирующим развитием открытых ландшафтов. Одновременно происходило постепенное понижение средних температур на поверхности Земли. Эти изменения природных условий оказали глубокое влияние на флору (см. начало этой главы) и фауну.
В неогене получили преобладание виды наземных животных, приспособленные к жизни в открытых и относительно сухих местообитаниях типа лесостепей, степей и саванн. В олигоцене вымерли многие виды, приспособленные к жизни в лесах, на влажных лугах, заболоченных участках и других подобных биотопах, характерных для палеогена. Эта судьба постигла многие группы непарнокопытных. Зато парнокопытные, начиная с олигоцена, широко распространились и быстро прогрессировали. Особенно это относится к наиболее новым их группам: оленям (Cervidae), жирафам (Giraffidae), вилорогам (Antilocapridae) и многочисленным полорогим (Bovidae). Успех этих групп прежде всего связан со специализацией зубов и пищеварительной системы к переработке грубых растительных кормов - злаков и других растений степей и саванн, а также с приспособлениями к быстрому бегу по относительно плотной почве травянистых равнин.
Из непарнокопытных в неогене успешно конкурировали с парнокопытными лишь лошади и носороги. Миоценовые и плиоценовые лошади были очень многочисленны и разнообразны. Обшее направление их эволюции (как и обычно, прослеживающееся в нескольких независимых и параллельных филетических линиях) характеризовалось дальнейшим увеличением размеров тела, происходившим коррелятивно удлинением морды и соответственно зубного ряда, увеличением размеров коренных зубов и развитием сложной системы гребней на их жующей поверхности, а также удлинением конечностей с дальнейшей редукцией боковых пальцев (см. рис. 81). При этом в разных филетических линиях преобразования различных указанных признаков происходили с разной скоростью и независимо друг от друга: в одних линиях быстрее прогрессировали преобразования зубов, в других - конечностей и т.п.
На основании анализа этих преобразований Г.Осборном был сформулирован так называемый принцип независимой эволюции различных признаков, согласно которому разные признаки организма могут изменяться в ходе эволюции в определенной степени независимо друг от друга. Разумеется, независимо эволюционировать могут лишь те признаки, которые не связаны друг с другом какими-либо коррелятивными взаимоотношениями. Принцип Осборна лежит в основе концепции "мозаичной эволюции" Г. де Вира (см. с. 249, 350).
Для миоцена были характерны трехпалые лошади (Parahippus, Merychippus, Hipparion и др.), у которых боковые пальцы были укорочены и касались субстрата, вероятно, лишь при более медленной ходьбе и при пересечении участков с влажной почвой, препятствуя увязанию. Центром эволюции лошадей оставалась Северная Америка, откуда эти непарнокопытные проникли в Евразию. В плиоцене появились формы (Pliohippus, Plesippus), строение конечностей и зубов у которых приближалось к таковым у современной лошади (Equus), известной начиная с плейстоцена. Отметим, что однопалость развивалась независимо и параллельно по меньшей мере в четырех обособленных филетических линиях лошадей (Astrohippus, Pliohippus, Hippidion - Onohippidion, Protohippus - Equus).
В неогене максимального расцвета достигли хоботные, расселившиеся из Африки в Евразию, затем в Северную и, наконец, в Южную Америку. Центральный эволюционный ствол хоботных был представлен мастодонтами (рис. 88), среди которых появились очень разнообразные формы: с коротким и длинным хоботом, различного строения бивнями и т.п. Причудливо выглядели лопаторылые мастодонты (Amebelodon, Platybelodon), у которых передняя часть нижней челюсти чрезвычайно удлинилась, образуя вместе с огромными уплощенными резцами своего рода "совок" или "лопату", выдвинутую вперед и достигавшую чудовищных размеров: животное высотой в холке 2,5 м имело нижнюю челюсть длиной более 2 м. По-видимому, лопаторылые мастодонты использовали свою необычайную нижнюю челюсть именно как лопату, подцепляя ею корм - может быть, со дна неглубоких полузаболоченных водоемов (сочные листья, клубни или корневодных растений).

Боковой ветвью эволюционного ствола мастодонтов были миоценовые динотерии (Deinotherium), у которых передняя часть нижней челюсти была отогнута вниз, а сидящие на ней бивни были направлены вниз и назад. Сейчас трудно представить, как использовали такие бивни эти крупные хоботные.
В миоцене от мастодонтов возникли настоящие слоны (Elephantidae). Их родиной, вероятно, была Южная Азия, откуда они в плиоцене расселились в Европу, Северную Америку и Африку. У слонов в отличие от мастодонтов и динотериев сохранилась лишь верхнечелюстная пара бивней, но особенно характерной чертой настоящих слонов является строение огромных коренных зубов. Их высокие и сжатые с боков коронки состоят как бы из многочисленных поперечных пластин, образующих на жевательной поверхности узор из узких поперечных гребней и желобков.
Обилию растительноядных млекопитающих в лесостепях и саваннах неогена соответствует и разнообразие хищников. В миоцене существовали уже все семейства современных хищных млекопитающих, большинство современных родов известно начиная с плейстоцена.

Среди ископаемых кошачьих (Felidae) интересна группа так называемых саблезубов (или правильнее "кинжалозубов" - Маchairodontinae), существовавшая с олигоцена до плейстоцена. Этих хищников иногда называют "саблезубыми тиграми", но к тиграм они были не более близки, чем к львам, леопардам или домашним кошкам (которые все являются представителями другого подсемейства кошачьих, собственно кошек - Felinae). Саблезубы (рис. 89) обладали огромными уплощенными с боков клыками, достигавшими у крупных форм длины 15 см; этими клыкам хищники и обязаны своим названием. Это были крепкие коренастые животные с мощной шеей и могучими когтистыми лапами, причем передние конечности были длиннее и сильнее задних. Такой тип телосложения совсем не соответствует способам охоты, характерным для современных кошачьих: едва ли саблезубы могли совершать дальние прыжки на добычу из засады или стремительно преследовать ее даже на коротких расстояниях. Огромные клыки и способность чрезвычайно широко открывать пасть, отводя нижнюю челюсть вниз и назад под углом, близким к прямому или даже несколько большим, указывают на то, что главным способом умерщвления добычи у саблезубов было нанесение сильных ударов клыками, глубоко вонзавшимися в тело жертвы и причинявшими тяжелые ранения. Вероятно, основной добычей крупных саблезубов (отметим, что среди них имелись и относительно мелкие виды) были крупные толстокожие растительноядные животные (хоботные, носороги и т.п.), на которых хищники нападали из засады.
Интересно, что саблезубые формы хищников возникали неоднократно и в разное время среди различных групп млекопитающих. Древнейшими из них являются эоценовые саблезубые креодонты Machaeroides - относительно мелкие хищники, питавшиеся, вероятно, кондиляртрами. В плиоцене в Южной Америке существовал сумчатый саблезуб (Thylacosmilus, рис. 90). Его добычей, вероятно, были такие крупные толстокожие животные, как токсодонты, мегатерии и т.п. В плейстоцене его вытеснили настоящие саблезубы (Smilodon), проникшие в Южную Америку из Северной вместе с другими видами высших млекопитающих. Возникновение саблезубых хищников в разных группах млекопитающих является одним из ярких примеров параллельной эволюции независимых филетических линий. Вымирание большинства крупных растительноядных форм к концу плейстоцена привело к вымиранию охотившихся на них саблезубов.

В неогене водные млекопитающие пополнились ластоногими (Pinnipedia), ископаемые остатки древнейшего представителя которых (Enaliarctos) известны из пограничных отложений олиго-цена и миоцена. Возможно, разные филогенетические стволы ластоногих возникли от примитивных представителей различных семейств наземных хищников: ушастые тюлени (Otariidae) и моржи (Odobaenidae) - от медвежьих, а настоящие или безухие тюлени (Phocidae) - от куньих. Общие особенности строения ластоногих сформировались параллельно и независимо в этих двух филогенетических стволах.
На огромных пространствах Северной Америки, Евразии и Африки, занятых лесостепями и степями с разной степенью увлажненности, в позднем миоцене и плиоцене получила широкое распространение фауна, которую называют "гиппарионовой" - по самому характерному ее представителю, трехпалой лошади - гиппариону (Hipparion, рис. 91). Многотысячные табуны гиппарионов паслись на лугах и остепненных участках, перемежающихся с рощами широколиственных листопадных деревьев и кустарников (дуб, бук, ясень, вяз, ольха, различные виды тополя, граб, клен, грецкий орех и др.) и разреженными лесами, включавшими и широколиственные, и хвойные деревья (сосны, секвойи, болотные кипарисы), прорезанных долинами рек и заболоченными низинами. Вместе с гиппарионами обитали разнообразные антилопы, олени, древние жирафы (например, палеотрагусы Palaeotragus, похожие на современных окапи, и тяжелые буйволообразные сиватерии - Sivatherium), хоботные (мастодонты и динотерии), носороги и страусы, близкие к современным африканским. В состав гиппарионовой фауны входили, конечно, и хищники (саблезубы, различные псовые, гиены, росомахи, куницы, виверры и др.), а также грызуны, зайцеобразные и обезьяны1.

Гиппарионовая фауна, рассматриваемая в целом, была в значительной степени предковой для современной фауны Африки, включающей представителей тех же основных групп животных. По аналогии с недавним прошлым африканских саванн и по обилию ископаемых остатков различных копытных в местонахождениях гиппарионовой фауны можно заключить, что в конце миоцена - плиоцене на огромных пространствах равнин Старого и Нового Света паслись неисчислимые стада гиппарионов, антилоп, оленей, хоботных и других растительноядных млекопитающих. Ю.А.Орлов высказал предположение, что в зоне умеренного пояса, где уже в неогене были отчетливо выражены сезонные изменения климата, эти гигантские стада копытных и сопровождающие их хищники должны были совершать весной и осенью далекие миграции, подобные миграциям африканских копытных, но в гораздо большем масштабе.
Четвертичный период
В плейстоцене происходило дальнейшее похолодание климата, сопровождавшееся постепенным отступанием теплолюбивой флоры и фауны в Евразии и Северной Америке к югу. Широкое распространение получили листопадные и хвойные леса, общий облик которых уже напоминал современные леса умеренного пояса. Состав фауны также приближался к современному; появились многие современные роды млекопитающих.
Однако в плейстоцене еще существовал целый ряд более древних форм и групп животных (мегатерии, глиптодонты, макраухении, токсодонты, мастодонты, динотерии, саблезубы и др.), особенно многочисленные в Америке. Появились и некоторые своеобразные виды, которым не было суждено пережить плейстоценовую эпоху. Среди них интересны гигантские бобры - трогон-терии (Trogontherium), имевшие более 2 м длины; носороги - эласмотерии (Elasmotherium), обладавшие огромным вздутием лобной области черепа, адаптивная роль которого остается неизвестной; большерогие олени - мегалоцеросы, или "ирландские лоси" (Megaloceros; рис. 92), достигавшие высоты в холке около 2 м, но особенно замечательные своими гигантскими ветвистыми рогами, размах которых превышал 4 м. Эти поразительные рога требовали, конечно, от организма немалых энергетических затрат и на их развитие в онтогенезе, и на поддержание головы с таким тяжелым вооружением. Вероятно, мегалоцеросы использовали свои рога для тех же целей, что и все остальные олени: как демонстрационный признак и "турнирное оружие" в борьбе между самцами в период гона (у самок рога отсутствовали). Развитие рогов было обусловлено половым отбором, который нередко (при отсутствии опасных хищников или при хорошей защищенности данного вида особенностями его образа жизни занимаемых местообитаний) приводит к формированию подобных демонстрационных признаков, выглядящих своего рода излишествами (достаточно вспомнить хотя бы хвост павлина или птицы-лиры, ослепительные окраски фазанов и райских птиц и т.п.).
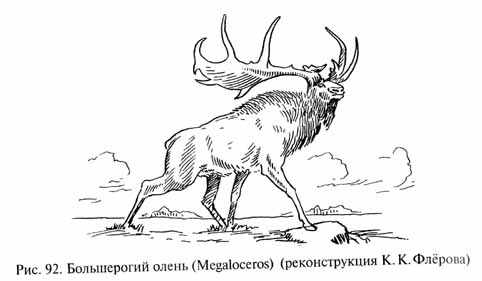
Из плейстоценовых отложений известны многочисленные находки крупных нелетающих страусоподобных птиц, большинство видов которых вымерло к концу плейстоцена или были истреблены человеком в совсем недавнее время. Эта судьба постигла в первую очередь крупных птиц, обитавших на островах: мадага-скарских эпиорнисов (Aepiornithes), новозеландских моа (Dinor-nithes) и др.
Как уже упоминалось, прогрессировавшее похолодание завершилось в плейстоцене развитием великого оледенения. Наступление ледников сопровождалось смещением климатических зон к экватору, вместе с которыми отступали теплолюбивые фауны и флоры, отчасти подвергавшиеся при этом вымиранию. В межледниковые эпохи, продолжавшиеся по нескольку десятков тысяч лет, происходили обратные смещения климатических зон и соответствующих флор и фаун.
В то же время в относительной близости к границам оледенения в Евразии и Северной Америке сформировался специфический приледниковый пояс с особыми физико-географическими условиями: резко континентальный климат с низким уровнем средних температур при сухом воздухе и значительной обводненности территории летом за счет талых ледниковых вод, с возникновением в низинах озер и болот. В этой обширной приледниковой зоне возник особый биоценоз - тундростепь, существовавшая все время оледенения и перемещавшаяся в соответствии с изменениями границ ледника к северу или к югу. Флора тундростепи включала различные травянистые растения (особенно злаки и осоки), мхи, а также мелкие деревца и кустарники, произраставшие главным образом в долинах рек и по берегам озер: ивы, березы, ольхи, сосны и лиственницы. При этом общая биомасса растительности в тундростепи была, по-видимому, очень велика, в основном за счет трав, что позволило расселиться на огромных пространствах приледникового пояса обильной и своеобразной фауне, которую называют мамонтовой.
Эта удивительная приледниковая фауна включала мамонтов, шерстистых носорогов, овцебыков, короткорогих бизонов, яков, северных оленей, антилоп-сайгаков и дзеренов, лошадей, куланов, грызунов - сусликов, сурков, леммингов, зайцеобразных, а также различных хищников: пещерных львов, пещерных медведи, волков, гиен, песцов, росомах. Состав мамонтовой фауны свидетельствует о том, что она произошла от гиппарионовой фауны, будучи ее северным приледниковым вариантом, тогда как современная африканская фауна является южным, тропическим производным гиппарионовой.
Для всех животных мамонтовой фауны характерны приспособления к жизни в условиях низких температур, в частности длинная и густая шерсть. Густой и очень длинной рыжей шерстью с длиной волос до 70-80 см был покрыт и мамонт (Mammonteus, рис. 93) - северный слон, обитавший 50-10 тыс. лет назад на огромных территориях Европы, Азии и Северной Америки.

Изучение представителей мамонтовой фауны значительно облегчается сохранением целых трупов или их частей в условиях вечной мерзлоты. На территории нашей страны был сделан целый ряд замечательных находок такого рода. Наиболее известен из них так называемый "березовский" мамонт, найденный в 1901г. на берегу реки Березовки в Северо-Восточной Сибири, а последняя находка - почти целый труп мамонтенка 5-7 месяцев от роду, обнаруженный в 1977г. на берегу ручья, впадающего в реку Берелех (приток Колымы).
По пропорциям тела мамонт заметно отличался от современных слонов, индийского и африканского. Теменная часть головы сильно выступала вверх, а затылок был скошен вниз, к глубокой шейной выемке, позади которой на спине возвышался большой горб, состоявший из жира. Вероятно, это был запас питательных веществ, использовавшийся во время голодного зимнего сезона. Позади горба спина была круто скошена вниз. Огромные бивни, до 2,5 м длины, закручивались вверх и внутрь. В содержимом желудков мамонтов были обнаружены остатки листьев и стеблей злаков и осок, а также побегов ив, берез и ольх, иногда даже лиственниц и сосен. Основу питания мамонта, вероятно, составляли травянистые растения.
Во многих местах, где прежде обитали мамонты: в Сибири, на Новосибирских островах, на Аляске, на Украине и др. - были обнаружены огромные скопления скелетов этих животных, так называемые "Мамонтовы кладбища". О причинах возникновения мамонтовых кладбищ было высказано немало предположений. Наиболее вероятно, что они сформировались, как и большинство массовых скоплений ископаемых остатков наземных животных, в результате сноса течением рек, особенно во время весенних половодий или летних разливов, в разного рода естественные отстойники (заводи, омуты, старицы, устья оврагов и т.п.), где целые скелеты и их фрагменты накапливались в течение многих лет.
Вместе с мамонтами обитали шерстистые носороги (Coelodonta), покрытые густой коричневой шерстью. Облик этих двурогих носорогов, так же как мамонтов и других животных этой фауны, был запечатлен людьми каменного века - кроманьонцами в их рисунках на стенах пещер. На основании археологических данных можно с уверенностью утверждать, что древние люди охотились на самых различных животных мамонтовой фауны, в том числе и на шерстистых носорогов и самих мамонтов (а в Америке на еще сохранявшихся там мастодонтов и мегатериев). В связи с этим высказывались предположения, что человек мог сыграть определенную роль (по мнению некоторых авторов - даже решающую) в вымирании многих плейстоценовых животных.
Вымирание мамонтовой фауны отчетливо коррелирует с концом последнего оледенения 10-12 тыс. лет назад. Потепление климата и таяние ледников резко изменили природную обстановку в прежнем поясе приледниковой тундростепи: значительно увеличились влажность воздуха и выпадение осадков, как следствие, на больших территориях развилась заболоченность, зимой возросла высота снегового покрова. Животные мамонтовой фауны, хорошо защищенные от сухого холода и способные добывать себе пищу на просторах тундростепи в малоснежные зимы ледниковой эпохи, оказались в крайне неблагоприятной для них экологической обстановке. Обилие снега зимой сделало невозможным добывание пищи в достаточном количестве. Летом же высокая влажность и заболачивание почвы, крайне неблагоприятные и сами по себе, сопровождались колоссальным возрастанием численности кровососущих насекомых (гнуса, столь обильного и в современной тундре), укусы которых изнуряли животных, не давая им спокойно Кормиться, как это происходит и ныне с северными оленями. Таким образом, мамонтовая фауна оказалась за очень короткий срок (таяние ледников происходило очень быстро) перед лицом резких изменений среды обитания, к которым большинство составлявших ее видов не смогло столь быстро приспособиться, и мамонтовая фауна как целое прекратила свое существование. Из числа крупных млекопитающих этой фауны сохранились до наших дней северные олени (Rangifer), обладающие большой подвижностью и способные совершать дальние миграции: летом в тундру к морю, где меньше гнуса, а на зиму на ягельные пастбища в лесотундру и тайгу. В относительно малоснежных местообитаниях на севере Гренландии и на некоторых островах Североамериканского архипелага сохранились овцебыки (Ovibos). Приспособились к новым условиям некоторые мелкие животные из состава мамонтовой фауны (лемминги, песцы). Но большинство видов млекопитающих этой замечательной фауны вымерли к началу голоценовой эпохи.
(По некоторым данным, в голоцене 4-7 тыс. лет назад на острове Врангеля еще сохранялась популяция измельчавших мамонтов) (См. кн.: Верещагин Н.К. Почему вымерли мамонты. - М.. 1979).
В конце плейстоцена произошло еще одно значительное изменение фауны, правда ограниченное территорией Америки, но остающееся до сих пор загадочным. В обеих Америках вымерло подавляющее большинство крупных животных, столь обильных там прежде: и представители мамонтовой фауны, и обитавшие в более южных районах, где не было никакого оледенения, мастодонты и слоны, все лошади и большинство верблюдов, мегатерии и глиптодонты. Видимо, еще в плиоцене исчезли носороги. Из крупных млекопитающих сохранились лишь олени и бизоны в Северной Америке и ламы и тапиры в Южной. Это тем более удивительно, что Северная Америка была родиной и центром эволюции лошадей и верблюдов, сохранившихся до нашего времени в Старом Свете.
Нет никаких признаков значительных изменений условий существования в конце плейстоцена на большей части территории Америки, не подвергавшейся оледенению. Более того, после появления в Америке европейцев некоторые из завезенных ими лошадей одичали и дали начало мустангам, быстро размножившимся в северо-американских прериях, условия которых оказались благоприятными для лошадей. Индейские племена, жившие охотой, не оказывали значительного влияния на численность огромных стад бизонов (и мустангов после их появления в Америке). Человек на уровне культуры каменного века едва ли мог сыграть решающую роль в вымирании многочисленных видов крупных плейстоценовых животных (за исключением, может быть, медлительных и малосообразительных мегатериев) на обширных территориях обеих Америк.
После завершения последнего оледенения 10-12 тыс. лет назад Земля вступила в голоценовую эпоху четвертичного периода, в течение которой установился современный облик фауны и флоры. Условия жизни на Земле ныне значительно более суровы, чем на протяжении мезозоя, палеогена и большей части неогена. И богатство и разнообразие мира организмов в наше время, судя по всему, существенно ниже, чем за многие прошлые геологические эпохи.
В голоцене все сильнее проявляется воздействие человека на среду обитания. В наше время с развитием технической цивилизации деятельность людей стала поистине важнейшим глобальным фактором, активно, хотя в большинстве случаев непродуманно и губительно, изменяющим биосферу.
В связи с формированием человека современного вида (Homo sapiens) и развитием человеческого общества в течение четвертичного периода А.П.Павлов предложил назвать этот период кайнозойской эры "антропогеном". Обратимся же теперь к эволюции самого человека.
Приматов можно кратко охарактеризовать как группу лесных теплолюбивых плацентарных млекопитающих, приспособившихся к лазающему древесному образу жизни, сохранив достаточно примитивную основу организации. Примитивные черты в строении приматов проявляются в сохранении ими пятипалой конечности, опирающейся при ходьбе на всю ступню (плантиградность); в сохранении ключиц, которые утрачиваются при совершенствовании бега, в отсутствии специализации питания (большинство приматов всеядны и питаются как растительной, так и животной пищей) и т. д. Сохранение некоторых примитивных признаков и отсутствие узкой специализации соответствующих органов способствовали высокой эволюционной пластичности приматов, проявивших в ходе своего филогенеза способность адаптироваться к самым различным условиям существования и способам использования природных ресурсов.
Важнейшие особенности приматов связаны с развитием приспособлений к древесной лазающей жизни. Прежде всего это совершенствование хватательной способности конечностей, с которым связано приобретение возможности вращать кисть и предплечье вокруг их продольной оси (способность к пронации и супинации кисти, т.е. к ее вращению ладонью вниз и вверх), а также развитие противопоставления больших пальцев всем остальным, облегчающего хватание. Эти приспособления значительно увеличили общую манипулятивную способность конечностей, т. е. способность различным образом действовать с удерживаемым пальцами предметом.
Лазание по веткам требует хорошо развитого осязания - для исследования поверхности опоры. В связи с этим на концевых фалангах пальцев развились мясистые пальцевые подушечки, кожа которых богата осязательными тельцами и нервными окончаниями, а когти преобразовались в ногти с плоской и тонкой когтевой пластинкой, защищающей конец пальца лишь сверху.
Но самую важную роль среди органов чувств при лазании приобретают глаза, поскольку прыжки с ветви на ветку требуют объемного (стереоскопического) зрения с точной оценкой расстояний и надежности опоры. Развитие стереоскопического зрения достигается перемещением глаз на переднюю (лицевую) поверхность головы, их сближением и все большим перекрыванием полей зрения. Это, собственно, и создает бинокулярный эффект: предмет рассматривается двумя глазами с разных сторон, и восприятие обоих глаз интегрируется в центральной нервной системе.
С другой стороны, на деревьях падает роль обоняния. Соответственно орган обоняния у приматов подвергается некоторой редукции.
Чрезвычайно важные изменения, связанные с адаптацией к лазанию, произошли в центральной нервной системе. Передвижение по веткам представляет собой один из наиболее сложных способов локомоции, требующий точной оценки расстояний и качества опоры и соответствующей коррекции движений. На деревьях каждый шаг и прыжок представляет собой, по удачному выражению Я.Я.Рогинского, "отдельный творческий акт". Все это требует высокого совершенства функций центральной нервной системы - как в анализе непрерывно меняющейся обстановки, так и в осуществлении тонкого мышечного контроля разнообразнейших оттенков движений и положения тела. При совершенствовании всех указанных функций естественный отбор благоприятствовал прогрессивному развитию головного мозга, особенно коры больших полушарий, в которой формируется сложная система борозд и извилин (у низших приматов кора имеет гладкую поверхность)1, а также мозжечка. Увеличение относительного и абсолютного размера головного мозга возможно только при соответствующих изменениях пропорций черепа, в котором относительно увеличивалась мозговая коробка и уменьшался челюстной отдел.
Кочевая жизнь на деревьях, которую ведет большинство обезьян, не позволяет самкам иметь одновременно много детенышей, так как они, цепляясь за шерсть матери, сковывают ее движения. Поэтому отбор благоприятствовал тенденции к уменьшению числа детенышей у приматов до 1-2 на самку за одну беременность. Для компенсации малочисленности потомства у обезьян сформировались сложные формы родительского поведения, обеспечивающие длительный уход за детенышами и защиту их родителями. Результатом уменьшения числа детенышей была и редукция числа млечных желез до единственной грудной пары.
Очень важной характеристикой приматов является их общественный (стайный) образ жизни. Эффективная организация жизни стаи требовала развития особых (социальных) форм поведения. Наиболее жизнеспособными оказывались те стаи, в которых наследственно закреплялись такие формы поведения, которые обеспечивали снижение взаимной агрессивности и помощь более слабым животным в своей стае. В стаях формировалась так называемая социальная иерархия, т. е. происходило выделение разных категорий особей: доминантов - вожаков, животных "второго ранга" - субдоминантов - и последующих уровней иерархии, что отвечало задаче оптимальной организации жизни стаи: совместного целенаправленного движения по охотничьим угодьям, сведения к минимуму конфликтов между разными особями, преодоления различных критических ситуаций. Усложнение поведения в результате общественной жизни требовало, с одной стороны, дальнейшего совершенствования головного мозга, с другой - развития системы сигналов, которая позволила бы передавать необходимую информацию между разными членами стаи.
Таким образом, давление отбора в направлении совершенствования высшей нервной деятельности, ориентации в пространстве, Манипулятивных способностей конечностей и социального поведения, т. е. в конечном итоге в направлении формирования важнейших особенностей человека, определялось спецификой организации и образа жизни приматов.
Ранние этапы эволюции приматов
Приматы возникли от примитивных насекомоядных млекопитающих (Insectivora), отдельные группы которых перешли к лазающей древесной жизни. К основанию эволюционного ствола приматов, возможно, близки древесные землеройки - тупайи (Tupaiidae), обитающие ныне в тропических лесах Малайского полуострова и Филиппинских островов.
Эволюционная ветвь насекомоядных, ведущая к приматам, обособилась, вероятно, еще до конца мезозоя. Из позднемело-вых отложений известны ископаемые остатки животных, которых ряд палеонтологов рассматривает уже в качестве примитивных приматов. Это были пургатории (Purgatorius) - мелкие зверьки с признаками приспособлений к лазающей древесной жизни. Вероятно, они имели ночную активность, как и другие мезозойские млекопитающие, и питались насекомыми и плодами деревьев.
Из самых нижних горизонтов палеоцена известны остатки лемуров (Lemuroidea) - наиболее примитивной группы среди современных приматов. Лемуры ныне обитают в тропических лесах Мадагаскара, Африки и Южной Азии. Они ведут сумеречный и ночной древесный образ жизни. Это небольшие животные, в среднем достигающие размеров кошки, с длинным хвостом, пушистым мехом и удлиненной мордочкой. Глаза у лемуров обращены еще не прямо вперед, как у более высокоразвитых приматов, а несколько косо - вперед и вбок. У лемуров сохраняется довольно хорошо развитое обоняние. Этих симпатичных зверьков иногда называют "полуобезьянами", подчеркивая, что они еще не приобрели всего комплекса признаков, характерного для высших приматов, и занимают по уровню организации промежуточное положение между последними и древесными землеройками.
От каких-то примитивных лемуров в первой половине палеогена возникли настоящие обезьяны - антропоиды (Anthropoidea). Вероятно, их обособление от лемуров было связано с переходом к дневной активности, сопровождавшимся усилением роли зрения, увеличением размеров и совершенствованием строения головного мозга, развитием стайного образа жизни и связанных с ним социальных форм поведения.
По мнению ряда антропологов, от общего корня с антропоидами произошли тарзиевые, или долгопяты (Tarsioidea), - своеобразная обособленная группа ночных насекомоядных приматов, в организации которых причудливо сочетаются примитивные И специализированные черты.
Древнейшие (фрагментарные) остатки антропоидов известны из среднеэоценовых отложений Алжира (Algeripithecus) верхнеэоценовых слоев Бирмы и Египта. В раннем олигоцене существовали уже достаточно разнообразные их представители. Наиболее известен из них парапитек (Parapithecus), близкий по уровню организации к низшим мартышкообразным обезьянам (Cercopithecidae) и, возможно, принадлежавший к группе, которая была предковой для более высокоразвитых антропоидов.
До сих пор остается нерешенной проблема происхождения широконосых обезьян (Platyrrhini) Нового Света, сохранивших в целом более примитивное строение, чем современные обезьяны Старого Света, объединяемые в группу узконосых (Catarrhini). Названия "широконосые" и "узконосые" связаны с характерными различиями этих групп по ширине перегородки между наружными отверстиями ноздрей. Возможно, широконосые обезьяны возникли от раннеолигоценовых парапитековых в Африке и проникли в Южную Америку уже после ее обособления от Северной. Древнейшие ископаемые остатки обезьян в Южной Америке (Branicella) известны из олигоценовых отложений, образовавшихся на несколько миллионов лет позднее, чем нижне-олигоценовые отложения Египта с остатками парапитека. Однако согласно другой гипотезе, предковый ствол широконосых обезьян мог обособиться от основного ствола антропоидов еще в эоцене вместе с изоляцией Южной Америки, и дальнейшая эволюция широконосых и узконосых обезьян шла независимо и параллельно.
Из нижнеолигоценовых отложений Египта известны кроме парапитеков ископаемые остатки более высокоразвитых обезьян, названных проплиопитеками (Propliopithecus). Проплиопитек, судя по особенностям строения его нижней челюсти и зубов, мог быть предком гиббонов (Hylobatidae), которых многие ученые рассматривают уже как низших представителей человекообразных, или антропоморфных приматов (Hominoidea, или Anthropomorpha). Помимо гиббонов к антропоморфным приматам относят семейства высших человекообразных обезьян (Pongidae, или Simiidae), к которым принадлежат современные шимпанзе (Pan), гориллы (Gorilla) и орангутаны (Pongo), и семейство людей (Hominidae).
Антропоморфным приматам присущи наиболее крупные в пределах отряда размеры тела (самцы гориллы при росте около 180 см достигают веса 200 кг) и характерные пропорции: относительно короткое туловище и длинные конечности. У человекообразных обезьян передние конечности заметно длиннее задних. Вероятно, Пропорции тела и полувыпрямленная осанка антропоморфов сложились в связи с особыми формами передвижения этих крупных обезьян по деревьям, которые получили названия "круриация" и "брахиация". При круриации обезьяна идет на задних конечностях по нижним толстым ветвям деревьев в вертикальном или близком к нему положении, охватывая стопами опорную ветвь и придерживаясь передними конечностями за верхние ветви. Брахиация представляет собой более специализированную форму передвижения: это своего рода быстрый "полет" под кронами деревьев с перехватыванием руками за ветви и свисающие лианы, подобно прыжкам гимнастов на трапеции. Брахиация особенно характерна для гиббонов, но в какой-то мере как вспомогательный способ передвижения могла использоваться и общими предками всех антропоморфов. Во всяком случае, использование круриации и в некоторой степени брахиации древнейшими антропоморфами позволяет объяснить некоторые особенности их строения1.
Антропоморфы обладают наиболее сложными формами поведения, в том числе и социального, среди всех приматов, чрезвычайно высокой способностью к разнообразным манипуляциям с предметами, вплоть до изготовления примитивных орудий, т.е. предварительной обработки какого-либо объекта, которая делает его пригодным для осуществления заранее задуманного действия (например, обработка ветви руками и зубами для последующего добывания с ее помощью плода). Однако человекообразные обезьяны не обнаруживают способности к настоящей трудовой деятельности, принципиальной особенностью которой является изготовление орудий не с помощью естественных органов, а посредством других предметов, которые используются как искусственные средства обработки. С другой стороны, человекообразные обезьяны весьма способны к научению: наблюдения ряда этологов в природных условиях показали, что в стаях шимпанзе происходит своего рода обмен опытом между разными обезьянами путем наблюдения и перенимания операций, "изобретенных" более старыми и опытными животными. Однако даже у высших обезьян в отличие от человека не обнаружено способности к продолжительной концентрации внимания, требующей подавления посторонних отвлекающих впечатлений и необходимой для перехода к настоящей трудовой деятельности.
Предпосылками для развития сложных форм поведения у антропоморфов было увеличение размеров головного мозга, достигающего у шимпанзе среднего объема 405 см3, а у горилл - 498 см3, и совершенствования его структуры. Соответственно мозговая коробка получила в их черепе еще большее относительное преобладание над лицевым отделом (рис. 94).

Эволюция человекообразных приматов
Некоторые антропологи полагают, что общим предком человекообразных приматов (антропоморфов) является проконсул (Proconsul), ископаемые остатки которого были обнаружены в раннемиоценовых отложениях Восточной Африки в Кении. Проконсул был типичной мозаичной формой, в организации которой соединились некоторые признаки низших обезьян, гиббонов и шимпанзе.
Расхождение филогенетических стволов, ведущих к двум семействам антропоморфов - человекообразным обезьянам (понгидам) и людям (гоминидам), произошло, возможно, уже в миоцене (по разным оценкам от 15 до 25 млн лет назад). Из верхнемиоценовых отложений Европы известны ископаемые остатки обезьян-дриопитеков (Dryopithecus), которые по основным особенностям строения и по размерам тела напоминали современных шимпанзе. Многие антропологи рассматривают дриопитеков как возможных непосредственных предков всех высших антропоморфов, т.е. понгид и гоминид.
В неогене антропоморфные приматы достигли своего высшего расцвета. Их ископаемые остатки широко представлены в местонахождениях гиппарионовой фауны. Это говорит о связи миоплиоценовых антропоморфов с характерными для этой фауны открытыми ландшафтами, тогда как большинство приматов и особенно низшие их представители являются обитателями тропических лесов, обильных в палеогене. Вероятно, расцвет антропоморфных приматов был связан с распадением единого лесного пространства на островные леса. Можно полагать, что древнейшие человекообразные обезьяны обитали в разреженных лесах на границах лесных массивов и лесостепей. В этих пограничных местообитаниях и произошла дивергенция понгид и гоминид: первые остались лесными обитателями, предки же вторых перешли к освоению все более открытых ландшафтов. Предпосылки для этого заключались, во-первых, в переходе к наземному образу жизни; во-вторых, во все большем использовании различных орудий для зашиты и нападения и соответственно в освобождении рук от участия в передвижении; в-третьих, в совершенствовании двуногого (бипедального) передвижения. Использование орудий (грубо обработанных камней и костей животных) вместе с высокоразвитым социальным поведением позволили древним гоминидам, не имевшим ни острых клыков, ни крепких когтей, ни мошной мускулатуры (по сравнению, например, с гориллами), не только обороняться от нападений крупных хищников, но и перейти регулярному добыванию животной пищи (лишь эпизодически используемой шимпанзе).
В качестве наиболее древних и примитивных представителей эволюционного ствола гоминид многие антропологи рассматривают рамапитека (Ramapithecus) и близкого к нему сивапитека (Sivapithecus), ископаемые остатки которых были обнаружены в верхнемиоценовых отложениях Индии, с абсолютным возрастом около 12 млн лет, а также в близких по времени возникновения отложениях в Китае, Кении и Венгрии. У рамапитековых, как и у гоминид, зубные ряды левой и правой сторон сзади расходятся друг от друга, а не идут параллельно, как у большинства обезьян; клыки относительно малы, а коронки крупных коренных зубов покрыты толстым слоем эмали. Вероятно, сивапитеки и рамапитеки были уже полностью наземными всеядными обезьянами. Они широко расселились в Африке, Европе и Южной Азии в конце миоцена и существовали, вероятно, с 14 до 8 млн лет назад. Не позднее этого времени произошла дивергенция линий, ведущих к высшим человекообразным обезьянам и к гоминидам (по мнению ряда антропологов, эта дивергенция произошла даже раньше - около 15 млн лет назад, на уровне дриопитеков).
К сожалению, пока еще нет палеонтологических данных о ближайших последующих этапах филогенеза гоминид. Зато большое количество ископаемых остатков высших антропоморфов известно начиная с верхнеплиоценовых отложений, имеющих возраст от 3,5-4 млн лет и моложе. Эти остатки принадлежат различным формам африканских австралопитековых (Australopithecinae): австралопитек (Australopithecus afarensis, A. africanus), парантроп (Paranthropus robustus), плезиантроп (Plesianthropustransvaalensis), зинджантроп (Zindjanthropus boisei). Многие антропологи полагают, что все эти формы являются видами одного рода Australopithecus.
Впервые остатки австралопитеков были обнаружены в Южной Африке в 1924г. С тех пор эти антропоморфы остаются в центре оживленных дискуссий.

Австралопитеки (рис. 95), обитавшие в Южной и Восточной Африке во второй половине плиоцена и начале плейстоцена, во Многих отношениях были гораздо ближе к людям, чем любые человекообразные обезьяны. Судя по строению их таза и скелета задних конечностей, австралопитеки постоянно использовали двуногое передвижение, хотя оно было значительно менее совершенным, чем у современных людей. Пропорции их черепа (см. рис. 94) и особенности зубной системы также более сходны с человеческими, чем с обезьяньими. Средний объем мозговой коробки австралопитеков, однако, оставался близким к таковому человекообразных обезьян - в среднем 520 см3, достигая в отдельных случаях 650 см3.
Вероятно, австралопитеки достаточно часто использовали мясную пищу, охотясь на различных животных с помощью камней и тяжелых костей крупных копытных.
Размеры тела этих приматов были сравнительно невелики: по расчетам, длина их тела составляла 133-154 см при среднем весе 36-55 кг. Средняя продолжительность жизни австралопитеков оценивается А.Манном в 17-22 года. По мнению этого ученого, в жизни австралопитеков большую роль играла передача опыта от одного поколения другому, что требовало достаточно длительного периода обучения детей.
Следует ли считать австралопитеков примитивными людьми или все же лишь высокоразвитыми человекообразными обезьянами и соответственно рассматривать их в составе понгид или же гоминид? Ведь объем их мозга и ряд особенностей строения оставались гораздо более близкими к состоянию обезьян, чем людей. Дискутируется в современной антропологии и другой вопрос: были ли австралопитеки прямыми предками людей или представляли слепую боковую ветвь, развивавшуюся параллельно ветви настоящих гоминид? Некоторые ученые предлагали рассматривать австралопитеков как подсемейство в пределах семейства понгид, другие - как подсемейство в составе гоминид (эта точка зрения получила наибольшее распространение), третьи - как особое независимое семейство среди антропоморфных приматов.
Дискуссия особенно обострилась после ряда замечательных палеонтологических находок, сделанных начиная с 1959г. английским антропологом Луисом Лики, его сыном Ричардом Лики и рядом других ученых в Восточной Африке, особенно в ущелье Олдовай (северная часть Танзании) и на восточном побережье озера Рудольф. Здесь в отложениях с возрастом 1-1,8 млн лет были обнаружены ископаемые остатки высокоразвитого австралопитека - зинджантропа (A. boisei), а также другого антропоморфного примата, которому дали название "человек умелый" (Homo habilis), противопоставляя его австралопитекам как представителя настоящих людей (род Homo). Основаниями для этого были некоторые прогрессивные черты в строении скелета конечностей и несколько больший объем мозговой коробки (680-700 см3), чем у типичных австралопитеков. Вместе со скелетами зинджантропа и "человека умелого" были найдены и примитивные каменные орудия, сделанные из грубо обколотых ударами галек кварца, кварцита, лавы (так называемая "олдовайская галечная культура"), а также кости животных, часть из которых была расколота древними людьми, вероятно, для добывания костного мозга. По мнению Лики, эти орудия принадлежали не зинджантропу, а примату, названному им "человек умелый". Лики рассматривал австралопитеков как слепую боковую ветвь эволюции, представители которой сохранили более примитивное строение, чем современные им древнейшие гоминиды Homo habilis.
Однако многие антропологи не согласны с принадлежностью "человека умелого" к роду Homo, указывая на морфологическую близость этого примата к прогрессивным австралопитековым и рассматривая его как одного из их представителей. В целом нет серьезных оснований "отлучать" австралопитеков от орудий галечной культуры, тем более что примитивные орудия были обнаружены и в более древних отложениях с возрастом 2-3 млн лет.
Однако на восточном берегу озера Рудольф были обнаружены остатки австралопитеков совместно с таковыми еще более высокоразвитого, чем "человек умелый", примата с емкостью мозговой коробки 800-900 см3. В отношении этой находки уже не могло быть сомнений в ее принадлежности к роду Homo.
Судя по этим данным, типичные плейстоценовые австралопитеки действительно существовали одновременно с более прогрессивными гоминидами и поэтому не могут рассматриваться как предки последних. Тем не менее остается вполне вероятным общее происхождение всех плейстоценовых гоминид, включая австралопитеков, от каких-то позднеплиоценовых предков, которыми вполне могли быть более ранние австралопитековые, пока еще мало известные. Во всяком случае, в морфологическом отношении австралопитеки занимают промежуточное положение между высшими гоминидами и человекообразными обезьянами (понгидами) и дают представление о первом важном этапе на пути гоминизации.
Что же касается вопроса о том, рассматривать ли австралопитеков как высших человекообразных обезьян или как наиболее примитивных людей, очевидно, решить его удалось бы при наличии четких критериев, определяющих принадлежность к людям. В качестве таковых указывались: постоянное использование двуногого передвижения, выпрямленное положение тела, увеличение первого пальца руки со сложной дифференциацией его мышц, увеличение головного мозга и относительных размеров больших полушарий, некоторые особенности зубной системы. По некоторым из этих показателей австралопитеки ближе к людям, по другим - к человекообразным обезьянам, занимая в целом промежуточное положение между теми и другими. М.И.Урысон отметил отсутствие морфологических критериев, с помошью которых можно было бы однозначно определить принадлежность ископаемых скелетных остатков к людям или к высокоразвитым обезьянам, и подчеркнул, что важнейшим показателем в этом плане являются найденные вместе со скелетными остатками орудия труда. Суди по их систематическому использованию австралопитеками, последних целесообразнее рассматривать как древнейших людей. Альтернативную точку зрения аргументировал А.А.Зубов, полагающий, что критерием должна служить не сама по себе трудовая деятельность, а тот "отпечаток", который она накладывает на морфологический облик гоминид, т. е. результат приспособления к трудовой деятельности различных органов (прежде всего головного мозга и передних конечностей).
Очевидно, переход к изготовлению орудий произошел на анатомической основе, характерной для австралопитеков и еще очень близкой к таковой высших человекообразных обезьян. Вероятно, на этом этапе гоминизации главную роль сыграло не столько увеличение объема мозга, размеры которого близки у австралопитеков и понгид, сколько изменение качества его работы, связанное со значительным возрастанием числа межнейронных связей и обусловившее существенные перестройки поведения, в частности широкое использование орудийной деятельности.
Трудовая деятельность стала важнейшим фактором дальнейшей эволюции человека. Поскольку использование орудий при высокоразвитой социальности, начинавшей формироваться еще у низших антропоидов, давало человеку огромные преимущества и позволяло осваивать новые местообитания и новые природные ресурсы, естественный отбор благоприятствовал таким изменениям организации древних людей, которые способствовали совершенствованию трудовой деятельности и социального поведения. Это были в первую очередь прогрессивные изменения структуры и массы головного мозга, а также мускулатуры и скелета передних конечностей, с развитием механизмов тонкой нервной координации движений. Отбор благоприятствовал развитию у людей таких форм поведения, которые облегчали их общение в процессе трудовой деятельности, а также в организации совместных охот или защиты от нападений хищников. В связи с этим совершенствовались способы обмена информацией, в частности система звуковой сигнализации, развитие которой привело к формированию членораздельной речи, а на ее основе - второй сигнальной системы, сыгравшей столь огромную роль в развитии человеческого интеллекта и культуры.
По мнению Я.Я.Рогинского, сообщества древних людей подвергались групповому отбору, благоприятствовавшему сохранению тех коллективов, в которых преобладали более развитые в социальном отношении индивиды. Это выражалось в совершенствовании тормозных механизмов мозга, позволявших снизить взаимную агрессивность, а также в развитии свойств, способствовавших обогащению знаний на основе своего и чужого опыта.
Важнейшей чертой человеческого общества является наличие фонда социальной или культурной информации, биологически не наследуемой и передающейся от поколения к поколению посредством обучения (а на более поздних этапах развития общества кодируемой письменно) и в форме созданных предыдущими поколениями орудий труда и других материальных и культурных ценностей. Рост и развитие этого социального фонда (или фонда материальной культуры) постепенно уменьшает зависимость человеческого общества от природы. Это не могло не привести к существенным изменениям самого характера эволюционных преобразований человека. Для любой человеческой популяции фонд материальной культуры, накопленный предшествовавшими поколениями, является, в сущности, важнейшей частью среды ее обитания. Естественный отбор приспосабливал человеческие коллективы к этой их специфической среде - отбор в пользу индивидов, более способных к обучению и трудовой деятельности, и групповой отбор в пользу коллективов, в которых преобладали индивиды с более развитым социальным Поведением. Зато снижалось давление отбора на совершенствование и поддержание приспособлений, повышающих устойчивость отдельных особей к неблагоприятным факторам среды, Поскольку фонд материальной культуры, социальная организация и трудовая деятельность опосредовали влияние неблагоприятных факторов внешней среды, играя своего рода защитную роль.
С другой стороны, развитие человеческого общества все в большей степени начинает регулироваться специфическими социальными закономерностями, которые сложным образом взаимодействуют с биологическими. В полной мере эта специфика эволюции человека стала проявляться лишь на более поздних этапах его филогенеза, особенно после появления человека современного вида, Homo sapiens.
Эволюция рода Homo
Филогенетическую историю гоминид, в ходе которой постепенно формировались особенности современного человека, разделяют на ряд последовательных стадий: 1) "предшественники людей", или протоантропы; 2) архантропы; 3) палеоантропы; 4) неоантропы.
Протоантропы были представлены австралопитеками, существовавшими в позднем плиоцене и плейстоцене 5,5-1,0 млн лет назад, но, возможно, возникшими еще раньше. Австралопитеки создали костяную и галечную олдовайскую культуру. Наиболее существенные морфологические преобразования на данном этапе гоминизации происходили в скелете и мускулатуре задних конечностей. Эти изменения были связаны с переходом к постоянному двуногому передвижению. Эта стадия антропогенеза была рассмотрена нами в предыдущем разделе.
Архантропы известны по многочисленным находкам, сделанным в самых различных уголках Старого Света. Первая из этих находок была сделана еще в 1891 г. Е.Дюбуа, обнаружившим на острове Ява ископаемые остатки существа, получившего название питекантроп или "обезьяночеловек". Первоначально разные архантропы, как и австралопитеки, рассматривались как представители разных родов: питекантропы с Явы, синантропы из ряда местонахождений на территории Китая, гейдельбергский человек в Европе, атлантропы в Северной Африке и др. Ныне большинство ученых считают всех архантропов принадлежащими к одному виду Homo erectus - "человек выпрямленный", в пределах которого выделяют до 9 подвидовых форм. До недавнего времени считали, что архантропы существовали на Земле в среднем плейстоцене, со времени гюнцского и до миндельского оледенения, т.е. 700-300 тыс. лет назад, однако новые данные (например, упомянутые выше находки, сделанные Р. Лики на восточном берегу озера Рудольф) и уточнение датировок расширили время существования архантропов до огромного диапазона 2,6-0,2 млн лет назад-В течение столь длительного времени архантропы не оставались неизменными. На этой стадии антропогенеза были сделаны важные шаги по пути морфофизиологического прогресса. Так. емкость мозговой коробки увеличилась с 750-800 см3 (у древнейших архантропов, подобных лантьянскому синантропу, существовавшему около 700 тыс. лет назад, или еще более древнему питекантропу с берегов оз. Рудольф) до 1225 см3 (у синантропов из пещеры Чжоукоудянь, живших около 300 тыс. лет назад).
Однако все архантропы сохраняли целый ряд очень примитивных (для человека) признаков (см. рис. 94): очень покатый лоб, переходящий в низкий черепной свод; сильно выступающий надглазничный валик, позади которого череп был сжат поперечно (заглазничное сужение); выступающие вперед челюсти; отсутствие подбородочного выступа. Общий облик архантропов (рис. 96) был уже несомненно человеческим, хотя телосложение оставалось очень грубым, а походка была более неуклюжей, чем у современных людей.
Архантропы питались и мясной, и растительной пищей, хотя первая в их рационе, вероятно, преобладала. Вместе с остатками некоторых архантропов были обнаружены каменные и костяные орудия, более совершенные, чем олдовайские, но в целом еще очень примитивные (культура раннего палеолита). В пещере Чжо-Укоудянь (в 60 км к югу от Пекина) вместе с многочисленными Ископаемыми остатками синантропов были обнаружены каменные и костяные орудия и следы костров. Многие найденные здесь же кости животных имели следы обжига. Использование огня для обогревания пещер, служивших жилищем, и для приготовления пищи, несомненно, было важным шагом вперед, свидетельствующим о достаточно высоких познавательных и манипулятивных возможностях архантропов. Различные архантропы жили не только в пещерах, но и в лесах, и в саваннах. Вероятно, продолжительность жизни этих древних людей была небольшой; немногие из них доживали до 30-32 лет.
Следующая стадия в эволюции гоминид, палеоантропы, представлена так называемыми неандертальцами (Homo neander-thalensis), видовое название которых связано с первой находкой ископаемых остатков этих людей в долине Неандерталь вблизи Дюссельдорфа. Неандертальцы, как и архантропы, были распространены практически по всей территории Старого Света и очень разнообразны. Они появились на Земле около 300 тыс. лет назад (во время миндель-рисского межледниковья) и просуществовали до первой половины вюрмского оледенения, т. е. примерно до 35 тыс. лет назад.
Палеоантропы достигли огромного прогресса в увеличении массы головного мозга. Объем мозговой коробки мужчин-неандертальцев в среднем составлял около 1550 см3, достигая 1600 см3. Размеры головного мозга, достигнутые неандертальцами, в ходе последующей эволюции при достижении стадии неоантропов далее не увеличивались, хотя происходили перестройки структуры мозга.
Несмотря на объемистую мозговую коробку, череп неандертальцев (см. рис. 94) сохранял еще многие примитивные особенности: покатый лоб, низкий свод и затылок, массивный лицевой скелет со сплошным надглазничным валиком, подбородочный выступ был почти не выражен, сохранялись крупные зубы. Пропорции тела палеоантропов (рис. 97) были в целом близки к таковым современного человека. По сравнению с архантропами у палеоантропов усовершенствовалось строение кисти. Средний рост неандертальцев составлял 151 - 155 см.
Палеоантропами была создана культура среднего палеолита. Неандертальцы хоронили своих умерших с погребальными обрядами, что позволяет предполагать у них наличие достаточно развитого абстрактного мышления.
Стадия неоантропов соответствует человеку современного вида - Homo sapiens (человек разумный). Древнейших неоантропов традиционно называют кроманьонцами по месту первой находки их ископаемых остатков в гроте Кро-Маньон, на территории французской провинции Дордонь. Кроманьонцы уже вполне соответствовали антропологическому типу современного человека (рис. 98), отличаясь лишь незначительными особенностями (несколько менее высокий свод черепа, сильнее развитая зубная система и др.). Кроманьонцы известны начиная со времени среднего вюрмского оледенения в позднем плейстоцене около 38-40 тыс. лет назад. Однако, по некоторым данным, организация неоантропов начала формироваться еще раньше, и древнейшие неоантропы могли существовать уже 40-50 тыс. лет назад.
Средний объем черепномозговой полости у неоантропов составляет 1500 см3, т.е., как мы уже отмечали, увеличение размеров головного мозга прекратилось после достижения стадии палеоантропов. Очевидно, этот объем мозга оказался достаточным для всего последующего усложнения высшей нервной деятельности человека, вплоть до наших дней. Более того, головной мозг современного человека, объем которого не превышает такового неандертальцев, по данным физиологов, сохраняет огромные ресурсы нервных клеток, с возможностью возникновения еще большего количества нервных связей, которые остаются неиспользованными в течение всей жизни индивида (еще один пример конструктивной избыточности структур организма, см. с. 138-140).
Основные морфологические преобразования, происшедшие в процессе формирования неоантропов, выражаются в некоторых структурных изменениях головного мозга и черепа (см. рис. 94), особенно его лицевого отдела (относительное уменьшение челюстей, образование подбородочного выступа, редукция надглазничного валика и заглазничного сужения, увеличение высоты черепного свода и т.п.).
Кроманьонцы были создателями культуры позднего палеолита, характеризующейся высоким совершенством обработки камня и кости. Именно кроманьонцы были творцами пещерных рисунков, запечатлевших животных мамонтовой фауны, а также древнейших скульптурных изображений и первых музыкальных инструментов. Можно поэтому утверждать, что с неоантропами возникает искусство.
Подчеркнем еще раз, что каждая из рассмотренных нами стадий эволюции человека включала большое количество вариаций - как в пространстве (в разных регионах), так и во времени. Характерные особенности следующей стадии не возникали внезапно и все сразу, но постепенно развивались у разных популяций, так сказать, "в недрах" предыдущей стадии антропогенеза. При этом различные признаки, в соответствии с правилом Осборна, изменялись своими темпами, и в разных популяциях возникали различные комбинации более прогрессивных и архаических особенностей.
Такая сложность общей картины антропогенеза при значительном количестве палеонтологических и археологических данных и некотором перекрывании во времени каждой пары последовательных стадий (т.е. сосуществовании поздних протоантропов с ранними архантропами, поздних архантропов с ранними палеоантролами и т.д.) создала предпосылки для разработки разных концепций эволюции человека.
Согласно теории пресапиенса, выдвинутой в 50-е гг. нашего века Г.Хеберером, А.Валуа, А.Тома и др., архантропы и палеоантропы не были предками неоантропов, все эти три вида рода Homo независимо друг от друга произошли от протоантропов в начале плейстоцена. Непосредственным предком неоантропов был так называемый "пресапиенс", который никогда не обладал такими характерными чертами архантропов и палеоантропов, как надглазничный валик, низкий лоб и черепной свод и т. д. Основные аргументы в пользу этой концепции связаны с трудностями, возникающими при попытках вывести неоантропов из некоторых подвидов палеоантропов (в частности, из западноевропейских, так называемых "классических" неандертальцев), обладающих очень массивным лицевым скелетом, сильно развитыми лобными пазухами, грубым и массивным посткраниальным скелетом. В качестве пресапиенса предлагались ископаемые остатки различных гоминид, начиная с пресловутого "пильтда-унского человека", позднее оказавшегося фальсификацией, и кончая среднеплейстоценовыми черепами из гротов Фонтешевад (Южная Франция) и Сванскомба (Южная Англия). Однако последующий анализ этих ископаемых остатков не подтвердил их принадлежности пресапиенсу. Один из черепных фрагментов из Фонтешевада принадлежал ребенку, а в детских черепах даже у низших антропоидов всегда сильнее выражены черты "сапиентности" (относительно большая мозговая коробка с куполообразным сводом, относительно слабо развитый лицевой отдел и надглазничный валик и т.п.). У второго черепного фрагмента из Фонтешевада определить наличие надглазничного валика невозможно. У сванскомбского черепа лоб и лицевая часть вообще не сохранились. В итоге концепция пресапиенса оказывается лишенной серьезных аргументов: при изобилии на всех континентах Старого Света остатков архантропов и палеоантропов, "преса-пиенсы" повсюду отсутствуют.
Значительно лучше обоснована теория последовательных стадий антропогенеза, разделяемая ныне большинством ученых. Согласно этой концепции, эволюция гоминид происходила в направлении от протоантропов к неоантропам через стадии архантропов и палеоантропов, с достижением на каждой стадии антропогенеза нового уровня гоминизации и, соответственно, с созданием новой, более совершенной культуры. Перекрывание стадии объясняется либо формированием новой стадии в каком-то одном центре внутри ареала предшествовавшей и дальнейшим постепенным вытеснением представителей предковой, менее совершенной формы в других регионах, либо независимой параллельной эволюцией разных филетических линий гоминид, каждая из которых могла проходить последовательные стадии своими темпами. Эти два варианта прохождения последовательных стадий антропогенеза получили соответственно наименования теорий моно- и полицентризма.
Согласно представлениям моноцентризма, каждый новый вид рода Homo, соответствовавший новой стадии антропогенеза, формировался в пределах какой-то определенной части ареала предкового вида и затем расселялся из этого центра, постепенно вытесняя предковую форму и отчасти гибридизируясь с нею. Так, в концепции широкого моноцентризма прародиной неоантропов считают обширную территорию, включавшую Переднюю Азию и, возможно, Юго-Восточную Европу (некоторые ученые в качестве прародины неоантропов рассматривают Африку). Основанием для этого является, с одной стороны, наличие ряда прогрессивных особенностей (слабо развитый надглазничный валик, более высокий лоб и др.) у неандертальцев, населявших указанные области, с другой - уже упоминавшиеся трудности, возникающие при попытках вывести неоантропов от ряда периферических популяций палеоантропов, в частности от классических неандертальцев Западной Европы. Причина этих затруднений в наличии у последних специфических особенностей, которые, возможно, возникли в связи с развитием приспособлений к жизни в суровых условиях приледниковой зоны.
Сторонники теории полицентризма (впервые обоснованной Ф. Вайденрайхом в 1939-1943 гг.) обращают внимание на следующие факты:
1) широкое расселение представителей каждой стадии антропогенеза в Старом Свете и дифференциация местных форм (подвидов) в разных регионах Европы, Азии и Африки;
2) наличие у представителей древних территориальных групп специфических особенностей строения, которые в некоторых случаях прослеживаются в данном регионе от стадии архантропа до неоантропа (например, уже у синантропов в Юго-Восточной Азии была выражена особая "лопатообразная" форма верхних резцов, которая характерна и для представителей современной монголоидной расы);
3) частая встречаемость явлений параллельной эволюции, хорошо аргументированная фактами по филогенезу различных групп организмов;
4) равномерное и параллельное развитие культуры (по археологическим данным) на всей территории Старого Света; при этом Не наблюдалось резких и внезапных изменений культуры, которых следовало бы ожидать при вытеснении, например, классических неандертальцев кроманьонцами, вторгшимися в Западную Европу, напротив, повсеместно среднепалеолитическая культура постепенно преобразуется в верхнепалеолитическую.
По этим причинам теория полицентризма считает более вероятной параллельную эволюцию нескольких филетических линий гоминид с независимым приобретением особенностей последующих стадий, от архантропов до неоантропов, на разных континентах Старого Света. Современные большие расы человека -европеоиды, негроиды, монголоиды и австралоиды - выводятся при этом от разных подвидов палеоантропов или даже архантропов.
В Новый Свет человек проник относительно недавно, около 25-30 тыс. лет назад, как предполагают, уже на стадии неоантропа.
Аргументом против крайнего полицентризма (понимаемого как длительная, полностью независимая и параллельная эволюция разных филетических линий) является биологическое единство современного человечества, представляющего собой единый биологический вид, в пределах которого происходило и происходит свободное скрещивание расовых группировок любого ранга. В этом отношении человек отличается от большинства других видов организмов, у которых внутривидовая дифференциация с возникновением подвидов обычно ведет к обособлению некоторых из них в качестве новых филетических линий, теряющих генетическую связь с родоначальной формой. Может быть, эта особенность человека как-то связана со специфическим характером его эволюции, определяемым наличием особой социальной среды и фонда материальной культуры, опосредующих взаимоотношения людей с внешним миром и позволяющих человеку приспосабливаться к изменениям внешних условий, не изменяя своих основных биологических характеристик.
Расы человека по своему происхождению соответствуют подвидам, т.е. крупным аллопатрическим популяциям, которые складываются в различных областях видового ареала, в какой-то мере обособленных друг от друга. Обычно подвиды приобретают под контролем естественного отбора особенности, являющиеся приспособительными к условиям их обитания. Некоторые отличительные особенности больших человеческих рас поддаются истолкованию как приспособительные признаки. Так, темная пигментация кожи у представителей негроидной и австралоидной рас, очевидно, является адаптацией к жизни в тропическом поясе, защищая организм от ультрафиолетового облучения. Точно так же курчавые волосы, образующие густую "шапку", предохраняют голову от перегрева, а характерные для южных рас пропорции тела с относительно коротким туловищем и длинными конечностями более благоприятны для увеличения теплоотдачи. Противоположный тип телосложения монголоидной расы позволяет, напротив, уменьшить теплоотдачу за счет уменьшения отношения поверхности тела к его массе, что может иметь приспособительную ценность в условиях резко континентального климата Центральной Азии, с сильными морозами и ветрами в зимний период. Характерный узкий разрез глаз представителей монголоидной расы, прикрытых продольными складками верхних век, возможно, уменьшает риск засорения глаз пылевыми частицами при сильном ветре. Труднее объяснить приспособительный смысл основных признаков европеоидной расы (светлая кожа, высокая переносица и длинный узкий нос, обильный волосяной покров и др.). Возможно, некоторые из них, как и вообще многие внешние отличительные расовые признаки у человека, не имея собственной приспособительной ценности, возникли коррелятивно как результат плейотропного эффекта генов или взаимодействия разных мор-фогенетических систем в онтогенезе при отборе каких-либо приспособительно важных, но менее заметных признаков (например, большей устойчивости к простудным заболеваниям или артритам, столь частым во влажном и прохладном климате Европы).
Так или иначе, но многие из расовых отличий сами по себе, вероятно, не имеют существенной адаптивной ценности. На этом основании ряд ученых считают, что закрепление таких признаков в процессе расогенеза происходило при участии дрейфа генов. Повышению роли дрейфа генов при обособлении малых популяций человека должно было способствовать снижение интенсивности естественного отбора по многим фенотипическим признакам, адаптивная роль которых была в значительной степени компенсирована развитием фонда материальной культуры.
Согласно теории полицентризма, современные расы человека возникли в результате длительной параллельной эволюции нескольких филетических линий на разных материках: европеоидная в Европе, негроидная в Африке, монголоидная в Центральной и Восточной Азии, австралоидная в Австралии. Однако если эволюция расовых комплексов и шла параллельно на разных континентах, она не могла быть полностью независимой, поскольку древние проторасы должны были скрещиваться на границах своих ареалов и обмениваться генетической информацией. В ряде областей сформировались промежуточные малые расы, характеризующиеся смешением признаков разных больших рас. Так, промежуточное положение между европеоидной и монголоидной расами занимают южносибирская и уральская малые расы, между европеоидной и негроидной - эфиопская и т. д.
С позиций моноцентризма современные человеческие расы Нормировались относительно поздно, 25-35 тыс. лет назад, в Процессе расселения неоантропов из области их возникновения.
При этом также допускается возможность скрещивания (хотя бы ограниченного) неоантропов во время их экспансии с вытесняемыми популяциями палеоантропов (как процесса интрогрессивной межвидовой гибридизации) с проникновением аллелей последних в генофонды популяций неоантропов. Это также могло способствовать расовой дифференциации и устойчивости некоторых фенотипических признаков (подобных лопатообразным резцам монголоидов) в центрах расообразования.
Существуют и компромиссные между моно- и полицентризмом концепции, допускающие расхождение филетических линий, ведущих к разным большим расам, на различных уровнях (стадиях) антропогенеза: например, более близких друг к другу европеоидов и негроидов уже на стадии неоантропов с первоначальным развитием их предкового ствола в западной части Старого Света, тогда как еще на стадии палеоантропов могла обособиться восточная ветвь - монголоиды и, может быть, австралоиды.
В целом, эволюция рода Homo наиболее точно соответствует модели сетчатой эволюции (см. с. 79-80). Во всяком случае, какую бы из рассмотренных нами концепций антропогенеза ни принимать, фактом является биологическое единство современного человечества, будущее развитие которого (после ликвидации расовых барьеров, имеющих историческое и социальное происхождение), вероятно, пойдет по пути постепенного слияния всех рас.
Обзор филогенеза организмов, сделанный в третьей части книги, позволяет выявить ряд закономерностей, проявляющихся лишь в масштабе макроэволюции и характеризующих ее специфику по отношению к микроэволюции. Среди них наиболее обращает на себя внимание неравномерность темпов макроэволюции, отмечавшаяся многими учеными (А.Н.Северцов, Д.Симпсон, О.Шиндевольф и др.). Формирование основной совокупности признаков нового крупного таксона или, другими словами, становление нового типа организации - типогенез - происходит постепенно, но в относительно краткие (в масштабах макрофилогенеза) сроки - от нескольких миллионов до 20-30 млн лет. Сформировавшийся в период типогенеза основной план организации данного таксона в дальнейшем остается постоянным у представителей разных филетических линий в течение значительных промежутков времени (многие десятки и даже сотни миллионов лет), составляющих период типостаза. Лишь немногие филетические линии в пределах данного таксона могут спустя длительное время вступить в новый период типогенеза того же масштаба. Чередование периодов типогенеза и типостаза придает макроэволюции своеобразный пульсирующий характер.
В то же время микроэволюционные процессы идут непрерывно - как в период типогенеза, так и во время типостаза. На их основе происходит адаптивная радиация - разветвление филетических линий с возникновением новых видов, связанное с развитием различных частных приспособлений. Закон адаптивной радиации, согласно которому эволюция любого филогенетического ствола сопровождается его разветвлением на большее или меньшее число обособленных филетических линий, был сформулирован независимо друг от друга В.О.Ковалевским и Г.Осборном. Адаптивная радиация является основной формой эволюционных изменений в период типостаза, происходит она и при типогенезе.
Для иллюстрации этих эмпирических обобщений рассмотрим филогенез высших наземных позвоночных - амниот. Типогенез организации рептилий происходил во второй половине раннекарбоновой и в среднекарбоновой эпохе, в течение примерно 25-30 млн лет. Последующие 300 млн лет в филогенезе пресмыкающихся происходила интенсивная адаптивная радиация, но в большинстве филетических линий общий тип организации пресмыкающихся не изменился (период типостаза). Некоторые филетиче-ские линии высших зверообразных рептилий (териодонтов) в позднепермское время вступили в новый период типогенеза, продолжавшийся до позднего триаса (30-35 млн лет). За это время сложились основные черты организации млекопитающих. В последующие 200 млн лет в филогенезе млекопитающих наблюдалась главным образом адаптивная радиация; более поздние преобразования общего значения (развитие живорождения, совершенствование аппарата передвижения, усложнение головного мозга и др.) стали признаками подклассов высших млекопитающих - сумчатых и плацентарных. Точно так же типогенез птиц протекал на протяжении второй половины юрского периода (30-35 млн лет), тогда как в последующие 140 млн лет происходила интенсивная адаптивная радиация этого класса, сопровождавшаяся некоторыми "доводками" основных особенностей организации птиц.
Вторая группа общих закономерностей макроэволюции связана с проблемой направленности эволюционного процесса. На уровне микроэволюции эта закономерность практически не проявляется. Однако в макрофилогенезе различных групп организмов обнаруживаются некоторые устойчивые тенденции (общие направления) эволюционных преобразований, как бы "проступающие" сквозь разнообразие частных вариантов, которые возникают в процессе адаптивной радиации. При этом для разных филетических линий в пределах данного крупного таксона характерна параллельная эволюция (или эволюционный параллелизм), с независимым приобретением разными видами сходных признаков. Напомним рассмотренные в третьей части книги многочисленные примеры параллельной эволюции самых различных групп организмов - проангиоспермов, членистоногих, кистеперых рыб, зверообразных пресмыкающихся и т.п.
В качестве наиболее общего проявления направленности макроэволюции рассматривается общее усовершенствование (повышение уровня) организации, или морфофизиологический прогресс, выражающийся в повышении степени дифференциации и интеграции организма, рационализации устройства его систем, интенсификации функций и в некоторых других показателях. Морфофизиологический прогресс в различных формах наблюдается в подавляющем большинстве крупных филогенетических стволов, хотя конкретные его проявления у разных групп существенно различаются, будучи связаны с особенностями их организации и адаптации.
В отличие от микроэволюции макроэволюция необратима. Закон необратимости эволюции был сформулирован в 1893 г. палеонтологом Л. Долло в несколько абсолютизированной форме: организм ни целиком, ни даже отчасти не может вернуться к состоянию, уже осуществленному в ряду его предков. В действительности по отношению к отдельным признакам эволюционная реверсия возможна (посредством обратных мутаций и некоторых других механизмов), но для организма как целого закон необратимости эволюции, безусловно, соблюдается. Даже при возвращении данной филетической линии в среду обитания, когда-то оставленную предками современных форм, при повторном развитии адаптации к этой исходной среде обычно не происходит восстановления утраченных органов и их аналоги развиваются заново - обычно из других элементов. Например, опорные структуры хвостового и спинного плавников у ихтиозавров и китообразных образованы плотной соединительной тканью в отличие от хрящевых или костных скелетных элементов в плавниках рыб.
Все перечисленные закономерности (пульсирующий характер и неравномерность темпов макроэволюции; ее направленность, выражающаяся в поступательном морфофизиологическом прогрессе, устойчивых эволюционных тенденциях и параллелизмах; адаптивная радиация; необратимость эволюции) обнаруживаются в макрофилогенезе практически любой крупной группы организмов, с достаточной полнотой представленной в палеонтологической летописи. Теория макроэволюции должна дать научное объяснение всем этим эмпирически установленным закономерностям. Но прежде чем мы приступим к анализу этих проблем, необходимо остановиться на механизмах эволюционных преобразований индивидуального развития - онтогенеза, которые являются своего рода связующим звеном между элементарными механизмами эволюции, рассмотренными во второй части книги, и проблемами макрофилогенеза.
Дифференцировка организма в онтогенезе
Одной из основных проблем онтогенетики, решение которой должно сыграть важную роль и для эволюционной теории, остается сущность явлений онтогенетической дифференцировки, т. е. тех процессов, которые ведут к развитию разнокачественности различных фенотипических структур, возникающих в онтогенезе, в конечном итоге, из единственной клетки - зиготы. Даже в организации столь просто устроенных животных, как кишечнополостные, насчитывается около 10 разных типов клеток, а в организме млекопитающих - порядка 1000 типов клеток. Клетки разных типов группируются в ткани и формируют многотканевые комплексы со сложной пространственной структурой - органы.
По современным представлениям, в процессе дробления зиготы на бластомеры и при последующих делениях соматических клеток каждая дочерняя клетка в норме получает 100 % наследственной информации, т.е. полный набор генов, точно соответствующий исходному генотипу зиготы. Однако соматические клетки резко различаются по строению и функциям (достаточно сравнить, например, нейрон, мышечную клетку и эритроцит в одном организме).
Можно считать установленным, что различия соматических клеток определяются дифференциальной активностью генов (другими словами, в одних типах клеток активны одни, а в других - другие участки хромосом). Неактивные в данной клетке гены блокируются какими-то входящими в состав хромосом веществами, какими именно - еще точно не выяснено. Предполагают, что этими веществами (репрессорами) являются те или иные хромосомные белки (многие ученые считают репрессорами белки-гистоны).
Очевидно, при эквивалентности генома всех зародышевых клеток их первоначальная дифференцировка должна определяться разнокачественностью их цитоплазмы. В период дробления практически не происходит роста клеток между их последовательными делениями. Поэтому в разные бластомеры попадают различные и, вероятно, разнокачественные участки цитоплазмы зиготы-Возможно, разнокачественность цитоплазмы и определяет активацию различных генов в разных бластомерах. Деятельность активированных генов приводит к изменениям цитоплазмы бластоме-ров - специфическим в разных клетках. Измененное цитоплазма-тическое окружение взаимодействует с геномом клетки, вызывая избирательную активацию новых групп генов и репрессируя другие группы. Так запускается цепь последовательных взаимодействий между геномом и цитоплазмой, протекающая по принципу обратной связи (положительной для одних и отрицательной для других генных комплексов), в результате которых клетка дифференцируется в определенном направлении (рис. 99). Однако пока еще мало известно о конкретных взаимодействиях между ядром и цитоплазмой на разных этапах клеточной дифференцировки.
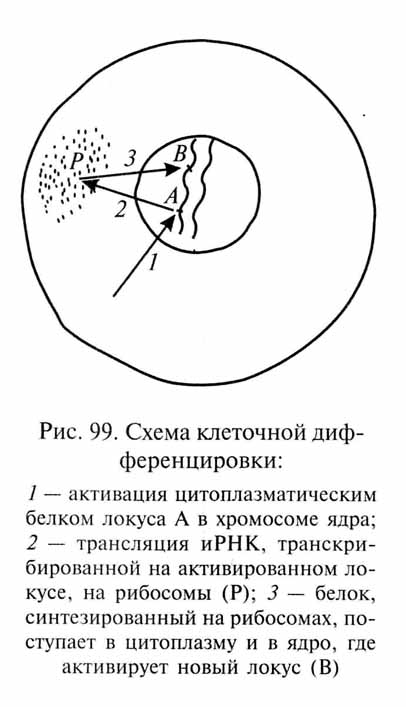
Онтогенез отнюдь не сводится к дифференциации разных типов клеток. Важнейшим аспектом индивидуального развития многоклеточных организмов является установление общего плана их строения, т. е. оформление пространственной структуры организма и определенного расположения его частей. У всех более сложных многоклеточных организмов к этому добавляются процессы органогенезов - формирование дифференцированных органов. Все ^эти процессы осуществляются посредством взаимодействия друг с другом отдельных клеток и клеточных комплексов.
О сущности этих взаимодействий известно, пожалуй, еще меньше, чем о молекулярных механизмах клеточной дифференцировки, хотя исследования в этой области ведутся уже давно. Одной из Наиболее разработанных концепций является теория эмбриональной индукции, созданная трудами Г.Шпемана и его учеников.
Под эмбриональной индукцией понимается такое взаимодействие между двумя клеточными комплексами зародыша, при котором один из них - организатор или организационный центр - Направляет (индуцирует) дифференциацию другого - реагирующей ткани - в определенном направлении. В нормальных условиях, т. е. в живом зародыше, присутствие организатора необходимо Для формирования из реагирующих тканей специфических структур взрослого организма. Так, у зародышей позвоночных крыша первичной кишки, образованная зачатком хорды, индуцирует в расположенной над ней эктодерме развитие нервной системы; глазной бокал, растущий по направлению к покровам от зародышевого головного мозга, индуцирует развитие хрусталика глаза; зачаток внутреннего уха (слуховой пузырек) индуцирует развитие вокруг себя слуховой капсулы и т.п. Согласно теории эмбриональной индукции, дифференциация органов в индивидуальном развитии представляет собой цепь индукционных взаимодействий, в которую последовательно включаются новые и новые организаторы, и каждый предшествующий этап "запускает в действие" следующий.
Характер взаимодействий между морфогенетическими системами в процессах эмбриональной индукции может изменяться в филогенезе. Например, у эмбрионов ряда костных рыб (Amia, Salmo) невромасты сейсмосенсорных рецепторов (или органов боковой линии) в области головы играют роль индукторов для формирования накладных костей черепа. Однако у хрящевых рыб невромасты сейсмосенсорных рецепторов в области головы имеются, но накладные кости черепа не развиваются. Следовательно, индукционные взаимодействия здесь разорваны: либо невромасты потеряли свойства индукторов, либо реагирующие ткани утратили свою реактивность. В то же время у наземных позвоночных накладные кости черепа имеются, а органов боковой линии нет. Очевидно, в этом случае вместо старого механизма индукции возник новый. С другой стороны, некоторые индукционные системы поразительно консервативны, сохраняясь у очень далеких друг от друга групп - таков первичный хордомезодермальный организатор у всех хордовых.
Природа эмбриональной индукции еще не вполне выяснена. Индукция может основываться на непосредственном взаимодействии поверхностных мембран клеток организатора и реагирующей ткани, или на химической передаче индуцирующего действия, или на совокупности пространственных и химических взаимодействий. По некоторым данным, в химической индукции существенную роль может играть белковый компонент рибонуклеопротеи-
дов индуктора.
Явление эмбриональной индукции, несомненно, играет важную роль в онтогенезе, но отнюдь не исчерпывает всех форм взаимодействий между клеточными комплексами зародыша, обеспечивающих развитие структурной упорядоченности организма.
Недавно (80-90-е гг. XX в.) было показано, что важную роль в контроле процессов онтогенетического формирования пространственной организации тела зародыша играет особая категория генов-регуляторов, именуемых гомеобокссодержащими, или гомеобоксными. Гомеобокс представляет собой эволюционно консервативный генетический блок - последовательность из 180 нуклеотидов, обнаруженный в геноме филогенетически столь далеких друг от друга организмов, как позвоночные, иглокожие, насекомые и нематоды. Гомеобокс кодирует "гомеодомен" из 60 аминокислот. Протеины, содержащие этот гомеодомен, регулируют транскрипцию других генов. Гомеобоксные гены являются важными регуляторами, контролирующими онтогенетические программы, которые определяют формирование специфики разных сегментов вдоль передне-задней оси тела. Очевидно, мутации гомеобоксных генов могут иметь очень глубокие последствия для организма, приводя к изменениям соответствующих программ развития. Значительное большинство исследованных мутаций гомеобоксных генов (например, у мух-дрозофил) летальны, что вполне естественно для крупных мутаций. Эти обстоятельства определяют высокий эволюционный консерватизм гомеобоксных генов, сохраняющих значительную степень общности у филогенетически далеко разошедшихся групп организмов.
Проявление мутаций в фенотипе
Мутации проявляются фенотипически и подвергаются действию отбора, начиная с той стадии онтогенеза, когда мутантные гены оказываются активированными. Очевидно, общий фенотипический эффект данной мутации в среднем будет тем меньше, чем позднее в онтогенезе начинается функционирование мутантного гена. При этом фенотипический эффект мутации отнюдь не сводится к действию мутантного гена на признаки, им непосредственно контролируемые. Плейотропный (множественный) эффект влияния гена на фенотипические признаки складывается из взаимодействия различных морфогенетических систем в развивающемся организме, которые сами по себе могут иметь совершенно разную генетическую основу. Прекрасным примером этого может служить фенотипический эффект летальной мутации мышей, известной под названием "врожденная гидроцефалия". При этом у новорожденных мышей, погибающих от кровоизлияния в мозг, отечная лобная часть черепа заполнена геморрагической жидкостью; отсутствуют плоские накладные кости черепа; гипофиз имеет аномальную форму, выражены аномалии век и усов (вибрисс); морда укорочена. Сам же мутантный ген не влияет ни на один из этих признаков: он обусловливает лишь аномалии строения хрящевых клеток. Нарушения роста хрящевого эмбрионального черепа приводят к его укорочению, в результате которого мозг оттесняется кверху, что препятствует нормальному оттоку цереброспинальной жидкости. Это последнее приводит в конечном итоге ко всем перечисленным выше результатам (кровоизлияние в мозг, отсутствие плоских костей, аномалии век и др.). Мутация, обусловливающая сама по себе лишь аномалию хрящевых клеток, благодаря взаимодействиям различных зародышевых структур на эпигенетическом уровне, влечет за собой целую серию вторичных и третичных фенотипических эффектов, не относящихся непосредственно к компетенции мутантного гена.
Поскольку мутации со значительным фенотипическим эффектом (рано выявляющиеся в онтогенезе) имеют гораздо меньше шансов на сохранение, чем более поздно выявляющиеся малые мутации, онтогенез в целом приобретает тенденцию оставаться консервативным из поколения в поколение. В наибольшей степени это относится к основным процессам формообразования (установление общего плана строения, морфогенез важнейших органов), которые протекают чрезвычайно сходно у всех родственных видов в пределах очень крупных таксономических групп, таких, как классы и даже типы. С этим связано действие эмпирического обобщения, известного как "закон зародышевого сходства", сформулированного одним из крупнейших эмбриологов XIX в. К.Э. фон Бэром: "Развитие всех животных идет таким образом, что прежде всего определяется основной тип, причем гистологическая и морфологическая дифференцировки очень малы и только начинаются. Эмбрион при дальнейшем развитии этих дифференцировок переходит из своей основной формы в вариацию ее, т. е. из основной формы с характером данного класса переходит в таковую с характером отряда или семейства этого класса или в еще более ограниченную форму, пока наконец не появляются особенности индивидуума" (рис. 100).
Взаимоотношения онтогенеза и филогенеза
Проблему взаимоотношений индивидуального развития организмов и их эволюции затрагивал уже Ч.Дарвин, подчеркивавший влияние естественного отбора на все стадии индивидуального развития и формирование под контролем отбора специфических личиночных приспособлений. Позднее эту проблему разрабатывали Ф. Мюллер, Э. Геккель, В.Гарстанг, Г. де Вир и другие ученые.
В эволюционно-морфологическом плане соотношение онтогенеза и филогенеза наиболее полно было рассмотрено А.Н.Северцовым в его теории филэмбриогенезов (1912-1939). Согласно этой концепции филогенез не следует рассматривать как ряд сменянющих друг друга статических взрослых состояний организмов последовательных поколений (понимание филогенеза как эволюции взрослого состояния до сих пор очень распространено благодаря концентрации внимания ученых-неэмбриологов на взрослых организмах).
По А.Н.Северцову, филогенез представляет собой генетический ряд онтогенезов: эволюционируют не только фенотипы взрослых организмов, но и их онтогенезы, или кратко - филогенез есть эволюция онтогенеза. Изменчивость организмов проявляется на всех стадиях индивидуального развития и служит материалом для действия естественного отбора.
А.Н.Северцов разработал классификацию разных типов филогенетических изменений онтогенеза, которые он прежде всего разделил на ценогенезы и филэмбриогенезы. Ценогенезы представляют собой приспособления развивающегося организма к специфическим условиям эмбрионального или личиночного развития, которые не сохраняются во взрослом состоянии и не сказываются на особенностях последнего. Таковы, например, роговые зубы и жаберный фильтрационный аппарат личинок бесхвостых амфибий, плацента и зародышевые оболочки у эмбрионов млекопитающих и т.п. С другой стороны, филэмбриогенезы - это изменения онтогенеза, связанные с эволюционными преобразованиями каких-либо признаков взрослого организма. Филэмбриогенезы могут происходить как на самых поздних стадиях индивидуального развития (надставка, или анаболия), так и на средних (уклонение, или девиация) или на ранних (архаллаксис) стадиях. Из сказанного выше следует, что эволюция посредством анаболии в целом более обычна, чем посредством девиации и тем более архаллаксиса. При этом следует иметь в виду, что между разными формами филэмбриогенезов нет отчетливых границ. Более того, некоторые эволюционные изменения, первоначально возникшие как изменения поздних стадий онтогенеза (анаболии), могут в ходе дальнейшей эволюции сместиться на более ранние стадии, т. е. вторично преобразоваться в девиации и архаллаксисы. Это может быть связано с необходимостью более раннего выявления в онтогенезе нового адаптивного признака. Такой эффект достигается посредством более ранней онтогенетической активации соответствующих генов. Только на тех стадиях онтогенеза, которые не подверглись ценогенезам или филэмбриогенезам, сохраняется сходство с состоянием соответствующих стадий онтогенеза предков. Такое повторение предкового состояния - рекапитуляция - может сохраняться только для отдельных эмбриональных структур или процессов, но не для всего организма в целом, как полагал Э. Геккель. Обобщение Геккеля, известное как "основной биогенетический закон" (онтогенез есть сжатое и сокращенное повторение хода филогенеза), не имеет универсального характера и реально наблюдается лишь как частный, хотя и достаточно широко распространенный случай соотношений индивидуального и филогенетического развития - только для отдельных органов при условии их эволюционных преобразований по способу анаболии, т. е. изменения конечных стадий онтогенеза. Рекапитуляция целых стадий филогенетического развития для всего организма, которую постулировал "основной биогенетический закон" Э.Геккеля, крайне маловероятна.
В работах А.Н.Северцова и его учеников (Б.С.Матвеев, В.В.Васнецов, С.В.Емельянов и др.) были собраны многочисленные примеры различных форм эволюционных преобразований онтогенеза. Укажем лишь некоторые из них. По способу анаболии шла эволюция конечностей лошадиных (Equidae), что доказывается рекапитуляцией в онтогенезе конечностей современной лошади стадий Постепенной редукции боковых лучей конечности, весьма сходных с соответствующими филогенетическими стадиями (рис. 101). По способу анаболии, видимо, происходила и эволюция производных покровов у завропсид: ранние стадии морфогенеза пера птиц полностью соответствуют морфогенезу роговой чешуи рептилий, а последующие стадии развития пера как бы продолжают этот морфогенез дальше состояния, достигнутого у пресмыкающихся в роговых образованиях их кожи.

В качестве девиации А.Н.Северцов рассматривал преобразование брызгальца (древней жаберной щели, находившейся у рыбообразных предков наземных позвоночных между челюстной и подъязычной дугами висцерального скелета) в полость среднего уха тетрапод.
Примером архаллаксиса может служить значительное увеличение числа сегментов (сомитов) мезодермы (и соответственно их производных - позвонков, мышечных миомеров и т.п.) у змей по сравнению с их предками - ящерицами. Сегментация мезодермы происходит на ранних стадиях онтогенеза, почти одновременно с ее обособлением как зародышевого листка (стадия нейруляции).
Наконец, примеры ценогенезов назвал автор этого последнего термина Э. Геккель: это развитие амниона, аллантоиса, хориона и других эмбриональных и личиночных органов. Нужно при этом отметить, что Геккель понимал термин "ценогенез" гораздо шире, чем А.Н.Северцов: для Геккеля ценогенезами были любые изменения индивидуального развития, искажавшие ("фальсифицировавшие") консервативные признаки и процессы (или палингене-зы, по Геккелю), а не только провизорные эмбриональные органы, как понимал этот термин А.Н.Северцов. (Провизорные органы - структуры, существующие и функционирующие лишь в течение эмбрионального периода). В частности, в качестве ценогенезов Геккель рассматривал изменения темпов индивидуального развития различных органов, или гетерохронии. Как показали многочисленные более поздние работы (Ф.Кайбель, А.Н.Северцов, В.Гарстанг и др.), гетерохронии являются чрезвычайно распространенной формой эмбриональной изменчивости и предоставляют богатый материал для различных эволюционных преобразований - как ценогенезов, так и филэмбриогенезов. Гетерохронии могут изменять темпы эмбриональной дифферен-цировки различных органов и приводить к более ранней или более поздней их закладке в онтогенезе. Ускорение процесса эмбрионального развития какого-либо органа Геккель назвал акцелерацией, замедление - ретардацией.
Педоморфоз
Важное эволюционное значение может иметь педоморфоз - одна из форм эволюционных преобразований онтогенеза, основанная на гетерохрониях. Педоморфоз представляет собой генетически обусловленное (наследственное) сохранение во взрослом состоянии признаков более ранних стадий онтогенеза. Термин "педоморфоз" ("уподобление детенышу") был введен В.Гарстангом в 1922г.; его долгое время использовали преимущественно для обозначения сохранения во взрослом состоянии личиночных признаков. При наличии в жизненном цикле личиночной стадии, существенно отличающейся по строению, физиологии и образу жизни от взрослого организма, у некоторых видов в определенных условиях происходит задержка (ретардация) морфогенеза соматических структур при сохранении нормальных темпов развития половой системы. Личинки приобретают способность к размножению - половому (неотения) или партеногенетическому (педогенез). Эти явления представляют собой ненаследственные модификации онтогенеза: в других условиях личинки могут нормально метаморфизировать, превращаясь во взрослую стадию. Широко известным примером неотении являются аксолотли (рис. 102) - неотенические личинки американских саламандр - амбистом (Ambystoma tigrinum и др.). Неотения является результатом подавления активности эндокринных желез, стимулирующих метаморфоз (у земноводных - щитовидная железа). Воздействуя на аксолотлей соответствующими гормональными препаратами, можно добиться нормального метаморфоза - аксолотль превращается во взрослую амбистому.
При педоморфозе происходит наследственное изменение (уменьшение чувствительности соматических тканей личинки к гормонам, стимулирующим метаморфоз), что приводит к полной утрате взрослой стадии онтогенеза: жизненный цикл укорачивается, последней в нем становится стадия, которая была прежде личиночной. Такой педоморфоз произошел, например, в эволюции некоторых групп хвостатых земноводных (протеи, сирены, рис. 103). На основе педоморфоза сформировалась организация аппендикулярий (Appendiculariae) - одного из классов подтипа низших хордовых - оболочников (Tunicata). Вероятно, предки аппендикулярий имели жизненный цикл со свободноплавающей личинкой и прикрепленной неподвижной взрослой стадией, подобно современным асцидиям (Ascidiae). В результате педоморфоза у предков аппендикулярий прикрепленная взрослая стадия онтогенеза была утрачена, и строение современных представителей этой группы во многом остается близким к строению личинок асцидий (имеются хорда, сегментированные мышцы, спинной нервный ствол и др., рис. 104).

В последние десятилетия получило распространение широкое понимание термина "педоморфоз" как эволюционного изменения онтогенеза, связанного с сохранением во взрослом состоянии признаков любых более ранних стадий индивидуального развития, в том числе и неличиночных. При таком широком понимании термина, введенном С.Гулдом, педоморфоз включает также явление, которое описано Л.Больком в 1926 г. под названием "фетализация" - сохранение у взрослых организмов отдельных признаков эмбриональных стадий. Больк обратил внимание на наличие ряда фетализированных признаков у современного человека; к ним относятся, в частности, общие пропорции черепа с характерным куполообразным сводом и преобладанием мозговой коробки над челюстным аппаратом (см. рис. 94), а также некоторые другие особенности (форма ушной раковины, характер волосяного покрова). Среди этих признаков приспособительное значение имеет, вероятно, лишь фетализация общих пропорций черепа, которая создала у предков человека предпосылки, необходимые для вмещения и защиты головного мозга при прогрессировавшем увеличении его размеров. Фетализация других признаков могла развиться коррелятивно.
Некоторые аналогичные проявления фетализации наблюдаются у ряда пород домашних животных (собаки, кошки, рогатый скот), у которых в черепе, по сравнению с их дикими родственниками и предками, относительно большая мозговая коробка и относительно слабо развит челюстной аппарат. По наблюдениям К.Лоренца, для домашних животных характерно сохранение во взрослом состоянии определенной инфантильности поведения, благодаря которой эти животные признают "авторитет" человека как "патрона", "вожака стаи". Вероятно, в процессе искусственного отбора человек отбирал наиболее легко приручающихся животных, а таковыми были как раз те особи, которые сохраняли во взрослом состоянии черты инфантильности, т. е. обладавшие в некотором смысле "фетализированным поведением". Такое поведение должно основываться на соответствующих особенностях функционирования нервных центров. А фетализация последних может быть коррелятивно связана с указанными чертами строения черепа и других морфологических признаков, также приобретавших фетализированный облик. Так или иначе определенная степень фетализации поведения и ряда морфологических признаков характеризует многих домашних животных. Вероятно, отсутствие подобных особенностей поведения затрудняет приручение других диких животных, и в работе по созданию новых пород домашних животных следует учитывать необходимость отбора особей с наиболее инфантильным (фетализированным) поведением. Явления педоморфоза описаны и у растений: согласно А.Л.Taxтаджяну, у цветковых растений фетализированы многие органы спорофита (цветок, листья), а организация мужского и женского гаметофитов подверглась общему педоморфозу. (Нужно отметить, что А.Л.Тахтаджян, С.Гулд и ряд других ученых используют для обозначения педоморфоза или некоторых его форм термин "неотения", что приводит к путанице ненаследственных модификаций [неотения] и наследственных изменений онтогенеза [педоморфоз]).
Выделяют две основные формы педоморфных эволюционных изменений. Первая из них - акцелерационный педоморфоз, или прогенез, представляет собой результат ускоренного онтогенетического развития органов размножения, тогда как соматические структуры сохраняют прежние темпы морфогенеза. При этом организм в целом сохраняет ювенильный (или личиночный) облик, приобретая способность к размножению. После достижения половой зрелости обычно снижаются темпы морфогенеза и роста соматических структур, что приводит к обшей миниатюризации организма. Вероятно, таков был путь эволюции предков аппендикулярий. Возможно, посредством этого же эволюционного механизма были редуцированы мужской и женский гаметофиты у голосеменных и покрытосеменных растений. Вторая форма - ретардационный педоморфоз, или ретрогенез, представляет собой замедление темпов морфогенеза соматических систем организма при нормальном созревании половой системы и росте.
Вероятно, механизмы ретрогенеза могут быть различны на разных стадиях индивидуального развития в зависимости от факторов, контролирующих морфогенез на разных стадиях (например, при участии гормонов или еще до начала функционирования эндокринных желез). При этом особенно благоприятные условия для педоморфных преобразований создает личиночное развитие с метаморфозом, поскольку личинка представляет собой не только особую стадию онтогенеза, но и особый тип организации, отличающийся от взрослого организма и приспособленный к жизни в определенных условиях внешней среды, обычно в иных, чем взрослые особи того же вида. В связи с этим целесообразно различать три онтогенетических уровня ретрогенезов. При ретардации онтогенеза на эмбриональных стадиях происходит фетализация, приводящая к сохранению во взрослом состоянии эмбриональных признаков (как это было описано Л.Больком для человека). Еще один хорошо известный пример фетализации - задержка гистогенеза скелета на эмбриональных стадиях, что приводит к сохранению у взрослых позвоночных животных некоторых групп (круглоротые, хрящевые рыбы, в значительной степени современные хвостатые и бесхвостые амфибии) хряща как основной скелетной ткани. Задержка соматического онтогенеза у личинок (вплоть до полного подавления метаморфоза) приводит к ларвизации, или личиночному педоморфозу. При большей или меньшей степени ларвизации сформировалась организация таких групп хвостатых амфибий, как скрытожаберники (Cryptobranchidae), амфиумы (Amphiumidae), протеи (Proteidae) и сирены (Sirenidae). Наконец, морфогенез некоторых соматических органов может быть задержан уже на ранних постметаморфозных (или постнатальных) стадиях. В этом случае у взрослого организма сохраняются некоторые признаки ювенильных стадий онтогенеза - ювенилизация, проявления которой не столь ярки, как в двух других формах ретрогенезов, но зато наиболее обычны (например, некоторое облегчение скелета, редукция надглазничного валика, уменьшение размера зубов и другие изменения, происходившие при становлении организации неоантропов).
Основная эволюционная роль любого педоморфоза (и вообше гетерохронии) заключается в формировании новых комбинации фенотипических признаков, причем эти преобразования могут происходить без существенных генетических изменений, поскольку генетические механизмы возникновения гетерохронии сравнительно просты, основываясь на изменениях активности отдельных генов. Поэтому гетерохронии являются широко распространенной формой наследственной изменчивости онтоге-незов, предоставляющей обильный материал для действия естественного отбора.
Онтогенез и целостность организма
В предыдущих разделах мы рассматривали механизмы эволюционных преобразований онтогенеза. Однако необходимо принимать во внимание и противоположный аспект - обеспечение в онтогенезе целостности организма, с которой связана эволюционная устойчивость важнейших морфогенетических систем. Этот аспект онтогенеза также оказывает глубокое воздействие на характер эволюционных преобразований. Целостность организма обеспечивается разнообразными корреляциями, представляющими собой системы упорядоченных взаимодействий различных структур в онтогенезе. Коррелятивно связанными друг с другом могут быть вообще любые структурные и функциональные характеристики организма. Механизмы коррелятивных связей весьма различны. И.И.Шмальгаузен выделил три основных онтогенетических уровня корреляций.
1. Генетические корреляции основываются непосредственно на процессах, происходящих на уровне генома (биосинтез белков - например, при контроле нескольких оперонов общим геном-регулятором). В фенотипе генетические корреляции проявляются, например, в виде плеиотропного действия генов, т. е. влияния данного гена на различные признаки и свойства. Так, мутация "ко-роткоухость" у домовой мыши сопровождается укорочением нёба и уменьшением высоты морды; мутация "полидактилия" у кур приводит к одновременному удвоению первого пальца как на задних, так и на передних конечностях; мутация "белоглазие" у дрозофилы сказывается также на плодовитости мух (снижая ее) и продолжительности жизни (уменьшение) и т.п.
Некоторые генетические корреляции связаны с образованием внутри генотипа генных комплексов "супергенов", интегрированных различными способами. Один из таких способов - возникновение хромосомных мутаций типа инверсий, затрудняющих Кроссинговер в мейозе с хромосомой, имеющей "нормальную" Последовательность генов. Отдельные гены, входящие в состав Подобного супергена, могут контролировать развитие разных фе-Нотипических признаков. Отбор будет благоприятствовать сохранению таких супергенов, если объединяемые в них генные комплексы обеспечивают более высокую приспособляемость особей.
Наконец, как генетическую корреляцию можно рассматривать также эффект сцепления генов, локализованных в одной хромосоме поблизости друг от друга, хотя эта корреляция относительно легко нарушается кроссинговером и, вероятно, не имеет большого эволюционного значения.
2. Морфогенетические корреляции представляют собой результат взаимодействия различных зачатков в ходе эмбрионального развития. Примеры такого взаимодействия дают, в частности, явления эмбриональной индукции. Изменение индуктора влечет за собой соответствующие изменения индуцируемых структур. Если последние, в свою очередь, являются вторичными индукторами, цепь коррелятивных изменений продолжается далее (пример подобной цепи первичных и вторичных индукторов представляют соотношения глазного бокала, хрусталика и роговицы глаза).
3. Морфофункциональные, или эргонтические, корреляции являются результатом взаимодействий различных структур в морфо-функциональных системах организма в постэмбриональном периоде онтогенеза. Так, ослабление мышечной активности при вынужденной длительной неподвижности конечности приводит к уменьшению кровоснабжения соответствующих мускулов. Изменения в функционировании желез внутренней секреции сказываются на функциональном состоянии различных органов, что нередко сопровождается и структурными изменениями последних. Например, усиление секреции андрогенов корой надпочечников приводит к общей маскулинизации организма, т. е. проявлению и усилению мужских особенностей, даже у генетически женских индивидов: усиленный рост скелета, развитие мускулатуры, развитие некоторых мужских и редукция женских вторичных половых признаков. Все подобные взаимосвязи носят характер функциональных корреляций.
Выделение трех указанных категорий корреляций, в сущности, условно. Это ярко проявляется в рассмотренном выше плейо-тропном эффекте мутации гидроцефалии у мышей. Разнообразные патологические изменения головного мозга, черепа, глазных век, вибрисс и др. возникают на основе морфогенетических корреляций (взаимосвязи в ходе онтогенеза), сама же мутация непосредственно сказывается лишь на процессах формирования эмбрионального хрящевого скелета. Очевидно, в этом случае тесно связаны явления генетических и морфогенетических корреляций. В свою очередь, морфогенетические корреляции нередко переходят в корреляции функциональные. Так, недоразвитие у молодых животных некоторых скелетных мышц, основанное на генетических причинах или определяемое условиями развития, приводит к недоразвитию отростков и гребней на поверхности костей, служащих для прикрепления этих мышц (например, для височных мышц млекопитающих - венечные отростки нижней челюсти). Таким образом, три онтогенетических уровня корреляций тесно взаимосвязаны, представляя в целом общую систему интеграции организма в его индивидуальном развитии.
Изучение коррелятивных систем организмов имеет важное значение для понимания процессов эволюционных преобразований. Филогенетические изменения относительных размеров различных структур могут определяться постоянным соотношением скоростей их роста. Два признака могут изменяться в эволюции так, что направление и скорость изменений одного зависят от изменений другого. Например, по данным Р.Робба, в филогенезе лошадей сохранялось постоянное соотношение удлинения морды и всего черепа в целом, хотя абсолютные скорости этих процессов были различны. В результате изменение пропорций черепа в эволюции лошадей в сторону удлинения морды происходило, вероятно, не посредством специальных мутаций, ответственных именно за эти конкретные изменения относительной и абсолютной длины морды, а коррелятивно - благодаря общему увеличению размеров тела и головы в филогенезе лошадей при постоянном соотношении длины морды и длины черепа в целом. Таким образом, увеличение размеров тела коррелятивно сказывалось на соответствующем удлинении морды по отношению к общей длине черепа. Следовательно, в этом и других подобных случаях филогенетические изменения конкретного морфологического признака, которые констатируются палеонтологами, происходили без каких бы то ни было специально ответственных за эти изменения мутаций. По определению Симпсона, в подобных случаях признак эволюционирует не генетически, а лишь морфогенетически. Подобные ситуации необходимо учитывать и при определении темпов эволюционных изменений различных признаков.
Благодаря наличию корреляций отбор, благоприятствующий какому-либо признаку, будет автоматически способствовать сохранению и распространению в популяциях всего комплекса коррелятивно связанных признаков, которые сами по себе могут иметь как нулевую (нейтральные признаки), так и слабо отрицательную приспособительную ценность. Поэтому нельзя ограничиваться действием отбора на какой-либо один признак - эффект отбора всегда определяется по общей приспособительной ценности всех взаимосвязанных изменений. При искусственном отборе, который селекционеры ведут обычно по отдельным интересующим их признакам, в результате происходит общее снижение приспособленности организмов, вплоть до потери ими жизнеспособности вне специфических условий их содержания человеком.
Формирование систем корреляций, совершенствующих интеграцию организма, в филогенезе происходило параллельно с усложнением (дифференциацией) организации, являясь важной составной частью эволюционных преобразований по пути морфо-физиологического прогресса. Можно выделить два основных пути формирования коррелятивных связей между фенотипическими признаками. Первый из них -- возникновение корреляций как случайный побочный результат перестроек морфогенетических систем, обусловленных возникновением каких-либо мутаций. В этом случае корреляции признаков на уровне фенотипа первоначально не имеют приспособительного значения (как описанная Ч.Дарвином корреляция глухоты, голубой окраски глаз, белой шерсти и мужского пола у кошек).
Второй путь формирования корреляций имеет в отличие от первого приспособительный характер. Если какие-либо фенотипические признаки связаны друг с другом участием в общем адаптивном комплексе, особенно если они взаимно необходимы для успешного функционирования какого-либо приспособления, отбор будет благоприятствовать возникновению между ними отношений, имеющих характер эргонтических корреляций. Первоначально эти корреляции могут иметь модификационную природу, т.е. особенности строения и функций двух структур развиваются под взаимным влиянием или под воздействием связывающей их активности. Примером таких взаимоотношений может служить хорошо известная корреляция между состоянием мышечных волокон и снабжающих мышцу кровеносных капилляров в связи с уровнем мышечной активности. При ее усилении возникает так называемая "рабочая гипертрофия" мышцы: ее кровоснабжение возрастает путем расширения капилляров и усиления кровотока, а в мышечных волокнах увеличивается объем саркоплазмы (цитоплазмы мышечных волокон) и возрастает количество миофибрилл, что приводит к возрастанию силы сокращения мышцы.
Если стабилизирующий отбор благоприятствует закреплению таких взаимосвязей между разными структурами, появляются предпосылки для своего рода "заглубления" этих корреляций --их перехода на более ранние этапы онтогенеза. Иными словами, отбор благоприятствует закреплению мутаций, которые обусловили бы более глубокую связь данной комбинации признаков - на уровне морфогенеза и далее на уровне генотипа. Это может быть достигнуто посредством различных генетических механизмов, простейшим из которых является, вероятно, объединение генов, ответственных за развитие адаптивно взаимосвязанных признаков, в одну группу сцепления посредством соответствующих хромосомных перестроек - транслокаций и инверсий. Далее активность разных генов может быть подчинена контролю общего гена-регулятора. Так возникают генетические комплексы, обеспечивающие устойчивое совместное наследование адаптивного комплекса фенотипических признаков.
Как полагает Р.Ридл, этот механизм может лежать в основе формирования устойчивых генетически закрепленных систем, совокупность которых формирует общий тип организации, характерный для крупного таксона и остающийся практически неизменным у всех составляющих его видов. Чем древнее данная система, тем жестче ее генетическая обусловленность и тем она эволюционно консервативнее.
И.И.Шмальгаузен показал, что стабилизирующий отбор благоприятствует развитию канализованности процессов морфогенеза: для сохранения постоянного фенотипического облика важных адаптивных признаков их морфогенез оказывается под контролем генов-модификаторов, которые способствуют достижению одного и того же онтогенетического результата при разной генетической основе. Гены-модификаторы блокируют фенотипические проявления продолжающих накапливаться в генофонде вида новых мутаций. Поэтому (несколько парадоксальным образом) в эволюции аппарат наследственности преобразуется в значительно большей степени, чем основные фенотипические структуры. Это является одной из причин различий в темпах молекулярной и морфологической эволюции (см. с. 71). В таких случаях эволюционно наиболее устойчивой оказывается система онтогенетических коррелятивных связей.
Важность интегрирующей роли стабилизирующего отбора наглядно проявляется в ситуациях, когда какие-либо органы или системы организма, достигшие в ходе предшествовавшей эволюции высокой степени сложности и совершенства, теряют свое прежнее адаптивное значение в связи с изменениями внешних условий или образа жизни. В таких случаях неизменно происходит постепенная дезинтеграция данной системы - распадение коррелятивных связей между ее компонентами, приводящее к недоразвитию некоторых из них, а затем и всего органа в целом. Так произошла, например, редукция глаз у многих животных, освоивших постоянное обитание в пещерах (в том числе у протеев, населяющих карстовые пещеры Югославии, см. рис. 103) или роющий образ жизни (многие пресмыкающиеся - амфисбены, Amphisbaenia; слепозмейки, Typhlopidae, и др.). При переходе от активного подвижного образа жизни к неподвижному прикрепленному оказываются ненужными, подвергаясь дезинтеграции и редукции, многие сложные органы, связанные с ориентацией (органы чувств) и передвижением (мышцы и скелет локомоторного аппарата), упрощается и редуцируется общая интеграционная система организма - нервная. Такие изменения произошли, например, в эволюции асцидий (см. рис. 104) - группы примитивных хордовых - оболочников, перешедших к сидячему образу жизни. Аналогичное упрощение организации характерно также для некоторых эндопаразитов. Особенно замечательна в этом отношении организация усоногого рачка Sacculina, во взрослом состоянии паразитирующего на крабах. Тело саккулины, располагающееся под брюшком краба (рис. 105), является лишь вместилищем половых желез и лишено каких бы то ни было других сложных органов. От тела отходят тонкие ветвящиеся отростки, оплетающие внутренние органы хозяина-краба, с помощью этих отростков паразит питается. У саккулины нет ни одной из сложно дифференцированных систем органов, характерных для членистоногих животных (нервной, рецепторной, дыхательной, кровеносной, пищеварительной, выделительной, локомоторной).

Под контролем стабилизирующего отбора эволюционные преобразования онтогенеза протекают таким образом, чтобы сохранялась и поддерживалась целостность организма. Напомним, что эволюция не сводится к преобразованиям биологических макромолекул, равно как и к изменениям частоты встречаемости разных аллелей в генофондах популяций, но представляет собой совокупность преобразований всех уровней биологической организации. При этом, как мы увидим далее, организационный уровень
Сальтационная концепция рассматривает микро- и макроэволюцию как качественно различные, не связанные друг с другом процессы, происходящие на уровне видов; новые крупные таксоны возникают сразу - как новые виды, посредством особых механизмов видообразования, обусловливающих быстрые и значительные преобразования - сальтации - предковых форм. Напротив, с позиций редукционистской концепции макроэволюция не имеет никаких собственных механизмов и полностью сводится ("редуцируется") к микроэволюционным изменениям, являясь лишь их обобщенной картиной, рассматриваемой в широкой исторической перспективе. Наконец, по представлениям системной концепции, макроэволюционные преобразования интегрируются из микроэволюционных изменений, но не сводятся к простой сумме последних, поскольку макроэволюция характеризуется целым рядом общих закономерностей, не проявляющихся на уровне микроэволюции; интеграция макроэволюционных преобразований определяется действием специфических механизмов макроэволюции. Рассмотрим эти концепции подробнее.
Сальтационная концепция макроэволюции
В третьей части книги мы неоднократно подчеркивали прерывистость палеонтологической летописи, в которой новые крупные таксоны часто появляются впервые как бы уже сложившимися - их наиболее древние известные представители уже обладают основным комплексом признаков (общим планом строения) нового отряда, класса и типа. Достаточно вспомнить древнейших известных представителей бесхвостых амфибий (Triadobatrachus), черепах (Proganochelys), змей (Dinilysia) или появление в кембрии представителей большинства типов многоклеточных животных. Это побуждало многих палеонтологов (Э.Коп, О.Шиндевольф, А.Б.Ивановский и др.) рассматривать макроэволюцию как качественно особый процесс по отношению к микроэволюции -"происхождению видов". Эта точка зрения была поддержана также некоторыми генетиками и другими биологами (Ю.А.Филипченко, Р.Гольдшмидт, Л.И.Корочкин и др.), разделяющими позиции мутационизма (см. с. 44). Согласно этой концепции микроэволюционные процессы - преобразования генофондов популяций, основанные на малых мутациях и контролируемые естественным отбором, - приводят лишь к формированию новых видов, представляющих частные варианты прежнего плана строения.
Напротив, макроэволюционные процессы преобразуют сам общий план строения, причем эти существенные перестройки организации происходят не постепенно, складываясь из мелких изменений, а сразу - посредством одного или немногих крупных преобразований в ходе смены всего нескольких поколений. Такие скачкообразные преобразования, в результате которых возникают виды с комплексом признаков новых крупных таксонов, называют сальтациями или макрогенезом. Некоторые современные сальтационисты, например В.И.Назаров, разделяя взгляды мутационизма начала XX в., полагают, что видообразование всегда является сальтационным, а микроэволюционные процессы обусловливают лишь внутривидовую изменчивость.
Главная проблема, стоящая перед сторонниками сальтационизма, - поиски специфических факторов и механизмов макрогенеза. Для ее решения охотно привлекают самые разнообразные факторы, которые хотя бы в первом приближении кажутся подходящими: крупные ("системные") мутации, сразу приводящие к значительным изменениям фенотипа; "взрывы мутаций" в результате различных катастрофических факторов земного или внеземного происхождения; "горизонтальный перенос" генетической информации между разными филетическими линиями; перестройка ранних стадий онтогенеза, существенно изменяющая протекание его последующих стадий; разрушение биоценозов в результате катастроф общепланетарного значения и т.п. Некоторые из этих факторов уже были рассмотрены нами в предшествовавших разделах, где было показано, что ни системные мутации (с. 122), ни взрывы мутаций в результате общепланетарных катастроф (с. 164) не могут рассматриваться как определяющие факторы макроэволюции.
При этом наиболее важным аргументом против существенной роли системных мутаций в макроэволюции является не столько редкость и низкая жизнеспособность таких мутантов, сколько принципиальная невозможность объяснить с их помощью возникновение и гармоничные эволюционные преобразования сложных адаптивных морфофункциональных систем организма, компоненты которых имеют разное происхождение. Так, дыхательная система у высших наземных позвоночных (амниот) включает легкие и их дыхательные пути (развивающиеся в онтогенезе как выросты эн-тодермального пищеварительного тракта), скелетные и мышечные структуры грудной клетки (мезодермального происхождения); дыхательная система снабжается кровеносными сосудами, ее работа интегрируется нервными центрами головного и спинного мозга. Совершенно очевидно, что никакая макромутация заведомо не может обусловить возникновение такой комплексной гетерогенной системы и ее координированные перестройки при становлении новых типов организации в макрофилогенезе - уже просто потому, что такие комплексы развиваются как в эволюции, так и в онтогенезе из разных зачатков, контролируемых разными генетическими структурами и разными морфогенетически-ми системами. И реально в филогенезе наблюдаются независимые преобразования разных компонентов таких систем (принцип независимой эволюции различных признаков, по Г.Осборну). Любопытно, например, что в эволюции ядовитых ужей (Boiginae) даже увеличение размера ядовитых зубов и формирование на них желобков для проведения яда происходило независимо друг от друга.
В последние десятилетия среди сторонников сальтационизма получили широкое распространение представления о новом факторе макроэволюции - так называемом горизонтальном (или латеральном) переносе генетической информации между разными видами - посредством гибридизации или переноса плазмид (в частности, при вирусной трансдукции). Горизонтальный перенос как фактор макроэволюции привлекателен тем, что одновременно позволяет объяснить и возникновение сальтаций, и происхождение эволюционных параллелизмов. Однако среди факторов горизонтального переноса гибридизация, ведущая к возникновению аллополиплоидов, представляет собой как по своей сущности, так и по результатам микроэволюционный процесс: этим путем могут возникать новые виды, но нет никаких оснований видеть в нем особый способ возникновения крупных таксонов. Что же касается переноса генетической информации плазмидами, то даже микроэволюционная его роль до сих пор не установлена вполне достоверно. Лабораторные достижения в области генной инженерии не могут служить доказательством реальности подобных процессов в природных условиях. Хотя имеются некоторые сведения о естественном переносе генетической информации между разными видами прокариот, для эукариот убедительных данных такого рода пока нет. Однако даже если бы такой перенос оказался реальностью, речь может идти лишь о незначительных фрагментах генетического кода, обеспечивающих в клетках другого вида синтез какого-либо нового белка - и только. Перенос этим путем от одного вида к другому крупных генетических комплексов, обеспечивающих формирование сколько-нибудь сложных фенотипических структур, представляется совершенно невозможным, поскольку такие структуры образуются в результате сложных взаимодействий разных морфогенетических систем, контролируемых многими генами, зачастую имеющими разную локализацию в геноме, причем фенотипический эффект любого гена в значительной степени определяется его генетическим окружением.
В сущности, результаты "латерального переноса" генетической информации принципиально не отличаются от обычных мутаций - ни по масштабу возникающих изменений, ни по их скорости. Как и для мутаций, чем больше масштаб интродуцированно-го в клетку чужеродного генетического фрагмента, тем меньше шансов на выживание развивающихся из таких клеток организмов и оставления ими потомства.
По мнению некоторых ученых, можно избежать этих трудностей, если заменить макромутации или горизонтальный перенос наследственной информации в роли основных факторов макроге-неза на "преобразования онтогенеза", происходящие за столь же краткие эволюционные сроки, т. е. в пределах смены немногих или даже всего двух поколений. При этом полагают, что в основе такого изменения онтогенеза могут лежать мутации, которые существенно не нарушают интеграцию генома, но их эффект начинает проявляться уже на ранних стадиях онтогенеза, и вследствие этого они оказывают влияние на различные морфогенетические системы, приводя тем самым к значительным перестройкам фенотипа. Однако и в этом случае не удается преодолеть трудностей, стоящих перед теорией макрогенеза. Любое наследственное изменение онтогенеза может возникнуть лишь в результате соответствующей мутации. О масштабе мутации приходится судить по ее фенотипическим результатам независимо от того, произошло ли при этом значительное изменение самого генома или нет. Поэтому все сказанное выше о макромутациях остается в силе и для "изменений онтогенеза". Хотя данная мутация не нарушает интеграцию генома, при ее значительном фенотипическом эффекте очень велика вероятность гибельных нарушений интеграции тонко скоординированных морфогенетических систем развивающегося организма. Типичный пример подобного изменения онтогенеза представляет фенотипический эффект мутации врожденной гидроцефалии у домовой мыши, который мы рассматривав в предшествующей главе. В этом случае происходит именно ранняя перестройка онтогенеза, вызванная мутацией, которая не нарушает интеграцию генома, но благодаря взаимодействиям разных морфогенетических систем влечет за собой целый ряд фенотипических преобразований, которые в итоге приводят к гибели организма.
Начиная с Э.Копа (1887) для объяснения механизма сальта-ционных изменений в макрофилогенезе пытались привлекать онтогенетические гетерохронии, особенно педоморфозы. Вероятно, гетерохронии развиваются за относительно краткие (в масштабах макроэволюции) сроки, но чем больше систем организма подвергается гетерохрониям, тем больше регуляторных генов должны подвергнуться мутациям. В некоторых случаях эти эволюционные перестройки онтогенеза действительно сыграли важную роль в формировании организации крупных таксонов (аппендикуля-рии среди низших хордовых, голосеменные и покрытосеменные растения; см. предыдущую главу). Однако педоморфозы и другие гетерохронии не могут претендовать на роль универсального механизма макроэволюции, поскольку, в сущности, они представляют собой не способ возникновения качественных новообразований, а некоторую перекомбинацию уже существовавших особенностей организации разных стадий онтогенеза. Педоморфоз позволяет избавиться от специализированного состояния признаков взрослых стадий, выводя на онтогенетический уровень взрослого фенотипа ювенильные, личиночные, зародышевые признаки - обычно более пластичные, чем соответствующие особенности взрослой стадии.
К сальтационной концепции примыкают взгляды сторонников неокатастрофизма, которые видят причину радикальных изменений биосферы Земли в глобальных катастрофах, приводивших к гибели значительную часть существовавших прежде видов организмов и стимулировавших "взрывную" эволюцию других видов. Причины подобных катастроф указывались самые разные: усиление вулканизма, возрастание фона космической радиации из-за относительно близкой к Солнечной системе вспышки cверхновой звезды, столкновение Земли с астероидом или кометой и т.п. В более новых вариантах гипотез предполагают не прямое, а косвенное влияние космических факторов на биосферу: кризисные условия приводят к распаду экосистем, что, по мнению В.А.Красилова, приводит к снижению конкуренции и повышает шансы на выживание макромутантов - "многообещающих уродов", по Р.Гольдшмидту.
Различные "катастрофические гипотезы" были рассмотрены нами в третьей части книги. Анализ причин предполагавшихся катастроф обычно обнаруживал сомнительность или невозможность глобального воздействия соответствующих факторов. Кроме того, с позиций неокатастрофизма невозможно объяснить, почему при предполагаемых глобальных катастрофах одни виды подвергались быстрым и значительным преобразованиям, другие вымирали, а третьи, жившие в тех же местообитаниях и подвергавшиеся воздействию тех же самых факторов, оставались неизменными. Напомним, например, что рубеж мезозойской и кайнозойской эр преодолели, словно бы "не заметив" предполагаемой глобальной катастрофы, такие группы, как костистые рыбы, черепахи, ящерицы, змеи, отряды птиц и млекопитающих и т.д.
Распад экосистем в кризисных условиях не может приводить к снижению конкуренции и повышать шансы на выживание макромутантов, поскольку значительное ухудшение условий существования неизбежно должно привести к противоположному результату - резкому обострению борьбы за существование и, в частности, к обострению конкуренции из-за нарастания дефицита необходимых ресурсов. Всякого рода кризисные ситуации приводят к возникновению катастрофического отбора, благоприятствующего выживанию и размножению особей данного вида, наиболее устойчивых к экстремальным условиям (развитие гипер-адаптаций, см. с. 108 и 140). Катастрофический отбор должен выбраковывать макромутантов, у которых нарушения интеграции систем организма неизбежно снижают жизнеспособность, особенно эффективно.
Ко всему прочему, макроэволюционные преобразования, приводящие к формированию основного комплекса признаков новых крупных таксонов, не проявляют обязательной приуроченности к эпохам биосферных кризисов. Напомним,что в середине мезозоя, когда на Земле отнюдь не было катастрофических изменений, сформировалась организация таких крупных таксонов, как тип Диатомовые водоросли, класс Покрытосеменные растения, класс Птицы и подклассы Сумчатые и Плацентарные млекопитающие. Очевидно, возникновение новых крупных таксонов может происходить при относительно стабильных условиях, исторически предшествуя радикальным перестройкам экосистем.
В заключение необходимо подчеркнуть, что представления о сальтациях (какими бы причинами эти сальтации не объясняли) вступают в очевидное противоречие с реально наблюдаемыми (по палеонтологическим данным) темпами макрофилогенеза. В предыдущей части книги мы видели, что для всех групп животных и растений, ранние этапы эволюции которых с достаточной полно той представлены в палеонтологической летописи, быстрота формирования нового типа организации относительна - обычно этот процесс занимает несколько десятков миллионов лет. Очевидно, здесь не может идти и речи о сальтации, происходящей в пределах смены немногих поколений. Сальтационизм, в сущности, основан на ошибочном (крайне гипертрофированном) представлении о темпах типогенеза.
Таким образом, по общей совокупности современных данных сальтационизм представляется лишенным серьезных оснований.
Редукционистская концепция
Сальтационизму противостоят две другие концепции макроэволюции - редукционистская и системная, которые утверждают единство всего эволюционного процесса: филогенетические преобразования любого масштаба происходят на основе микроэволюционных изменений. Микроэволюционные процессы идут непрерывно на любом отрезке филетических линий, и никаких иных изменений организмов, кроме микроэволюционных, не происходит. Эти общие представления совпадают у редукционистской и системной концепций.
Согласно собственно редукционистсткой концепции, нет никаких особых факторов и механизмов макроэволюции, отличающихся от микроэволюционных. Макроэволюция здесь полностью сводится (редуцируется) к микроэволюционным изменениям, являясь лишь их обобщенной картиной, рассматриваемой в широкой исторической перспективе (Д.Симпсон, Б.Ренш, Э.Майр и др.). Эта концепция имеет в основном генетико-экологический характер, а в методологическом отношении является приложением к анализу эволюционного процесса принципа редукционизма, утверждающего полную сводимость (редукцию) свойств любой системы к свойствам ее элементов и следующую отсюда возможность обратного выведения свойств системы из свойств ее элементов. Основываясь на этом принципе, полагают, что все явления, процессы и закономерности макрофилогенеза полностью сводятся к микроэволюционным процессам, т. е. обусловлены действием элементарных эволюционных механизмов в пределах видовых популяций. Действительно, как мы видели во второй части книги, совокупность основных эволюционных факторов - мутационного процесса и естественного отбора в изменяющихся условиях внешней среды - необходима и достаточна для приспособительной эволюции организмов; влияние дополнительных эволюционных факторов (конкуренция, гибридизация, изоляция популяций, колебания их численности, дрейф генов и др.) обусловливает все разнообразие микроэволюционных процессов, ведущих к видообразованию. Некоторые закономерности и правила, обнаруживающиеся в макрофилогенезе, также находят удовлетворительное объяснение в кумулятивном (накапливающем) действии естественного отбора, интегрирующем результаты микроэволюционных изменений.
Так, упоминавшийся выше принцип независимой эволюции отдельных признаков обусловлен дискретностью наследственной информации и мутационного процесса: эволюционные изменения разных признаков могут происходить независимо друг от друга в той мере, в которой они кодируются разными комплексами генов и контролируются разными морфогенетическими системами, не связанными корреляциями. Следствием принципа независимой эволюции отдельных признаков является мозаичная эволюция -соединение в организации промежуточных форм, как в мозаике, примитивного состояния одних признаков и продвинутого - других. Классическим примером мозаичной формы является археоптерикс. При параллельной эволюции нескольких родственных филетических линий нередко в разных линиях эволюционно обгоняют друг друга разные признаки, т.е. у различных видов возникают разные мозаичные комбинации признаков - явление, названное О.Абелем "перекрест специализаций". Типичным примером перекреста специализаций была эволюция параллельных филетических линий лошадиных непарнокопытных. С позиций редукционистской концепции легко находит объяснение и адаптивная радиация. Эта закономерность макрофилогенеза определяется действием на популяции дизруптивного отбора, благоприятствующего разным направлениям изменчивости и приводящего к распадению первоначального генофонда на несколько дочерних. Адаптивной радиации способствуют также такие дополнительные микроэволюционные факторы, как конкуренция, изоляция популяций, колебания их численности и дрейф генов.
В механизмах наследственности и морфогенеза коренится также сущность закона необратимости эволюции. Появление нового или исчезновение старого наследственного признака прежде всего связано с возникновением мутации определенного гена, а для подавляющего большинства сколько-нибудь сложных структур и функций - с перестройкой целых генетических систем, их контролирующих. При закреплении нескольких последовательных изменений, не связанных общим геном-регулятором и не входящих в одну морфогенетическую систему, вероятность возникновения совпадающих обратных изменений всех затронутых элементов, относящихся к разным генетическим и морфогенетическим системам, очевидно, исчезающе мала. Такой возврат для организма в целом практически невозможен, как это и констатирует упоминавшийся уже нами закон Л.Долло.
Следует, однако, отметить, что формулировка Долло излишне абсолютизирована, не допуская никакой реверсии не только для целых организмов, но и для отдельных их признаков. В действительности по отношению к отдельным признакам организма реверсия к предковому состоянию, утраченному в предшествующей эволюции, в некоторых случаях возможна. Во-первых, могут происходить обратные мутации отдельных генов. Разумеется, при этом возврат фенотипического признака к утраченному предковому состоянию возможен обычно лишь на уровне микроэволюции - если в генотипе еще не закрепились другие мутации, которые могут сказаться на фенотипическом проявлении данного гена. Если же такие мутации закрепились, то при возврате гена к первоначальному состоянию его фенотипический эффект в новой генетической среде будет совсем иным, чем у предков данного организма. Во-вторых, возврат отдельных признаков к предковому состоянию может быть достигнут при так называемой реализации латентных (скрытых) потенций генома. В этом случае гены (или полигенные системы), контролировавшие какие-то признаки у предков, сохраняются и у потомков, но их действие блокируется влиянием генов-модификаторов или других морфогенетических систем. В результате предковый признак в фенотипе не проявляется, хотя в геноме сохраняются структурные гены, ответственные за его развитие. Следовательно, геном сохраняет латентную потенцию к формированию исчезнувшего фенотипического признака, характерного для предков данного организма. В таком латентном состоянии признак может сохраняться в генофондах последовательных поколений, вероятно, очень долго. Спустя ряд поколений действие факторов, блокирующих морфогенез латентного признака, может быть снято. На уровне генома это происходит благодаря репрессии соответствующего гена-модификатора, а в ходе морфогенеза - в результате взаимодействия разных морфогенетических систем; последнее может произойти даже под влиянием внешних факторов, воздействующих на протекание морфогенетических процессов. Тогда давно исчезнувший признак предков вновь проявится в фенотипе потомка. Вероятно, таков механизм развития некоторых атавизмов (появление у отдельных особей данного вида каких-либо древних, давно утраченных предковых особенностей, например трехпалость у лошадей, развитие хвоста, сплошного волосяного покрова или добавочных млечных желез у человека и т.п.).
Эволюционная реверсия по отдельным признакам возможна также благодаря педоморфозам. Для отдельных органов, эволюционные преобразования которых осуществлялись путем анаболии, в эмбриональном состоянии сохраняется некоторое сходство с состоянием соответствующих структур у предков (рекапитуляция). Педоморфоз "выводит" эмбриональные признаки на уровень взрослого состояния. Тогда эмбриональные рекапитуляции также становятся признаками взрослого организма, и состояние соответствующих органов оказывается близким к таковому у далеких предков. Так, у аппендикулярий располагающиеся в хвосте хорда, спинной нервный ствол и мышцы (см. рис. 104) напоминают соответствующие признаки предков оболочников, которые до их перехода к прикрепленному образу жизни на дне моря были, по-видимому, в значительной степени сходны с современными бесчерепными - ланцетниками.
Эволюционные параллелизмы отчасти объясняются конвергенцией - приобретением сходных адаптации при обитании в сходных условиях внешней среды (см. с. 230). Однако далеко не все параллельные изменения в близких филетических линиях поддаются этому простому объяснению, поскольку некоторые сходные морфофункциональные изменения у разных групп развиваются в связи с разными приспособлениями (это явление было названо параконвергенцией) или же сами по себе являются адаптивно
нейтральными.
Среди факторов микроэволюции направляющим является лишь естественный отбор. Отбор, по выражению Д.Симпсона, "оппортунистичен" - он направляет эволюцию по любому пути, доступному для данной группы организмов и обеспечивающему ее биологический успех посредством развития и совершенствования соответствующих приспособлений. Следовательно, проявления направленности в макрофилогенезе не могут быть полностью объяснены действием известных факторов микроэволюции.
Точно так же и темпы макроэволюции не обнаруживают корреляции с темпами микроэволюционных изменений. Напомним, что в периоды типостаза происходило интенсивное видообразование при отсутствии существенных макроэволюционных преобразований, и при самых высоких темпах видообразования происходящие изменения обычно не затрагивают общий план строения. Таким образом, ускорение микроэволюционных изменений отнюдь не обязательно приводит к возрастанию скорости макроэволюции. Даже такие радикальные преобразования генофондов, как обоснованная Э.Майром "генетическая революция" (см. с. 108), могут привести лишь к быстрому обособлению нового вида, но не к макроэволюционным преобразованиям.
Д.Симпсон назвал феномен ускорения темпов макроэволюции при становлении организации нового крупного таксона "квантовой эволюцией" и предложил для его объяснения гипотезу смены адаптивных зон. Под адаптивной зоной понимается либо определенный тип среды обитания с соответствующим набором жизненных ресурсов (например, морское дно, приливно-отливная зона, толща воды в открытом океане и т.п.), либо определенный способ использования конкретных жизненных ресурсов (например, "адаптивные зоны собачьих и кошачьих хищных млекопитающих"). Причиной квантовой эволюции Симпсон считал дискретность адаптивных зон: при переходе из одной зоны в другую, т. е. при освоении новой среды обитания, нового образа жизни, нарушается "равновесие между организмом и средой" и возникает "неадаптивная фаза" эволюции, которая не может продолжаться долго: оказавшаяся в "неадаптивной фазе" группа организмов либо вымирает, либо под усиленным давлением отбора ускоренно переходит в новую адаптивную зону. При этом происходят крупные изменения организации, необходимые для формирования нового адаптивного типа.
Однако новый крупный таксон может сформироваться и в прежней адаптивной зоне, которую занимали его предки. Например, среда обитания и используемые ресурсы у ящериц очень близки или практически те же самые, что и у их предков - эозухий. С другой стороны, если происхождение нового крупного таксона действительно связано с освоением новой адаптивной зоны, часто старая и новая адаптивные зоны не разграничены резко, между ними имеется промежуточная адаптивная зона, где длительное время существуют приспособленные к ее условиям виды предкового и дочернего крупных таксонов. Удачным примером таких соотношений является происхождение змей, возникших от одной из групп ящериц. Адаптивные зоны современных ящериц и змей в целом четко различаются: ящерицы питаются главным образом мелкими пищевыми объектами, преимущественно беспозвоночными; змеи - крупной добычей и в основном - позвоночными; различны и способы овладения добычей у ящериц и змей. По существующим представлениям, обособление змей произошло при освоении их предками скрытного и отчасти роющего образа жизни. Позднее основной филогенетический ствол змей вновь вернулся к жизни на поверхности почвы, что было связано с приобретением нового способа питания (нападение на относительно крупную добычу, проглатываемую целиком с использованием максимально развитого кинетизма черепа). Однако в промежуточных адаптивных зонах и ныне обитают многочисленные группы роющих ящериц, утративших конечности (рис. 106), и роющих змей. Из этого следует, что постулированного Симпсоном усиленного давления отбора, побуждавшего предков змей ускоренными темпами покинуть промежуточные адаптивные зоны, очевидно, не было. При этом особенно интересно, что современные виды роющих ящериц и змей, занимающие промежуточные адаптивные зоны, сохраняют основные черты организации своего крупного таксона: промежуточные адаптивные зоны заняты, но промежуточных современных форм между этими двумя группами в них нет! Очевидно, смена адаптивных зон не может служить объяснением квантовой эволюции.

Сторонники концепции прерывистого равновесия ("пунктуализм", см. с. 110-112) считают особым фактором макроэволюции так называемый видовой отбор, под которым понимается отбор по признакам, единообразным в пределах данного вида, но различающимся у близких конкурирующих видов. При этом виды как бы уподобляются отдельным особям, время их существования - жизни особи, а дифференцированное вымирание (обусловленное видовым отбором) - дифференцированной гибели особей. Это сравнение само указывает на слабые стороны данной концепции: отбор как вероятностный фактор эффективен лишь при достаточно большой выборке; при малой выборке (меньше 50 единиц при коэффициенте отбора 0,01; см. с.91) на первый план выходит влияние случайных факторов. Очевидно, число близких видов, различающихся единообразными видовыми признаками друг от друга, в этих масштабах невелико и совершенно недостаточно для эффективного действия видового отбора. Отбор по устойчивым видовым признакам, имеющим приспособительное значение, при конкуренции близких видов, конечно, происходит, но, в сущности, он представляет собой не особый "видовой", а обычный отбор, оперирующий многочисленными особями каждого вида - носителями соответствующих видовых признаков. Этот отбор, как и любая форма естественного отбора, благоприятствует любому адаптивному направлению изменчивости и поэтому сам по себе не может быть источником длительной направленности эволюции. К тому же если бы видовой отбор как таковой был реален, он приводил бы лишь к крайне медленным эволюционным преобразованиям, поскольку материалом для него являются наиболее устойчивые видовые признаки.
В последние годы получили распространение взгляды о биоиенотической регуляции как специфическом факторе макроэволюционных преобразований. В условиях стабильных биоценозов механизмы ценотического гомеостаза (см. с.117) являются источником сил стабилизирующего отбора. В стабильных экосистемах продолжаются микроэволюционные процессы. Условия для макроэволюционных преобразований создаются при перестройках биоценозов, при нарушении систем их гомеостаза. Вероятно, биоценотические взаимодействия действительно могут играть важную роль в регуляции микроэволюционных процессов. Однако повышение интенсивности видообразования в период становления новых биоценозов отнюдь не всегда приводит к формированию новых крупных таксонов. С другой стороны, как уже отмечалось, макроэволюционные преобразования могут происходить, вероятно, и в относительно устойчивых экосистемах с использованием незанятых экологических ниш или "свободных лицензий". Перестройка ценозов может быть не причиной, а следствием формирования новой группы организмов, как это было, например, при возникновении и распространении покрытосеменных растений.
Подводя некоторые итоги, мы можем заключить, что в рамках редукционистской концепции получают удовлетворительное объяснение такие характеристики макроэволюции, как независимая эволюция признаков, мозаичная эволюция, адаптивная радиация, необратимость эволюции целостных организмов и возможность частичной реверсии отдельных признаков. Однако редукционистская концепция недостаточна для решения основных проблем макроэволюции - ее пульсирующих темпов, направленного характера и морфофизиологического прогресса. Это не означает, что макроэволюция является особым процессом, происходящим независимо от микроэволюции, как это утверждают сторонники сальтационизма. В каждый исторический момент в любом филогенетическом стволе происходят только микроэволюционные изменения, весьма разнообразные, но сами по себе ведущие лишь к более или менее быстрому обособлению новых филетических линий, т.е. более или менее быстрому видообразованию. Однако организация макроэволюции не сводится к совокупности микроэволюционных изменений - в том же смысле, в каком не сводятся свойства любой целостной системы, имеющей определенную организацию, к свойствам ее отдельных элементов. На этом строится третья - системная концепция макроэволюции.
Системная концепция макроэволюции
В основе этой концепции лежит методологический подход, обоснованный В.А.Энгельгардтом и получивший название интегратизм. Интегратизм использует результаты редукционистского подхода, являясь в определенном смысле его дальнейшим развитием и представляя собой анализ механизмов, обеспечивающих интеграцию структурных элементов в целостную систему. Всякая целостная система является не просто суммой каких-то структур" ных элементов, но результатом их интеграции, т. е. такого объединения, при котором свойства системы, основанные на особенностях ее организации, качественно отличаются от свойств элементов. При интеграции некоторые свойства элементов утрачиваются или преобразуются, формируя системные связи и новые системные свойства. При этом из одних к тех же структурных элементов могут сформироваться качественно разные системы, специфика которых определяется различиями их организации (например, многочисленные изомеры органических соединений).
Согласно системной концепции, микроэволюционные изменения являются структурными элементами эволюционного процесса, а макроэволюция -- системно организованный процесс, организация которого выражается в его историчности и определенных закономерностях, отсутствующих в микроэволюционных изменениях. В соответствии со сказанным макроэволюция представляет собой не просто сумму микроэволюционных изменений, но результат их интеграции.
Определенная интеграция микроэволюционных изменений осуществляется естественным отбором, который формирует на их основе приспособления организмов к условиям изменяющейся внешней среды. Но, как мы уже видели, отбор не может сам по себе обусловить такие проявления организации макроэволюции, как ее пульсирующие темпы и направленный характер. Эволюция, как мы уже подчеркивали, представляет собой преобразования всех уровней организации живых систем - от молекул биоорганических соединений до биосферы в целом. В этой сложной иерархии разных уровне и биологической организации особое место занимают целостные организмы, обладающие наиболее жесткими внутренними связями среди обособленных биологических систем (организмы - популяции - биологические виды - экосистемы - биосфера).
Важнейшую роль организационной основы живых существ для определения направлений эволюционных преобразований подчеркивал еще Ч.Дарвин, указавший, что "при определении характера вариации, орган изация или строение существа, испытывающего влияние, обыкновенно бывает гораздо более важным элементом, чем характер изменяющихся условий. Доказательством служит появление почти сходных изменений при различных условиях и появление различных изменений при условиях, по-видимому, одинаковых. Мы имеем еще лучшее доказательство в том, что близко параллельные разновидности часто получаются от самостоятельных рас или даже от самостоятельных видов, а также в частом повторении одного и того же уродства у одного и того же вида. К сожалению, эта мысль Ч.Дарвина (которую сам Дарвин считал, вероятно, важной, поскольку возвращался к ней в своих разных книгах) редко привлекала внимание ученых, что может быть связано с длительным доминированием в эволюционистике редукционистского генетико-экологического подхода, в котором при концентрации внимания на изменениях генофондов популяций природа организма оттесняется на второй план.
Понимание эволюционной роли организационной основы целостного организма связано с проникновением в биологию системного подхода на основе работ Л.Берталанфи и И.И.Шмальгаузен в 30-е гг. XX в.
Разнообразные биологические системы относятся к двум основным категориям, различающимся общим характером своей дифференциации и интеграции, который определяется, с одной стороны, степенью разнокачественности системных элементов, с другой же - степенью жесткости связей между ними. Корреляционные, или жесткие, системы состоят из разнокачественных элементов, объединяемых жесткими связями - корреляциями. В таких системах изменение одного элемента сопровождается соответствующими изменениями других. К этой категории принадлежат целостные организмы и большинство внутриорганизменных систем, образующих иерархию, в которой системы низших рангов являются элементами систем более высокого ранга (клетки - органы - морфо-функииональные адаптивные комплексы и системы органов). В целостном организме формируются специальные интегрирующие системы: у многоклеточных животных - нервная, сосудистая и эндокринная, причем нервная система является общим интегрирующим центром; в клетке аналогичную роль играет ядро. Стохастические, или дискретные, или корпускулярные, системы состоят из многочисленных взаимозаменяемых элементов, связи между которыми лабильны, т. е. неустойчивы, не жестки. В таких системах нет общего интегрирующего центра; утрата или изменение отдельных элементов не влечет за собой обязательных коррелятивных изменений других элементов. При отсутствии жестких внутрисистемных связей и большом количестве элементов в стохастических системах причинные взаимосвязи выявляются в виде стохастических (вероятностных) процессов. К этому типу относится большинство надорганизменных биологических систем - популяции, биологические виды, экосистемы и т.п. Только в стохастических системах может действовать естественный отбор, который, как известно, сам является вероятностным фактором.
Нужно отметить, что между жесткими и стохастическими системами нет резких границ. Во внутриорганизменных корреляционных системах кроме характерных для них жестких связей между некоторыми элементами могут существовать лабильные связи, наряду с разнокачественными присутствуют и множественные взаимозаменяемые элементы (например, многочисленные клетки разных типов, имеющиеся в любом органе многоклеточного организма). С другой стороны, и в некоторых надорганизменных дискретных системах возникает некоторая дифференциация элементов, например, в биоценозах разные виды занимают различные экологические ниши и разное положение в трофических цепях. Соответственно связи между ними приобретают некоторые черты корреляционных: изменения одного элемента сказываются на других. В целом степень жесткости связей между элементами системы пропорциональна степени дифференциации и соответствующему возрастанию сложности и упорядоченности системной организации. По этим показателям организменные системы значительно превосходят надорганизменные. Особенно отчетливо это различие проявляется при сравнении целостного организма и популяции. Микроэволюция представляет собой итог стохастических процессов в популяциях, и ее специфика определяется свойствами последних. Напротив, возникновение проявлений организации макроэволюции связано с жесткими биологическими системами и в первую очередь с целостным организмом.
В последующих главах мы рассмотрим, как решаются основные проблемы макроэволюции с позиций системной концепции.
Главные направления эволюционного процесса А.Н.Северцов выделил три главных направления эволюционных преобразований: 1) морфофизиологический прогресс - повышение общего уровня организации, ее усложнение; 2) морфо-физиологический регресс (противоположное направление) - понижение и упрощение общего уровня организации; 3) идиоадаптация - развитие частных приспособлений, не изменяющих общий уровень организации. Позднее для обозначения этих направлений эволюции разными авторами были предложены другие наименования. Некоторые из них укоренились благодаря большей краткости и унификации звучания. Эволюционные преобразования организмов, ведущие к морфофизиологическому прогрессу и имеющие важное общее значение для организма в целом, А.Н.Северцов назвал ароморфозами. По аналогии с этим И.И.Шмальгаузен предложил называть регрессивные изменения катаморфозами, а термин "идиоадаптации" заменить на алломорфозы. В соответствии с этими терминами были предложены (А.Л.Тахтаджяном, А.А.Парамоновым и Б. Реншем) новые названия трех главных направлений эволюционного процесса: морфофизиологический прогресс стали именовать орогенезом (или анагенезом), морфофизиологический регресс - катагенезом, развитие частных приспособлений - аллогенезом (или кладогенезом).
(Еще одним особым направлением эволюции А.Н.Северцов первоначально считал ценогенезы (зародышевые приспособления), но позднее он сам признал это неоправданным, так как различные ценогенезы соответствуют тому или другому из других главных направлений эволюции).
Примеры указанных главных направлений эволюции можно найти в филогенезе основных групп хордовых животных. В целом по пути морфофизиологического прогресса шла эволюция подтипа позвоночных. Оболочники - группа, несомненно процветающая и в наше время, претерпели морфофизиологический регресс в связи с переходом их предков к прикрепленному образу жизни. Развитие идиоадаптации - всевозможных частных приспособлений - характеризует эволюцию любого крупного таксона (например, разнообразнейшие формы клюва у птиц, разные варианты строения конечностей у млекопитающих и т.п.). Многие группы длительное время эволюционировали именно путем идиоадаптаций, не изменяя общего уровня организации (например, любой из отрядов современных земноводных или пресмыкающихся, такие отряды млекопитающих, как грызуны, насекомоядные и т.п.). Понятие идиоадаптации (или алломорфозов) в концепции А.Н.Северцова объединяет очень широкий круг эволюционных изменений организмов - от самых незначительных частных приспособлений к специфическим условиям существования и образу жизни отдельных видов до адаптации общего значения, создающих предпосылки для значительного расширения среды обитания или освоения качественно новой адаптивной зоны (обширного круга местообитаний, нового способа использования ресурсов среды). Примерами таких "частных адаптации общего значения" являются кожное дыхание у земноводных, раковина моллюсков, особенности осевого скелета и мускулатуры тела змей, связанные с особым способом передвижения этих пресмыкающихся. Удачным примером подобного приспособления является усиленное развитие кутикулы в покровах нематод. Гибкая, прочная и химически устойчивая кутикула позволяет этим червям освоить широкий круг разнообразных местообитаний, включающих толщу органических осадков на дне морей и пресных водоемов, почву, эндопаразитизм в растениях и животных. Сильное развитие кутикулы стало отличительной чертой класса нематод и оказало существенное влияние на самые различные особенности их организации: оно обусловило редукцию мерцательного эпителия, утрату кольцевых мышц в кожно-мышечном мешке (поскольку прочная кутикула затрудняет перистальтические червеобразные движения); соответственно изменился характер передвижения нематод, вместо перистальтики возникли боковые изгибания тела; редукция же мерцательного эпителия сделала необходимым развитие нового типа органов выделения - нефридии заменились на шейные железы'.
Очевидно, эволюционное приобретение таких адаптации общего значения не соответствует ни собственно идиоадаптациям - частным приспособлениям, не оказывающим широкого влияния на организм в целом, ни ароморфозам, ни катаморфозам, поскольку общий уровень организации при этом существенно не меняется.
Такие эволюционные преобразования, которые значительно увеличивают адаптивные возможности данной группы (позволяя расширить старую или занять новую адаптивную зону), оказывают существенное влияние на организм в целом, но не изменяют общий уровень организации, были названы эпектоморфозами. Обеспечивая возможность существования в обширной адаптивной зоне, эпектоморфозы сохраняются длительное время в ходе последующей адаптивной радиации, становясь признаками крупных таксонов. Соответствующее направление эволюционных изменений было названо эпектогенезом. Соотношения между разными направлениями эволюционного процесса можно проиллюстрировать схемой (рис. 107), в основе которой лежит аналогичная схема А.Н.Северцова, модифицированная в связи с введением эпектогенеза.

Необходимо отметить, что все указанные формы эволюционных преобразований организмов (аро-, эпекто-, алло- и катаморфозы) определяются как таковые лишь в масштабе макрофилогенеза, когда приспособительный характер эволюционных изменений и их значение для организма как целого становятся вполне очевидными. На уровне же микроэволюции происходят как приспособительные (направляемые отбором), так и неприспособительные (нейтральные) изменения генофондов популяций; их дальнейшая эволюционная судьба и значение для организма, как правило, еще недостаточно ясны. Поэтому целесообразнее вообще не употреблять указанные термины для обозначения микроэволюционных изменений и использовать их лишь на макроэволюционном уровне.
Направляющие факторы эволюции
С точки зрения сторонников "оппортунистической эволюции", за различными направлениями эволюционных изменений не стоит никаких закономерных и организующих тенденций, кроме действия естественного отбора, который организует изменчивость организмов лишь в направлении развития приспособлений к изменениям среды обитания. С этих позиций главные направления эволюционного процесса (аро-, эпекто-, алло- и катагенез), по сути дела, равнозначны - в том смысле, что каждое из них является лишь средством для достижения успеха данной группы организмов в борьбе за существование (такой точки зрения придерживался и А.Н.Северцов).
Действительно, среди движущих факторов эволюции лишь естественный отбор обладает организующим воздействием на изменчивость организмов, и при этом отбор действительно лишен определенной направленности, что подчеркивал еще Ч.Дарвин. Но Дарвин указал и на фактор, определяющий конкретные направления эволюционных преобразований: "Природа условий имеет в определении каждого данного изменения подчиненное значение по сравнению с природой самого организма". Хотя эволюция организмов основывается на вероятностных процессах - возникновении мутаций (фенотипическое проявление которых неадекватно вызвавшим их появление изменениям внешних условий) и естественном отборе, "природа организма", т. е. организационная основа живых систем, ограничивает проявления случайности в эволюции определенными рамками. Иначе говоря, системная организация канализует филогенез, т.е. направляет эволюционные преобразования в определенные русла, и для любой конкретной группы организмов выбор возможных путей эволюции ограничен. Концепция жестко запрограммированной (номогенетической) эволюции основана на абсолютизации направляющей роли в эволюционном процессе организационной основы живых систем, тогда как концепция оппортунистической эволюции - на абсолютизации направляющей роли естественного отбора. Обычно истину следует искать где-то между крайними точками зрения.
Конкретные направления филогенетических преобразований различных групп организмов определяются взаимодействием сил естественного отбора и исторически сложившейся организации этих групп. Поэтому можно говорить о двух категориях направляющих факторов эволюции: внеорганизменных (силы отбора) и организменных.
Для любого данного вида особенности его организации создают предпосылки (преадаптации) к развитию определенных приспособлений и препятствуют развитию других, "разрешают" некоторые направления эволюционных преобразований и "запрещают" другие направления. Совокупность этих позитивных и негативных характеристик эволюционных возможностей данной группы и обозначена как организменные направляющие факторы эволюции. Эти факторы можно разделить (несколько условно) на три категории, в соответствии с уровнем их проявления в онтогенезе: 1) генетические, 2) морфогенетические, 3) морфофизио-логические (морфофункциональные).
Действие двух первых категорий организменных направляющих факторов в полной мере проявляется уже на уровне микроэволюции. Как уже отмечалось (ч. II, гл. 1), каждый генотип и генофонд каждого вида характеризуется определенным набором возможных ("разрешенных") мутаций, или спектром мутационной изменчивости, который ограничен не только качественно, но и количественно, т. е. определенной частотой возникновения каждого типа мутаций. При этом некоторые мутации оказываются для данного генотипа (и генофонда) невозможными (запрещенными) - например, синие и зеленые окраски глаз для мух-дрозофил или синие окраски цветов для розоцветных растений. Причиной этого является отсутствие в генотипе соответствующих биохимических предпосылок.
Так как генофонды родственных видов сохраняют наборы гомологичных генов, унаследованных от общего предка, в них закономерно появляются гомологичные мутации (см. с. 65). Гомологичные мутации могут послужить основой для параллельных эволюционных изменений у близкородственных видов, относительно недавно дивергировавших от общего предка. Однако с течением времени в генофондах обособленных видов неизбежно накапливаются разнокачественные (негомологичные) мутации; это происходит даже при действии стабилизирующего отбора, когда фенотипический эффект мутаций структурных генов блокируется генами-модификаторами. У разных видов, генофонды которых были изолированы друг от друга достаточно длительное время, сохраняются гомологичные структуры фенотипа, но их генетический контроль может значительно (и даже практически полностью) различаться. Поэтому параллельная эволюция филетических линий, давно дивергировавших от общего предка (до уровня разных родов, семейств и т.д.), основывается не столько на гомологичных мутациях, сколько на действии двух других категорий организменных направляющих факторов.
(Гомология - сходство структур, основанное на общности их происхождения. Соотношение гомологичных структур, принадлежащих к разным уровням иерархической организации биологических систем (в том числе генетическая и фенотипическая гомология), имеет сложный и неоднозначный характер).
Некоторые мутации, возможные для данного генотипа в биохимическом отношении (т. е. разрешенные на генетическом уровне), в конечном итоге, однако, приводят к гибельным для развивающегося организма последствиям в виде нарушений морфогенеза (летальные мутации, например, морфогенетические последствия мутации врожденной гидроцефалии у домовой мыши, см. с. 325). Каждый онтогенез может быть изменен лишь определенным образом, т. е. в рамках соответствующего спектра возможных онтогенетических изменений. Это еще более сужает выбор возможных направлений эволюционных преобразований.
Наконец, существуют также морфофизиологические эволюционные ограничения и запреты, действие которых (так же как и соответствующих преадаптации) в полной мере проявляется лишь в масштабах макроэволюции, являясь одной из специфических причин ее направленного характера. Они обусловлены разнообразными взаимосвязями в пределах морфофизиологических систем и между этими системами в фенотипе взрослых организмов. При этом мутации и перестройки онтогенеза, которые могли бы привести к соответствующим изменениям фенотипа, сами по себе вполне возможны, и мутантные особи с определенной частотой могут появляться в популяциях данного вида. Однако возникающие при этом изменения фенотипа (даже, казалось бы, обладающие высокой приспособительной ценностью!) не могут быть использованы для формирования новых адаптации из-за своего несоответствия морфофизиологической организации данного вида. Такие преобразования остаются неосуществимыми до снятия соответствующих морфофизиологических запретов.
Так, например, в эпидермисе земноводных могут развиваться ороговения -- имеются необходимые для этого биохимические предпосылки, отсутствуют и морфогенетические запреты для этого процесса. И действительно, локальные ороговения эпидермиса развиваются в покровах некоторых видов амфибий (например, роговые когти у шпорцевых лягушек или у самцов когтистых тритонов, роговые "зубы" у головастиков многих видов бесхвостых амфибий). Однако для земноводных оказалось невозможным сформировать на этой основе такие ороговения покровов, которые могли бы эффективно защищать организм от обезвоживания в воздушной среде и в соленых водоемах, как у рептилий, птиц и млекопитающих. Это обусловлено необходимостью для земноводных сохранять постоянно влажную поверхность кожи, которая используется как дополнительный орган газообмена, в первую очередь для выведения из организма углекислого газа (подробнее см. ниже).
Морфофизиологические эволюционные ограничения и запреты обусловлены необходимостью гармонических перестроек систем организма, интегрированных адаптивно (т.е. входящих в общий адаптивный комплекс), функционально или хотя бы топографически. В филогенезе действие таких ограничений проявляется в форме различных координаций (т.е. филогенетических корреляций) между различными структурами и системами организма. Под топографическими координациями понимаются наиболее простые сопряженные эволюционные изменения органов, тесно связанных пространственно. Например, увеличение размеров глаз невозможно без соответствующих перестроек черепа, изменений положения мышц, сосудов и нервов в глазнице и височной области. Динамические координации представляют собой филогенетические взаимоотношения органов, связанных друг с другом в онтогенезе функциональными корреляциями. Примером эволюционных ограничений, основанных на таких координациях, является невозможность усиления какой-либо группы мышц без соответствующего усиления структур скелета и некоторых других мышечных групп, поскольку это сделало бы согласованную работу скелетно-мышечной системы механически несовершенной. Так, нет смысла в развитии мощных бедренных мышц при сохранении более слабых мышц голени, так как последние не могут эффективно передавать на субстрат силу сокращения первых. В то же время мышцы голени не могут значительно усиливаться у животных, приспособленных к быстрому бегу, поскольку это существенно увеличило бы момент инерции конечности. Это эволюционное ограничение требует развития у быстро бегающих животных характерной конструкции конечностей, в которых основная масса мышц располагается в проксимальных отделах (плечо, бедро), а сила их сокращения передается к опоре через тонкие и легкие дистальные отделы (предплечье, голень, стопа) посредством системы сухожилий.
И.И.Шмальгаузен выделял еще так называемые биологические координации, под которыми понимаются сопряженные изменения органов и отдельных структур, не связанных друг с другом непосредственно какими-либо корреляциями в онтогенезе, но входящими в общий адаптивный комплекс (например, эволюционные взаимосвязи между строением жевательных мышц, зубов, челюстных костей и челюстных суставов, обусловленные определенным способом питания). Координированные эволюционные изменения этих гетерогенных структур определяются естественным отбором.
Ключевые признаки и каскадные взаимодействия
В целостном организме различные органы и структуры взаимосвязаны различными формами корреляций и координации, и цепи этих взаимосвязей сложно переплетаются друг с другом. При этом в организации многих групп организмов возникают своего рода узкие места - такие ключевые морфофункциональные особенности организации, которые через каскад различных коррелятивных и координационных взаимосвязей оказывают определяющее влияние на особенности функционирования многих зависимых систем организма.
В координационном каскаде взаимосвязанных систем состояние каждого последующего уровня определяется предшествующим, начиная от ключевой, или лимитирующей, системы. Преобразования ключевой системы могут оказывать как негативное ("запрещающее"), так и позитивное ("разрешающее") воздействие на эволюционные изменения зависимых систем организма. В первом случае они обусловливают возникновение морфофизиологических эволюционных запретов, препятствующих развитию определенных адаптации и направлений эволюционных преобразований, во втором, наоборот, снимают подобные запреты и ограничения, существовавшие прежде.
Очень интересны каскадные морфофизиологические взаимосвязи в организации земноводных, определяемые особенностями их дыхательной системы. Земноводные используют для вентиляции легких нагнетательный насос, образованный подъязычным аппаратом и его мускулатурой и представляющий собой модифицированный жаберный насос предков наземных позвоночных - кистеперых рыб. Несовершенство этого дыхательного насоса, расположенного впереди легких (при отсутствии эффективных механизмов, способных изменять объем самих легочных мешков), приводит к неполноте опорожнения легких при выдохе, к сохранению там некоторого количества застойного воздуха, к смешению отработанного и свежего воздуха в ротоглоточной полости (рис. 108). В результате хотя легкие у земноводных могут обеспечить организм достаточным количеством кислорода, но они неспособны эффективно освобождать его от углекислого газа. Это сделало необходимым развитие у земноводных дополнительного пути выведения углекислого газа - через кожу. Как мы уже упоминали, дыхательная функция кожи требует постоянной влажности покровов за счет деятельности кожных желез и запрещает интенсивное ороговение эпидермиса. Это резко ограничивает круг доступных для земноводных местообитаний как на суше (где амфибии активны, как правило, лишь при повышенной влажности), так и в воде: из-за высокой проницаемости покровов земноводные не могут осуществлять эффективную осморегуляцию в гипертонической среде соленых водоемов (где их организм обезвоживается, теряя влагу через покровы), и море для земноводных недоступно. Напротив, в гипотонической среде пресных водоемов через покровы в организм земноводных непрерывно поступает избыточное количество воды, которую должны удалять из организма органы выделения. Это препятствует развитию приспособлений для экономии воды в выделительной системе амфибий. Кровеносная система земноводных должна обслуживать два органа газообмена - легкие и кожу, причем кровь от них поступает к сердцу по разным магистральным сосудам (легочная вена, впадающая в левое предсердие, и кожные вены, несущие кровь через систему полых вен в правое предсердие). Это определяет необходимость смешивания обоих потоков крови в желудочке сердца и невозможность эффективного разделения артериальной и венозной крови. В результате этого для земноводных невозможно развитие эффективных систем терморегуляции и высокого уровня метаболизма, достижение гомойотермии для земноводных запрещено.

Очевидно, эволюцию дыхательной системы земноводных можно интерпретировать как пример инадаптивного варианта эволюционных преобразований, по В.О.Ковалевскому. В организации земноводных несовершенный способ вентиляции легких оказался ключевым эволюционным запретом для развития важных адаптации во многих зависимых системах организма.
Но нередко и весьма совершенное само по себе приспособление может сыграть роль ключевого запрета по отношению к развитию определенных адаптации других систем организма. Эффектный пример такого рода представляют каскадные взаимосвязи в организации насекомых, также определяемые особенностями их дыхательной системы. Органы дыхания насекомых образованы сложной системой трахей - разветвленных воздухоносных трубочек, пронизывающих все тело; тончайшие трахейные стволики достигают практически всех клеток. По мнению М.С.Гилярова, именно совершенство трахейной системы позволило насекомым освоить широкий круг наземных местообитаний в условиях дефицита влаги. Вместе с тем трахейная система создала предпосылки для значительной интенсификации газообмена, вплоть до достижения у некоторых видов насекомых временной гомойотермии при полете.
С другой стороны, как было показано В.Н.Беклемишевым, трахейная система определенным образом ограничивает адаптивные и эволюционные возможности насекомых, накладывая отпечаток на всю их организацию. Крайне разветвленные трахеи делают ненужным участие кровеносной системы в газообмене, и ее периферические отделы в значительной степени редуцируются. Но при таком устройстве дыхательной и транспортной систем затруднено снабжение массивных органов, что ограничивает возможности увеличения размеров тела. Огромное большинство насекомых имеет мелкие размеры (у крупных современных видов жуков, палочников, стрекоз длина тела не превышает 13-15 см, лишь у некоторых тропических палочников она достигает 26 см). "Эволюционные эксперименты" насекомых в области крупных форм были неудачны: самые крупные известные насекомые, стрекозы-меганевры, достигавшие около 70 см в размахе крыльев при длине тела около 30 см, вымерли еще в раннепермское время.
Таким образом, у насекомых специфические особенности распределительной и дыхательной систем стали причиной возникновения эволюционного запрета на увеличение размеров тела. Небольшие размеры тела, в свою очередь, определяют многие особенности организации, поведения и экологии насекомых, которые достигли высокого совершенства в "мире малых форм". Малые размеры тела ограничивают количество клеток в организме, в частности в центральной нервной системе. Для насекомых характерна автономизация периферических отделов нервной системы (замыкание рефлекторных дуг на уровне периферических ганглиев) и преобладание наследственно закрепленных автоматизированных форм поведения. Трахейная система ограничивает также возможности экологической экспансии насекомых: при огромном разнообразии форм и адаптации насекомые совершенно не смогли освоить наиболее древнюю и обширную среду жизни - толщу морской воды. Вероятно, это связано с высокой плавучестью тела насекомых, пронизанного густой сетью воздухоносных ходов.
Причины параллельной эволюции
Очевидно, виды, происходящие от общего предка и сохраняющие гомологичные гены, морфогенетические системы и структуры фенотипа, должны обладать по этим причинам значительным сходством направляющих эволюцию организменных факторов. Закономерным результатом этого является широкое распространение явлений параллельной эволюции родственных филетических линий, известное как одно из проявлений направленности филогенеза. При этом, как мы уже отмечали, гомология генов имеет существенное значение только для близкородственных видов, тогда как параллельная эволюция таксонов более высокого ранга определяется в первую очередь сходством морфогенетических и морфофизиологических факторов.
Длительной параллельной эволюции независимых филогенетических стволов в сильной степени способствуют унаследованные от общих предков каскадные взаимодействия между системами органов. Изменения ключевой системы сказываются на состоянии всего каскада зависимых от нее систем, создавая предпосылки для определенных направлений эволюционных преобразований и запрещая другие. После формирования какого-либо ключевого приспособления начинается адаптивная радиация. Возникшие при этом родственные филетические линии наследуют вместе с ключевым признаком весь комплекс обусловленных им системных взаимосвязей и соответственно весь набор эволюционных потенций, т.е. возможных направлений эволюционных изменений и эволюционных запретов. Неизбежным следствием этого является возникновение параллельных эволюционных преобразований в зависимых от ключевого признака системах организма у родственных филетических линий.
Таким образом, исторически сложившаяся организация данного таксона ограничивает для составляющих его видов выбор возможных направлений эволюции определенными рамками. Если естественный отбор длительное время благоприятствует развитию какой-либо категории адаптации (например, переработке пищи челюстями у хищников, питающихся крупной добычей, как у предков млекопитающих), направленность эволюционных преобразований в пучке родственных филетических линий получает, так сказать, "двойное обеспечение" - со стороны как организменных, так и внеорганизменных факторов. Это является причиной возникновения устойчивых эволюционных тенденций и параллельной эволюции независимых линий в филогенезе многих групп организмов.
Одним из важнейших проявлений направленности макроэволюции является поступательный характер и широкое распространение эволюционных преобразований по пути морфофизиологи-ческого прогресса. Эволюционный прогресс и связанные с ним проблемы мы рассмотрим в следующей главе.
Однако непредвзятый обзор общей картины развития жизни на Земле убедительно доказывает реальность происходившего в истории множества групп организмов постепенного усложнения и усовершенствования организации. Эволюция "от простого к сложному" и обозначается большинством ученых как эволюционный прогресс.
Основы современных представлений об эволюционном прогрессе были заложены А.Н.Северцовым, указавшим прежде всего на необходимость разграничивать понятия биологического и морфофизиологического прогресса. Под биологическим прогрессом понимается успех данной группы организмов в борьбе за существование, каким бы путем этот успех ни был достигнут. Биологический прогресс проявляется в увеличении численности особей данного таксона, расширении области его географического распространения и распадении на таксоны более низкого ранга (адаптивной радиации). Соответственно биологический регресс, характеризующийся противоположными показателями, означает неудачу данной группы организмов в борьбе за существование.
С другой стороны, морфофизиологический прогресс представляет собой эволюционное усложнение и усовершенствование организации. С позиций системного подхода морфофизиологический прогресс характеризуют как процесс качественной трансформации биологических систем в направлении от менее организованных форм к более организованным.
Согласно А. Н.Северцову, морфофизиологический прогресс является одним из возможных путей (наряду с другими выделенными им главными направлениями эволюции) для достижения биологического прогресса. Эволюционный "выбор" того или другого из этих путей определяется соотношением особенностей организации данной группы, ее образа жизни и характера изменений условий внешней среды.
Однако в целом ряде отношений морфофизиологический прогресс занимает особое положение среди главных направлений эволюционного процесса. Это определяется, во-первых, удивительно широким распространением арогенеза в эволюции организмов: усложнение и усовершенствование организации прослеживаются в той или иной форме в самых различных ветвях подавляющего большинства основных филогенетических стволов земных организмов. Противоположное направление - катагенез - встречается в целом несравненно реже, чем арогенез, и катагенез всегда связан со специфическими изменениями образа жизни - переход от активной подвижности к прикрепленной или эндопаразитической жизни. Во-вторых, основные проявления морфофизиологического прогресса характеризуются у разных групп известной общностью, которую, правда, не всегда легко обнаружить среди разнообразнейших частных проявлений. Наконец, в-третьих, особой чертой арогенеза является его поступательный характер - интеграция последовательных достижений на пути морфофизиологического прогресса, как бы далеко они ни были разделены по времени (поскольку арогенез в эволюции данного филогенетического ствола обычно чередуется с периодами алло- и эпектогенеза).
Критерии морфофизиологического прогресса
А.Н.Северцов связывал морфофизиологический прогресс с повышением дифференциации организма и интенсификацией функций (в частности, с повышением интенсивности обменных процессов и энергии жизнедеятельности организма). Позднее различными учеными (И.И.Шмальгаузен, Б.Ренш, Д.Хаксли, К.М.Завадский, А.П.Расницын) к этим двум критериям арогенеза были добавлены другие. Наиболее важными из них являются: усовершенствование интеграции организма; рационализация его устройства, т. е. упорядочение организации, ведущее к оптимизации функционирования; повышение уровня гомеостаза, т.е. способности поддерживать постоянство внутренней среды организма; возрастание объема информации, извлекаемой организмом из внешней среды, и совершенствование ее обработки и использования. Другие критерии арогенеза, предлагавшиеся разными учеными, либо являются частными производными от перечисленных основных его критериев, либо не всегда коррелируют с морфофизиологическим прогрессом. Конкретные проявления морфофизиологического прогресса в разных филогенетических стволах существенно различаются в соответствии с исторически сложившимися особенностями организации разных групп и различным характером взаимоотношений с внешней средой. Поэтому часто оказывается трудно сравнивать уровни морфофизиологического прогресса, достигнутые разными группами. Ч.Дарвин полагал даже, что "попытки сравнивать в отношении высоты организации представителей различных типов совершенно безнадежны; кто решит, кто выше, каракатица или пчела"2. Действительно, необходимые для подобных сравнений методы количественной оценки высоты организации еще не разработаны. Однако на качественном уровне сравнение по основным критериям арогенеза все же возможно: едва ли может возникнуть сомнение, что общий уровень организации насекомых выше, чем, например, у многоножек, и у любых членистоногих он выше, чем у кольчатых червей, а у последних - выше, чем у плоских червей, и т.п.
При этом важно подчеркнуть, что лишь совокупность названных основных критериев, не менее четырех из них, характеризует общий морфофизиологический прогресс (арогенез), т.е. повышение организации в филогенезе данной группы организмов. Арогенез происходил, например, в филогенезе позвоночных, членистоногих, моллюсков; среди таксонов меньшего ранга общий арогенез характерен для млекопитающих, птиц, насекомых, головоногих моллюсков.
В то же время во многих филогенетических стволах происходили прогрессивные изменения лишь по отдельным основным критериям. Так, например, челюстной аппарат змей значительно сложнее дифференцирован и совершеннее интегрирован, чем у их предков - ящериц. У змей он включает значительно больше подвижных костных элементов и обслуживающих эти движения мускулов, чем у ящериц. Это связано с приспособлением змей к проглатыванию целиком крупной добычи - в процессе глотания левая и правая половины верхней и нижней челюсти могут двигаться независимо, как бы "переступая" по телу жертвы и постепенно все дальше охватывая проглатываемый объект, причем кости змеиных челюстей несколько расходятся друг от друга, удерживаясь прочными и эластичными связками. Однако это несомненное усложнение и усовершенствование челюстного аппарата змей по сравнению с ящерицами все же нельзя расценивать как проявление общего морфофизиологического прогресса, так как по своему значению для организма в целом эти эволюционные достижения имеют частный характер: общий уровень организации змей не выше, чем у ящериц. Об этом свидетельствует и уровень их энергетических и обменных процессов, и сходство гомеостаза, ре-цепторных систем, высшей нервной деятельности и др.
Даже столь важное проявление общей интенсификации функций организма, как повышение уровня обменных и энергетических процессов, может происходить в отрыве от других аспектов морфофизиологического прогресса и в этом случае само по себе не приводит к общему повышению уровня организации. Так, среди ракообразных наиболее интенсивным обменом веществ обладают щитни (Triops cancriformis), сохраняющие примитивную и очень консервативную организацию - эти рачки являются одним из примеров персистируюших форм, поскольку их строение не претерпело заметных изменений с триасового периода (т.е. около 200 млн лет).
Для ситуаций, когда совершенствование организации происходит лишь по отдельным критериям, целесообразно использовать термины частный прогресс (предложенный А.Л.Тахтаджяном) и адаптационный прогресс (Н.В.Тимофеев-Ресовский и др.). В зависимости от степени воздействия возникающих изменений на организм в целом и от широты формирующихся адаптации, частный прогресс соответствует либо эпектогенезу (развитие эпекто-морфозов, подобных аппарату передвижения и кинетическому черепу змей), либо аллогенезу (развитие алломорфозов: ядовитых желез у змей, различных типов зубной системы у разных групп млекопитающих и т.п.).
Ароморфозы
Как уже отмечалось, А.Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологиче-скому прогрессу, ароморфозами. Согласно А.Н.Северцову, ароморфозы - это такие изменения строения и функций органов, которые имеют общее значение для организма в целом и поднимают энергию его жизнедеятельности на новый качественный уровень. Как следует из приведенных самим А.Н.Северцовым примеров, конкретное содержание ароморфозов не сводится лишь к энергетическому аспекту совершенствования организации, но охватывает любые морфофизиологические преобразования, соответствующие указанным основным критериям арогенеза (дифференциации, интеграции, рационализации и оптимизации, интенсификации функций, повышению уровня гомеостаза, возрастанию усваиваемой информации и усовершенствованию ее обработки в организме). Так, несомненными ароморфозами в эволюции позвоночных животных были: развитие механизма активной вентиляции жабер (жаберного насоса) у древнейших позвоночных посредством движений висцерального жаберного скелета, приобретение челюстного аппарата (с перестройкой передних жаберных дуг), интенсификация вентиляции жабер при развитии жаберной крышки у костных рыб, приобретение последними плавательного пузыря - гидростатического аппарата, позволяющего рыбам регулировать свою плавучесть; развитие у предков высших наземных позвоночных- - амниот зародышевых оболочек (амниона, серозы, аллантоиса), обеспечивающих возможность откладки яиц на суше; развитие мощного всасывающего (разрежающего) дыхательного насоса грудной клетки у рептилий; формирование летательного аппарата у птиц; развитие живорождения и выкармливания детенышей молоком у млекопитающих; совершенствование головного мозга у птиц, млекопитающих и человека.
Ароморфозы имеют очень высокое общее приспособительное значение, повышая независимость организма от внешней среды. В связи с этим ароморфозы ставят организмы в качественно новые отношения с внешней средой, позволяя осваивать ее ресурсы на новом уровне. Поэтому ароморфозы сохраняются в филогенезе длительное время, становясь признаками крупных таксонов. Возникающие при этом частные приспособления различных видов не затрагивают общих ароморфных признаков.
Поскольку развитие ароморфозов открывает новые возможности для освоения ресурсов внешней среды, некоторые ученые пытались, так сказать, экологизировать концепцию арогенеза, т.е. придать ароморфозам экологическое истолкование или найти экологические критерии арогенеза. В современной научной и учебной литературе можно встретить определения ароморфозов как таких изменений организации, которые позволяют освоить новую адаптивную зону (т.е. новую среду обитания или новый способ использования ее ресурсов). Иногда при этом подчеркивают, что новая адаптивная зона должна быть шире, чем у предковой группы. Однако при таком подходе, в сущности, искажается сущность концепции А.Н.Северцова, согласно которой ароморфозы - это прежде всего морфофизиологические усовершенствования, повышающие общий уровень организации, т.е. ведущие к морфофизиологическому прогрессу. В предыдущей главе было показано, что освоение новых адаптивных зон, в том числе и более широких, чем исходные, отнюдь не обязательно связано с ароморфозом и может быть достигнуто также на пути эпектогенеза или даже катагенеза.
Между разными типами изменений организмов, связанных с различными направлениями эволюционного процесса, нет резких границ. Более того, преобразования общего значения (т.е. аро- и эпектоморфозы) обычно развиваются в филогенезе из частных приспособительных изменений - алломорфозов. Так, одним из важнейших ароморфозов в эволюции позвоночных было развитие легких - органа воздушного дыхания, позволившего освоить сушу как среду обитания и (у высших наземных позвоночных) резко повысить уровень метаболизма, вплоть до достижения гомойотермии. Напомним, однако, что легкие возникли у древних хоановых рыб как дополнительный орган дыхания, который позволяет компенсировать дефицит растворенного в воде кислорода, периодически возникавший в специфических местообитаниях этих рыб (см. с. 188). В этих условиях легкие сформировались как типичный алломорфоз. У потомков древних кистеперых рыб - примитивных амфибий, начавших осваивать сушу как среду обитания, легкие стали основным органом дыхания. Для земноводных усложнение строения легких и преобразования подъязычного аппарата в роли дыхательного насоса приобрели значение эпектоморфоза. Эти преобразования оказали глубокое влияние на всю организацию земноводных и открыли для них доступ в новую обширную адаптивную зону, но не привели к существенным изменениям общего уровня организации, который у земноводных по всем основным критериям в целом не выше, чем у высших костных рыб. Наконец, для высших наземных позвоночных - амниот - высокая степень дифференциации легких и дыхательных путей и возникновение механизма всасывающего дыхательного насоса грудной клетки стали важным ароморфозом, способствовавшим значительной интенсификации газообмена и общему повышению уровня организации. Таким образом, как показал Б.С.Матвеев, один и тот же орган может иметь значение и частной адаптации, и адаптации общего значения - в зависимости от степени своего развития и общей роли в организме. В эволюции алломорфоз может преобразоваться в эпектоморфоз, а этот последний - в ароморфоз.
Причины поступательного характера арогенеза
Особое положение морфофизиологического прогресса среди разных направлений макроэволюции определяется прежде всего его поступательным характером, т. е. интеграцией последовательных ароморфозов, ведущей к постепенному общему повышению уровня организации. Эта особенность и придает эволюции по пути арогенеза черты направленности. Вопрос о причинах поступательного характера арогенеза остается одной из центральных проблем теории макроэволюции.
Ламарк, которому принадлежит заслуга первооткрывателя прогрессивной эволюции ("градации"), рассматривал ее как проявление имманентных свойств организмов, изначально заложенных в них Творцом. Идеалистическое понимание эволюционного прогресса разделяли такие крупные ученые, как К. фон Бэр, Г.Ф.Осборн, Э.Коп, Л.С.Берг, П.Тейяр де Шарден и др. Основой всех идеалистических концепций эволюционного прогресса является допущение изначально присущих организмам стремления и способности развиваться в направлении усложнения и совершенствования, далее не анализируемых и представляющих собой своего рода имманентные метафизические сущности.
Примером подобных идей может служить концепция П.Тейя-ра де Шардена, согласно которой любая энергия имеет психическую природу. В каждом материальном элементе энергия делится на две составляющие: "тангенциальную", обеспечивающую взаимодействия друг с другом элементов одного и того же порядка и соответствующую разным формам "обычной" физической энергии, и "радиальную", "которая влечет его в направлении все более сложного и внутренне сосредоточенного состояния". Радиальная энергия способствует возрастанию сложности и упорядоченности организации, т. е. является движущей силой прогрессивной эволюции.
Рациональным зерном в подобных концепциях является понимание закономерного (хотя и абсолютизируемого в метафизическом духе) характера эволюции по пути морфофизиологического прогресса. Этого понимания, к сожалению, недостает некоторым современным неодарвинистским концепциям, концентрирующим внимание на процессах микроэволюции и подчеркивающим "оппортунистический характер" отбора.
Как уже отмечалось, естественный отбор не несет в себе никаких предпосылок, которые направляли бы эволюцию именно по пути морфофизиологического прогресса. Это подчеркивал еще Ч.Дарвин, полагавший, что эволюционный прогресс является одним из следствий борьбы за существование: отбор способствует совершенствованию организмов по отношению к условиям их существования; при неуклонно нарастающем общем усложнении биотической среды (благодаря дивергенции видов) это ведет к постепенному общему повышению организации.
Развивая идею Ч.Дарвина, некоторые современные ученые, например Ю.М.Оленов, полагают, что усложнение биотической среды в качестве движущей силы прогрессивной эволюции выражается в усложнении структуры биоценозов и межвидовых взаимодействий. Усложнение биоценозов и биосферы в целом, несомненно, происходило в истории Земли, однако между этими процессами и арогенезом нет простых и однозначных соотношений. Общий характер организации экосистем (определяемый обилием видов, входящих в их состав, различными формами взаимодействий между ними, соотношениями процессов биосинтеза и деструкции органических веществ и путями передачи продуктов биосинтеза в биоценозе), вероятно, не претерпел принципиальных изменений по крайней мере с конца палеозоя, когда в основном была заселена вся суша и сложились наземные экосистемы (водные экосистемы сформировались еще раньше). Разумеется, происходили изменения видового состава биоценозов, во многих случаях -- значительные, возникали новые виды и новые высшие таксоны растений и животных, изменялся уровень биопродукции и распада органических веществ, многие экосистемы распадались и формировались новые. Особенно крупные преобразования биоценозов (биоценотический кризис) произошли на рубеже мезозоя и кайнозоя (см. с. 257). Однако нет серьезных оснований полагать, что современные экосистемы организованы принципиально иначе (не в частностях, а по общей структуре) и сложнее, чем мезозойские.
В то же время на протяжении мезозоя и кайнозоя в разных филогенетических стволах возникали ароморфозы, далеко продвинувшие эволюцию по пути морфофизиологического прогресса (достаточно упомянуть формирование организации птиц и млекопитающих, появление человека). Нужно еще раз подчеркнуть, что эти ароморфозы отнюдь не были приурочены к каким-либо значительным изменениям условий внешней среды. Например, формирование организации птиц происходило во второй половине юрского периода, когда физико-географические условия не претерпели значительных перемен. С другой стороны, существенные преобразования биоценозов часто происходили уже после возникновения новых групп (например, во второй половине мезозоя в процессе расселения по Земле покрытосеменных растений).
Таким образом, арогенез не обнаруживает отчетливой зависимости от преобразований экосистем. К морфофизиологическому прогрессу, как и к другим проявлениям направленности эволюции, в полной мере можно применить уже упоминавшееся мнение Ч.Дарвина, что характер преобразований организмов определяется в гораздо большей мере природой их организации, чем характером изменений внешних условий (хотя, как мы видели, сам Дарвин не связывал это положение с эволюционным прогрессом). Логично заключить, что специфика орогенеза в значительной степени определяется свойствами самих организмов.
В то же время все ароморфозы имеют отчетливо приспособительный характер, будучи связаны с теми или иными конкретными адаптациями - совершенно различными у разных групп организмов. Следовательно, морфофизиологический прогресс, как и прочие направления эволюции, идет под контролем естественного отбора. Но при этом, в отличие от других направлений эволюции, арогенез имеет поступательный характер: последовательные ароморфозы, связанные с разными адаптациями, интегрируются, что выражается в обшем повышении уровня организации.
Г.Стеббинс сделал попытку объяснить поступательный характер эволюционного прогресса в рамках редукционистской концепции "оппортунистического" действия естественного отбора. Он предположил, что повышение уровня организации является побочным результатом развития некоторых "оппортунистических" (т.е. возникающих ненаправленно) адаптации, достигнутый же уровень организации сохраняется - вплоть до нового случайного его повышения - просто благодаря приспособительному значению соответствующих характеристик организма.
Однако из допущения принципиальной равнозначности и ненаправленности возникновения любых адаптации следует, что равно вероятно возникновение новых адаптации, как повышающих, так и понижающих общий уровень организации, и их эффект в отношении уровня организации должен уравновешиваться. С другой стороны, при изменениях внешних условий приспособительное значение признаков организма неизбежно значительно изменяется, поэтому представления о сохранении достигнутого уровня организации как следствии приспособительного значения его характеристик являются упрошенными и неточными. Наконец, понимание эволюционного прогресса как случайного побочного результата равнозначных "оппортунистических" адаптации игнорирует реальную разнокачественность приспособлений, и в частности специфику ароморфозов и эпектоморфозов как изменений, имеющих общее значение для организма в целом. Иными словами, концепция Стеббинса полностью игнорирует эволюционную роль системной организации целостных
организмов.
Для выяснения причин поступательного характера морфофизиологического прогресса необходимо выяснить специфику ароморфных адаптации по сравнению с наиболее обычными, т. е. алломорфными.
Чтобы интегрироваться естественным отбором в поступательном процессе прогрессивной эволюции, разные ароморфозы, связанные с различными адаптациями, должны, во-первых, иметь общие черты и, во-вторых, обеспечивать важные преимущества в борьбе за существование. Общими для любых ароморфозов являются именно те основные критерии морфофизиологического прогресса, которые мы рассматривали в начале этой главы. А особые преимущества ароморфных изменений определяются важнейшими общими особенностями организации и функционирования живых систем.
Для существования организмов в изменчивой внешней среде необходимым условием является их специфическое обособление от последней при контролируемом живыми системами обмене веществ и энергии, обеспечивающем сохранение динамического равновесия организмов. Эволюционные изменения, соответствующие главным критериям морфофизиологического прогресса (дифференциация, интеграция, рационализация и др.) способствуют выполнению этого основного условия существования живых систем, повышая независимость внутренней среды организма от внешних воздействий. Тем самым ароморфозы повышают общую устойчивость живых существ в изменчивых условиях. Поэтому они имеют универсальное значение для разных групп организмов, что и определяет широкое распространение и поступательный характер арогенеза в разных филогенетических стволах.
Ограничения на пути арогенеза и ключевые ароморфозы
Хотя морфофизиологический прогресс в наибольшей степени характерен для групп организмов, образ жизни которых требовал повышения активности основных биологических функций (питание, газообмен, передвижение, размножение), даже в филогенезе автотрофных прикрепленных растений происходили несомненные ароморфные изменения. Таковы дифференцировка вегетативного тела у высших растений на различные ткани и органы, развитие проводящей, покровной и механической систем, возникновение семени и т.п. Эти преобразования соответствуют по крайней мере первым пяти основным критериям арогенеза. Проявления морфофизиологического прогресса в эволюции неподвижно прикрепленных растений не противоречат сказанному выше о регрессивных изменениях у прикрепленных животных. Катагенез наблюдается при переходе от активного подвижного образа жизни к прикрепленному, тогда как при постоянно прикрепленном образе жизни проявляется действие общей закономерности эволюции - развития в направлении арогенеза, хотя и значительно слабее выраженного, чем у активно подвижных организмов.
Специфические приспособления, связанные с конкретной средой обитания и образом жизни данной группы, нередко накладывают разнообразные морфофизиологические ограничения и запреты на дальнейшее развитие в направлении арогенеза. Для высших наземных растений автотрофность определяет относительно невысокую общую интенсивность обмена веществ и энергии; необходимость извлечения из почвы воды и минеральных солей требует развития корневой системы, которая, подобно якорю, прочно удерживает растение на месте; прикрепленный образ жизни делает ненужным развитие приспособлений для передвижения, органов чувств и общего координационного центра (нервной системы). Более того, естественный отбор благоприятствует сохранению относительно слабой интеграции растительного организма, поскольку с ней связана более высокая способность к регенерации (восстановлению утраченных частей тела) и к вегетативному размножению, чрезвычайно важная для растений, поскольку они не могут активно защищаться или избегать растительноядных животных и разного рода механических повреждений.
Точно так же во многих филогенетических стволах животных специфические адаптации могут сдерживать или препятствовать дальнейшему общему морфофизиологическому прогрессу. Например, сдерживает арогенез значительное развитие органов пассивной защиты, подобных раковине моллюсков, панцирю черепах и т.п. Однако эти ограничения не являются абсолютными: у головоногих моллюсков раковина была преобразована в гидростатический аппарат, а затем редуцировалась, и современные головоногие, как известно, достигли значительной высоты организации, вероятно не уступающей по сложности и совершенству "среднему уровню" позвоночных.
Выше мы уже рассматривали комплекс морфофизиологичес-ких ограничений и запретов в организации земноводных, связанных каскадом корреляций с ключевым запретом - несовершенным механизмом вентиляции легких. У предков высших позвоночных - амниот - развился эффективный механизм вентиляции легких - всасывающий дыхательный насос грудной клетки. Развитие этого ароморфоза повлекло за собой каскад важных перестроек организации, который можно схематизированно представить следующим образом: 1) исчезла необходимость кожного дыхания; 2) в покровах усилились процессы ороговения эпидермиса, и это позволило ликвидировать непрерывные потери влаги в воздушной среде за счет испарения с поверхности влажной кожи и обеспечило лучшую механическую защиту; 3) одновременно открылись возможности для последующей дифференциации производных покровов (развитие роговых чешуи рептилии, перьев птиц, волос млекопитающих); 4) ороговение эпидермиса позволило также решить проблему осморегуляции в водной среде, защитив организм от обезвоживания в морской воде и от переобводнения в пресной; 5) в связи с этим появилась возможность сформировать в выделительной системе физиологический механизм экономии воды за счет ее обратного всасывания из мочи и синтеза концентрированных продуктов белкового обмена, все эти преобразования открыли доступ амниотам в широкий круг наземных местообитаний, включая самые засушливые, а также в море; 6) исчезла необходимость смешивать в желудочке сердца кровь от легких и кожи, в результате появилась возможность для разделения артериальной и венозной крови в сердце в нутрижел уд очковой перегородкой, что, в свою очередь, привело к перестройкам отходящих от сердца артериальных стволов и открыло возможность повышения уровня метаболизма и развития гомойотермии; 7) утрата ротоглоточной полостью и подъязычным аппаратом роли дыхательного насоса позволила развить механически более совершенные конструкции черепа (у земноводных череп должен быть широким, низким и плоским для эффективной работы подъязычного дыхательного насоса).
Ароморфозы, существенно изменяющие взаимодействия различных систем организма, снимая существовавшие прежде эволюционные запреты и ограничения на пути арогенеза и открывая новые возможности функционирования и эволюционных преобразований всего каскада зависимых систем, были названы ключевыми. Ключевые ароморфозы открывают путь другим важным изменениям организации, которые сами также могут быть ароморфозами, но не могут развиться без предварительного осуществления ключевого ароморфоза.
Ключевые ароморфозы возникали на всех этапах развития жизни. Так, важнейшим ключевым ароморфозом было приобретение предками эукариот в середине протерозоя актино-миозиновой сократительной системы, обеспечивающей цитоплазматическую подвижность. Ее появление могло быть связано с хищническим образом жизни и развитием фагоцитоза у предков эукариот. Развитие подвижности цитоплазмы потребовало обособления ядерных структур от остальной цитоплазмы, с этим связано возникновение ядерной оболочки из специализированной части цитоплазматического ретикулума. Актино-миозиновая система внутриклеточной подвижности позволила решить проблему клеточного выделения и осморегуляции - сформировался аппарат пульсирующих вакуолей, функция которых основана на использовании этих же сократительных белков. Сократительные белки функционируют и в аппарате микротрубочек, формирование которого было, вероятно, первоначально связано с обслуживанием органелл клеточного передвижения - жгутиков. Только после развития актино-миозиновой системы стало возможно возникновение упорядоченного механизма клеточного деления - митоза.
Ключевые ароморфозы (как и другие ключевые изменения организации, см. с. 367) оказывают глубокое воздействие на направления и темпы макроэволюционных преобразований; эти вопросы будут рассмотрены в следующей главе.
Д. Кэмпбелл обратил внимание еще на один важный аспект эволюционного прогресса: по мере совершенствования организации биологические системы приобретают возрастающую способность контролировать собственную эволюцию. Это выражается, в частности, в приобретении клетками специальных комплексов энзимов (ДНК-полимераза, ДНК-лигаза, репликаза, мутаза, ре-комбиназа, транслоказа, инвертаза и др.), которые катализируют самые различные изменения молекулы ДНК, определяя возникновение соответствующих мутаций. Согласно Кэмпбеллу, гены, эволюционировавшие в определенном направлении, часто связаны в геноме с особым механизмом, повышающим способность мутировать именно в этом направлении.
В связи с этим важно отметить, что высшие организмы эволюционируют иными и притом более экономичными способами, чем более примитивные. Если основные приспособительные эволюционные изменения у прокариот связаны с глубокими биохимическими преобразованиями и происходят при выбраковке отбором огромного числа особей в каждом поколении, то у эукариот - с морфологическими преобразованиями при относительно незначительных биохимических изменениях, и только для высших животных характерны приспособительные изменения поведения, позволяющие в определенной степени снизить необходимость морфологических перестроек. Очевидно, в ходе арогенеза изменяется сам характер эволюционных преобразований.
Вероятно, повышение возможностей контроля со стороны организмов за своими эволюционными преобразованиями и становление все более гибких форм "эволюционного реагирования" организмов на изменения внешней среды можно рассматривать как еще один важный показатель арогенеза. Однако заметные сдвиги по этому показателю можно обнаружить лишь в самой широкой исторической перспективе, сравнивая качественно различные уровни организации.
Сопоставив все критерии и проявления морфофизиологиче-ского прогресса, можно заключить, что общим итогом и сущностью арогенеза является повышение независимости организма от внешней среды. Морфофизиологический эволюционный прогресс является закономерностью эволюции, обусловленной важнейшими общими свойствами живых систем и прежде всего целостных организмов: необходимостью поддерживать системный гомеостаз при обмене веществ и энергии с внешней средой. Таким образом, закономерное совершенствование организации живых систем в эволюции является следствием специфических особенностей самой этой организации.
Однако при этом необходимо учитывать и важнейшую роль естественного отбора. Как мы видели, ароморфозы развиваются на основе определенных приспособлений и сами являются особого рода приспособлениями, повышающими независимость организма от внешних условий. Единственным же фактором, обусловливающим развитие любых приспособлений, является естественный отбор. Очевидно, отбор представляет собой движущую силу арогенеза, без участия которой невозможна реализация никаких тенденций эволюционного развития, определяемых особенностями системной организации живых существ.
Вероятно, квантовый эффект в макрофилогенезе является внешним выражением действия нескольких эволюционных механизмов, основанных на системных свойствах организмов и объединяемых пороговым характером своего проявления.
Эволюционные преобразования корреляционных систем
Самый простой вариант такого механизма представляют эволюционные перестройки функциональных (эргонтических) корреляционных систем фенотипа. В целостной корреляционной системе изменение одного компонента требует соответствующих изменений других компонентов. Например, невозможно изменить форму суставной поверхности на одной кости без соответствующих изменений другой кости, сочлененной с первой (см. также примеры динамических координации, с. 366). В сущности, коррелятивные связи в морфофункциональных системах организма сами являются причиной возникновения сил мощного направленного отбора, требующих гармонических перестроек целого по принципу "все или ничего".
При этом очевидно, что одновременное или строго последовательное возникновение всех необходимых мутаций разных структурных генов, которые могли бы обеспечить гармоничное преобразование всех затронутых компонентов фенотипа, чрезвычайно маловероятно. Поэтому на первых этапах эволюционных преобразований таких корреляционных и координационных систем важную роль могут играть модификационные (ненаследственные) изменения взаимосвязанных элементов (рис. 109). Хотя такие изменения остаются в пределах прежней нормы реакции, они в совокупности могут привести к существенным преобразованиям общего фенотипического облика в очень короткие филогенетические сроки. Для палеонтолога, изучающего ископаемые остатки организмов, такие преобразования будут выглядеть как макроэволюционные.

Проиллюстрировать действие этого механизма проще всего на примере модификаций, компенсирующих различные повреждения организма, возникающие в естественных условиях или в эксперименте. Г. Понтер и Д.Уорд обнаружили среди отловленных в море окунеобразных рыб, принадлежащих к разным видам (Chaetodipterus faber, Micropogon undulatus и др.), несколько особей, у которых был полностью утрачен хвостовой плавник (вероятно, в результате нападения хищника), причем отсутствие хвоста было компенсировано разрастанием назад спинного и анального плавников и их мускулатуры. Эти рыбы были вполне жизнеспособны и, очевидно, прожили уже значительное время после утраты хвостового плавника. Конечно, в данном случае произошло не наследственное изменение организации, а лишь комплекс моди-фикационных ненаследственных изменений, компенсирующих повреждение. Однако можно полагать, что если утрата хвостового плавника будет обусловлена мутацией, то организм сможет компенсировать ее неблагоприятные последствия посредством аналогичных модификационных перестроек.
Интересные модификации скелетно-мышечной системы возникают у четвероногих млекопитающих в ответ на экспериментальное приучение к постоянному передвижению на задних конечностях. У обезьян-макаков это приводило к возникновению характерного для человека S-образного изгиба позвоночника; у крыс - к некоторому укорочению (брахицефализации) черепа, а также к усилению ягодичных мышц и изменениям формы таза, связанным с увеличением угла между лобковой и подвздошной костями. Особенно любопытно, что эти модификации в определенной степени сходны с изменениями соответствующих структур в эволюции предков человека, несомненно, связанными с переходом к двуногому передвижению.
В этом отношении интересны также результаты экспериментов И.Гармса с илистыми прыгунами (Periophthalmus) - окунеобразными рыбами (рис. ПО), обитающими в тропической приливо-отливной мангровой зоне. Во время отливов илистые прыгуны регулярно остаются на суше, где они могут переползать и даже совершать прыжки, отталкиваясь передними плавниками. Гармс выдерживал илистых прыгунов длительное время на суше и подвергал их воздействию повышенной концентрации тироксина (гормон щитовидной железы). В результате у рыбок уплотнялись ткани внутренних органов и кожа, усиливалось ороговение эпидермиса, удлинялись конечности, значительно уменьшались отолиты (слуховые камешки во внутреннем ухе, которые у рыб обычно имеют очень крупные размеры). Эти адаптивные модификации сходны с соответствующими изменениями, происходящими в онтогенезе земноводных при метаморфозе, который стимулируется повышенной секрецией тироксина щитовидной железой. Можно полагать, что эти изменения, приспосабливающие организм земноводных к жизни на суше, также могли первоначально представлять собой модификации, возникавшие в ответ на повышенную тироксинизацию, а эта последняя была результатом мутации, обусловившей усиление функции щитовидной железы.
Если возникшие подобным образом изменения фенотипа имеют адаптивную ценность, стабилизирующий отбор будет благоприятствовать наследственному закреплению новой комбинации фенотипических признаков и автономизации их онтогенетического развития. При этом отбор способствует закреплению и распространению мутаций, обеспечивающих формирование этих признаков в онтогенезе без модифицирующего влияния внешних условий - т. е. генохопий соответствующих модификаций. В связи с этим напомним подмеченный Р. Гольдшмидтом параллелизм фе-нотипического проявления мутаций и модификаций (см. с. 61). Дальнейшие перестройки онтогенеза в направлении автономизации развития этих признаков приведут ко все более ранней их закладке в эмбриогенезе. Отбор также будет способствовать новому расширению нормы реакции в том же направлении для создания нового "запаса прочности", нового резерва модификационной изменчивости, который может быть использован в последующей эволюции.
Таким образом, целостные системы организма, связанные функциональными корреляциями, преобразуются в эволюции как целое. Поэтому, как показал Л.Ридл, их преобразования должны происходить гораздо быстрее, чем при отборе независимых случайных вариаций отдельных признаков.
Каскадные корреляции и ключевые изменения
Важное значение для возникновения высоких темпов типоге-неза имеют каскадные взаимодействия между разными системами организма, рассмотренные в двух предшествовавших главах. Ключевые изменения являются своего рода "спусковым механизмом", запускающим весь каскад эволюционных преобразований в зависимых системах, имеющих важное адаптивное значение (как мы уже отмечали, эти преобразования были и раньше возможны на генетическом и морфогенетическом уровнях, но запрещены на морфофизиологическом, см. с. 365). После приобретения ключевого изменения при сильном давлении отбора, благоприятствующего развитию этих важных адаптации, эволюционные изменения должны происходить с максимально возможной скоростью. Таким образом, ключевые изменения организации являются своего рода "эволюционным триггером", значительно увеличивающим темпы макроэволюционных преобразований.
Происходящие при этом микроэволюционные изменения ничем не отличаются от обычного видообразования, и темпы микроэволюции в периоды каскадных макроэволюционных преобразований могут оставаться на среднем уровне. Однако взаимозависимость перестроек разных систем организма, скоординированных и разрешенных ключевым преобразованием, сама становится источником для возникновения мощного давления направленного отбора, благоприятствующего изменениям в заданном направлении. Скоординированность и направленность преобразований системной организации значительно ускоряют макроэволюцию.
Одним из наиболее известных примеров является каскад эволюционных преобразований у предков рептилий, связанный с развитием у них ключевого ароморфоза - механизма дыхательных движений грудной клетки, ставшего высокоэффективным всасывающим легочным насосом. Как мы видели в предыдущей главе, развитие этого насоса ликвидировало весь комплекс мор-фофизиологических ограничений и запретов, существующих в организации амфибий и обусловленных недостаточностью примитивного нагнетательного легочного насоса (подъязычного аппарата) для выведения из легких углекислого газа.
Возникновение направленности и в силу этого - высоких темпов макроэволюционных преобразований может быть обусловлено не только каскадными взаимодействиями, но и другими формами координации между различными системами организма - без ясно выраженного доминирования одной системы над другими (хотя квантовый эффект при этом, очевидно, будет выражен слабее). Такой характер макроэволюции соответствует обоснованной Т. Кемпом модели взаимосвязанного прогресса, согласно которой формирование организации нового крупного таксона происходит путем серий взаимосвязанных изменений различных систем организма. По этой модели происходило развитие основных признаков организации млекопитающих в филогенезе высших синапсидных рептилий — териодонтов. При этом отсутствие общего межсистемного ключевого признака не исключает каскадных взаимодействий в отдельных системах.
Так, сложный комплекс преобразований челюстного аппарата у териодонтов (см. рис. 62) был обусловлен ключевой адаптацией - совершенствованием переработки пищи челюстями.
Эти преобразования включают дифференциацию зубов на резцы, клыки и трехбугорчатые щечные зубы ("коренные"), увеличение размеров зубной кости нижней челюсти и ее венечного отростка, на который прикреплялась височная мышца (роль последней усиливалась при развитии механизма пережевывания пищи). Развитие этих признаков непосредственно повлекло за собой изменение обших пропорций черепа (увеличение объема височных ям, вмещающих жевательную мускулатуру, привело к относительному увеличению заглазничной части черепа), развитие вторичного челюстного сустава, разрастание зубной кости и редукцию остальных костей нижней челюсти и преобразование костей первичного челюстного сустава в косточки среднего уха. Пережевывание пищи требует жесткого давления на перерабатываемый корм, что привело к утрате примитивной подвижности нёба и верхней челюсти (кинетизма черепа). С другой стороны, задержка пищи для переработки в ротовой полости приводит к задержке дыхания, крайне нежелательной при развитии гомойо-термии. Это вызвало необходимость развития вторичного нёба, разделившего носовой ход и собственно ротовую полость. Кроме того, вторичное нёбо укрепляет верхнюю челюсть, что также важно при пережевывании пищи. Развитие всех этих преобразований было обусловлено указанной ключевой адаптацией. Весь комплекс изменений развивался скоррелированно и направленно, что доказывается многочисленными параллелизмами в разных филетических линиях териодонтов. Это определило высокие темпы типогенеза.
Ускорению темпов типогенеза могут способствовать и другие эволюционные механизмы, обладающие пороговым характером проявления. Например, таковым является механизм морфофунк-циональной преадаптации, посредством которой в филогенезе происходит смена адаптивной роли органов. После прохождения стадии преадаптационного порога перед данной группой организмов открываются новые адаптивные возможности, которые реализуются в относительно краткие (в масштабах макроэволюции) сроки. Одновременно происходит перестройка всего комплекса структур, связанных корреляциями с органом, приобретшим новую адаптивную роль.
Возникновению квантового макроэволюционного эффекта могут также способствовать дискретные изменения программ онтогенеза. Взаимодействие различных факторов в морфогенезе часто имеет пороговый характер. В этом случае сравнительно незначительные генетические перестройки, обусловленные мутациями генов-регуляторов (особенно типа гомеобоксных генов, контролирующих экспрессию целых генетических систем) могут привести к существенным преобразованиям фенотипических признаков. Например, при обработке эмбриональных зачатков кожи ретиноевой кислотой (витамин А) зачатки роговых чешуи на конечностях у эмбрионов птиц преобразуются в перья, а зачатки волос у эмбрионов млекопитающих - в железистые структуры.
В эволюции корреляционных систем изменения их концевых звеньев и боковых ветвей могут происходить более или менее независимо друг от друга - вплоть до соответствия принципу независимой эволюции разных признаков по Г. Осборну. Скорость этих изменений определяется только факторами микроэволюции, в первую очередь частотой мутаций и давлением отбора. Такие изменения происходят наиболее часто и являются основой для процессов видообразования и адаптивной радиации, не затрагивающих основной план строения крупного таксона.
Гораздо реже возникают изменения на более глубоких уровнях организации корреляционных систем. Их следствием являются координированные изменения всех взаимосвязанных компонентов. В результате происходят дискретные преобразования целого комплекса признаков, в том числе в общем плане строения. Эти преобразования также осуществляются через обычные микроэволюционные механизмы, но их координированный и направленный характер обусловливает высокие темпы макроэво-люционных перестроек. Р. Ридл показал, что темпы эволюционных преобразований интегрированных целостных систем могут быть на несколько порядков выше, чем эволюция, основанная на комбинировании независимых вариаций отдельных компонентов.
Очевидно, при необходимости быстрых координированных перестроек гетерогенного адаптивного комплекса отбор должен благоприятствовать возникновению корреляционных связей между разными его компонентами и подчинению разных признаков общему контролю в онтогенезе, вплоть до формирования целостных генетических комплексов. При этом и на молекулярном уровне формируется блочно-иерархическая организация генома, позволяющая существенно увеличить скорость эволюции за счет действия отбора на целостные блоки в геноме, контролируемые общими генами - операторами и регуляторами (см. с. 90). Иначе говоря, длительно действующий направленный отбор может благоприятствовать более глубокой интеграции систем организма, которая способствует значительному убыстрению макроэволюции, поскольку эволюционные преобразования целостной системы происходят несравненно быстрее, чем комбинирование благоприятных мутаций по отдельным признакам.
Таким образом, можно заключить, что высокие темпы типогенеза определяются прежде всего самим характером происходящих изменений организации, причем осуществляется своего рода самоконтроль темпов макроэволюции со стороны организма.
Причины макроэволюционного типостаза
Все формы корреляционных и координационных связей в целостном организме при возникновении изменений в ключевых системах способствуют комплексному характеру макроэволюци-онных преобразований, что многократно повышает темпы типогенеза. С другой стороны, стабильность общего плана строения в период типостаза обеспечивается разными механизмами системного гомеостаза, действующими на всех уровнях организации, - канализация морфогенеза под воздействием генов-модификаторов, регуляция возникающих изменений при взаимодействиях разных морфогенетических систем, мультифункциональность и функциональная пластичность фенотипических структур и др. (Мультифункциональность - выполнение данным органом или структурой нескольких функций, связанных с разными адаптациями. В этом смысле термин "мультифункциональность" иногда заменяют на "мультиадаптивность").
Вообще степень эволюционного консерватизма данной системы в целом пропорциональна ее способности к авторегуляции. Например, в скелетно-мышечном комплексе мускулы обладают более высокой онтогенетической пластичностью, чем кости; в онтогенезе строение и функции мышц относительно легко модифицируются в соответствии с изменениями нагрузок. К тому же мышцы обладают высокой функциональной лабильностью, т.е. способны обеспечивать различные оттенки движений сокращением разных пучков мышечных волокон в различных комбинациях. Иными словами, мышцы обладают высокими авторегуляционными возможностями. Следствием этого является возможность для данного мышечного комплекса удовлетворительно функционировать при разнообразных адаптивных требованиях, соответствующих различным направлениям отбора. Скелетные же элементы обладают авторегуляционными способностями в значительно меньшей мере, чем мышцы. Поэтому они не могут адекватно реагировать адаптивными модификациями на существенные изменения условий функционирования и должны отвечать на изменения направления отбора наследственными перестройками, основанными на соответствующих мутациях. В результате этого в эволюции скелетные компоненты в целом оказываются менее консервативными, чем мышечные.
При этом целостные морфофункциональные системы (например, скелетно-мышечные комплексы конечностей или челюстного аппарата) проявляют более высокий эволюционный консерватизм, чем отдельные их компоненты (в данном примере кости и мышцы). Это связано с более значительными возможностями авторегуляции систем высокого ранга, в которых авторегуляционные приспособления отдельных элементов дополняются соответствующими свойствами всей системы. Еще Галену было известно, что целый череп прочнее по отношению к ударам, чем его отдельные кости. Причиной этого являются амортизационные свойства шовных соединений между костями.
У многоклеточных организмов авторегуляционные возможности основных морфогенетических систем, обеспечивающих в онтогенезе формирование общего плана строения данной группы, в целом значительно выше, чем таковые генотипа или отдельных фенотипических признаков. Поэтому, как уже было отмечено в главе об эволюции онтогенеза, аппарат наследственности преобразуется в макрофилогенезе значительно быстрее, чем основные фенотипические структуры. Наиболее устойчивы и консервативны в макроэволюции эпигенетические корреляционные системы организма, обладающие наибольшими авторегуляционными возможностями.
Таким образом, корреляционные системы организма, представляющие собой гомеостаты, проявляют тенденцию к эволюционной стабильности. Можно допустить, что нормальное функционирование таких систем происходит в определенных режимах, действующих в пределах основных состояний или типов их конструкции, промежуточные состояния между которыми функционально неэффективны и поэтому эволюционно неустойчивы. Это обусловливает дискретность макроэволюционных преобразований, т.е. их квантовый характер. Макроэволюция представляет собой переход от одного устойчивого состояния морфофункциональных, морфогенетических и генетических систем организмов к другому их устойчивому состоянию.
В целом можно сказать, что характерные пульсирующие темпы макроэволюции, т.е. чередование типогенеза и типостаза в макрофилогенезе, являются следствием основных свойств корреляционных биологических систем, и в первую очередь - целостного организма.
Жизнь представляет собой форму существования сложных органических соединений - нуклеопротеидов, структурно организованных в виде биологических систем разного ранга. Структурными единицами живой материи являются интегрированные целостные системы - организмы. Сложность живых систем сама по себе повышает их уязвимость, снижает устойчивость перед лицом изменений внешней среды. Разрушению и гибели живых систем при неблагоприятных изменениях внешних условий противостоят два их важнейших свойства: 1) способность к саморегуляции (системный гомеостаз) и 2) способность к самовоспроизведению, основанная на свойствах молекул нуклеиновых кислот (редупликация) и ведущая к размножению систем данного класса.
Эволюция организмов представляет собой процесс исторических преобразований всех уровней организации биологических систем - от молекулярного до биосферного. Эволюция является неизбежным следствием, вытекающим из основных свойств организмов - размножения и редупликации аппарата наследственности. В изменчивых внешних условиях эти процессы неизбежно сопровождаются возникновением мутаций, поскольку устойчивость любой системы имеет свои пределы. Наличие наследственных вариаций строения и функций организмов при борьбе за существование ведет к возникновению естественного отбора - преимущественного выживания и преимущественного оставления потомства организмами, лучше приспособленными к условиям существования. Результатом действия естественного отбора является приспособительная эволюция организмов. Можно сказать, что эволюция есть форма существования организмов в изменяющейся внешней среде.
Эволюционные изменения разных уровней иерархической организации живых систем (молекулярного - организменного - популяционного) взаимосвязаны, но их связи не имеют простого и однозначного характера. Лабильность этих связей допускает определенную степень независимости и специфику преобразований разных уровней. Естественный отбор непосредственно воздействует на фенотипы целостных организмов в популяциях данного вида. Адаптивные преимущества определенных фенотипов повышают шансы на их выживание и оставление ими потомства по сравнению с хуже приспособленными вариантами. Это приводит к преимущественному воспроизведению соответствующих генотипов и, как следствие, к распространению в популяции генетических комплексов ("супергенов") и отдельных аллелей, обусловливающих развитие адаптивных фенотипических признаков. Результатом этого являются направленные преобразования генофондов популяций, которые представляют собой основное содержание микроэволюционных процессов. При достаточно длительной изоляции разных популяций друг от друга изменения в их генофондах обычно приобретают различные направления, что приводит в конечном итоге к невозможности обмена генетической информацией между ними и тем самым к обособлению изолированных популяций друг от друга в качестве самостоятельных новых видов. Видообразование может происходить как постепенно, в течение длительных сроков, так и сальтационно (при смене немногих поколений) в результате "генетических революций" в популяциях основателей, при полиплоидизации, гибридогенезе, стасигенезе.
Помимо основных факторов эволюции (мутации и отбор) на характер протекания микроэволюционных процессов оказывают влияние различные дополнительные факторы (изоляция популяций, изменения их численности, дрейф генов, молекулярный драйв, гибридизация).
В связи с некоторыми особенностями организации генетического кода (вырожденность, наличие молчащих участков ДНК, повторные последовательности нуклеотидов) и функциональной эквивалентностью ряда аминокислот, на молекулярном уровне эволюции биологических систем происходит более или менее непрерывное накопление относительно нейтральных в адаптивном отношении наследственных изменений, не отражающихся на фе-нотипическом облике и не ведущих к развитию приспособлений организмов ("молекулярная эволюция"). При изоляции друг от друга разных популяций накопление в их генофондах различных изменений может привести к обособлению независимых филетических линий, т. е. к возникновению новых видов, различия между которыми первоначально не имеют приспособительного характера. Однако основным содержанием эволюции организмов является развитие приспособлений к изменениям условий внешней среды, которое может происходить только под воздействием естественного отбора. Сложные приспособления, которые могут функционировать достаточно эффективно лишь в "готовом виде", формируются на основе уже существовавших прежде органов и структур при смене их адаптивной роли в процессе морфофунк-циональной преадаптации. Эволюционное возникновение качественных новообразований основано на дифференциации полимерных и избыточных структур, имеющихся на всех уровнях структурно-функциональной организации живых систем (повторяющиеся последовательности нуклеотидов, дуплицированные гены и целые хромосомы, метамерные структуры фенотипа и т.п.). Все филогенетические преобразования осуществляются на основе микроэволюционных изменений популяционных генофондов. Однако в широкой исторической перспективе макрофилогенеза при формировании организации новых крупных таксонов обнаруживается ряд общих закономерностей, которые не наблюдаются в микроэволюционных процессах и не следуют непосредственно из действия факторов микроэволюции. Таковы необратимость и направленность макроэволюции, поступательный характер морфофизиологического прогресса, чередование периодов типогенеза и типостаза. Макро- и микроэволюция соотносятся друг с другом как высший и низший уровни иерархической системной организации эволюционного процесса: макроэволюционные преобразования складываются из микроэволюционных изменений, но к ним не сводятся - в том же смысле, в котором любая целостная система не сводится просто к сумме своих структурных элементов. Если особенности протекания микроэволюционных процессов определяются свойствами популяций - биологических систем стохастического типа, с нежесткими связями взаимозаменяемых компонентов, то закономерный и направленный характер макроэволюции обусловлен свойствами целостных организмов - биологических систем корреляционного типа с высокой степенью интеграции разнокачественных элементов.
Особенности организации конкретных групп организмов создают предпосылки для развития определенных типов приспособлений и исключают развитие других типов, делают возможными одни направления эволюционных преобразований и запрещают другие. Закономерный поступательный характер эволюции по пути морфофизиологического прогресса определяется отбором на совершенствование механизмов, повышающих независимость организма от внешней среды, при контролируемом самим организмом обмене веществ и энергии со средой. Длительная устойчивость общего плана строения крупных таксонов в периоды типостаза связана с совершенствуемыми стабилизирующим отбором авторегуляционными механизмами в основных системах организмов. Если же преобразования этих систем все же происходят, корреляционные и координационные связи вовлекают в них различные другие системы организма, и вся совокупность этих преобразований развивается за относительно краткие (в масштабах макрофилогенеза) сроки. Высокие темпы типогенеза обусловлены различными организменными факторами, характеризующимися пороговым действием (триггерный эффект изменений ключевых признаков при каскадных взаимодействиях различных систем организма, морфофункциональная преадаптация, дискретные изменения программ онтогенеза и др.).
При этом отбор является основным движущим фактором эволюции, без участия которого невозможна реализация каких бы то ни было потенций развития, обусловленных системными свойствами организмов. Отбор движет эволюцию и придает эволюционным преобразованиям характер приспособлений к изменениям внешней среды, а организменные направляющие факторы определяют конкретные направления и формы происходящих эволюционных перестроек.
Закономерный и направленный характер макроэволюции позволяет поставить вопрос о возможности прогнозирования эволюции. Решение этого вопроса связано с анализом соотношений необходимых и случайных явлений в эволюции организмов. Как известно, в философии категориями необходимости и случайности обозначают разные типы связи явлений. Необходимые связи определяются внутренней структурой взаимодействующих явлений, их сущностью, коренными особенностями. Напротив, случайные связи имеют внешний характер по отношению к данному явлению, будучи обусловлены побочными факторами, не связанными с сущностью этого явления. При этом случайное, конечно, не беспричинно, но его причины лежат вне причинно-следственного ряда, определяющего сущность данного явления. Случайность и необходимость относительны: случайное для одного причинно-следственного ряда является необходимым для другого, и при изменении условий случайные связи могут превратиться в необходимые, и наоборот. Статистическая закономерность представляет собой выявление необходимых, т. е. внутренних, существенных связей среди многочисленных внешних случайных взаимодействий.
Очевидно, надежные прогнозы принципиально возможны для процессов, основанных на необходимых связях компонентов данной системы. С другой стороны, для систем, в которых действуют статистические закономерности (т. е. имеется множество независимых причинно-следственных рядов, но лишь немногие из них имеют существенный, а большинство - случайный характер) возможен лишь вероятностный прогноз. Наконец, прогнозы мало достоверны или практически недостоверны для систем, подчиненных действию нескольких независимых и примерно равнозначных рядов причинно-следственных отношений, поскольку развитие таких систем принимает случайный характер.
Как мы видели выше, возникновение мутаций и естественный отбор определяются основными (сущностными) свойствами живых систем, существующих в изменчивой внешней среде. В свою очередь, эти факторы с необходимостью обусловливают дивергентную и приспособительную эволюцию организмов. Поступательный и закономерный характер эволюции по пути морфофи-зиологического прогресса также является необходимым результатом направляемой отбором эволюции, вытекающим из основных свойств живых систем. Однако он проявляется лишь как общая тенденция в масштабе макроэволюции, т. е. как своего рода статистическая закономерность в филогенезе многочисленных групп организмов, поскольку сказывается влияние многих других причинно-следственных рядов, связанных с изменениями внешней среды. Такой же характер имеют и другие закономерности макроэволюции.
По отношению к развитию жизни на Земле эти выводы сделаны, так сказать, "задним числом" и не имеют прогностического характера. Однако на их основе мы можем с высокой достоверностью предсказать, что после возникновения жизни на любой планете организмы должны претерпеть дивергентную, приспособительную и арогенетическую эволюцию.
Конкретные же направления эволюционных преобразований любой данной группы организмов определяются, с одной стороны, действием отбора (соответствующим характеру изменений внешней среды), с другой - совокупностью организменных направляющих факторов, допускающей возможность некоторого выбора из "разрешенных" вариантов эволюционных изменений ("спектр эволюционных потенций" таксона). Действие отбора имеет статистический характер, а изменения внешней среды лишь отчасти контролируются самими организмами (например, в пределах биоценотической регуляции), но в значительной степени не зависят от них. Таким образом, изменения разных компонентов системы "организм-среда" для каждого вида определяются взаимодействиями многочисленных независимых причинно-следственных рядов, что делает прогнозирование эволюционных преобразований конкретных видов организмов в естественных условиях малодостоверным. Однако при возможности контроля за изменениями условий задача упрощается: при достаточно полном знании организации данного вида теоретически возможно определить спектр его эволюционных потенций и прогнозировать направление эволюционных преобразований при данном направлении изменений условий по крайней мере на вероятностном уровне. Сколько-нибудь полный анализ такого рода еще не осуществлен ни для одного вида организмов и является одной из актуальных задач современной эволюционистики. Наиболее слабым звеном здесь остается недостаточное понимание сущности процессов эмбриональной дифференцировки, лежащих в основе сложных соотношений генотипа и фенотипа и эволюционных преобразований онтогенеза. Это одна из интересных перспектив дальнейшего развития науки.
Что же касается направленного воздействия на виды организмов с целью их изменения в интересах человека, то оно в разных формах на эмпирической основе осуществляется уже давно. Напомним, что практика селекционеров послужила важным пунктом в цепи рассуждений, которые привели Дарвина к открытию естественного отбора. В свою очередь, селекция получила в эволюционной теории научную основу, подкрепленную ныне мощным арсеналом современных методов исследования. Развитие молекулярной биологии в последние годы открыло новые возможности для направленного воздействия непосредственно на наследственную основу организмов с целью получения новых форм: разрабатываемые ныне методы генной инженерии позволяют перестраивать молекулу ДНК с соответствующими изменениями наследственных свойств организмов.
Эволюционистика далеко еще не разрешила всего огромного круга стоящих перед нею проблем и продолжает стремительное развитие. Помимо традиционного обобщения и переосмысливания данных, полученных в области других биологических наук, начинают формироваться ее собственные методы. Среди них следует упомянуть постановку экспериментов на природных популяциях различных видов для изучения действия естественного отбора, внутри- и межвидовых отношений и их эволюционной роли. Аналогичные проблемы решаются и на модельных лабораторных популяциях с использованием методов популяционной генетики. Развиваются методы математического моделирования различных эволюционных процессов. Вероятно, в недалеком будущем важную роль в решении эволюционных проблем будут играть методы генной инженерии и экспериментального вмешательства в онтогенез.
Среди центральных проблем современной теории эволюции следует назвать коэволюцию разных видов в естественных сообществах и эволюцию самих биологических макросистем - биогеоценозов и биосферы в целом. Продолжаются оживленные дискуссии о роли в эволюции нейтральных мутаций и дрейфа генов, о соотношениях адаптивных и неадаптивных эволюционных изменений, о сущности и причинах типогенеза и типостаза в макроэволюции, неравномерности ее темпов, морфофизиологическом прогрессе и т.д. Многое еще предстоит сделать даже в наиболее разработанных областях эволюционистики - таких, как теория отбора, учение о биологическом виде и видообразовании.
Насущной задачей эволюционистики является переосмысливание и интеграция новейших данных и выводов, полученных в последние годы в области молекулярной биологии, онтогенетики и макроэволюции. Некоторые биологи говорят о необходимости "нового синтеза", подчеркивая устарелость классических представлений синтетической теории эволюции, являющейся, в сущности, в основном теорией микроэволюции, и необходимость преодоления характерного для нее узкоредукционистского подхода. Однако важнейшие положения классической синтетической теории о естественном отборе, факторах микроэволюции, механизмах видообразования и адаптациогенеза, без всякого сомнения, останутся основой обновленной эволюционной теории. Интегрирующим началом в ней может стать системный подход, плодотворность которого уже продемонстрировали современные достижения в понимании механизмов макроэволюции. В связи с этим некоторые ученые предлагают именовать формирующуюся в результате современного синтеза эволюционную теорию "системной". Приживется ли это название, покажет будущее.
Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. Палеолит. - М., 1978.
Афанасьев С.Л. Геохронологическая шкала фанерозоя и проблема геологического времени. - М., 1987.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. - 3-е изд. - М., 1964.
Белоусов Л. В. Введение в общую эмбриологию. - М., 1980.
Берг Л.С. Труды по теории эволюции. 1922-1930. -Л., 1977.
Беркнер Л., Маршалл Л. Кислород и эволюция // Земля и вселенная. - М., 1966. - № 4.
Б е р н а л Д. Возникновение жизни. - М., 1969.
Б л я х е р Л.Я. Проблема наследования приобретенных признаков. - М., 1971.
Б у н а к В. В. Род Homo, его возникновение и последующая эволюция. - М., 1980.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. - Л., 1987.
Вегенер А. Возникновение материков и океанов. - М., 1925.
Воробьева Э.И. Проблема происхождения наземных позвоночных. - М., 1992.
Галл Я. М. Становление эволюционной теории Чарлза Дарвина. - СПб., 1993.
Георгиевский А. Б. Эволюция адаптации: Историко-методоло-гическое исследование. - Л., 1989.
Гиляров М.С. Закономерности приспособлений членистоногих к жизни на суше. - М., 1971.
Грант В. Эволюционный процесс: Критический обзор эволюционной теории. - М., 1991.
Грант В. Эволюция организмов. - М., 1980.
Дарвин Ч. Происхождение видов путем естественного отбора. - СПб., 1991.
Дорн А. Происхождение позвоночных животных и принцип смены функций. - М.; Л., 1937.
Д р у щ и ц В. В. Палеонтология беспозвоночных. - М., 1974.
Дубинин П. П. Общая генетика. - 2-е изд. - М., 1976.
Завадский К.М. Развитие эволюционной теории после Дарвина (1859-1920 годы). - М.; Л., 1973.
Завадский К. М., Колчинский Э.И. Эволюция эволюции: Историко-критические очерки проблемы. - Л., 1977.
Иванов А. В. Происхождение многоклеточных. - Л., 1968. Иорданский Н.Н. Макроэволюция: Системная теория. - М.,
1994.
Иорданский Н.Н. Эволюция комплексных адаптации: Челюстной аппарат амфибий и рептилий. - М., 1990.
Кальвин М. Химическая эволюция. Молекулярная эволюция, ведущая к возникновению живых систем на Земле и на других планетах. -М., 1971.
К и м у р а М. Молекулярная эволюция: Теория нейтральности. - М..
1985.
Ковалевский В. О. Собрание научных трудов: В 3 т. - М., 1960.
Колчинский Э.И. Эволюция биосферы: Историко-критические очерки исследований в СССР. - Л., 1990.
К о у э н Р. История жизни. - Киев, 1982.
Красилов В. А. Происхождение и ранняя эволюция цветковых растений. - М., 1989.
Красилов В.А. Эволюция и биостратиграфия, - М., 1977.
Криштофович А. Н. Палеоботаника. - 4-е изд. - Л., 1957.
К э и н А. Вид и его эволюция. - М., 1958.
Кэррол Р. Палеонтология и эволюция позвоночных: В 3 т. - М..
1992.
Л а м а р к Ж. Б. Философия зоологии // Избр. произв. - М., 1955. -
Т. 1.
Левонтин Р. Генетические основы эволюции. - М., 1978.
М а и р Э. Зоологический вид и эволюция. - М., 1968.
М а и р Э. Популяции, виды и эволюция. - М., 1974.
Медников Б.М. Дарвинизм в XX веке. - М., 1975.
М е и е н С. В. Основы палеоботаники. - М., 1987.
Меттлер Л., Г р е г г Т. Генетика популяций и эволюция. - М.,
1972.
Мирзоян Э.И. Развитие учения о рекапитуляции. - М., 1974.
Мозелов А. П. Философские проблемы теории естественного отбора.-Л., 1983.
М о н и н А. С. История Земли. - Л., 1977.
Мюллер Ф., Г е к к е л ь Э. Основной биогенетический закон. -М.; Л., 1940.
Новая глобальная тектоника (Тектоника плит). - М., 1974.
Оленов Ю.М. Некоторые проблемы эволюционной генетики и дарвинизма. - М.; Л., 1961.
Оно С. Генетические механизмы прогрессивной эволюции. - М.,
1973.
Опарин А. И. Возникновение и начальное развитие жизни. - М"
1966.
Основы палеонтологии: Справочник для палеонтологов и геологов СССР: В 15т. /Под ред. Ю.А.Орлова. - М., 1958.
Развитие и смена беспозвоночных на рубеже мезозоя и кайнозоя: Мшанки, членистоногие, иглокожие / Под ред. А.Н.Соловьева. - М., 1980.
Развитие и смена органического мира на рубеже мезозоя и кайнозоя: Позвоночные. - М., 1978.
Развитие и смена простейших, кишечнополостных и червей на рубеже мезозоя и кайнозоя / Под ред. В.Н.Шиманского. - М., 1981.
Р а т н е р В. А., Жарких А. А., Колчанов Н.А. и др. Проблемы теории молекулярной эволюции. - Новосибирск, 1985.
Решетов В. Ю. Третичная история высших приматов: (Предыстория человека) // Итоги науки и техники: Стратиграфия и палеонтология. - М., 1986.-Т. 13.
Рогинский Я. Я. Проблемы антропогенеза. - 2-е изд. - М., 1977. Рубайлова П. Г. Формирование и развитие теории естественного отбора: Исторический очерк. - М., 1981.
Р у т т е н М. Происхождение жизни. - М., 1973. Р э ф ф Р., К о ф м е н Т. Эмбрионы, гены и эволюция. - М., 1986. Севериов А. Н. Главные направления эволюционного процесса. - 3-е изд. - М., 1967.
Северцов А. Н. Морфологические закономерности эволюции. - М.; Л., 1939.
Северцов А. С. Направленность эволюции. - М., 1990. Северцов А. С. Основы теории эволюции. - М., 1987. Симпсон Д. Г. Темпы и формы эволюции. - М., 1948. Современная палеонтология / Под ред. В. В. Меннера, В. П. Макриди-на. - М., 1988.
Соколов Б. С. Органический мир Земли на пути к фанерозойской дифференциации. - М., 1975.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. - М., 1982.
Старобогатов Я. И. Проблема видообразования // Итоги науки и техники: Общая биология. - М., 1985. - Т. 20.
С у т т Т. Проблема направленности органической эволюции. - Таллин, 1977.
Т а т а р и н о в Л. П. Морфологическая эволюция териодонтов и общие вопросы филогенетики. - М., 1976.
Татаринов Л. П. Очерки по теории эволюции. - М., 1987. Тахтаджян А.Л. Происхождение и расселение цветковых растений. - М., 1970.
Тейяр де Шарден П. Феномен человека. - М., 1987. Т и м о ф е е в - Р е с о в с к и и Н.В., Воронцов Н.Н., Яблоко в А. В. Краткий очерк теории эволюции. - М., 1969. Т о к и н Б. П. Общая эмбриология. - 3-е изд. - М., 1977.
Ушаков С. А., Я с м а н о в И. А. Дрейф материков и климаты Земли. - М., 1984.
Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции метазоа // Труды Палеонтол. ин-та АН СССР. - М., 1987. - Т. 226.
Федотов Д.М. Эволюция и филогения беспозвоночных животных. - М., 1966.
Филипченко Ю.А. Изменчивость и методы ее изучения. - 5-е изд. - М., 1978.
Филипченко Ю.А. Эволюционная идея в биологии: Исторический обзор эволюционных учений XIX в. - 3-е изд. - М., 1977.
Фокс С., Д о з е К. Молекулярная эволюция и возникновение жизни. - М., 1975.
Xесин Р. Б. Непостоянство генома. - М., 1986.
Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения современной генетики // Классики современной генетики. - М.; Л., 1968.
Шварц С. С. Экологические закономерности эволюции. - М., 1980.
Шеппард Ф.М. Естественный отбор и наследственность. - М., 1970.
Шиманский В. Н., Соловьев А. Н. Рубеж мезозоя и кайнозоя в развитии органического мира. - М., 1982.
Шишкин М.А. Морфология древних земноводных и проблемы эволюции низших тетрапод. - М., 1973.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. - 3-е изд. - М., 1982. - (Сер. Избр. труды).
Шмальгаузен И. И. Проблемы дарвинизма. - Л., 1969.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. - 2-е изд. - М., 1983. - (Сер. Избр. труды).
Шмальгаузен И. И. Факторы эволюиии.Теория стабилизирующего отбора. - 2-е изд. - М., 1968.
Я блоков А. В., Юсуфов А. Г. Эволюционное учение: Дарвинизм. - 3-е изд. - М., 1989.
Beyond neo-darwinism. An introduction to the new evolutionary paradigm / Eds. M.-W. Ho and P.T.Saunders. - London e. a., 1964.
Dobzhansky Т., Ayala F.J., Stebbins G.L., Valentine J.W. Evolution. - San Francisco, 1977.
Fisher R. A. The genetical theory of natural selection. - 2-nd ed. - New York, 1958.
Frazzetta Т.Н. Complex adaptation in evolving populations. -Massachusetts, 1975.
G о t h a n W., W е у 1 a n d H. Lehrbuch der Palaobotanik. 3. Aufl -Berlin, 1973.
Gould S.J. Ontogeny and phylogeny. - Cambridge, 1977.
H a r 1 a n d W. В., A r m st ro ng R. L., С ox A.V. e. a. A geological time scale 1989. - Cambridge, 1990.
Huxley J. Evolution. The modern synthesis. - 2-nd ed. - London, 1963. Kemp T.S. Mammal-like reptiles and the origin of mammals. - London^
M a rg u 1 i s L. Origin of eukaryotic cells. - New Haven, 1970.
М а у r E., P r о v i n e W. B. The evolutionary synthesis'. - Cambridge, 1981.
M u 11 e r A. H. Lehrbuch der Palaozoologie. 3. Aufl. B. 1-3. - Jena, 1976-1985.
R e n s с h B. Neuere Probleme der Abstammungslehre: Die transspezifische Evolution. 3 Aufl. - Stuttgart, 1972.
R i e d 1 R. Order in living organisms. - New York, 1978.
Simpson G.G. The major features of evolution. - 3-rd ed - New York, 1953.
Stanley S. M. Macroevolution: Pattern and process. - San Francisco, Stebbins G. L. The basis of progressive evolution. - Chapel Hill, 1969.