В истории изучения природы попытки объяснить целое через часть
на пути редукционизма неоднократно сменялись попытками объяснить части
через целое в системном подходе. В современном естествознании эти
тенденции вылились, соответственно, в господствующую идеологию дарвинизма
с молекулярной генетикой как её острием, и в
геосферно-биосферную систему взглядов, призванную выяснить возможности
устойчивого развития.

Рифейские бактериальные
строматолиты, Австралия. |
Сумма
больше слагаемых
Чтобы корректно оперировать понятием „жизнь“, нужно сначала разобраться
с понятием системы. Каковы же её характеристики?
Во-первых, любая система состоит из отдельных компонентов,
взаимодействующих между собой определённым образом. Например, простейший
одноклеточный организм состоит из 4-х компонентов: мембраны,
изолирующей клетку от окружающей среды, наследственного материала
в виде ДНК, рибосомы (аппарата для синтеза
белка) и цитоплазмы, т. е. внутренней клеточной
среды, где протекают все процессы метаболизма.

Верхний
цианобактериальный слой современного галофильного мата. Жгут Microcoleus
chthonoplastes, состоящий из более, чем 20 трихомов, одетых
одной тонкой слизистой, местами складчатой
оболочкой. |
Теперь плавно переходим
к главному тезису: свойства системы не являются простой суммой свойств
составляющих её компонентов. На этом уровне возникает новое качество.
Вспомним, например, изначальную дискретность жизни: хотя все клеточные
компоненты и исполняют конкретные биологические функции, жизнь является
свойством системы компонентов, объединённых в организм, и только
в виде организмов нам известна.
Следующее, на что нужно обратить внимание, — это неизбежная
иерархичность систем. Чтобы правильно исследовать систему, требуется
трёхуровневый анализ. То есть необходимо не только учитывать все
входящие в систему элементы, но и понять, в какую большую
систему она вписывается. Любой организм существует не сам по себе,
а лишь как составная часть какой-либо экосистемы, включающей
среду обитания. А разные экосистемы являются частями биосферы, включающей
в себя биоту (живое) и географическую оболочку Земли.
И именно эта большая система имеет определяющее значение
для существования жизни вообще.
„На заре
жизни на Земле существовало лишь прокариотное сообщество,
и становление биосферы происходило под действием только бактерий.
Все остальные события — лишь модификация биосферной системы, созданной
в период исключительного господства „невидимых“ (Г.
З.)
Кратко упомянем ещё два очень важных свойства систем,
к которым мы ещё вернёмся: первое — в систему можно
поместить только совместимые с ней элементы, при этом
для соответствия имеют значение только функциональные свойства.
И второе — функциональные системные компоненты могут быть
с успехом заменены другими со сходными функциями.
Если кто-то считает, что мы зря разбирались с основами анализа систем,
то пусть попробует вывести свойства биосферы из свойств информационных
наследственных молекул! Это будет так же некорректно, как начать
поиски „универсального общего предка“ без чёткого представления о среде
обитания и геохимических процессах, в которые он должен
вписаться, чтобы обеспечить себе длительное существование в масштабах
эволюционного времени. Чтобы понять, как возникла и эволюционировала жизнь,
нужно следовать логике анализа больших систем „сверху вниз“, от общего
к частному, что мы дальше и попытаемся сделать
при рассмотрении эволюции микробных сообществ.
Жизнь на
старте
Начнём с того, что первичную биосферу Земли сформировали бактерии или, более
строго, прокариоты (безъядерные одноклеточные
организмы). Представить себе прокариотную биосферу ранее, чем
3,5 млрд лет назад, крайне затруднительно из-за отсутствия
соответствующих геологических данных — за исключением одиночных
местонахождений метаморфизированных архейских пород. Поэтому современные
реконструкции носят несколько зыбкий характер и строятся
по аналогии.
„Эволюция
живого обычно представляется в виде генеалогического дерева, развивающегося
от немногих предковых форм, которые должны были бы вымереть как менее
приспособленные. На самом же деле в природе сосуществуют
представители всех основных групп организмов — от примитивных
до самых сложных, — когда-либо появлявшихся
в истории биосферы“ (Г. З.)
Все
свободноживущие организмы в качестве источника энергии используют
окислительно-восстановительные реакции. Простейшим микробным
сообществом можно считать сообщество
гидрогенотрофных микроорганизмов.
В качестве восстановителя они используют водород, образующийся
при реакции воды с перегретыми горными породами,
т. е. в
условиях достаточно высоких температур. Такие организмы сейчас широко
встречаются среди представителей глубинной микрофлоры, населяющей поры горных
пород примерно
до глубины 3 км. Очевидно, что на такой глубине
и в таких условиях эти реликтовые формы в незапамятные времена
могли жить, игнорируя ультрафиолетовое излучение, губительное
на поверхности при отсутствии озонового слоя.
К сожалению, из этой группировки не удаётся вывести фотоавтотрофов,
которые стали основными производителями органического вещества на Земле.
Именно эти организмы открыли на необратимо остывающей планете новый внешний
источник энергии — солнечный свет. Фотоавтотрофы начали „работать“
при температуре от нуля до примерно 60°С.
Сине-зелёные
чемпионы
Важнейшими фотоавтотрофами стали цианобактерии, известные как
сине-зелёные водоросли. Эти удивительные самодостаточные
организмы, ошибочно называемые водорослями, сейчас обитают на нашей планете
везде — в тропиках, горах, морях, даже на атомных полигонах.
С водорослями и растениями вообще этих бактерий объединяет наличие
пигмента хлорофилла, благодаря которому они осуществляют реакцию
фотосинтеза.

Реконструкция
строматолитовой постройки. |
С появлением
цианобактерий наша биотическая система становится автономной, с замкнутыми
циклами всех биогенных элементов. Цианобактерии служат первичными
производителями органического вещества и свободного кислорода
в атмосфере, а хемотрофные бактерии — деструкторами, разрушающими
органику и возвращающими её в биотический цикл. При этом
неполное разрушение приводит к накоплению углерода осадочных горных
пород — керогена.
Первые цианобактерии, сходные по строению с современными, жили, согласно
палеонтологическим данным, 2,7 млрд лет назад. Интересно, что эти
невообразимо древние организмы с успехом идентифицируются
по современным определителям — так мало изменений претерпели они
за миллиарды лет существования! Приведённая выше цифра довольно условна,
потому что, например, мелкие кокковидные сине-зелёные, ныне
населяющие океан, трудно различить даже в микроскоп. Поэтому подобные
организмы могли существовать в ещё более древние времена,
но современными методами бактериальной палеонтологии их следы пока
обнаружить нельзя.
Возраст палеонтологических находок сине-зелёных в древних
горных породах на суше равен 2,2 млрд лет. Возможно, они обитали
также на влажном грунте, в эфемерных водоёмах и в пористых
породах (как в современных пустынях и Антарктиде),
где их останки не могли сохраниться. Настоящими памятниками
сине-зелёным водорослям стали строматолиты — поразительные
„каменные ковры“, представляющие собой окаменевшие продукты жизнедеятельности
древних цианобактериальных сообществ — как продуцентов, так
и разрушителей.

Вероятно, остатки
чехлов, заключавших по несколько нитей цианобактерии
из Хубсугульских фосфоритов. |
Упомянутыми
сообществами была создана полноценная биосфера современного геохимического типа.
Функционировала она достаточно долго —
2–3 млрд лет —
и за это время сформировала устойчивый живой покров планеты.
И если в качестве критерия эволюционной продвинутости брать
численность и устойчивость организмов, то именно цианобактериальное
сообщество нужно считать абсолютным чемпионом.
Эволюционная
пирамида
Следующий этап в развитии нашей системы — появление эукариот
(организмов с полноценным клеточным ядром)
и многоклеточных организмов. Время их появления точно
не установлено, однако можно с уверенностью сказать, что ранее
1-го млрд лет назад эукариоты главных ролей на Земле
не играли.
Сразу отметим, что эукариоты и многоклеточные возникли не на пустом
месте, но вписались в прокариотную биосферу, как среду своего
обитания. Эукариотная биота была значительно „уже“ прокариотной биосферы: только
бактериальные организмы по-прежнему выносили экстремальные
условия — высокие температуры, солёность и т. д. Такие
реликтовые микробные сообщества, сохранившиеся до сих пор
в высокоминерализованных озёрах, морских лагунах и гидротермах, могут
служить моделями древней биосферы.

Современный
галофильный мат. Захоронённые слои с цистами зелёной водоросли
Dunaliella salina Teod., на поверхности которых видны характерные
бугорки. |
В результате мы видим значительное сужение базиса жизни. „Новое“, как это
и полагается согласно системным представлениям, вписывается в рамки
уже существующего, а сохранение „старого“ является необходимым условием
дальнейшего поступательного развития системы. То есть эволюции. Если стоять
на этих позициях, то вместо обычного эволюционного дерева перед нами
отчётливо вырисовывается настоящая эволюционная пирамида (или, по
настроению, Вавилонская башня).
Новый
«старый» мир
В новом эукариотном мире, возникшем „над“ прокариотной биосферой, появились
группы, способные выполнять те же функции, что и прокариоты. Прежде
всего, это относится к водорослям, подобно цианобактериям обладающим
способностью к фотосинтезу.
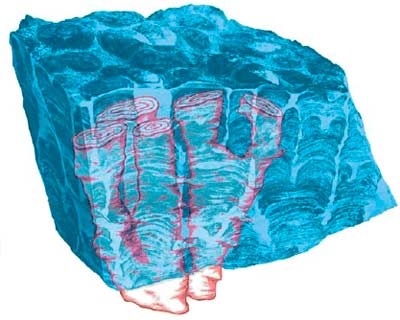
Строматолитовый монолит
с полированными поверхностями распилов. Хорошо видны тёмные
и светлые полосы — следы нарастания отдельных столбиков
за счёт выделения цианобактериями известкового материала
(образец из коллекции Центрального сибирского
геологического музея при Объединённом институте геологии, геофизики
и минералогии СО РАН, г.
Новосибирск) |
Они начинают завоёвывать
океан и вытеснять цианобактериальные сообщества с мелководий морей
и озёр. Но, в отличие
от сине-зелёных,
у водорослей есть один крупный недостаток: они не в состоянии
фиксировать атмосферный азот.
Эту проблему биосфера решила на уровне сообщества: избыточный синтез
безазотистого органического вещества эукариотами стал источником для жизни
азотфиксирующих бактерий, разрушителей органики. Это служит наглядным
примером того, что главным действующим лицом в природе выступает отнюдь
не отдельный организм или вид, но сообщество как кооперативное
единство.
Таким образом, приоритет в любом сообществе имеют трофические,
т. е. пищевые, цепи, создающие в нём „обмен веществ“, аналогичный
метаболизму клетки. Все группировки организмов в сообществе выполняют
сеть взаимодополняющих химических реакций, причём один организм может быть
заменён другим, отличным по происхождению и виду,
но осуществляющим такую же функцию. Индивидуум — ничто,
общество — всё! А сообщество в целом ограничивается
требованиями геофизической среды, и прежде всего — транспортом
вещества между элементами. Удовлетворяет этим требованиям только единая
замкнутая система функционально комплементарных организмов. В природе нет
„универсальных солдат“, которые в одиночку справлялись бы
со всеми задачами.

Схема первобытной
прокариотной биосферы. |
Первые
жертвы эволюции
Существенно ограничивала бактериальный мир принципиальная несовместимость
строения прокариот с питанием твёрдым субстратом. Проход твёрдой частицы
через мембрану нарушал у них работу самой клетки.

Современный
галофильный мат. Цисты Dunaniella salina (справа)
и смятые клетки пурпурной бактерии Thiocapsa (слева)
среди створок диатомовых водорослей. |
Поэтому
„хищный образ“ жизни был не для прокариот. Изменения мембранных
структур у первых простейших одноклеточных эукариот, способных захватывать
твёрдые частицы, стали крупнейшей инновацией, предшествующей прочим изменениям
в архитектуре их клеток. С этого времени оболочки клеток могли
перевариваться внутри организмов, и бактерии получили статус „жертв“.
И если раньше отмирание прокариот происходило вследствие исключительно
внутренних причин, то теперь картина была совершенно иной:
на охоту — вернее, на „пастбище“ — отправились первые
„бактероядные“ хищники…
В дальнейшей эволюции сообществ каждый шаг в усложнении организации живых
существ сопровождался образованием новых ниш для бактерий.
Так, благодаря сформированному пищеварительному тракту животные,
с точки зрения кишечной микрофлоры, представляют собой просто ходячие
трубчатые „ферментёры“. При этом для всякого усложнения пищевых цепей
и систем незыблемым правилом остаётся то, что они начинаются
с первичных производителей органического вещества, т. е. с
цианобактериально-водорослево-растительного звена,
а заканчиваются деструкторами. Круг жизни всегда должен быть замкнут.
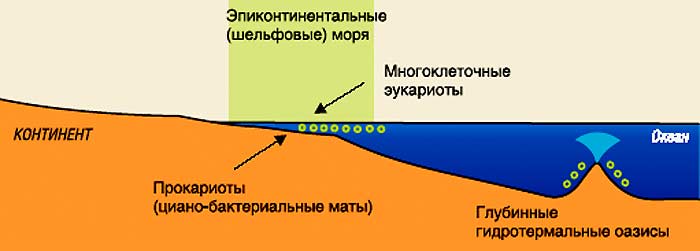
Первые этапы
пространственной экспансии жизни. |
Параллельные
миры пересекаются
И что же, в конце концов, мы видим, бросая взгляд с вершины
прогрессивной морфологической эволюции? Необратимые изменения геосферы диктовали
свои законы биоте.

Современные
бактериальные строматолиты,
Австралия. |
Появлялись, усложнялись и исчезали
организмы, но микробы во все времена как были, так и оставались
базисом планетарной системы поддержания жизни. Именно поэтому столь чётко
и слаженно работает наша огромная
геосферно-биосферная машина,
включающая в себя комплекс сопряжённых геобиохимических циклов
и процессов, которые объединены глобальным планетарным круговоротом
вещества и энергии.
Вот вам и эволюционный прогресс! При всей нашей морфологической
продвинутости мы, вкупе с животными, обеспечиваем лишь
скромные 3% от круговорота углерода на нашей планете! Просто
несопоставимо с тем, что приходится на долю микробов.
Но о них мы вспоминаем, только когда заболит горло, скиснет
молоко или „зацветёт“ любимый купальный водоём.
„Реликтовые
микробные сообщества, развивающиеся в экстремальных условиях
(например, в гидротермах или содовых
озёрах), представляют собой аналоги экосистем прошлого. Развитие
таких сообществ в местах, где „трава не растёт и скот
не пьёт“, означает, что эволюция и усложнение организмов
сопровождались сужением базы жизни“ (Г. З.)
И
ещё не было прецедента, чтобы „зелёные“ забили тревогу по поводу судьбы
редкого вида маленьких цианобактериальных стоиков. А впрочем… Разве
нуждаются в нашем признании и одобрении „сильные“ мира сего?
«Альтернативная» жизнь
Когда в 1977 г. франко-американская экспедиция обнаружила вблизи
Галапагосских островов, в безднах океана, поля гигантских двустворчатых
моллюсков, плотные стаи креветок и сплетения червевидных вестиментифер,
напоминающие ожившую макаронную фабрику, учёный мир был поражён. Никто
не ожидал встретить на многокилометровой глубине, под громадным
давлением, в беспросветной тьме, настоящие оазисы жизни. Ныне они открыты
во всех океанах — на глубинах от 400
до 7000 м.

Колония мидий, 1688 м,
Срединно-Атлантический хребет. |
В
океанических рифтовых
долинах-расщелинах раскалённая лава,
с температурой около 1200°С, поднимается из земных недр
к поверхности. Навстречу ей по трещинам сочится морская вода.
Она нагревается
до 500–850°С, обогащается соединениями
металлов и серы и „фонтанирует“. Эти источники называют
гидротермами, вентами или „чёрными курильщиками“
(из-за тёмной
взвеси сернистого
железа).

Высокотемпературный
„чёрный курильщик“, 600 м, Восточно-Тихоокеанское
поднятие. |
При отсутствии света сообщество
организмов, образующееся вокруг гидротерм, использует химическую энергию.
Его основу составляют хемосинтезирующие серные бактерии, которые производят
органические вещества за счёт энергии, выделяемой при окислении
соединений серы. Всё сообщество живёт на иждивении этих бактерий.
Вестиментифера рифтия строит трубки до трёх метров длиной, откуда
высовывает алый жаберный султан. Почти всё её тело занимает особый губчатый
орган — трофосома, где содержится до 10 млн серных бактерий.
У кольчатых червей альвинелл такие бактерии сидят прямо на поверхности
тела,
а у двустворок — в жабрах. Все эти животные служат
кормом креветкам, крабам и рыбам. Всего же ныне насчитывается более
500 видов гидротермальных животных.

Скопление креветок
и крабов, 3660 м, Срединно-Тихоокеанский
хребет. |
В 1979 году советский
геолог-тектонист Л. Зоненшайн выдвинул гипотезу о том,
что Уральские горы образовались на месте „схлопнувшегося“ океана,
и знаменитые южноуральские месторождения медного колчедана являются
ископаемыми „чёрными курильщиками“
силурийско-девонского возраста!
Действительно, со временем российские учёные нашли в них всю
ископаемую биоту, типичную для гидротерм. Постепенно „докопались“
и до более древних, докембрийских, „курильщиков“. Возник вопрос:
а не зародилась ли жизнь среди горячих серных котлов? Соединения
серы и поныне поставляют энергию клеткам, причём не только
у гидротермальных организмов. Может быть, это дань далёкому прошлому, когда
„семена“ жизни вызревали вблизи горячих серных источников?
В восьмидесятые годы в Байкале с помощью глубоководных аппаратов „Пайсис“
было обнаружено сообщество организмов, сформировавшееся в районе выброса
термальных вод.
„Микробное
сообщество возникает не путём расхождения и приобретения способностей
у определённых видов, но собирается из филогенетически удалённых,
„не родственных“ организмов, условия для которых задаются ландшафтом.
В этом смысле эволюция микробных сообществ — основная движущая сила
биогеохимической эволюции биосферы — находится в области
недарвиновских представлений“ (Г.
З.)
Источник тёплой воды с повышенной минерализацией
расположен в северной котловине Байкала в районе бухты Фролиха
на глубине
более 400 м. Вокруг фролихинской гидротермы
раскинулись „поляны“
(маты), покрытые белыми нитчатыми плёнками
из цианобактерий и нитевидных
сульфат-редуцирующих
бактерий, плотные поселения из корковых губок, в массе встречаются
различные черви, моллюски, ракообразные и рыбы. При этом если
для глубоководной фауны Байкала в целом характерно преобладание мелких
животных, то в районе вента крупные животные — не редкость.
Голубой цвет грунта в районе вента говорит об отсутствии кислорода
в непосредственной близости ко дну. Однако количество живых организмов
вокруг источника огромно. Биота фролихинского вента является аналогом
океанических донных сообществ, населяющих относительно малопрогретые
метаново-сульфатные просачивания. Этот гидротермальный оазис
с его удивительной „альтернативной“ жизнью — ещё одна уникальная черта
маленького пресноводного „океана“ Байкала.
В статье
использованы фотографии, сделанные с помощью сканирующего электронного
микроскопа, из „ATLAS of Microorganisms from Ancient Phosphorites
of Khubsugul (Mongolia)“ (2000, Huntsville, Alabama,
USA), изданного по результатам совместного проекта
Палеонтологического института РАН и NASA–Marshall Space Flight
Center (NASA/TP–2000–209901).
Об
авторе:
Заварзин Георгий Александрович — действительный член РАН,
доктор биологических наук, заведующий отделом микробных сообществ Института
микробиологии РАН.
„Наука из первых
рук“