
АКАДЕМИЯ НАУК СССР Серия «Человек и окружающая среда»
А.А. ЗАХАРОВ
МУРАВЕЙ, СЕМЬЯ, КОЛОНИЯ
ИЗДАТЕЛЬСТВО «НАУКА» Москва 1978
Ответственный редактор доктор биологических наук К. В. АРНОЛЬДИ
В книге рассказывается о жизни муравьев. Рассмотрены принципы организации семьи, ее состав и жизнедеятельность (согласованность действий, рабочий день, распределение пищи и т. д.). Особое внимание уделяется причинам возникновения колоний и комплексов, состоящих из десятков и сотен гнезд. Затронута проблема использования полезных видов муравьев для биологической защиты леса.
ВВЕДЕНИЕ
Вряд ли найдется человек, который хоть раз не останавливался возле муравейника, завороженный таким далеким и в то же время необъяснимо близким нам миром этих удивительных насекомых.
Дистанция, разделяющая нас, огромна. Заботы этих хлопотливых существ могут показаться некоторым просто недостойными серьезного разговора. Но есть в этих заботах нечто такое, что заставляет посмотреть на братьев наших меньших внимательнее и с уважением. И не потому, что их все еще много вокруг нас, что они освоили большую часть суши и почти везде стали самой сильной группой насекомых. Наверное, главное не в этом, хотя во многих случаях интерес человека к муравьям объясняется именно той важной ролью, которую играют муравьи в природе.
Нас привлекает в муравьях прежде всего то, что во многих их действиях мы можем найти какое-то подобие наших собственных дел и проблем. Муравьи относятся к тем немногим живым существам, которые не только сами приспосабливаются к среде обитания, но и активно перестраивают окружающий мир применительно к своим нуждам, своим задачам. Муравьи — вечные строители. Гнезда многих видов поражают своими размерами, сложной и рациональной архитектоникой. Дороги, тоннели, разбросанные по территории убежища для тлей и червецов, грибные сады... Разнообразные способы запасания и хранения пищи, фактическое приручение ряда видов насекомых. И все это при почти абсолютном доминировании инстинктов. Муравьи — отважные воины. К тому же они многочисленны и умеют постоять за себя. Именно благодаря этому муравьи оказались вне конкуренции среди беспозвоночных. Многие «рекорды» мира насекомых принадлежат муравьям: наибольшая продолжительность жизни особи, максимальные плотность поселения и биомасса на территории, самая высокая скорость обучения и т.п. А за всем этим сложная и отлаженная организация муравьиной семьи, позволяющая объединить и направить усилия тысяч и даже миллионов индивидов на решение главной задачи: обеспечение благополучия семьи.
Подчиняясь этой главной задаче, миновав на долгом пути эволюции множество тупиков, муравьи смогли достичь вершин организации. Суета муравейника — это всего лишь наши мимолетные впечатления, от которых не остается ни следа при ближайшем рассмотрении. На самом деле муравьев отличает четкое взаимодействие всех особей, сохраняющих при этом свою индивидуальность, но в то же время подчиненных властному диктату общины.
Миновала в науке пора описательных работ. Расширяя масштабы хозяйственной деятельности, человек повсеместно вовлекает муравьев в сферу своего непосредственного влияния, изменяет при этом свойственную им среду обитания. Одни муравьи превращаются в такой ситуации в наших противников, других мы начинаем использовать как помощников. И тут оказалось, что муравьи требуют самого серьезного отношения, и когда помогают, и когда вредят. Чисто академический интерес к муравьям сменился острой необходимостью глубокого познания особенностей их жизни, разработки биологических основ охраны и использования полезных видов и борьбы с вредными.
В изучении муравьев теперь участвуют ученые различных специальностей: лесоводы и медики, математики и экологи, специалисты по проблемам управления, бионике и защите растений. Фронт изучения муравьев постоянно расширяется, в качестве объектов исследования используются все новые и новые виды. Разнообразнее становится тематика работ. В ряде стран изучением муравьев заняты целые лаборатории.
По мере углубления наших знаний о муравьях выявляются общие принципы образа их жизни, организации. Наиболее характерное, свойственное всем муравьям качество — их «социальность», обязательное существование только сообществами (семьями, общинами). Именно социальность во всем разнообразии ее проявлений и форм у муравьев позволила им занять столь почетное место в мире беспозвоночных животных. Эволюция муравьев — это прежде всего процесс развития общинного образа жизни.
Ряд характеристик муравьиной общины свойствен и другим общественным насекомым: пчелам, общественным осам, термитам. Некоторые моменты имеют и общебиологическое значение. Поэтому изучение образа жизни муравьев никогда не было узкоспециальной задачей. Во многих отношениях муравейник представляет собой удобныи модельный объект для исследования принципов организации биологических систем, структуры популяции и ценозов и ряда важных для науки и практики проблем.
О муравьях много написано. Поэтому в данной книге не ставилась задача дать еще один развернутый очерк обо всех сторонах их жизни. Основное внимание сосредоточено здесь на способах и механизмах регуляции жизни муравьев в общине на различных уровнях развития социальности. Особый интерес представляет в этом плане переход семьи к существованию в нескольких гнездах, а также возникновение объединений муравейников. Лежащая в основе этих явлений организационная структура муравейника и представляет основную тему предлагаемой книги.
1. МУРАВЕЙ И МУРАВЕЙНИК
Говоря о муравьях, мы почти всегда подразумеваем муравейник. И это не удивительно. Одиночных муравьев в природе нет. Возникновение муравьев неразрывно связано с возникновением семьи (сообщества, общины) этих насекомых. Семья* [Звездочкой обозначены термины, которые даны в специальном словаре в конце книги.] — постоянное, многолетнее объединение отдельных насекомых, взаимодействующих друг с другом, зависящих друг от друга и от общины в целом. Связь муравья с семьей столь велика, что изолированный одиночка неизбежно погибает. Поэтому муравейник нередко воспринимается как неделимое целое. Такое впечатление создается еще и потому, что в муравейнике, так же как и в семьях других общественных насекомых: термитов, пчел и ос — каждое насекомое (индивид) выполняет лишь часть задач, необходимых для сохранения, роста и размножения вида. Общинный уклад жизни сказался как на морфологии отдельных муравьев, так и на их поведении.
Одновременно с развитием общественного образа жизни у муравьев происходило формирование в семье отдельных морфологических и функциональных групп*, связанных с выполнением различных задач, стоящих перед общиной. В ходе длительной эволюции выработался специфический для вида облик насекомых, состав, размеры и организация семьи, характер взаимодействия муравьев в гнезде и на кормовом участке, особенности индивидуального и группового поведения, пищевые связи, строение гнезда и многое другое. Все это составляет неповторимый облик каждого из нескольких тысяч видов муравьев, известных в настоящее время.
СОСТАВ СЕМЬИ
Употребление термина «семья» применительно к населению муравейника обусловлено происхождением сообщества муравьев. Эти сообщества возникли в результате последовательного усиления связей родителей со своим непосредственным потомством, а не из случайных скоплений особей, которые наблюдаются у многих беспозвоночных. Все последующие этапы развития общественного уклада жизни у насекомых — это этапы развития исходной семьи и внутрисемейных отношений отдельных насекомых. Семейная сущность такого сообщества очевидна для каждого вида независимо от того, сколь высокого уровня и сложности организации общины достиг данный вид в результате эволюции. Поэтому применительно к населению муравейника, улья «семья» — самый точный, единственно правильный термин.
Говоря о семье общественных насекомых, мы вправе употреблять и более пространные термины — «сообщество» или «община».
Семья муравьев — многолетнее, четко организованное сообщество индивидов, состоящее из половых особей (самцов и самок), а также рабочих муравьев, которые являются недоразвитыми, бесплодными в обычных условиях самками. Роль самцов сводится к оплодотворению молодых крылатых самок. Самцы обычно появляются в муравейнике незадолго до брачного лета и вскоре после спаривания погибают. Самка оплодотворяется один раз и получает при этом огромный запас спермы, которая в ее организме расходуется постепенно в течение всей жизни. Продолжительность жизни муравьиной самки максимальна для мира насекомых — до 20 лет. Оплодотворенные самки сбрасывают крылья и либо основывают новую семью, либо остаются в своем муравейнике. Иногда молодых самок принимают в другие, уже существующие семьи своего вида.
После оплодотворения для самки-основательницы наступает самый опасный период жизни — период одиночного существования. Самка должна выбрать место для Гнезда, подготовить первую камеру нового муравейника и приступить, спустя некоторое время, к яйцекладке.
У некоторых муравьев камера имеет выход на поверхность, и самка, как только выведутся личинки, периодически выходит наружу охотиться. Выходы самки продолжаются до тех пор, пока из личинок не вырастут первые рабочие. У большинства видов муравьев самка после яйцекладки постоянно находится в изолированной от внешнего мира камере, выкармливая первых рабочих и существуя сама за счет дегенерации мышц и собственного жирового тела. Никакого дополнительного питания в это время она не получает.
Одиночные самки, крылатые и уже сбросившие крылья, являются легкой добычей для многочисленных врагов: птиц, насекомоядных млекопитающих, пресмыкающихся, амфибий, хищных беспозвоночных и для других видов муравьев. Из сотен вылетевших из гнезда самок лишь единицам удается соорудить первую камеру и приступить к основанию муравейника. Еще меньше тех, чьи усилия приводят к образованию новой семьи. Поэтому в ходе эволюции многие муравьи выработали способы, позволяющие сократить до минимума или полностью исключить наиболее уязвимый период одиночного существования самки.
Далеко не все муравьи способны основывать новые семьи самостоятельно. У некоторых видов молодая самка использует для этого уже существующую небольшую семью другого вида муравьев. Проникая в чужое гнездо, она убивает живущую там самку и занимает ее место. Рабочие муравьи этой семьи обычно начинают ухаживать за пришелицей и выкармливать ее потомство — муравьев другого вида. Постепенно рабочие вида-хозяина* вымирают, и в гнезде остаются только новая самка и ее потомство. Иногда рабочие вида-хозяина ведут себя агрессивно по отношению к чужой самке. В таких случаях, если семья небольшая, она убивает не только местную самку, но и взрослых рабочих. В гнезде остаются только куколки. Выходящие из них молодые рабочие уже лояльны к чужой самке. С ними самка и основывает муравейник. Используя для организации нового муравейника семью вида-хозяина, муравьи проходят стадию так называемого временного социального паразитизма. К таким видам относятся, например, столь широко распространенные и общеизвестные обитатели наших лесов, как рыжие лесные муравьи, кроваво-красный муравей-рабовладелец, пахучий муравей-древоточец. Обычно видами-хозяевами служат бурый лесной муравей, краснощекий и серый песчаный муравьи, а для пахучего муравья-древоточца — желтый земляной муравей.
Однако и этот путь имеет свои слабые стороны. Во-первых, молодая самка должна найти небольшое гнездо определенного вида-хозяина, что само по себе задача нелегкая и связана с риском. Период одиночного существования самки хотя и сокращается, но не исключается полностью. Во-вторых, поединок с местной самкой не всегда заканчивается победой пришелицы. Наименее опасный вариант — найти семью, в которой собственная самка погибла, но и в этом случае возможность занять ее место очень мала, так как осиротевшие рабочие в такой ситуации очень быстро становятся нетерпимыми к любым самкам.
В связи с этим, по-видимому, у муравьев и появился принципиально новый способ умножения числа семей путем деления материнской общины и обособления ее частей. При этом возможны различные варианты деления: например, деление семьи пополам — гесмозис и выделение небольшого отводка — почкование*, который в дальнейшем вырастет во взрослый муравейник. Здесь самки-основательницы утрачивают свое значение при расселении вида, так как расселяются уже сформировавшиеся семьи. Правда, самка-основательница имеет и свои преимущества: она может улететь за несколько километров, а расстояние, на которое может переместиться отводок, исчисляется десятками, в лучшем случае сотнями метров. Тем не менее деление семьи становится основным способом расселения у многих муравьев.
Естественно, что возможность образования новых семей у муравьев путем деления появилась и развилась лишь как следствие специфической организации семьи муравьев, ее состава и характера взаимодействия особей и их групп. На последующих этапах развития социального образа жизни у муравьев можно выделить ряд узловых моментов, определивших то или иное направление их развития. К таким моментам относятся число самок в семье, тип гнезда, исходный принцип организации, фуражировки и другие характеристики.
Число яйцекладущих самок имеет важное значение для жизни и структуры семьи муравьев. В зависимости от этого различаются семьи моногинные* — с одной самкой — и полигинные* — с несколькими самками. Полигинные семьи могут содержать десятки и сотни яйцекладущих самок. Моногиния у муравьев, как и у других общественных насекомых, связана с однородностью состава семьи, обособленностью монотонного муравейника от других семей того же вида. Полигинные семьи, неоднородные по составу, проявляют поэтому значительную терпимость к особям из других муравейников. Поэтому у полигинных видов нередко существуют объединения взаимосвязанных семей — колонии* и федерации*.
К чисто моногинным видам относятся обитающие в Южной Америке муравьи-кочевники, широко распространенные в СССР садовые муравьи, муравьи-фаэтончики и многие другие.
Появление в семье моногинного вида нескольких оплодотворенных самок приводит к столкновениям между самками и рабочими особями, в результате чего в гнезде остается одна самка. Нередко несколько молодых самок-основательниц совместно основывают новый муравейник. Они подготавливают общую гнездовую камеру, сообща ухаживают за яйцами и личинками, которые содержатся, вместе. Но после появления первых взрослых рабочих между самками начинается ожесточенная борьба, в результате в живых остается только одна самка.
Поскольку возникновение муравейника и дальнейшее его существование во многом зависит от самок, можно было бы предположить, что они в основном и определяют жизнь муравейника. Такое положение вещей казалось настолько очевидным, что во многих работах прошлого и начала нашего века самку называли «царицей». У термитов самка и самец, постоянно живущие в семье, и сейчас именуются «царской парой». Однако в действительности оказалось, что хозяевами положения в общине являются все-таки рабочие муравьи. Чем больше в муравейнике самок, тем «непочтительнее» отношение к ним рабочих. Рабочие муравьи переселяют самок из одной части гнезда в другую, передают на обмен в другие гнезда, убивают тех, чья плодовитость стала слишком низкой. Рабочие контролируют и воспроизводство особей в семье: уничтожают лишних личинок или изменяют режимом кормления соотношение каст в семье.
Подавляющее большинство населения муравейника составляют рабочие особи, которые выполняют разнообразные функции, связанные с обеспечением нормальной жизнедеятельности семьи. Рабочие муравьи строят и охраняют гнездо, в котором обитает семья, добывают и доставляют в муравейник пищу, чистят и кормят самок и молодь, охраняют кормовой участок и т. д. Численность рабочих муравьев в семьях различных видов колеблется от нескольких десятков у наиболее примитивных до сотен тысяч и миллионов у видов, достигших вершин общественной организации.
ПРОФЕССИИ МУРАВЬЕВ
Семья — это объединение, в котором каждая особь выполняет лишь часть функций, связанных с сохранением и воспроизводством. По мере роста муравьиной общины и укрепления ее целостности разделение функций становится все более глубоким: число профессий рабочих муравьев возрастает, а специализация каждой особи сужается (для обозначения различий в выполняемых муравьями функциях специалисты, изучающие муравьев, — мирмекологи используют термин полиэтизм*).
У некоторых видов специализация проявилась в морфологическом различии особей; например, каста рабочих муравьев разделилась на солдат и рабочих. Однако у большинства муравьев разделение функций не связано с морфологическими различиями. Основами для специализации рабочих муравьев стали их физиологическое состояние и психические наклонности. Рабочие муравьи из одного гнезда, очень похожие друг на друга внешне, сильно отличаются друг от друга по возрасту и физиологическому состоянию; они в разной степени инициативны и любознательны. Одни из них смелы и агрессивны, другие отличаются робостью. Один муравей находчив, но нетерпелив, другой — может, как автомат, многократно повторять однообразные действия. Соответственно распределяются и профессии среди муравьев.
Это явилось предпосылкой для выделения в пределах касты рабочих муравьев так называемых полиэтических групп*, т. е. групп особей, занятых выполнением определенного круга обязанностей. Многочисленными опытами исследователей из разных стран установлено, что более инициативные, с быстрой реакцией муравьи выполняют работы, связанные с решением разнообразных задач в нестандартных ситуациях. Они становятся разведчиками или охотниками. Эти же муравьи гораздо хуже справляются с однообразными, не требующими активного начала обязанностями. Зато особи, имеющие замедленную реакцию и минимум любознательности, склонны к выполнению именно таких функций, например сбор пади (сладких выделений) тлей. Сборщик пади изо дня в день неторопливо ходит по одной и той же дороге к одному и тому же дереву, где на ветке имеется колония тлей. Муравей всасывает сладкие капельки и, наполнив зобик, так же неторопливо возвращается по давно известной дороге в гнездо.
В семьях всех общественных насекомых происходит смена работ, выполняемых особью в течение жизни. В большинстве случаев такая смена сводится к обязательным переходам насекомого от одной профессии к другой по мере его старения, что получило название возрастного полиэтизма. Молодые особи находятся внутри гнезда и ухаживают за самками и молодью, потом роют ходы или строят ячейки, а спустя какое-то время становятся фуражирами и добывают для семьи пищу.
У муравьев возраст тоже влияет на выполняемые рабочим обязанности, но связь эта не такая четкая. Уже через несколько дней после выхода из кокона перед рабочим муравьем появляется возможность выбора профессии. Одни муравьи ухаживают за молодью, а другие — участвуют в строительстве гнезда, становятся фуражирами, третьи — чистят помещения и т. д. Занятия муравья в этом случае не связаны с возрастом. Значительная часть особей может почти всю жизнь оставаться при исполнении одних и тех же обязанностей, например в роли нянек или свиты самки. Другие очень быстро, в течение нескольких дней, проходят как бы ознакомительный курс внутри гнезда и становятся фуражирами. Закрепление особи в одной функции получило название кастового, или постоянного, полиэтизма.
Естественно, не только наклонности индивидов определяют их обязанности в муравейнике. Муравей не может жить в одиночку. Каждый муравей — часть большой общины, в которой от нескольких сот до нескольких миллионов особей. Чтобы муравьиная семья могла успешно существовать, каждая профессиональная, или функциональная группа особей* в ее составе должна быть достаточно представительной и хорошо выполнять свои. функции. Обычно большинство особей занимается делом, в значительной мере соответствующим их индивидуальным данным. Но бывают ситуации, когда муравейник несет огромные потери и может пострадать в основном какая-нибудь одна из функциональных групп. Например, при химических обработках лесных насаждений могут погибнуть все муравьи-фуражиры, бывшие в то время на поверхности, а рабочие внутри гнезда фактически не пострадают. И тогда уцелевшим приходится менять профессии. Подобную ситуацию моделировала в садках польский мирмеколог Я.Добжанская. Экспериментальные семьи набирались из рабочих какой-либо одной функциональной группы, им придавали самку и личинок. Муравьи были вынуждены перестраиваться и выполнять все работы, необходимые для нормального существования семьи. При этом было заметно, что муравьи, набранные из одной функциональной группы, не одинаково успешно справляются с другими обязанностями. Так, муравей-охотник неуклюже и весьма неуверенно выполняет функции няньки. Тем не менее в большинстве случаев семья, составленная из рабочих особей одной функциональной группы, выживала. Понятно, что подобные ситуации бывают в жизни муравьев исключительно редко и, как правило, способности муравья соответствуют функциям, которые он выполняет.
Задачи, стоящие перед муравьями одной общины, существенно отличаются. Их можно рассмотреть на примере внегнездовых муравьев-фуражиров, обеспечивающих семью пищей. У аралокаспийского муравья-жнеца, собирающего семена пустынных растений существуют два типа фуражиров — активные и пассивные*. Фуражиры первого типа действуют на кормовом участке по-одиночке, выполняя задачи разведчиков. Их немного, не более 3% от общего числа фуражиров. Каждый разведчик имеет индивидуальный поисковый участок, который он знает и который регулярно обследует. Обнаружив на участке созревшие семена, разведчик в тот же день или на следующий производит мобилизацию* на сбор этих семян тысячи муравьев, которые по собственной инициативе на фуражировку не выходят. Мобилизованные муравьи относятся ко второй группе фуражиров, не имеющей у большинства видов определенной специализации и привязанности к какому-либо участку на территории, используемой муравейником. Они выполняют разнообразную работу в гнезде (чистка гнезда, рытье ходов и т. д.) и на поверхности делают то, на что их мобилизуют особи первой группы (сбор семян и вынос из муравейника шелухи).
Муравьи второй группы добычу на территории не ищут, а будучи приведенными к ней, не могут мобилизовать других рабочих. Они представляют собой в какой-то мере постоянный мобильный резерв «широких специалистов», пригодных к выполнению нескольких функций, но не владеющих инициативой и играющих роль пассивных исполнителей. Поэтому вторую группу фуражиров мы назвали пассивными фуражирами. Соответственно фуражиры первой группы, ведущие самостоятельный поиск, активизирующие и мобилизующие других муравьев для решения конкретных задач, будут активными фуражирами. Распространяя эти понятия на внутригнездовых рабочих, мы можем говорить об активных и пассивных особях. Поскольку такое деление основано на индивидуальных свойствах муравьев, данные группы являются полиэтическими, что отражает тип деятельности, а не конкретную функцию особо. В пределах одной полиэтической группы может быть несколько функциональных групп особей, выполняющих задачи, разные по конкретному содержанию, но сходные по типу поведения исполнителей.
Активные и пассивные фуражиры имеются в семьях большинства видов (у некоторых видов в наличии только активные фуражиры). Число активных и пассивных функциональных групп зависит от особенностей биологии вида: размеров семьи и сложности ее организации, типа гнезда и т. д. Например, с появлением наземного купола из хвои и веточек у рыжих лесных муравьев сформировалась функциональная группа муравьев-наблюдателей, впервые обнаруженная П.И.Мариковским. В эту группу входят наиболее опытные особи, мобилизующие семью на защиту муравейника при непосредственной угрозе гнезду. У многих видов появились муравьи-санитары, выносящие хитин жертв и пустые коконы на окраину охраняемой территории семьи. Более конкретные функции возникли и у части пассивных фуражиров некоторых видов. Особенно четко это выражено у муравьев, использующих тлей и других насекомых, выделяющих сладкую падь (рис. 1). Падь стала одной из основных составляющих рациона муравьев, которые в связи с этим охраняют тлей от врагов, расселяют их и даже сооружают для них специальные укрытия. Такой союз (трофобиоз*) выгоден обеим сторонам и сопровождается в конечном счете значительным увеличением численности как муравьев, так и связанных с ними насекомых.

Рис.1. Муравьи-древоточцы на колонии тлей.
Пассивные фуражиры превращаются при этом в сборщиков пади и, что представляет собой особый интерес, закрепляются за одной из колоний насекомых, выделяющих падь. Таким образом, постоянная специализация влечет за собой и территориальное закрепление муравьев. Тип деятельности пассивного фуражира при этом не меняется. Сборщик пади изо дня в день следует по одной и той же дороге к определенной колонии тлей и обратно, выполняя однообразную и лишенную инициативы работу. В первый раз к данной колонии его проводят или приносят другие муравьи. Перенесенный с дороги на 1 м в сторону, сборщик пади может потеряться я, если останется предоставленным самому себе, вообще не найдет пути в муравейник.
Сборщики пади всегда остаются резервом семьи. В случае столкновений обитателей муравейника с кем-либо из соседей, на «войну» мобилизуются сборщики пади, а активные фуражиры, занятые в других секторах кормового участка, останутся на своих местах. Следовательно, субординация рабочих муравьев двух полиэтических групп остается одинаковой у разных видов. В упомянутых выше опытах Я.Добжанской для семей муравьев, составленных из особей одной функциональной группы, был единственный вариант нежизнеспособной семьи — семья, включающая только сборщиков пади. Подобные семьи быстро погибли. Таким образом, спор об «узких» и «широких» специалистах муравьи решают в пользу первых.
Показательно, что у видов, не имеющих четкой кастовой дифференциации рабочих особей, функция, выполняемая муравьех в семье, не связана с его размерами. В тех же случаях, когда имеются четкие касты или различия в размере, профессии муравьев достаточно прочно связаны с кастовой принадлежностью или размерной группой, к которой относится индивид. Так, у степных медовых муравьев имеются крупные и мелкие рабочие, причем последние в несколько раз меньше первых. Одной из особенностей данных муравьев является наличие в их семьях «медовых бочек» — муравьев, запасающих пищу для семьи в зобике. «Бочками» служат самые крупные рабочие, а небольшие особи выполняют функции фуражиров.
На фоне многообразия профессий, имеющихся в семье муравьев, и сложности поведения, требуемой для решения ряда задач (овладение добычей, сооружение гнездового купола правильных очертаний и т. п.), особое значение приобретает появившийся у муравьев постоянный полиэтизм. Муравей получает возможность накапливать опыт, обучаться разнообразным приемам овладения добычей и борьбы с муравьями других видов, транспортировки добычи в гнездо. Он начинает легко ориентироваться на участке. Все это особенно важно для разведчиков, охотников, сборщиков строительного материала, носильщиков, наблюдателей и других полиэтических групп активных фуражиров. Следует отметить, что в связи со значительной продолжительностью жизни муравьев (рабочая особь может жить несколько лет) накопление, опыта становится важным не только для индивида, но и для семьи в целом. Постоянный полиэтизм приобретает еще большее значение у видов, ведущих групповую охоту и транспортировку добычи.
В семье одновременно присутствуют особи нескольких поколений, при взаимодействии которых происходит обучение начинающих фуражиров более опытными. Старые муравьи являются хранителями важной для жизни семьи информации. Например, у рыжих лесных муравьев каждый фуражир начинает свою внегнездовую деятельность на периферии охраняемой территории семьи. В дальнейшем он постепенно переходит на все более близкие к гнезду индивидуальные поисковые участки, а заканчивается этот путь на куполе, где муравей несет службу в качестве наблюдателя.
Каждый вышедший из фуражиров наблюдатель имеет достаточно полное представление об одном из секторов охраняемой территории гнезда. Он несет службу в той части купола, которая примыкает к этому сектору. Если в результате какой-нибудь катастрофы погибнут все фуражиры, действующие в настоящий момент на одной из дорог муравейника, информация о данном участке не будет утрачена семьей. Это проверено экспериментально.
У нескольких семей обыкновенного лесного муравья на одной из дорог изымались все фуражиры. В силу специфики организации общины у данных муравьев переход фуражиров с других дорог почти исключен. Да он и не решил бы проблемы сохранения прежней структуры территории, прилегающей к подопытной дороге, так как фуражиры с других дорог ее не знают. Однако в эксперименте в течение двух-трех дней структура участка была восстановлена во всех деталях. Это произошло благодаря сохранившимся на куполе муравьям-наблюдателям, часть которых, как установлено мечением, вернулась на территорию и восстановила ее исходную структуру. Когда же вместе с фуражирами были удалены и муравьи, находившиеся на прилегающей к подопытной дороге стороне купола, ситуация резко изменилась.
Через несколько дней, выделив из своего состава новый контингент фуражиров, семья постепенно вновь овладела потерянной частью территории, но исходная дорожная сеть и многие колонии тлей в подопытном секторе были потеряны.
В обычных условиях муравьи-наблюдатели гарантируют восстановление старой дорожной сети ~ основы охраняемой территории у муравьев при весенней активизации семей. Это — одна из причин удивительного постоянства кормовых дорог некоторых видов, возобновляющихся из года в год на протяжении многих лет.
Помимо специализации индивидов в пределах семьи каждого вида существует не менее разнообразная и четкая специализация различных видов муравьев, приспособившихся к обитанию в тех или иных условиях. Видовые профессии связаны прежде всего с пищевой специализацией муравьев. Тип питания во многом определяет и образ жизни муравьев, и профессии рабочих в общине, а также соотношение полиэтических и различных функциональных групп. Зерноядные виды сооружают гнезда со специфической структурой и имеют характерные циклы фуражировочной активности, приуроченные к срокам созревания семян различных растений. Это обычно довольно медлительные, миролюбивые, даже робкие муравьи. У них развита система массовой мобилизации пассивных фуражиров немногочисленными разведчиками. Последних мало, так как они ведут поиск не отдельного зернышка, а значительных площадей с созревшим урожаем семян, для чего многочисленные разведчики не нужны.
Хищные муравьи, наоборот, отличаются агрессивным нравом, быстротой реакции и высокой мобильностью фуражиров. Здесь преобладают активные особи, действующие зачастую в одиночку на значительном удалении от гнезда. Например, у одного из австралийских муравьев-бульдогов одиночные охотники в поисках добычи уходят от гнезда на расстояние до 2 км. Наибольшее разнообразие функциональных групп у муравьев-полифагов, употребляющих разнообразную пищу. Здесь же и относительно уравновешенное соотношение пассивных и активных фуражиров в гнездах.
У видов-полифагов достигается наиболее высокая постоянная динамическая плотность особей* на кормовом участке, в связи с чем значение мобилизации у этих муравьев уменьшается.
«Профессиональный облик» муравьев формируется и в ходе адаптации вида к специфическим условиям обитания. Много ярких примеров этого дают экстремальные условия обитания, в частности пустыня. Так, например, у обитающего на сыпучих песках бледного бегунка одним из основных занятий вне гнезда является отгребание песка от входа в муравейник. Этим постоянно занимаются сменяющие друг друга группы рабочих особей.
Для успешного существования муравьиной общины обязательно присутствие в ней всех характерных для данного вида специализаций (профессий) и оптимальное для конкретных условий соотношение представителей различных профессий. У муравьев имеются разнообразные возможности оперативно контролировать и регулировать состав муравейника, чем обеспечивается высокая жизнестойкость и пластичность общины, способность активно противостоять неблагоприятным изменениям среды и стабилизировать численность. Это недоступно большинству других насекомых.
ВЗАИМОДЕЙСТВИЕ МУРАВЬЕВ В СЕМЬЕ
Семья муравьев — объединение сотен, тысяч, а иногда и миллионов индивидов, усилиями которых сооружается гнездо, выкармливается многочисленное потомство, охраняется муравейник и его кормовой участок от посягательств агрессивных соседей. Все эти задачи остались бы невыполненными, а сама семья распалась, если бы составляющие семью муравьи не координировали своих действий, и их усилия не подчинялись общей задаче — сохранению и процветанию семьи.
Понятно, что вся жизнь семьи и каждого ее члена должна быть строго регламентирована и регулироваться весьма совершенными механизмами, выработанными в ходе длительной эволюции муравьев как общественных насекомых. Действие регулирующих механизмов основывается на постоянной и необходимой связи всех особей в единой системе. Такая связь, осуществляемая по двум каналам — пищевому и сигнальному, обеспечивает достаточный уровень координации поведения муравьев, подчиняя его общим задачам.
Основа существования семьи — обмен пищей.Муравьи регулярно обмениваются пищей — кормят друг друга. Сытый кормит голодного, тот передает часть полученной пищи следующему и т. д. (рис. 2). Обмен пищей, или трофаллаксис*, охватывает всех без исключения членов семьи. В нем участвуют не только взрослые муравьи, но и личинки и даже яйца. Одна из форм трофаллаксиса — взаимное облизывание. При этом муравьи слизывают выделения различных желез, играющие важную роль в развитии каждой особи. Пища, передаваемая изо рта в рот, также содержит вырабатываемые муравьями ферменты. Изменения содержания различных ферментов в пищевой цепи может ускорить или, наоборот, замедлить рост и развитие молоди, стимулировать взрослых муравьев (имаго) к переходу в ту или иную профессиональную (функциональную) группу. Недостаточная мощность пищевого потока означает голодание семьи и стимулирует фуражиров на поиск добычи.
Рис. 2. Обмен жидкой пищей у садовых муравьев.
Обмен пищей жизненно необходим каждому муравью, и это обстоятельство обусловливает взаимное тяготение их друг к другу. Пищевой поток, объединяющий всех членов муравьиной общины, является стержнем, сохраняющим ее как целое. Потребность каждого в партнерах по трофаллаксису стала основой не только для анатомических и физиологических особенностей муравьев, но и для развития сложных форм поведения, обеспечивающих бесперебойность пищевого потока, а в дальнейшем - разнообразие форм взаимодействия муравьев в различных ситуациях.
Выполнение особью задач семьи - закон муравейника. Функционирование каждого рабочего муравья направлено на обеспечение благополучия семьи. Действия муравья определяются в конечном счете общественной необходимостью, любой вид его деятельности носит ярко выраженный социальный характер. Это проявляется всюду. Рабочие-фуражиры добывают пищу в количествах, во много раз превышающих их индивидуальные потребности. Сбор строительного материала для сооружения гнезда у муравьев вообще беспредметен в приложении к особи и имеет смысл лишь в общине. В семьях многих муравьев имеется специальная группа муравьев-носильщиков, переносящих личинок, куколок, молодых рабочих, а иногда и самок из одной части гнезда в другую или же в родственный муравейник. Для видов, создающих в гнездах оптимальный гигротермический режим, поддержание такого режима становится одной из основных задач семьи.
Появились различные теории, в которых делается попытка объяснить безотказность муравьиной организации гипертрофированным развитием отдельных черт их психики и поведения. Например, по теории смены стимулов, выдвинутой французским энтомологом Р. Шовеном, муравей действует фактически как жестко запрограммированный семьей автомат, реакции, тип поведения которого заранее определены для каждой точки пространства. Охотник, выйдя из гнезда, не смотрит на добычу, пока не придет в предназначенную для него зону поиска добычи. Там он только ищет добычу, найдя и захватив ее, действует далее как транспорт и т. п. При этом место охоты и расстояние до него заданы в гнезде. Индивидуальная инициатива при выполнении функции, таким образом, отсутствует.
Весьма привлекательным для ряда исследователей оказался тезис об исключительном «альтруизме» рабочих муравьев, самоотверженно и без всякого вознаграждения денно и нощно пекущихся о товарищах по гнезду и даже чужих яйцах, личинках, куколках (т. е. о потомстве других особей-яйцекладущих самок).
Такие односторонние подходы не оправданы и не подтверждаются фактами. Многочисленные опыты и наблюдения за муравьями в природе показывают, что семья не в состоянии дать индивиду полную программу действий в гнезде или на участке. Его поведение всегда носит печать индивидуальности. Семья же, ее потребности лишь побуждают особь к действию, причем обычно в самой общей форме (нужна пища, холодно и т. п.). Задача и место ее выполнения конкретизируются при индивидуальном взаимодействии муравья с другими членами семьи в процессе их жизнедеятельности. Для смены стимулов не остается места.
«Альтруизм» рабочих муравьев также не выдерживает критики. Все формы ухода муравьев друг за другом и за молодью базируются на обязательной взаимности или вознаграждении. Кормя личинку, муравей получает от нее капельку насыщенной ферментами пищи. Но вот в гнезде появились жучки ламехузы, выделяющие очень привлекательные для муравьев вещества. И тут муравьи забывают о бескорыстной любви к личинкам и переключаются на уход за ламехузами - чистят их, кормят, переносят из гнезда в гнездо. Забытые личинки между тем могут погибнуть.
В каждом конкретном случае поступки муравья отражают прежде всего его собственное состояние (возрастные особенности поведения, выполняемую функцию, норму реакции на внешние раздражители, степень усталости и т. д.). И все же муравейник процветает. В основе этого процветания лежит соответствие между исходной предрасположенностью взрослого муравья к той или иной деятельности и потребностью семьи в различных функционерах. Если в муравейнике не хватает охотников, то подходящие для этого молодые рабочие быстрее обычного завершат внутригнездовую «школу» и выйдут на участок. Если обнаружится нехватка нянек, обычно самых молодых муравьев, часть бывших нянек вернется к исполнению своих прежних обязанностей. Однако муравьев, не соответствующих по складу характера дефицитной профессии, подобные перестройки обычно не затрагивают.
Оказанное выше не принижает роли семьи, но показывает, что гармоничное взаимодействие общины и индивида достигается путями, не требующими машинизации поведения рабочего муравья. Для индивида остаются значительные возможности выбора занятий и инициативы во время исполнения своих обязанностей. И если в первом случае чрезвычайные условия могут продиктовать особи и не совсем подходящее для нее место в жизни семьи, то индивидуальная инициатива функционеров просто необходима для самого существования муравейника.
Взаимодействие особей разных полиэтических групп и возрастов строится по иерархическому принципу. Активные фуражиры-разведчики у многих видов не только обнаруживают источники пищи, но и являются «поводырями» для групп пассивных фуражиров. В группе особей, мобилизованных из гнезда, разведчик играет роль вожака, лидера. Самостоятельно, без помощи разведчика пассивные фуражиры часто не могут найти обнаруженную им добычу, даже если путь к ней промечен следовыми феромонами. Однако роль лидера непродолжительна во времени и не относится исключительно к данной группе особей. Каждый разведчик потенциально является таким лидером и может мобилизовать тех или любых других пассивных фуражиров, но не мобилизует муравьев-разведчиков. У жнецов рода мессор в роли лидеров, увлекающих рабочих-сборщиков семян, одновременно могут выступать несколько разведчиков.
На поверхности купола гнезд рыжих лесных муравьев, как уже говорилось, постоянно находятся муравьи-наблюдатели, следящие за возможной опасностью и отличающиеся быстротой реакции и агрессивностью. Обычно это самые опытные члены семьи. По их тревоге все занятые на куполе рабочие муравьи почти мгновенно переключаются на оборону. Муравьи-наблюдатели играют таким образом роль активаторов, мобилизующих при необходимости массы рабочих особей на защиту гнезда.
Доминирование активных фуражиров проявляется во всех ситуациях, когда муравьи двух полиэтических групп действуют совместно. Точно так же более старые, опытные муравьи доминируют по отношению к молодым особям. Каких-либо элементов иерархии среди одновозрастных особей внутри группы пассивных фуражиров не замечено. А вот у активных иерархия проявляется в отношениях муравьев на территории, где рабочие конкурируют из-за поисковых участков, расположенных ближе к гнезду, в очередности выхода новичков в фуражиры.
Организация совместных действий.Перед семьей стоит масса задач, выполнение которых возможно только при объединении усилий нескольких муравьев. Согласованность действий участвующих в совместном деле - необходимое условие успеха. Координация действий нужна уже при общении двух особей. Во время взаимной чистки, кормления, переноски муравьями друг друга или индивидуальной мобилизации два муравья вступают в непосредственный контакт, что облегчает выработку согласованных действий.
При уходе за молодью задача облегчается, так как здесь имеется только одна активная сторона - взрослый муравей. Однако в ряде случаев требуется согласование поведения большого числа муравьев. Возможности координированных действий важны и для конкурентоспособности вида. Характерно, что воинственность муравьев прямо связана с умением особей объединяться в группы во время сражений.
Привлечение находящихся на территории фуражиров к совместной охоте или транспортировке добычи происходит обычно посредством реагирования муравьев на характерные движения и позы других особей одной общины. Это явление, получившее название кинопсис* и впервые описанное немецким мирмекологом Р.Штегером в 1931 г., особенно развито у муравьев с многочисленными семьями и высокой плотностью особей на участке. Муравей, обнаружив добычу, начинает возбужденно бегать вокруг нее, принимает агрессивную позу, атакует. Привлеченные этими почти стереотипными движениями другие муравьи устремляются к добыче. Если добыча крупная, несколько охотников совместными усилиями умерщвляют ее и тащат в гнездо. При появлении опасности сигнал тревоги прежде всего распространяется с помощью кинопсиса практически моментально и на значительные расстояния. Стоит резко наклониться над муравьиной дорогой, как тут же на метр-полтора в обе стороны все муравьи как по команде примут угрожающую позу. Еще одно неосторожное движение, и на противника обрушиваются многочисленные ядовитые струйки, десятки и сотни защитников муравейника набросятся на пришельца.
Ни одно из других известных средств передачи информации у муравьев, например феромоны тревоги, не в состоянии обеспечить столь быстрое распространение сигнала на такое расстояние.
Многое зависит от умения муравьев объединить свои усилия при транспортировке добычи в гнездо. Прежде считалось, что каждый рабочий действует при этом, самостоятельно. Поэтому при коллективной транспортировке предмет перемещается в нужном направлении даже медленнее, чем при индивидуальной. Однако такой вывод говорит лишь о том, сколь ошибочным может быть поверхностное наблюдение «издалека», не сопровождаемое тщательным анализом ситуации. Между тем детальные исследования выявили, что замедление происходит только при групповой транспортировке мелкой добычи. Примерно так же, как если бы трое людей несли, в руках один стакан воды. Когда же груз достаточно велик по своим размерам, муравьи тянут его согласованно в направлении гнезда. Английский мирмеколог Д.Садд измерил усилия, прилагаемые двумя муравьями. Оказалось, что в паре мощность каждого муравья значительно выше, чем у тех же особей, действующих поодиночке.
Имеются задачи, решение которых возможно только при постоянной координации действий множества рабочих особей. К числу таких задач относится сооружение муравейника, особенно купола гнезда, и поддержание его правильной формы и размеров. Среди различных типов гнезд муравьев наибольший интерес представляют два: секционные гнезда* и гнезда типа капсулы (рис. 3). Первый тип без наземных построек (за исключением небольших кратеров*, образованных из выброшенной на поверхность почвы). Второй тип гнезд развился из гнезд первого типа. Его отличает наземный купол и компактность размещения основных гнездовых помещений. Еще более выраженные гнездовые капсулы* имеются у многих термитов. Содержание наземного купола - задача несравнимо более сложная, нежели рытье разрозненных ходов в толще почвы. Например, гнезда рыжих лесных, тонкоголосых и садовых муравьев, несмотря на значительные размеры, всегда имеют плавные очертания, характерную для данного вида архитектонику, приспособленную к конкретным условиям обитания. Понятно, что ни один муравей не в состоянии представить себе гнездо в целом. Но как же тогда семье удается соорудить огромное по сравнению с размерами его строителей гнездо, соблюдая при этом устойчивость формы купола. Серией оригинальных экспериментов Р.Шовену удалось выявить механизмы этого явления. Обязательным условием здесь оказываются постоянные контакты муравьев-строителей по всей поверхности купола. Эти контакты должны быть равномерными.
Если такие контакты отсутствуют или их равномерность искусственно нарушена, купол начинает расти хаотично даже при равном поступлении строительного материала во все части гнезда.
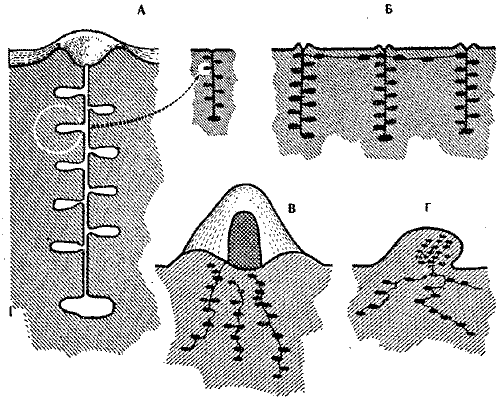
Рис. 3. Некоторые типы гнезд муравьев
Секционные гнезда: А - одосекционное гнездо (степной медовый муравей);
Б-полисекционное гнездо (пустынный дерновый муравей), секции соединяются
поверхностными тоннелями
Гнезда типа капсулы: В - гнездо с наземным
куполом из хвои и веточек (рыжие лесные муравьи), в центре купола внутренний
конус;
Г - земляная кочка (желтый лесной муравей)
Таким образом, согласованное поведение проявляется при взаимодействии любого количества муравьев. Обычно оно достигается посредством групповой иерархии* и массового подражания большинства особей муравьям-активаторам. Самые общие задачи решаются путем постоянного контакта всех участников.
Взаимное обучение муравьев.В той или иной степени обучение происходит у всех животных, какую-то часть жизни обитающих совместно, будь то птичьи базары или стаи рыб. Чем устойчивее такое объединение, прочнее связи между его членами, тем больше возможности обучения. В этом плане муравейник, в котором одновременно обитает несколько поколений рабочих муравьев, - удобная среда для появления разнообразных форм индивидуального и группового обучения.
Опыты по выяснению способностей муравьев к обучению были начаты еще в прошлом веке. С тех пор методы их проведения значительно усовершенствовались, хотя общее число опытов остается небольшим. Серьезными препятствиями для постановки и особенно толкования результатов таких опытов является сложность однозначного формулирования вопроса на «языке» муравьев и адекватной оценки их ответа, а также бытующая предубежденность в отношении интеллектуальных возможностей насекомых вообще. Тем не менее уже доказано, что муравьи быстрее всех других насекомых, а также земноводных, пресмыкающихся (лягушек, черепах) и многих птиц обучаются в лабиринте. Они хорошо отличают фигуры по их форме, например треугольник от четырехугольника, и в состоянии различать число углов при выборе пути, ведущего к кормушке. При этом им по силам инвариантные задачи (для них не имеют значения конкретная форма и размер треугольника или квадрата, а лишь совпадение числа углов).
В основе способности муравьев к обучению лежит хорошая память, используемая рабочими во время фуражировки. Наиболее простой пример обучения - повторный приход фуражира к месту, где он уже находил добычу. Г.М.Длусский показал на муравьях мирмика, что у каждого охотника есть места, которые он регулярно обследует, помня, что когда-то там ему попадалась добыча. При появлении новых удачных мест предыдущие не забываются.
В процессе обучения большое значение имеют реакции подражания. Это ключ к опосредованному обучению, т. е. обучению на опыте соседа. В жизни муравьев подражание играет большую роль благодаря почти постоянному контакту в гнезде и на участке. Муравьи воспринимают сигналы и смысл поступков не только членов своей семьи и особей своего вида, но и других видов муравьев. Еще в прошлом веке французский мирмеколог А.Форель, автор первых капитальных трудов по биологии муравьев, описывал эту ситуацию при сражении между кроваво-красными муравьями-рабовладельцами и луговыми муравьями. Стоило рабочим лугового муравья дать сигнал отступления (серия яростных ударов антеннами по телу своих собратьев), как охотники муравьев-рабовладельцев сразу же стремительно бросались на отступающих противников и отнимали куколок.
Интересен и тот факт, что в естественных поселениях муравьев охотники одного вида, доминирующего на территории, используют сигналы разведчиков других видов для обнаружения и захвата найденной теми добычи.
Очень важно, что муравьям доступно не только индивидуальное, но и групповое обучение. В семье происходит также регулярное обучение более старыми особями молодых муравьев. Одним из результатов такого обучения является удивительное постоянство дорог муравьев к колониям тлей, которые сохраняются в течение ряда лет. Примеров, подтверждающих передачу сигнальной информации от поколения муравьев к поколению, немало. При этом может полностью меняться исходный стереотип поведения обучаемого. Такое происходит, например, в семьях кроваво-красного муравья-рабовладельца, который похищает из гнезд бурого лесного муравья куколок. Вышедшие из этих куколок рабочие выполняют в муравейнике «рабовладельца» те же функции, что и у себя. Кроме того некоторые рабочие начинают помогать своим похитителям и во время грабительских набегов, участвуя в похищении и транспортировке куколок из гнезд своего вида.
Значение согласованности действий и взаимного обучения муравьев возрастает по мере увеличения численности семьи, так как происходит более узкая специализация особей, а это создает предпосылки для их более успешного обучения и приобретения профессионального опыта. Отлаженная система передачи информации в семье делает индивидуальные достижения рабочих муравьев достоянием муравейника, что увеличивает его шансы в борьбе за существование.
2. СТРУКТУРА МУРАВЕЙНИКА
Облик муравьев различен. Среди нескольких тысяч видов муравьев есть и карлики размером до 2 мм, и великаны, у которых рабочие достигают 2 см. Белесые, почти прозрачные и глянцево-черные, одноцветные, двухцветные и т. д. Разнообразны форма головы и пропорция тела, скорость передвижения и способы добывания пищи. Различаются не только отдельные особи, составляющие семью, но и семьи - и по численности муравьев, и по организации общины. У одних видов это единственная самка и несколько десятков рабочих, у других - миллион рабочих и сотни самок. Чем больше население муравейника, тем сложнее организовать ее четкую жизнедеятельность. Семья может достигнуть такого уровня численности, когда существование ее как единого целого становится просто невозможным. И тогда в недрах общины начинают образовываться более мелкие внутрисемейные структурные единицы - колонны*.
ОБОСОБЛЕННОСТЬ КОЛОНН
Чтобы сформировались колонны, необходимо несколько определенных условий. Во-первых, должна быть определенная численность колонны. Например, семья рыжих лесных муравьев не может быть слишком малой численности, так как в этом случае она оказывается нежизнеспособной. Минимум численности жизнеспособной семьи где-то около 100 тыс. Гнездо семьи из 100 тыс. муравьев имеет диаметр около 60 см и высоту 40-45 см. Такой муравейник еще слишком мал, чтобы иметь колонны (они появляются позже при численности семей 180-200 тыс. особей). Число колонн в муравейнике невелико: две, реже три (60-100 тыс. муравьев в одной колонне).
Второе необходимое условие - достаточно выраженная разобщенность колонн, прежде всего пространственная. Без этого колонны существовать не могут. Разобщение необходимо и в гнезде, и на кормовом участке. Это установлено и проверено на разных видах муравьев. В основе обособленности колонн лежит характерный для муравьев принцип вторичного деления территории*, выявленный в 1956 г. польским мирмекологом Я.Добжанской. Каждый охотник действует на элементарном поисковом участке, с которым он связан длительное время. Если он и меняет участок, то это происходит только в пределах одного из секторов охраняемой территории семьи, точнее - в пределах одной дороги. Этим обусловлено постоянство сочетаний рабочих муравьев на кормовых дорогах, по которым муравьи попадают к своим поисковым участкам и колониям тлей. Сами дороги также отличаются большим постоянством, оставаясь практически неизменными 10-13 лет. Эти цифры означают лишь максимальные сроки регулярных наблюдений за одним и тем же гнездом.
Каждая дорога берет свое начало в определенной точке гнезда. Начальные точки дорог рассредоточены обычно довольно равномерно по периметру гнездового купола. Логично предположить, что муравьи, действующие на различных дорогах, придерживаются и в гнезде секторов*, прилегающих к этим дорогам. Экспериментальная проверка на гнезде обыкновенного лесного муравья, произведенная нами осенью 1969 г. и летом 1970 г. в Приокско-террасном заповеднике, подтвердила такое предположение.
Фуражиры на дорогах подопытного гнезда были помечены групповыми метками. Ночью, когда все фуражиры собрались в куполе, семью усыпили эфиром. Далее был послойно разобран купол и зафиксировано местонахождение каждого меченого муравья. Оказалось, что рабочие, связанные с определенной дорогой, сосредоточиваются в прилегающей к этой дороге части гнезда. Единственным местом, где рабочие из разных колонн свободно смешиваются, оказался внутренний конус муравейника. Муравьи-строители также придерживаются своих секторов на поверхности купола. Привязанность муравьев к своим секторам гнезда оказалась очень прочной. В ходе разборки муравейника весь строительный материал купола и оказавшиеся в нем муравьи были полностью перемешаны. Когда разборка гнезда и регистрация муравьев были закончены, этот материай вместе с начавшими просыпаться муравьями собрали на прежнее место и придали ему форму, приблизительно соответствующую доопытной. И в этих условиях фуражиры и строители нашли свои секторы муравейника.
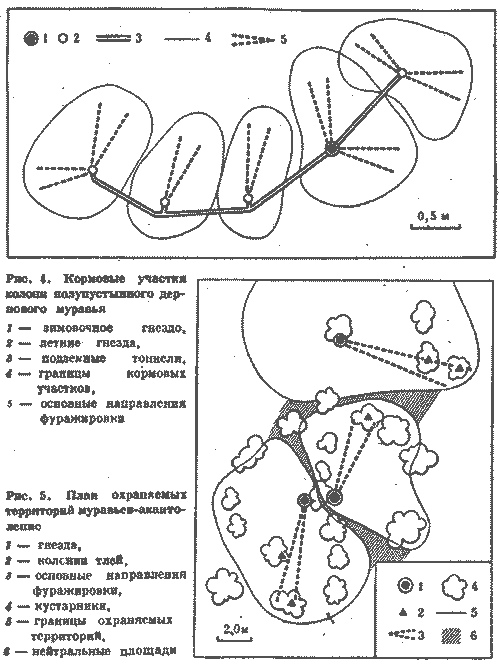
Еще нагляднее связь колонн с определенной частью гнезда у видов, сооружающих секционные гнезда, о которых говорилось выше. Там действует четкий принцип: каждой секции гнезда соответствует колонна. Пространственная разобщенность колонн сохраняется и на территории. Это вытекает уже из постоянства на территории кормовых дорог муравейника или кратеров - выходов из секций на поверхность (рис. 4, 5). Каждая колонна имеет самостоятельный выход или группу выходов на кормовой участок. На территории характер отношений колонн проявляется прежде всего во взаимодействии потоков фуражиров разных дорог.
С этой целью в июне 1974 г. был поставлен специальный опыт на средней по размерам семье обыкновенного лесного муравья (диаметр купола - 0,9 м, высота гнезда - 0,6 м). Семью составляли четыре колонны примерно одинаковой мощности, о чем можно судить по сопоставимым размерам дорог (17-20 м) и интенсивности движения на них (рис. 6). После мочения на всех дорогах 2 тыс. фуражиров в зоне дороги II была установлена непреодолимая для муравьев металлическая стенка, вынуждающая поток с дороги II выйти на соседнюю дорогу I. Протяженность стенки в противоположную от дороги I сторону, к югу, превышала 15 м. Через несколько часов поток II обошел стенку с севера и вышел на полотно дороги I.
Можно было бы предполагать, что произойдет слияние двух потоков муравьев. Однако это не случилось. Войдя в вынужденное соприкосновение с потоком II, поток I отодвинулся от него к северу на 20 см. Миновав препятствие, потоки сразу же расходились, попадая в собственные русла. Такое положение сохранялось 12 дней, после чего стенка еще трижды наращивалась, пересекая каждый раз дорогу I. При этом происходили сопряженные перемещения обоих потоков к северу с сохранением установившегося между ними расстояния в 20 см. Опыт длился месяц, после чего стенка была убрана, и уже на следующий день оба потока вернулись в исходное положение.
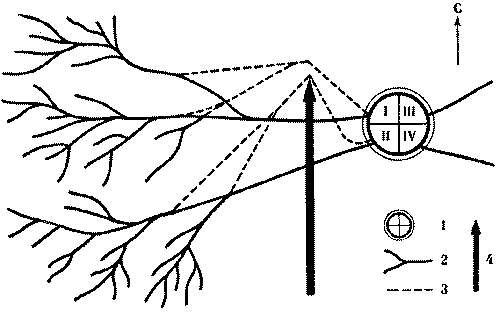
Рис. 6. Потоки фуражиров рыжего лесного муравья разных колонн «отталкиваются» при перемещении дорог.
I, II, III, IV - номера секторов и дорог гнезда; 1 - муравейник, 2 - положение дорог до и после опыта, 3 - положение дорог при максимальном смещении во время опыта, 4 - стенка
Этот опыт показал, что существует упругость дорожной сети муравейника, как бы натянутой между постоянными точками гнезда и кормового участка. Потоки фуражиров не смешиваются, они отталкиваются друг от друга. В этом выражается обособленность и целостность колонн и даже противостояние их на участке.
Обособленность колонн на территории сохраняется у рыжих лесных муравьев и в других частях кормового участка* семьи, где контактируют уже не потоки, а группы муравьев. Территория каждой колонны охраняется и отделена от охраняемых территорий* других колонн нейтральными пространствами* - зонами, не посещаемыми муравьями (так же, как отделены охраняемые территории самостоятельных муравейников).
Кормовые участки колонн стабильны и неприкосновенны и у видов, не имеющих дорог. У обитающих в пустыне Каракум зерноядных пустынных дерновых муравьев к каждой секции примыкает свой обособленный кормовой участок. В ряде случаев он отделен от других нейтральным пространством, как и у всех зерноядных муравьев. Сбор семян производится не ежедневно, а по мере их созревания. А так как каждая колонна собирает семена самостоятельно, то обычна ситуация, когда на поверхности работают фуражиры лишь части колонн, а выходы из остальных секций закрыты. Кормовые участки закрытых секций остаются в этот день не использованными ни фуражирами из других секций данного гнезда, ни другими семьями (рис. 7).
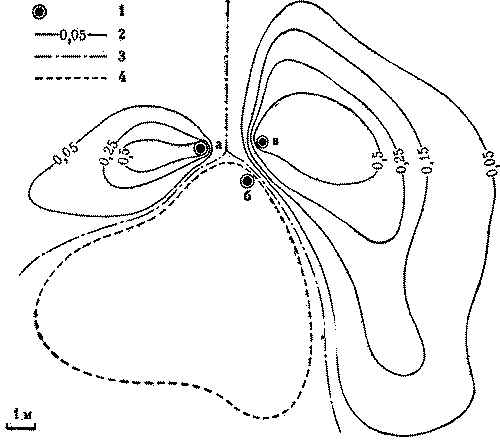
Рис.7. Использование кормового участка трехсекционного гнезда дернового пустынного муравья при работе двух секций 1 - кратеры секций а, б а в, г - изолинии динамической плотности особей на участках работающих секций а и б (особь/дм2мин), 3- границы кормовых участков секций, 4 - границы охраняемого участка неработающей секции б
Понятно, что только пространственной обособленности колонн в гнезде и на участке еще недостаточно для существования в муравейнике группировок особей. Пространственная обособленность должна сочетаться с такими важными характеристиками, как достаточно глубокая информативная изоляция и функциональная автономность колонн друг от друга. Естественно, что ни одно из перечисленных проявлений автономности колонн в рамках семьи не может быть полным, так как в этом случае семья просто перестала бы существовать как единое целое.
Информация может передаваться из колонны в колонну различными путями: посредством обмена пищей, различных сигналов (акустических, киноптических, химических, тактильных)*; обмена особями (личинками, куколками, взрослыми муравьями); распространения в гнезде специфического запаха, в результате чего возникает и поддерживается особый запах муравейника - гнездовой запах*.
Передача различных сигналов носят прежде всего оперативный характер, в то время как обмен пищей и особями дает информацию, накапливающуюся в семье и приводящую к изменению поведения со значительной задержкой. Специфический запах семьи служит как бы эталоном при опознании своих (т. е. из своего гнезда) и чужих муравьев в гнезде и на участке.
Изучая с помощью изотопов пути распространения пищи в семьях рыжих лесных муравьев, Р.Шовен установил, что пища, добываемая муравьями на одной из дорог, не передается в другие секторы гнезда, т. е., как только в семье возникают колонны, единый пищевой поток, охватывавший ранее всех членов семьи, распадается на несколько изолированных друг от друга потоков, каждый из которых ограничен пределами одной колонны. Такая пищевая, или трофическая, изоляция колонн имеет очень важные для дальнейшего развития муравейника последствия.
Трофическая изоляция колонн и их пространственная обособленность усиливаются почти полным отсутствием передачи сведений из колонны в колонну о том, что происходит на кормовом участке. Это было проведено рядом опытов. Один из них заключался в следующем. У одиночного муравейника с помощью ловушек в течение дня отлавливали всех фуражиров с одной из дорог, интенсивность движения на подопытной дороге при этом непрерывно снижалась и через несколько часов упала до нуля в результате полной выборки всех внегнездовых рабочих из данной колонны. Казалось бы, для муравейника отлов тысяч фуражиров равнозначен гибели массы муравьев на участке одной из колонн, что означает серьезную угрозу для всей семьи.
Однако бедственное положение на одной из дорог не отразилось на скорости и интенсивности движения и поведении муравьев на остальных дорогах. Там фуражиры работали в своем обычном ритме.
Такая же картина наблюдалась и на поверхности муравейника, в контрольных секторах купола. При изъятии всех фуражиров с одной из дорог у нескольких муравейников, отличающихся по размерам и числу дорог, заметная реакция в остальных колоннах отсутствовала. Все происходящее на кормовом участке одной колонны оставалось ее частным делом и совершенно не интересовало соседей, обитающих в том же гнезде и входящих в состав одной семьи.
Абсолютен ли этот тип изоляции? Конечно, нет. Но вместе с тем настолько глубок, что мы можем проделать и более бесцеремонные действия, не вызвав тревоги у соседей. Например, надев резиновые перчатки, отлавливаем с поверхности одного из секторов купола всех находящихся там муравьев. Здесь, правда, появляется опасность, что муравьи из соседних секторов заметят резкое движение руки и устремятся навстречу надвигающейся угрозе. Однако достаточно нехитрого приспособления, чтобы этого избежать. В нижней части купола и на гнездовом валу, по границам подопытного сектора, устанавливаем невысокие стенки (5-6 см высотой). Отлов муравьев производим только между стенками. Непосредственной реакции муравьев-наблюдателей из соседних колонн на пути сборщика и боевые позы защитников гнезда из подопытной колонны уже нет. Теперь, если не делать слишком резких движений, можно отловить всех внегнездовых муравьев одной из колонн без опасений, что будет захвачено сколько-нибудь значительное число особей из соседних колонн. Муравьи подопытной колонны реагируют на исчезновение собратьев и как бы дают сигнал, когда такая выборка закончена: они быстро закрывают все гнездовые выходы. Сектор как бы вымирает. А рядом в соседних секторах все идет своим чередом: бдительные наблюдатели патрулируют поверхность купола, строители укладывают в покровный слой гнезда очередные хвоинки, непрерывным потоком идут в гнездо и вновь выходят из него потоки фуражиров. Судьба забаррикадировавшейся в своем секторе колонны никого из них не беспокоит.
Из всего изложенного можно сделать вывод, что колонны, обитающие в одном гнезде, а у многих видов имеющие даже общую крышу над головой, действуют фактически обособленно друг от друга. Но очевидно, что такое положение попросту нереально, ибо тогда непонятно, почему они все-таки обитают в одном гнезде и что их там удерживает.
Вопрос об автономности и обособленности колонн не возник бы, если бы ее состав не позволял выполнять в пределах одной колонны все основные функции, которые свойственны одинарной семье (т. е. семье, не подразделенной на колонны). В каждой колонне имеются все характерные для вида функциональные группы муравьев (и внутригнездовых и внегнездовьгх), выращивается молодь. Полное укомплектование колонны муравьями разных профессий и позволяет ей обособиться.
ФАКТОРЫ СОХРАНЕНИЯ ЦЕЛОСТНОСТИ СЕМЬИ
Оказывается, что в ряде случаев четкая изоляция колонн сочетается с постоянными контактами и взаимодействием в других сферах. Естественно, что эти контакты строго регламентированы. Большинство контактов имеет чисто «профессиональный» характер: строители общаются со строителями, внутригнездовые рабочие - с внутригнездовыми, фуражиры - с фуражирами. Зоны регулярного соприкосновения и взаимодействия муравьев из разных профессиональных групп пространственно разделены.
У рыжих лесных муравьев внутригнездовые рабочие контактируют во внутреннем конусе гнезда, где производится важнейшая для поддержания целостности семьи операция - регулярный обмен личинками, куколками и молодыми рабочими. Это удивительное явление чем-то напоминает меновую торговлю, но объектами обмена служат сами муравьи. По-видимому, это единственное, чем могли бы меняться колонны, ведь обмена пищей между ними нет (рис. 8).
Происходит это так. Во внутреннем конусе*, являющемся настоящей инкубационной камерой муравейника (в нем поддерживается постоянная температура), обычно сосредоточен весь расплод гнезда. Каждая колонна держит молодь в своем секторе внутреннего конуса, который, однако, невелик, и расплод разных колонн оказывается рядом. И муравьи-носильщики, беспрепятственно входя в чужой сектор, забирают то личинку, то куколку и уносят их к себе. Так поступают и носильщики остальных колонн. В результате внутри муравейника идет весьма интенсивный обмен особями. Энтомолог из ФРГ Г.Кнайтп подсчитал, что в разгар сезона обмен производится каждые две минуты. Это величина достаточно большая, хотя, если учесть, что в крупном гнезде рыжих лесных муравьев одновременно может воспитываться несколько сот тысяч личинок, обмениваемая часть состава колонн относительно невелика.
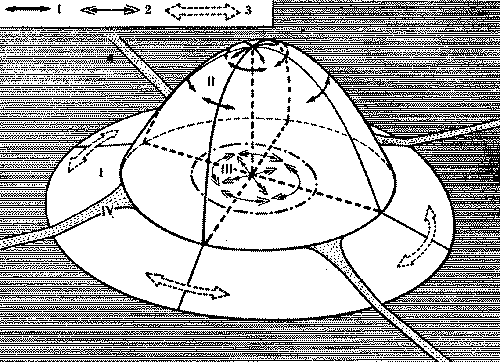
Рис. 8. Общая схема взаимодействия колонн в гнезде рыжих лесных муравьев, состоящем из четырех секций. I - гнездовой вал, II - поверхность купола. III - внутренний конус, IV - кормовая дорога. 1 - контакты муравьев-наблюдателей и строителей на поверхности купола, 2 - обмены молодью во внутреннем конусе, 3 - контакты фуражиров на гнездовом валу.
Отсутствие в гнездах многих видов внутреннего конуса не исключает внутригнездовых обменов. В секционных гнездах для этого служит система поверхностных тоннелей и камер, связывающая все входящие в состав гнезда секции. Вообще носильщики из других колонн проникают не только в «общие» помещения гнезда, но и дальше, собирая часть молоди, даже если она содержится в покровном слое или в верхних камерах секций.
Что же дают обмены особями? Они препятствуют отчуждению колонн, позволяют сохранить между ними тесные родственные отношения, исключают вражду. При обмене особями происходит скрытый обмен пищей. Все это приводит к некоторому выравниванию состава колонн как по происхождению, так и но состоянию. Сам факт устойчивого во времени существования муравейников, состоящих из нескольких колонн, говорит о том, что выработанный муравьями уровень обменов достаточен для сохранения целостности семьи.
Однако взаимодействие колонн внутри гнезда не сводится к одним обменам. Гнездо или хотя бы его общие части (внутренний конус, система поверхностных ходов и камер и т. п.) строятся муравьями из разных колонн сообща. При этом основная задача контактов сводится к обеспечению сопряжения отдельных элементов гнезда. Подобную роль играют постоянные контакты и на поверхности купола. Здесь на стыке соседних секторов взаимодействуют муравьи-наблюдатели и строители гнезда, а также фуражиры, доставляющие в муравейник материал для его сооружения. Их контакты обязательны, ибо без них муравьи не в состоянии возвести купол правильных очертаний. Это проверено экспериментально Р. Шовеном.
Ограничив с помощью нескольких стенок контакты между отдельными частями гнезда, ученый нарушил равномерность контактов строителей на поверхности купола. В результате различные части купола начинали расти неравномерно и купол утратил присущие ему округло-конические очертания. Такая картина наблюдалась даже в тех случаях, когда в каждый ограниченный стенками квадрант гнезда поступало одинаковое количество строительного материала.
Муравейник как бы накрыт контактной сетью с ячейками определенного размера. Ячейки меньше на маковке гнездового купола - здесь муравьев обычно больше и контактируют они чаще, поэтому центральная часть растет быстрее. Ближе к земле ячейки увеличиваются, скорость роста замедляется. В старых, приходящих в упадок гнездах рыжих лесных муравьев число строителей уже недостаточно для обеспечения необходимой частоты контактов. И это сразу же сказывается на форме купола. Он становится неровным, бугристым. Функция частоты контактов здесь гипертрофирована. Но, с другой стороны, представление об этакой живой форме для растущего муравейника, пожалуй, в наибольшей мере соответствует возможностям муравьев создавать впечатляющие по размерам и совершенству архитектоники целостные сооружения. Ведь общего плана муравейника нет ни у кого из его строителей.
В свое время, изучая процесс строительства галерей у термитов, известный французский энтймолог Р.Грассе выдвинул теорию стигметрии, объясняющую стереотипность элементов термитника различиями в действиях строителей на разной высоте oт пола. Такой подход поясняет и связь между размерами особей и размерами ходов и камер. Но это касается технологии сооружения отдельных элементов гнезда, соизмеримых с самими строителями. Муравейник же превосходит муравья в тысячи раз. Вот и остается муравьям разве что «взяться за руки», чтобы выстроить купол правильной формы.
Одновременно, по-видимому, решается и другая очень важная задача - соответствие размеров гнезда численности семьи. В слишком маленьком гнезде тесно, содержать очень большое - оказывается не под силу. Поэтому размеры гнезда, как правило, оптимальны для конкретной семьи. В природе задача оптимизации гнезда решается радикально: либо надстраивается купол, либо сооружаются новые секции. Уменьшение семьи влечет за собой уменьшение гнезда. Если муравьев поместить в гипсовый садок неподходящих размеров, то они либо концентрируются в части камер, если садок велик, либо «лишние» особи уходят из садка.
У самых различных видов муравьев, объединяемых лишь одним принципом общего вторичного деления территории*, фуражиры составляют определенную часть семьи, в среднем около 13%. Также постоянен и процент особей, занятых уходом за самками и расплодом (25-30%). На долю строителей и муравьев-наблюдателей на поверхности купола у рыжих и тонкоголовых муравьев приходится 10-12%. На постоянстве последней величины и основывается связь между численностью семьи и размерами гнезда. Эта зависимость обнаружена и у других видов, например у строящего земляные кочки желтого земляного муравья. Даже в тех случаях, когда муравьи строят лишь небольшие временные купола для согревания личинок, сохраняется четкая связь между численностью семьи и размерами гнезда.
Имеется зона регулярных контактов и потоков фуражиров. Это гнездовой вал. Частота контактов невысока и сводится к периодическому обмену тактильными сигналами*, а также к постоянному наблюдению за ситуацией в ближайшей части соседнего сектора гнездового вала. Сильное возбуждение муравьев в пограничной зоне одного из секторов привлекает туда часть фуражиров из соседней колонны. При этом отдельные фуражиры могут увлечься и попасть в чужой поток фуражиров. Такие переходы возможны только на гнездовом валу или в непосредственной близости от него. На пространственной обособленности колонн на кормовом участке все это не сказывается: потоки продолжают отталкиваться друг от друга, а сам участок остается поделенным между колоннами. Основанное на общей бдительности взаимодействие фуражиров помогает муравьям заблаговременно выявить угрозу гнезду. Надо отметить, что это всегда реакция отдельных особей или небольших групп муравьев. Чтобы активизировалась колонна в целом, источники возбуждения должны действовать непосредственно в ее гнездовом секторе. Действия колонн при этом не согласуются. Единственным местом на поверхности гнезда, воздействие на которое может вызвать общую ответную реакцию семьи, является маковка купола.
Следует отметить, что схемы взаимодействия муравьев перечисленных выше функциональных групп различны. Во внутреннем конусе каждая колонна обменивается молодью непосредственно со всеми остальными колоннами. Чтобы установить это, мы провели следующий опыт. Куколок с разноцветными метками выложили в несколько приемов на разные кормовые дороги гнезда.
Муравьи тут же начали заносить куколок в гнездо. Почти одновременно меченые куколки начали поступать в соседнее родственное гнездо, куколки с каждой новой меткой составляли определенную часть от общего числа выложенных на соответствующей дороге.
Обмен по принципу «каждая с каждой» отражает и поддерживает определенный тип взаимодействия колонн в семье муравьев. Здесь действуют многосторонние связи без какого-либо предпочтения кому-либо из партнеров.
Возможность многостороннего контакта имеется и у особей, занятых на поверхности купола. Правда, не у всех, а лишь у тех муравьев, которые функционируют на самой маковке, где смыкаются границы всех секторов. Здесь обычно максимальное число наблюдателей, так как это самое ответственное место муравейника, маковку, восстанавливают в первую очередь. Эта часть гнезда оформляется сразу же, даже если место, над которым оно должно находиться, оказалось в результате разрушения на уровне почвы.
Возможно, в какой-то мере маковка воплощает целостность муравейника.
Действительно, разделить верхнюю часть купола на две, сделать его двуглавым - значит положить начало глубокому и долгому процессу перестройки всей организации семьи. Нередко это приводит даже к разделению муравейника на два.
На остальной поверхности купола колонны контактируют и обмениваются сигнальной информацией только по эстафете. Преднамеренной передачи информации из колонны в колонну нет; муравьи воспринимают как сигналы тревоги специфические позы других особей в минуты опасности. Не имеет значения, принял ли такую позу муравей из своей или соседней колонны. Состояние возбуждения поддерживается специальными веществами-феромонами тревоги, или торибонами, выделяемыми муравьями при появлении угрозы.
Эстафетные контакты фуражиров разных колонн на гнездовом валу связаны с ролью взаимодействия особей данной функциональной группы в жизни семьи. В результате такого взаимодействия устанавливаются радиальные структуры кормового участка, а для сохранения целостности семьи контакты фуражиров значения не имеют.
Вообще гнездовой вал играет у многих видов муравьев особую роль. Здесь организационная структура семьи трансформируется в пространственную структуру кормового участка и начинается раздел сфер влияния колонн вне гнезда: определяются направления кормовых дорог или потоков фуражиров в тех случаях, когда постоянных дорог нет. Дальнейшее размежевание колонн на территории осуществляется при взаимодействии отталкивающихся друг от друга потоков фуражиров. В остальных частях кормового участка границы поддерживаются группами фуражиров, численность которых в пограничных зонах обычно выше, нежели в среднем по участку.
В итоге складывается достаточно сложная и вместе с тем рациональная система постоянного взаимодействия колонн в гнезде и на территории, схематично изображенная на рис. 8. И хотя колонны обособлены и во многом развиваются почти независимо друг от друга, они все-таки составляют единое целое - семью.
Подытоживая изложенное в этом разделе, следует назвать два фактора, наиболее важных для сохранения семьи муравьев: общее гнездо и регулярные обмены особями Гнездо важно не только как жилище, общая крыша. В таком гнезде присутствует и постоянно выравнивается единый запах семьи. Ведь именно по запаху муравьи отличают своих от чужих. Пока муравейники имеют одинаковые или близкие запахи, воины между ними не будет. Встретившись на куполе или вдали от него рабочие муравьи первым делом обмениваются легкими ударами антенн. В булавах антенн находятся хеморецепторы, с помощью которых муравей и определяет запах различных предметов, в том числе и запах муравьев. Свой будет отпущен или привлечен к какой-нибудь деятельности. Чужому не поздоровится; его либо разорвут на части, либо изгонят.
Обмены как средство сохранения лояльных отношений колонн дополняют общность гнезда и общность гнездового запаха.
Однако обмены оказываются крайне необходимыми при выполнении еще одного определяющего условия устойчивости семьи: сравнимости колонн по численности.
СРАВНИМОСТЬ КОЛОНН ПО ЧИСЛЕННОСТИ НАСЕЛЕНИЯ
Сколько-нибудь выраженной иерархии колонн в пределах семьи нет. В секционных гнездах это обусловлено тем, что все секции единого гнезда равны между собой. Одинаковы по численности и колонны, населяющие секции. Это показано на разных видах, относящихся к различным родам и подсемействам.
Численность населения одной секции стабильна для каждого вида в конкретных условиях обитания. А поскольку отношения группировок муравьев одного вида отражают в конечном счете соотношение численности этих группировок, то основа для доминирования какой-либо из колонн в семьях, обитающих в секционных гнездах, отсутствует.
У стабилизированной семьи рыжих лесных муравьев все обстоит сложнее, так как с сооружением купола численность муравьев перестает зависеть от емкости подземной части гнезда. Купол можно надстроить, увеличив его диаметр и высоту. Увеличился объем купола - возросли возможности гнезда вместить дополнительное количество особей. Поэтому какой-то определенной численности колонны здесь нет.
В гнезде одного размера может быть разное число колонн. Так, у северного лесного муравья в гнезде с диаметром купола 1 м оказывается и две, и пять колонн. Естественно, что в первом случае численность каждой колонны выше. Но внутри семьи колонны, как правило, соизмеримы. И чем старше муравейник, чем меньше вокруг других гнезд, так или иначе влияющих на его рост, тем ровнее длина дорог, интенсивность движения на них и общая численность колонн. В идеальной ситуации, когда одиночный муравейник расположен в равномерном по составу, возрасту и продуктивности насаждении, кормовой участок приобретает форму круга.
Такие семьи обладают наибольшей устойчивостью во времени, и только серьезное вмешательство извне может вывести их из стабилизированного состояния. В этом случае чем больше колонн в муравейнике, тем меньше вероятность последующих серьезных изменений в структуре семьи. И все же такие изменения происходят. Гнездо растет, и у него появляются новые дороги. На некотором расстоянии от старого муравейника возникают новые муравейники, которые тоже начинают расти. Наступает время, и здесь образуются колонны.
Что же приводит к этим изменениям? Ведь очень часто новые колонны возникают задолго до того момента, когда уже имеющиеся достигли верхнего предела численности. Такой предел еще не известен, однако в совершенно одинаковых условиях по соседству существуют муравейники с разной численностью колонн. Например, у каждого по три колонны, но колонны первого гнезда насчитывают по 50-60 тыс. особей, второго - 90- 100 тыс., а третьего -150-200 тыс. особей. Можно было бы ожидать, что новая колонна возникнет прежде всего в последнем, наиболее мощном муравейнике. Однако в действительности очень часто дело обстоит иначе. Следовательно, абсолютная численность еще не определяет ситуации.
Чтобы выяснить, в чем причина, был проведен следующий опыт. В 1969 г. мы подобрали шесть близких по размерам муравейников обыкновенного лесного муравья, имевших по три равных между собой дороги. Территория каждого подопытного муравейника была тщательно закартирована до начала опыта. Все шесть муравейников были одиночными. Четырем из них на одну из дорог в течение июля-августа выкладывали в больших количествах куколок того же вида, взятых из гнезд, не участвовавших в опыте. В несколько приемов в каждом случае на подопытную дорогу было выложено по 100 тыс. куколок.
Вышедшие из куколок рабочие муравьи влились в состав соответствующей колонны, которая таким образом резко усилилась. Два других муравейника в это же время также были усилены куколками, но одновременно все колонны усиливались на 90-100 тыс. особей каждая.
Реакция муравейников в первом и втором вариантах оказалась различной. Равномерное усиление всех колонн привело к росту гнезд и также равномерному увеличению дорожной сети каждой колонны. Усиление одной из колонн нарушило имевшееся до опыта равновесие. Кормовой участок такой колонны резко расширился, дорога удлинилась и появились новые ответвления. Контрольные дороги не изменились. Наконец, одна из усиленных колонн разделилась и выделила дочернюю семью*. Отводок* был сооружен в сентябре того же года, через месяц после усиления колонны куколками.
Из приведенного опыта следует несколько выводов. Обмены, которые производят муравьи внутри гнезда, не обеспечивают выравнивания численности колонн в случае резкого усиления одной из них. Если одна из колонн вследствие каких-либо причин начинает расти быстрее остальных, различия в численности накапливаются. В определенный момент подобная несоразмерность может послужить причиной выделения отводка наиболее сильной колонной.
Интересно, что в различных экспериментах и при естественном ходе событий дочернюю семью всегда выделяет самая мощная колонна, резко выделяющаяся по численности особей. Есть все основания расценивать образование отводка как средство сохранения структуры семьи: проблема внутрисемейной регуляции решается путем расселения и создания новой семьи.
Образование отводка - один из путей выравнивания колонн по численности населения, наиболее радикальный, но не единственный. Второй, более обычный путь - формирование новой колонны. Здесь тоже обязательным условием является нарушение численного равновесия колонн.
Образование новой колонны теснее связано со структурой кормового участка, с дорожной сетью муравейника. Возникновение колонны у рыжих лесных муравьев невозможно без появления новой дороги. Число разветвлений у той или иной дороги роли не играет. Важно не число ответвлений на территории, а число потоков, выходящих из гнездового купола. Поэтому процесс разделения колонны начинается всегда на кормовом участке, схематично он изображен на рис. 9. Из схемы видно, что разделение в большей степени зависит от возможности радикального раздвоения исходного кормового участка колонны. Чем шире занимаемый участком сектор, тем проще использующей его колонне разделиться на две. Число колонн и связанных с ними дорог у рыжих лесных муравьев весьма постоянно. Еще в 30-х годах появились работы немецкого энтомолога Р.Штегера, проследившего топографическое постоянство кормовых дорог в течение 13 лет. Имеются подобные данные и у других авторов, работавших с муравьями разных родов. Смещения отдельных ответвлений дорог обычно не связаны с изменением в составе колонн. А иногда разделение колонны на две почти не отражается на периферийных отрезках дорог (именно этот вариант показан на рис. 9, в).
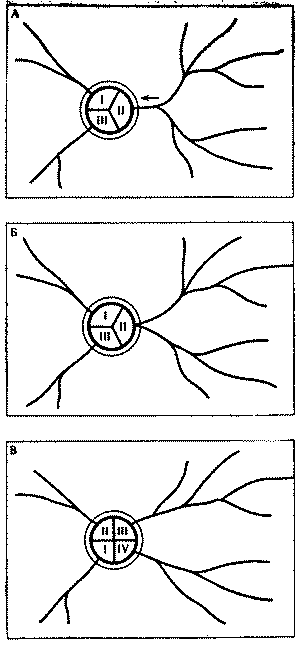
Рис. 9. Процесс образования новой колонны при раздвоении дороги
А - наметившееся разделение наиболее сильной и разветвленной дороги (стрелка
показывает направление перемещения развилки);
Б - положение накануне
разделения колонны на две: развилка переместилась к основанию купола;
В -
колонна разделилась, ответвления стали самостоятельными колоннами
I, II,
III, IV - номера колонн (дорог, секторов)
Увеличение колонн - процесс, направленный как бы внутрь муравейника, ведущий к умножению связей в семье, укрепляющий ее как единое образование. А это означает, что данный процесс в какой-то мере противодействует почкованию* - выделению дочерних муравейников. Действительно, чем больше колонн, тем прочнее каждая из них связана с семьей, тем труднее одной из них достичь условий, необходимых для выделения отводков.
Наибольшее значение имеют при этом два показателя: размер семьи, тесно связанный с размерами муравейника и кормового участка, и число колонн. В результате взаимодействия двух тенденций вырабатывается оптимальная зона образования отводков. Причем эта зона даже не видоспецифична вообще, а отражает специфику вида лишь в определенных условиях. Так, в подмосковном ельнике у обыкновенного лесного муравья максимум образованных отводков падает на муравейники с диаметром 1,2-1,3 м, состоящие из четырех колонн. А в сосняках у того же вида диаметр наиболее плодовитого гнезда-1-1,1 м при трех колоннах.
Процессы разделения колонн и почкования противодействуют друг другу и имеют разные последствия для роста гнезда. Увеличение числа колонн - это путь к развитию крупных муравейников, появлению «сверхгнезд». Муравейники-гиганты почти всегда одиночные и всегда включают много колонн. Максимальное число колонн в средней полосе отмечено нами в Приокско-террасном заповеднике; семья малого лесного муравья имела 12 дорог. Охраняемая ею территория занимала более 1,5 га. Почкование, наоборот, сдерживает рост муравейника. Причем сдерживание происходит за счет самой мощной, т. е. наиболее быстро растущей колонны, что особенно важно. В итоге семья удерживается в пределах, соответствующих состоянию интенсивного выделения отводков.
Попробуем охарактеризовать этот размерный интервал с позиции оптимизации размеров семьи. Представим себе требования, которым должен соответствовать такой муравейник.
В общих чертах оптимальной организацией любого биологического объекта можно считать такую организацию, которая обеспечивает устойчивую плотность поселения на максимальном для данных условий обитания уровне. Это почти в равной мере применимо к различным уровням организации. Для муравьев мы вправе использовать такой подход по отношению к отдельной семье, комплексу муравейников* одного вида и даже к обитающим на одной территории поселениям муравьев разных видов, или, как их назвал в 1975 г. московский мирмеколог А.В.Демченко, к многовидовым ассоциациям муравейников*.
Для отдельного муравейника основным показателем оптимальности организации будет стабильность размеров семьи и гнезда во времени. Важны также вероятность гибели муравейника определенного размера, его возможности к дальнейшему росту и образованию отводков. Необходимо выбрать отрезок времени, соответствующий нашей задаче. Год для этой цели слишком малый срок: условия разных лет могут резко отличаться друг от друга, а это неизбежно сказывается на состоянии каждой семьи. Но пятилетнего периода уже достаточно, чтобы выявить общие тенденции развития муравейников. Вот что дал анализ пятилетних данных по 112 семьям северного лесного муравья, обитающего в ельнике-кисличнике в районе Солнечногорска Московской области.
Совсем маленькие муравейники (диаметр до 40 см), как правило, обречены. Если в течение двух лет такой муравейник не вырастет в более крупный, он погибнет. Погибает 83% подобных гнезд. Муравейники этого размерного класса не обладают устойчивостью. Среди муравейников покрупнее (диаметр 45-60 см) доля погибших меньше (24% за пять лет), но и здесь семья всегда перед альтернативой: либо вырасти, либо погибнуть. Лишь 4% семей сохраняется в исходном классе в течение пяти лет. Зато большинство муравейников (72%) вырастают, причем некоторые за это время достигают метра в диаметре.
Далее вероятность погибнуть уменьшается, но скорость роста начинает снижаться. Одновременно появляется и обратная тенденция: муравейники, продержавшись некоторое время в более высоком классе, вновь попадают в разряд более мелких. Это особенно заметно на примере крупных гнезд диаметром 1,25 м и более. Тенденции к росту резко падают с увеличением контрольного периода с одного до пяти лет. Такой муравейник может выйти в следующий класс на один-три года, но в течение пяти лет ни один из них не удерживается там и снова уменьшается в размерах, нередко даже ниже исходного уровня. Среди гнезд диаметром более 1,4 м в своем размерном классе удерживаются только из муравейников, а остальные - начинают деградировать.
Единственной устойчивой размерной группой оказались муравейники с диаметром купола 1,05-1,2 м. Большинство муравейников этого размера сохраняются в классе независимо от срока наблюдения. У них еще сильна тенденция к росту: почти 1/3 гнезд за пятилетие переходит в следующий размерный класс. Это значит, что муравейники такого размера, обретая устойчивость, сохраняют активность. Вероятность гибели муравейников данного класса за пятилетний период равна нулю. И лишь 7% гнезд уменьшается в размерах.
Но, пожалуй, самое интересное заключается в том, что муравейники именно этого диаметра образуют основную часть естественных отводков в комплексе. Следовательно, есть все основания рассматривать образование отводков как механизм поддержания численности семьи на определенном, оптимальном для данного вида в данных условиях уровне.
Что же это за уровень? Расчеты показали, что в гнезде северного лесного муравья диаметром 112,5 см (середина класса) обитает около 870 тыс. муравьев. Эти муравьи организованы, как правило, в четыре колонны (среднее число колонн - 4,11). -
Знакомясь с двумя механизмами стабилизации муравейников, мы не должны упускать из виду одну важную общую черту. Действие этих механизмов непостоянно. Они срабатывают периодически, как ответная реакция семьи на накопившиеся количественные диспропорции между составляющими ее колоннами. И образование отводка, и формирование новой колонны - специфические аварийные меры сохранения семьи. И хотя фактологическая сторона процессов достаточно прояснилась, один из наиболее интригующих, узловых моментов остается неясным: какие реальные силы заставляют самую могучую колонну в семье выделять отводок? Почему именно так решается этот важный для муравейника вопрос? Ведь заглядывая немного вперед, можно сказать, что образование отводков далеко не всегда выгодно материнскому муравейнику*. Казалось бы, усилившаяся колонна могла бы потеснить соседей по гнезду или просто расширить свой сектор.
К сожалению, ответов на эти вопросы еще нет. Очевидно только, что существенного передела секторов гнезда в обычных условиях не происходит, так же никогда не наблюдаются внутригнездовые муравьиные войны.
В природе мы можем застать самые различные этапы оптимизации муравейника. Этот процесс складывается из двух процессов: оптимизации размеров семьи и оптимизации ее организации. Они взаимосвязаны, дополняют друг друга, но не совпадают. Синхронности здесь тоже нет, потому что первый процесс почти непрерывен, а второй - четко дискретен. Поэтому первый процесс обычно идет быстрее. Численность колонн «догоняет» его скачками (и даже может опередить).
При этом увеличение числа колонн связано с формированием наиболее рациональной формы кормового участка - округлой.
Формированию кругового кормового участка могут препятствовать различные факторы. Такой участок маловероятен при неравноценности кормовых угодий вокруг муравейника. Распределение растений с колониями насекомых, выделяющих падь, также имеет значение. Особенно сильно сказывается размещение соседних муравейников. Конечно, не всяких, а того же или конкурирующих видов. Установив в 1932 г. наличие у муравьев охраняемой территории, английский эколог Ч.Элтон положил начало целому направлению в изучении жизни муравьев - первых насекомых, у которых данный вопрос был разносторонне изучен.
Конкуренция сводится к борьбе за территорию, реже - за места поселения. Кормовые участки расположенных поблизости муравейников вытянуты в противоположные стороны либо параллельными полосами. Нередко форма их причудлива: кормовые дороги с прилегающими полосками угодий как бы пробираются среди чужих земель. В таких условиях выравнять колонны путем увеличения их числа нельзя. И тогда выручают отводки. Вместе же оба процесса дают большинству семей достаточно богатые возможности для достижения устойчивого состояния.
КОЛОННА-МУРАВЕЙНИК В МУРАВЕЙНИКЕ
Достигая определенных размеров, семья муравьев начинает дробиться на части, каждая из которых вполне автономна и в состоянии полностью или почти полностью выполнять все необходимые для полноценного существования функции. Под одной крышей сосуществуют несколько вполне сформировавшихся муравейников? - Именно так и обстоит дело. Еще раз перечислим основные характеристики, указывающие на самостоятельность колонн. Сюда относятся пространственная и пищевая обособленность в гнезде и на кормовом участке, устойчивость состава, отсутствие передачи из колонны в колонну оперативной информации о состоянии и делах на территории, фактически автономное развитие.
У полигинных видов каждая колонна в состоянии перейти в самостоятельное гнездо. Это довольно часто происходит при сильном разрушении муравейников (например, кабанами). На месте разрушенного гнезда, на сохранившемся гнездовом валу или неподалеку от него, возникают несколько муравейников. Число таких гнезд обычно равно числу колонн в исходной семье. Иногда из семьи уходит только одна колонна, нередко часть колонн объединяется в одном из вторичных муравейников, а часть - в другом. При таких внутренних разделах соблюдается в общем-то лишь одно правило: гнезда меняются, семьи меняются, а колонны остаются, сохраняя свой кормовой участок и дорогу.
Но силы, ранее удерживавшие их в одном гнезде, сохраняются. По крайней мере первое время. Впрочем, понятие «первое время» здесь относительно, оно может растянуться и на несколько лет. Бывают и быстротечные варианты, когда уже через две-три недели все отношения между бывшими сородичами прекращаются. Это случается чаще при обосновании каждой колонной своего гнезда. Связи между групповыми вторичными муравейниками гораздо устойчивее.
Дальнейшая судьба вторичных муравейников различна. Колонны, прервавшие связи, становятся самостоятельными семьями и далее развиваются как одиночные муравейники. Если связи между гнездами устойчивы, события принимают почти фатальный характер. Вторичные гнезда исходно равны или почти равны друг другу, так как равны основавшие их колонны. Некоторое время, не более одного-двух лет, это равенство сохраняется. Между муравейниками существует оживленная дорога с интенсивным взаимным обменом особями. Обмен всегда равный - один к одному. Семьи продолжают придерживаться принципа, действовавшего, когда они еще были колоннами одного муравейника.
Но колонны развиваются с разной скоростью. И семьи тоже. Один муравейник растет немного быстрее, другой - медленнее или вообще не растет. Спустя некоторое время различия между ними становятся настолько существенными, что меньший становится чем-то вроде отводка у более крупного. И тогда более крупный поглощает, присоединяет его. Население маленького гнезда переселяется при этом в большое. Но опять же колонны остаются. Этапы такого воссоединения в одном гнезде схематично изображены на рис. 10. Отметим, что все это возможно только у относительно небольших одинарных муравейников. Если муравейники стали взрослыми и в них произошло разделение колонн, подобная ситуация уже исключена.
Рис.10. Воссоединение колонн, перешедших в отдельные гнезда после разрушения исходного муравейника
1 - исходный муравейник с двумя колоннами,
2 - равные по размерам
гнезда колонн непосредственно после обособления,
3 - нарушение
равенства колонн в результате их неравномерного роста,
4 - различия в
силе колонн достигают критической величины,
5 - колонны объединились
в более крупном муравейнике, вслед за этим часто следует деление усилившейся-
колонны.
На первый взгляд объединение, в общем гнезде при таких обстоятельствах противоречит изложенному выше принципу выравнивания колонн путем выделения отводка. Там выделение, а здесь та же неравноценность колонн ведет к объединению. Однако противоречия нет. Отводок должна выделить наиболее сильная колонна, входящая в муравейник. Но если колонна в муравейнике одна, ей не с кем сравнивать свою мощь. Система из двух одноколонных муравейников после усиления одного из них начинает несколько напоминать материнский муравейник и отводок. Но и это лишь внешнее подобие, так как отводок может возникнуть только у муравейника с несколькими колоннами, т. е. ситуация искусственная и именно поэтому неустойчивая.
И все-таки система существует один-два года?! В том-то и дело, что неустойчивой система становится лишь после нарушения равновесия разбежавшихся по отдельным гнездам колонн. Когда различия в размерах семей становятся кратными, поглощение меньшей происходит быстро, как правило во время весеннего или осеннего массового обмена между гнездами. Такие обмены нередко интенсивнее летних. Поэтому они и являются обычно как бы поворотным пунктом в жизни муравейников.
В объединении муравейников проявляется очень сильная тенденция муравьев к концентрации. Об этом мы уже говорили в первой главе. Она даже доминирует при отсутствии взрослых муравейников, например, после искусственного переселения отводков. Аналогичная ситуация складывается и в результате распадения семьи на несколько мелких. Концентрация муравьев протекает одинаково в обоих случаях. Следствием объединения семей-недоростков в одном гнезде оказывается последующий быстрый рост этого муравейника. Причин тут, видимо, немало: улучшение температурного режима в гнезде (в большом гнезде он успешнее выдерживается), повышение конкурентоспособности семьи и, что, наверное, самое главное, восстановление естественной структуры семьи. По-видимому, мы не ошибемся, если назовем подразделение большой семьи ряда видов на колонны обязательным условием их нормального существования и развития. Семья, состоящая из одной колонны, - всего лишь начальный этап жизни муравейников многих видов.
А что же происходит в тех случаях, когда «разбежавшиеся» колонны неопределенно долго остаются равными по численности? Этот вариант тоже ведь не исключен. Встречается и такое: два муравейника растут рядом, связанные обменной дорогой. Со временем в каждом из них возникают новые колонны, иногда дело доходит и до образования отводков. А давняя дорога продолжает существовать. Впрочем, не бесконечно долго. В конце концов муравейники-близнецы обособляются, и дорога между ними зарастает травой.
Таким образом, каждая колонна настолько индивидуальна и автономна, что всегда может начать самостоятельное существование. Но все-таки одноколонный муравейник, по-видимому, нерационален. Именно поэтому колонны образуются уже в небольших муравейниках. Общая численность населения такого гнезда находится на уровне 200 тыс., т. е. даже меньше численности одной колонны процветающего муравейника.
Имеется масса видов с небольшими семьями, которые никогда не достигают уровня, необходимого для формирования колонн. Сюда относятся муравьи с наиболее простой организацией и зачастую невыраженной структурой семьи. Для некоторых видов сама возможность образования внутрисемейных группировок особей открыта, но реализуется только в исключительных случаях. Есть и такие виды, у которых достижение семьей определенной (пороговой) численности служит сигналом к разделению ее пополам (гесмозису). Примерно так, как это происходит у медоносной пчелы. Только события, естественно, разворачиваются в гнезде или на поверхности почвы. Гесмозис у муравьев рода афеногастер описан французским энтомологом А.Леду в 1973 г. и рода мономориум - индийскими мирмекологами супругами Сеансами в 1971 г.
При делении старая самка вместе с частью рабочих покидает свой муравейник и основывает новый. В оставшемся гнезде из отложенных старой самкой яиц выкармливаются молодые самки, одна из которых после оплодотворения начинает яйцекладку. Эти виды монотонные, поэтому гесмозис связан с обеспечением самками обеих частей делящейся семьи. Этим обусловлено столь значительное сходство гесмозиса у данных видов муравьев и у пчел. Но гесмозис происходит и у некоторых полигинных муравьев, например у повсеместно распространенных у нас мирмика. Деление семьи мирмика, прослеженное А. В. Демченко, происходило по упрощенной схеме: самки, так же как и рабочие, делятся на две группы, после чего одна из группировок покидает старое гнездо и переходит в новое, сооружение которого начинается заблаговременно частью рабочих, муравьев.
Здесь, как и в гнездах формика, длительный процесс наращивания численности ведет к вызреванию в недрах процветающей семьи более мелких и соизмеримых с видовым оптимумом объединений индивидов. Различия в том, что семья не усложняется структурно, а распадается на две. Это происходит, очевидно, как следствие относительно слабых внутрисемейных связей. Отсутствует та могучая центростремительная сила, которая, как мы видели на примере рыжих лесных муравьев, не только доминирует над автономными силами колонн внутри гнезда, но и способна удержать колонны в рамках общей системы даже при его распадении. Вероятно, развитие механизмов, обеспечивающих сохранение целостности семьи при делении семьи на колонны, является тем эволюционным достижением, который по своему значению для дальнейшей судьбы муравьев намного превосходит даже сам принцип деления.
К этому вопросу мы еще вернемся, а пока лишь отметим, что в любом варианте деление общины происходит по дихотомическому принципу, что образующиеся при этом новые семьи или колонны - организационно завершенные и жизнеспособные единицы, способные к самостоятельному существованию. И, наконец, первая задача, которую решает в такой ситуации семья, - это оптимизация своей численности и организационной структуры. Образование новых муравейников в стратегию муравейника не входит, а является следствием решения первой задачи.
Необходимо отметить, что образование колонн резко повышает жизнеспособность каждого муравейника и комплексов муравейников. Это особенно важно для видов, сооружающих наземные купола из легких строительных материалов. Такие гнезда не защищают их обитателей от крупных млекопитающих: медведей, барсуков, кабанов, - а, наоборот, привлекают их. В этих условиях рассредоточение колонн по отдельным гнездам - единственное средство пассивной защиты, которое могут противопоставить муравьи. Небольшие муравейники звери почти не трогают. В результате одним из следствий массовых разорении муравейников кабанами оказывается увеличение общего числа гнезд при уменьшении их средних размеров.
В арсенале ряда видов имеются и некоторые другие средства выхода из критической ситуации, когда семья разрастается до таких размеров, что нормальное существование ее как единой системы индивидов становится невозможным. Речь об этом пойдет в последнем разделе данной главы.
СЕМЬЯ КАК СИСТЕМА КОЛОНН
Итак, можно охарактеризовать семью муравьев как систему, состоящую из нескольких идентичных по составу и сравнимых по численности особей подсистем - колонн, которые имеют постоянный состав, занимают определенные части гнезда. Эти части (секции, секторы) подобны друг другу. Кормовой участок, используемый каждой колонной, также постоянен.
Колонны сравнимы по численности, и их отношения строятся не на иерархическом принципе, а на многостороннем равнодействии всех входящих в систему подсистем. Возможность проявления иерархических отношений исключается, так как вслед за непомерным усилением одной из колонн неотвратимо следует либо разделение этой колонны на две, либо выделение части ее населения в отводок. Муравейник действует как целостная система, хотя какие-либо специальные образования, координирующие рост и жизнедеятельность колонны или управляющие ими как подсистемами целого, отсутствуют.
Перечисленные свойства присущи особому координационному типу систем, представляющему собой довольно низкий уровень интеграции. На этом уровне в наименьшей степени проявляется зависимость подсистем друг от друга и от системы в целом, что мы и видим на примере муравьев. Общесемейные реакции формируются путем прямого сопоставления колонн между собой и каждой из них со всеми остальными.
Однако имеются и некоторые серьезные отличия системных свойств муравейника от традиционных координационных систем, например популяции. Из них стоит выделить три.
Во-первых, это ограниченное число подсистем, входящих в муравейник. Обычно 2-5, максимально 12, в то время как в любой благополучной популяции тысячи и миллионы индивидов. Немногочисленность подсистем повышает удельный вес каждой из них, делает значимым для состояния семьи любое изменение их количества. Одновременно функциональные связи между всеми колоннами становятся обязательными и регулярными. Прекращение таких связей означает выпадение колонны из семьи, чего в нормальных условиях не бывает. Все это приводит к значительно большей организованности муравейника как одного из вариантов координационных систем.
Во-вторых, колонны объединяются не только функциональными связями, но и общим местом обитания - гнездом, которое является основным выражением интеграции их деятельности. В общем гнезде регулярные связи всех колонн обретают важное дополнительное свойство: они осуществляются по строго определенным путям. И если в муравейнике нет координирующего функционального образования, какой-нибудь колонны-координатора, то там имеется определенная зона (внутренний конус или группа поверхностных ходов и камер), где сосредоточены основные пути координации. Это дополняет вытекающие из первого положения системные возможности семьи муравьев, еще более укрепляя ее организационную целостность, делая ее функционально более эффективной.
В-третьих, система контролирует изменение числа колонн уже на начальном этапе, регулируя их численность и размеры. Благодаря этому появилась возможность сопряженного устойчивого роста подсистем и семьи как объединяющего их целого. Почти стихийный рост колонн преобразуется в результате их взаимодействия в зарегулированный рост семьи. На определенном уровне численности появляется стабильность. Ее обеспечивает высокая степень организации муравьиной общины. Без четкой организации существование муравьев в таких количествах было бы просто невозможно.
Для более полной характеристики муравейника как системы требуется привлечение данных о системных свойствах самих колонн, т. е. об их поведенческой структуре. Этот чрезвычайно емкий и очень интересный вопрос не входит в нашу задачу. Однако стоит упомянуть, что поведенческая структура колонн достаточно сложна и, как правило, более видоспецифична. Все моменты, связанные с разнообразием каст и размерных групп муравьев, со сменой профессий и взаимодействием профессиональных групп между собой, добычей пищи и ритмикой общей активности, с особенностями поведения муравьев в гнезде и на участке, регулируются в рамках отдельной колонны.
Об организационной структуре муравейников известно еще очень мало. Многие вопросы требуют тщательной разработки, другие - едва лишь затронуты исследователями. Ограничен и круг видов, используемых в качестве модельных для этих целей. Но все-таки уже сейчас можно, хотя бы в первом приближении, представить себе, сколь сложна и совершенна организация семьи муравьев, насколько своеобразен их мир, преодолевший не одно, казалось бы, непреодолимое препятствие - мир, где собранные по крупинкам достижения индивидов синтезированы в строгие и действенные формы организации.
ПОЛИКАЛИЯ У МУРАВЬЕВ
Для рациональной организации семьи и при перенаселенности гнезда у муравьев наблюдается не только образование колонн. В арсенале ряда видов имеются некоторые другие возможности выхода из критической ситуации. В настоящее время мы имеем сведения еще о двух типах внутрисемейной организации - это поликалия* и диффузные гнезда*.
Поликалия - обитание одной семьи в нескольких взаимосвязанных гнездах - очень широко распространена у муравьев, в первую очередь среди видов, в семьях которых имеется только одна самка. Помимо основного, самого крупного гнезда муравьи сооружают вспомогательные, находящиеся на некотором расстоянии от основного и выполняющие различные функции. Эти гнезда гораздо меньше по размерам и могут существовать различное время: несколько дней, в течение одного сезона, ряда лет или всего срока существования семьи.
Изучение поликалии начато еще в прошлом веке выдающимся французским мирмекологом А.Форелем. С тех пор описаны вспомогательные гнезда*, различающиеся по своим размерам, сроку существования и назначению. Небольшие временные гнезда муравьи строят при переселении на новое место, пряча там расплод, самок и внутригнездовых рабочих. Во второй половине лета большинство муравьев, обитающих в умеренном поясе, переключаются на сбор пади корневых тлей. При этом для каждой колонии тлей строится небольшое убежище, состоящее из нескольких камер различных размеров. В степи корневые тли - основной источник пади для муравьев. Здесь убежища тлей почти постоянны. В каждом убежище находится небольшой постоянный гарнизон рабочих, поддерживающих гнездо в порядке и охраняющих его. Убежища для тлей существуют около двух месяцев. Пахучие муравьи-древоточцы накрывают колонии тлей картонным чехлом. Постоянно укрывают колонии тлей в галереях рабочие желтого пахучего муравья.
Чтобы пережидать непогоду, муравьи сооружают микрогнезда, или, как их называл А.Форель, «станции» и «павильоны». Такие укрытия часто находятся в стороне от дорог. Еще одним типом вспомогательных гнезд являются кормовые почки, сооружаемые рыжими лесными муравьями, муравьями-древоточцами и некоторыми муравьями-жнецами на дорогах в тоннелях жилого муравейника. Кормовые почки невелики по размерам, в них не бывает молоди, функционируют такие гнезда только летом. В этот период в кормовой почке обитают от нескольких сот до нескольких тысяч фуражиров и строителей.
Временные вспомогательные гнезда облегчают муравьям только использование кормового участка, они не влияют на функции основного муравейника. Невелико значение этих гнезд и для рассредоточения жителей перенаселенного муравейника. Но их роль в удержании территории и более рациональном ее использовании существенна.
От старой сосны, в сгнившей сердцевине которой поселилась семья муравьев-древоточцев, в разные стороны расходятся тоннели. Чтобы обнаружить их, достаточно приподнять слой подстилки из опавшей хвои, тоннели могут быть довольно длинными, до 10-15 м. На разном удалении от гнезда из тоннелей имеются выходы на поверхность. Отсюда отправляются на охоту фуражиры. Заканчиваются тоннели в кормовых почках. Каждое гнездышко так же, как и сам муравейник, размещается в отмершей древесине: пнях, лежащих стволах деревьев. Они и служат опорными пунктами. Из кормовых почек действует основная масса фуражиров. Сюда же поступает большая часть добычи. При большом количестве добычи она поначалу накапливается в почках, а уж затем постепенно переносится в муравейник.
У многих муравьев отработана четкая эстафетная транспортировка пищи: одни рабочие приносят добычу в кормовую почку или к выходу из тоннеля, другие - транспортируют ее оттуда в муравейники. Эстафета экономична: она позволяет обходиться на участке минимумом опытных фуражиров. А транспортировку добычи по тоннелю может осуществить любой.
Большинство кормовых почек остаются неизменно в этом качестве. Но некоторые из них в дальнейшем служат основой для сооружения настоящего муравейника. В специальной литературе описано преобразование в жилые гнезда кормовых почек у рыжих лесных муравьев и пахучего муравья-древоточца. У формика это обычно связано с основанием нового отводка, а у пахучего муравья-древоточца - с разделением семьи.
Другой вариант поликалии связан с разделением гнезд на летние и зимние. Зимние - обычно более крупные, капитальные, с мощной подземной системой ходов и камер. Они расположены в укрытых местах: в пнях, в середине кустов и т. п. Летние гнезда находятся на открытой местности. Они служат муравьям в течение всего периода активности, а в зимние семья перебирается поздней осенью (на зиму в летних гнездах остаются единичные особи).
Летние и зимние гнезда были детально описаны на примере кроваво-красного муравья-рабовладельца одним из пионеров изучения поведения муравьев - Э.Васманом в 1910 г. Сходная картина прослежена у некоторых других видов. Летних гнезд, жилых и вспомогательных, обычно больше, чем зимних. Как правило, концентрация особей в зимовочных гнездах всегда выше. Но из всякого правила бывают исключения. Изучая в Финляндии биологию черноголового муравья, Р. Розенгрен установил, что у этого вида все происходит наоборот: летом муравьи обитают в большом муравейнике, а зимуют вне основного гнезда в небольших зимовочных камерах, сооружаемых вокруг муравейника на различном удалении от него.
Более сложные варианты поликалии возникают при единовременном использовании семьей нескольких жилых гнезд, одного - центрального и нескольких - периферийных. В этом случае отдельные гнезда несут различные функции. Сам факт разделения функций гнезд в по-ликалическом муравейнике не удивителен, ведь подобное разделение складывается уже при монокалии, т. е. при обитании семьи в одном гнезде. Но там различные функции имеют отдельные части одного гнезда (например, внутренний конус и гнездовой вал у рыжих лесных муравьев). Здесь же обособление частей гнезда пошло дальше и вылилось в принцип: гнездо - функция. Сравним единое сложное гнездо пятнистого муравья с поликалической системой муравьев-портных (рис. 11).
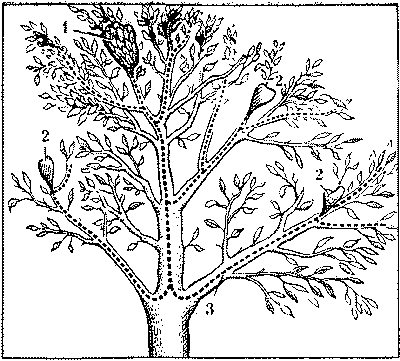
Рис. 11. Цоликалическии муравейник муравьев-портных на дереве
1 - центральное гнездо с самкой, 2 - периферийные гнезда, 3 - дороги
У пятнистого муравья гнездо состоит из трех частей: центрального гнезда, где живет единственная самка и ее свита; большого внутреннего гнезда (сферы), где находятся только рабочие - имаго; периферийного гнезда {сферы) с рабочими и молодью. В центральном гнезде муравьев-портных также находится самка и ее свита. Личинок младших возрастов уносят в разбросанные по кроне дерева периферийные гнезда, где их выращивают до взрослого состояния. Есть также небольшие гнезда, в которых находятся одни рабочие.
В обоих случаях добыча доставляется фур ажирами сначала в периферийные гнезда (или сферу) и лишь оттуда поступает в центральное гнездо. Этот принцип сохраняется у всех муравьев, перешедших к поликалической форме существования. Естественно, что между гнездами имеются оживленные дороги и постоянный обмен. Из центрального гнезда в периферийные идет пополнение молодью, обратно - поступают взрослые муравьи, а также пища. Функциональная цельность такого муравейника не вызывает сомнений.
Поликалия позволила муравьям увеличить численность общины и размер используемого кормового участка. Вся территория внутри кольца вспомогательных и периферийных гнезд используется одной семьей даже в тех случаях, когда в других частях кормового участка статус охраняемой территории отсутствует. В этом одно из преимуществ поликалии.
Вообще с переходом к поликалии конкурентоспособность семьи значительно возрастает. Одновременно муравьям гораздо легче приспособиться к среде: гнезда располагаются в разных условиях, появляется возможность маневра при изменении погоды или режима освещенности отдельных гнезд. Виды, освоившие поликалическую форму существования, смогли подняться на довольно высокий уровень численности населения семьи. Подсчет особей в семье муравьев-портных, произведенный Ф. Вандер-планком в 1960 г., показал, что в крупном поликалическом муравейнике обитает более 100 тыс. особей. По данным московского энтомолога А.М.Кравченко, численность семьи пахучего муравья-древоточца еще выше - более 300 тыс. особей.
Уровень численности особей в семье, которого достигают муравьи, освоившие очередной этап развития социальной организации, убедительнее всего показывает адаптивную ценность того или иного достижения. Ряд поликалических видов преодолел стотысячный рубеж численности, т. е. достиг и даже превзошел уровень, при котором у муравьев с наиболее многочисленными семьями (рыжие лесные муравьи) происходит образование колонн.
Это доказывает, что, хотя поликалия располагает несколько меньшими возможностями, нежели система колонн, она оказалась очень важным шагом в эволюции социального образа жизни у муравьев.
Диффузные гнездаможно рассматривать как один из вариантов поликалии, связанный с использованием муравьями готовых полостей и камер, небольшие размеры которых ограничивают возможное число обитающих в них особей. Как правило, диффузные гнезда встречаются у муравьев, особи которых имеют небольшие размеры. Семья как бы растекается по всевозможным укрытиям, легко меняет их, сохраняя при этом связи между отдельными частями. Вместе с рабочими по микрогнездам рассредоточены и самки. Семьи таких видов имеют необыкновенно пластичную, подвижную структуру. Они легко распадаются, и обитатели каждого микрогнезда могут стать основой новой большой семьи. Состав и численность населения микрогнезд могут меняться: увеличивается размер гнезда - там начинает собираться больше муравьев, при необходимости сменить жилище группировка может разделиться на несколько, чтобы потом вновь собраться в следующей обители.
Численность семьи бывает значительной - до 10 тыс. и более. Среди муравьев, обитающих таким образом, наиболее известен фараонов муравей, доставляющий массу хлопот санитарно-эпидемиологической службе у нас и за рубежом. Фараонов муравей ведет настоящее наступление на жилище человека, проникает в библиотеки и на предприятия пищевой промышленности. В больницах он способен совершить настоящие преступления, нарушая стерильность операционных и разнося инфекцию. Борьба с этим крошечным (около 2 мм) желтоватым муравьишкой крайне затруднена именно из-за его особенностей, обусловленных диффузно-кочевым образом жизни. Едва заметные щели, забытая корка хлеба, корешок книги - все пригодно для поселения фараонова муравья. Просачиваясь из квартиры в квартиру, из палаты в палату, он очень быстро осваивает различные помещения, постепенно становясь одним из самых распространенных и нежелательных спутников человека.
Надо сказать, что роль муравьев с диффузными гнездами в природе невелика. Они заселяют брошенные ходы жуков, трещины, плоды и даже шипы некоторых растений и т.п. Впрочем, приспособленность к жизни в чужих жилищах пригодилась не только фараонову муравью и не только в жилище человека. Семьи столь же мелкого по размерам блестящего муравья-малютки отлично приспособились к обитанию в гнездах рыжих лесных муравьев. Крошечные камерки размещаются в простенках между камерами гнезд муравьев-хозяев. Оттуда по ходам диаметром менее 1 мм муравьи-малютки похищают добычу хозяев и даже их яйца и личинок. И всюду, где есть гнезда рыжих лесных муравьев, встречается вид-приживалка. Муравьи этого вида уже не могут жить самостоятельно.
Муравьи с диффузными семьями имеют перед остальными муравьями то преимущество, что им не надо строить жилища. И поэтому их пора наступает тогда, когда появляются чужие жилища, где тепло и всегда есть корм.
Как мы убедились, муравьи использовали разнообразные приспособления, открывающие дополнительные возможности роста семьи за счет усложнения ее организационной структуры. В общих чертах смысл таких приспособлений сводится к решению одной задачи - формированию внутрисемейных организационных единиц и функциональной дифференциации самого гнезда. Соотношение этих составляющих различно в рассмотренных вариантах, различен их эффект. Но все происходит внутри одной семьи. Многие виды ограничились этими достижениями. Следующий этап развития социального образа жизни у муравьев - выход на подсемейный уровень организации. Предпосылки к возникновению подсемейных структур сложились в процессе развития организационной структуры отдельной семьи муравьев. Возможности их образования - в механизмах, регулирующих жизнь муравьиной общины.
3. МУРАВЕЙНИК И КОЛОНИЯ
В предыдущей главе мы подошли к тому моменту в жизни семьи муравьев, когда внутренние возможности общины отрегулировать свой состав и организационную структуру исчерпаны. Целостность семьи под угрозой. В этой критической ситуации происходит деление общины. Тут возможны два варианта: либо семья делится на две равные, сразу же обособляющиеся и становящиеся чужими части (гесмозис), либо из муравейника уходит небольшая часть населения и основывает отводок (почкование). Именно второй ведет к образованию подсемейных структур.
Дело в том, что отпочковавшаяся часть - дочерняя семья - еще долгое время не теряет связи с материнским муравейником. Эта часть организационно оформлена: в ней есть свои самки и рабочие муравьи всех функциональных групп. Появляется у нее и свое гнездо, но связи между старым и новым муравейниками сохраняются. В итоге возникает система родственных муравейников, объединяемых обменными дорогами и определенным типом отношений.
ВОЗНИКНОВЕНИЕ КОЛОНИИ
Образование отводка происходит обычно в определенные для каждого вида сроки. Вот как происходил начальный этап выделения нового муравейника у волосистого лесного муравья в 1967 г. в подмосковном ельнике (Солнечногорский лесокомбинат). В течение нескольких дней (15-21 сентября) муравьи проявляли повышенную активность. Огромное количество рабочих возбужденно перемещалось по куполу гнезда и на участке. Особо оживленно было в зоне одной из дорог, на которой предстояло сооружение отводка.
В разных местах кормового участка этой дороги собирались группы рабочих, от нескольких десятков до нескольких сот муравьев; 21 сентября под березой, что росла в 12 м от гнезда, началось сооружение нового муравейника, сначала укрывавшегося под сухими листьями.
Через несколько дней листва была покрыта небольшим плоским куполом из хвои диаметром около 15 см. Вокруг купола появился небольшой вал из свежей почвы:
муравьи сооружали подземные гнездовые помещения. Это были подготовительные работы, а само переселение началось 22 сентября. В тот день с утра на куполе материнского муравейника появилось множество крайне возбужденных рабочих особей, в основном крупных размеров. К 11 часам на поверхность купола вышло несколько десятков самок, каждая из которых была. окружена толпой снующих рабочих.
Измерение муравьев из свиты самок показало, что эти муравьи значительно крупнее рабочих, находящихся на поверхности купола в обычное время. Рабочие непрестанно облизывали медленно передвигавшихся по вершине гнезда самок.
Тем временем муравьи-носильщики начали курсировать между сооружаемым отводком и материнским муравейником и переносить в отводок находящихся на куполе взрослого гнезда самок и рабочих из их окружения. Образовалась широкая дорога, по которой в отводок непрерывным потоком поступали самки и рабочие особи. Рабочие, которых переносили носильщики, были внутригнездовыми с характерным для этой категории особей раздувшимся из-за увеличенного жирового тела брюшком. Ежеминутно в отводок доставлялось 25-30 особей. Фуражиры переходили туда самостоятельно. На следующий день картина повторилась. Затем возбуждение на куполе взрослого муравейника улеглось, но массовое переселение в отводок продолжалось до середины октября.
Переселенцы благополучно зимовали во вновь отстроенном гнезде, достигшем к этому времени 30 см в диаметре и 15 см в высоту.
Отводки основываются на различных расстояниях от материнского гнезда. У рыжих лесных муравьев удаление отводка колеблется в пределах от 10 до 120 м. Дочерние семьи кроваво-красного муравья-рабовладельца поселяются в 15-30 м от материнского гнезда, у тонкоголовых муравьев - в 5-25, у муравьев-жнецов - в 15-20, у муравьев-фаэтончиков в Туркмении - в 10-25 м.
В результате почкования возникает новое образование - колония, состоящая из материнского и одного или нескольких дочерних муравейников. Семьи, входящие в одну колонию, связаны дорогами, по которым производятся обмены. Эти дороги называются в отличие от кормовых дорог* муравьев обменными дорогами*, что отражает их основное назначение и функцию в колонии. Как уже говорилось, именно благодаря обменам и появилась возможность сохранения в течение длительного времени особых лояльных, так называемых колониальных отношений между муравейниками.
Почкование - не единственный способ возникновения колонии. Довольно часто колония появляется при установлении колониальных отношений между уже существующими семьями разного происхождения, но поселившихся по соседству. Происходит породнение чужих семей муравьев одного вида, что впервые описано Ц.И.Мариковским в 1962 г. Только таким путем формируются искусственные колонии рыжих лесных муравьев. Нами выяснено, что при породнении чужих семей в естественных и искусственных популяциях рыжих лесных муравьев связь между такими семьями устанавливается с помощью небольшого специального гнезда - промежуточной почки, сооружаемой на границе кормовых участков муравейников. В отдельных случаях на дороге, соединяющей соседние гнезда, строятся сразу две промежуточные почки. С обеих сторон к промежуточной почке подходят обменные дороги, по которым во встречных направлениях переносятся самки, рабочие муравьи и молодь. Некоторое время у такой почки существует небольшое население из одних рабочих особей. Значительная часть обменов производится здесь, а муравьи-носильщики, пришедшие на купол чужого гнезда, встречаются там довольно враждебно.
Через две-четыре недели, после проведения большого числа обменов, промежуточную почку покидают, и между породнившимися семьями устанавливается прямая связь. Но если дороги пересекались здесь под каким-то углом, то и после установления прямой связи обменная дорога долго еще сохраняет первичные изгибы, указывающие на места соприкосновения охраняемых территорий различных семей или колоний, связанных вторичными обменными дорогами. Поэтому вместе с другими показателями дорожная сеть может таким образом использоваться при анализе структуры и происхождения объединений муравьиных семей.
При породнении чужих семей бросаются в глаза два непременных условия, соблюдаемые в этом случае муравьями. Во-первых, породниться таким путем могут только чужие в данный момент семьи. Во-вторых, муравейники должны значительно отличаться по своей численности, Суть возникновения колонии посредством породнения чужих семей заключается в вовлечении более сильным, процветающим муравейником ослабевшего соседа в свою обменную систему.
Опять обмены! Обмены пищей между индивидами в колонне (семье). Обмен особями между колоннами одной семьи. Обмены - средство сохранения дружественных отношений между родичами и вовлечение в круг родичей новых особей и целых муравейников.
Ради чего это происходит? Только ради возможности построить побольше гнездо, в котором легче регулировать гигротермический режим? Это объяснение можно принять во внимание, если речь идет о концентрации муравьев в одном гнезде. Но ведь сейчас перед нами другая ситуация: оба муравейника остаются на своих местах.
Неужели колония возникает на тот крайний случай, когда в какой-нибудь черный для муравьев год семьи настолько ослабевают, что только слияние их в одну спасает уцелевших от гибели в пустеющих гнездах? Подобные слияния нередки.
Но причина в другом. Стратегия муравьев заключается в максимальном увеличении числа индивидов в системе, удлинении обменной цепи. Ведь чем она длинней, тем устойчивей система. А пространственная разобщенность семей в колонии при этом не так важна: уже при переходе к поликалии муравьи решили эту проблему даже без нарушения целостности семьи.
И если колониальные связи не устанавливаются между всеми соседними муравейниками, то, по-видимому, лишь потому, что имеется ряд других мощных и препятствующих процессу консолидации факторов. Снять действие этих факторов или хотя бы ослабить их противодействие - и этакий панмуравейник в пределах, например, лесного массива, пожалуй, мог бы стать реальностью. Впрочем, как мы увидим ниже, при определенных условиях такая возможность частично реализуется.
Некоторый из препятствующих объединению факторов уже обсуждались: прежде всего специфический гнездовой запах, четко делящий всех муравьев на своих и чужих. Этот барьер очень высок. Но все-таки он преодолим с помощью интенсивных обменов и небольшого временного гнезда диаметром 15-30 см и высотою 5- 7 см, без подземной части и дифференциации купола. Такое гнездо рыжие лесные муравьи могут построить за день. Очевидно, необходимо еще что-то.
Может быть, влияют размеры муравейников, абсолютные и относительные? Здесь еще многое неясно, но значение этого показателя, несомненно, велико. Ведь недаром для образования колонии требуется, чтобы семьи различались по численности. Попробуем оценить этот фактор в следующих разделах данной главы. А пока вни-- мательно поглядим на первое условие породнения чужих муравейников. Они должны быть чужими, т. е. не входить в одну обменную систему. Но почему обмен, порою весьма слабый, особями налагает вето на установление прямых связей между расположенными рядом гнездами? Не потому ли, что они и так уже входят в одну обменную систему? Если у одного из соседей связь с материнским гнездом оборвется, тогда прямые связи устанавливаются. Такие случаи отмечены в естественных комплексах муравейников. Чтобы яснее представить значение обменов особями для муравьев, возможно подробнее разберем их роль в жизни колонии.
МАТЕРИНСКАЯ И ДОЧЕРНЯЯ СЕМЬИ
Начальный этап в жизни колонии - просто большое переселение в довольно короткий срок, измеряемый у одних видов днями, у других - неделями. В это время из материнского муравейника поступает в отводок и пища. Дочерняя семья - фактически лишь часть материнской семьи, у которой нет ни собственного охраняемого участка, ни возможности выращивать молодь. Она еще ничего не дает в обмен на получаемых от материнской особей и пищу. Постепенно численность дочерней семьи растет, и характер ее отношений с материнской начинает меняться. На обменной дороге появляются муравьи-носильщики, доставляющие особей из отводка в материнское гнездо. Индивидуальным мочением носильщиков в колониях северного лесного муравья в Подмосковье и муравья-фаэтончика в Туркмении было установлено, что эти особи выходят налегке из своего гнезда и направляются а другое, где забирают объекты обмена (личинок, куколок или внутригнездовых рабочих) и переносят их в свой муравейник. Учитывая схему действия носильщиков, можно считать, что начало обоюдных обменов означает перелом в отношении материнской семьи к дочерней. Это как бы признание отводка отдельным муравейником. Признание еще далеко не полное.
Отводок обычно сооружается очень энергично. У фаэтончиков и муравьев-жнецов уже через две-три недели он имеет все основные элементы гнезда. У рыжих лесных муравьев через месяц-полтора отводок также принимает вид вполне законченного гнезда с внутренним конусом и подземной частью. Это позволяет поддерживать необходимый для развития молоди температурный минимум. Дальнейшие отношения между материнской и дочерней семьями будут зависеть от соотношения их сил (численности) и продуктивности. Сначала наступает период интенсивных взаимных обменов. У муравьев формика он совпадает с достижением семьей размеров автономного муравейника.
В это время у растущего муравейника начинает складываться и свой кормовой участок. Он еще маленький, проницаем для фуражиров материнской семьи, которые «по старой памяти» продолжают здесь охотиться. Ведь в абсолютном большинстве случаев отводок сооружается в пределах ее охраняемой территории, где-нибудь в периферийной зоне одной из дорог.
Обмен обычно почти равный. На этом этапе в какой-то мере еще выдерживается обменный принцип колонн в гнезде - один к одному. Но появляются и нарастают различия. Материнский муравейник, как более многочисленный, с лучшей терморегуляцией, раньше начинает фуражировку утром, и его носильщики раньше приходят в отводок. А прекращают свою работу носильщики материнской семьи позже носильщиков дочерней. В итога утром и вечером каждого дня материнская семья собирает некоторую «дань» с дочерней. Днем обмен выравнивается, но по итогам суток материнская семья всегда в выигрыше.
Точно так же обстоит дело и в зависимости от сезона. Весной и осенью материнская семья получает гораздо больше (нередко в несколько раз), чем отдает. Положение таким образом изменилось в принципе: отводок служит источником дополнительных особей для материнского муравейника. Но, учитывая небольшие размеры дочерней семьи, в абсолютных величинах прибыль эта невелика.
Весенне-осенние обмены могут быть очень интенсивными, чем, по-видимому, и объясняется рассмотренное выше положение, что именно в эти периоды решается вопрос, быть или не быть отводку. Если дочерняя семья после зимы или неблагоприятного лета оказывается к урочному моменту слишком маленькой, неравный обмен настолько истощает ее, что оставшихся в гнезде муравьев уже недостаточно для жизни отдельной семьей. И тогда ее остатки просто возвращаются в материнскую общину. Таким образом отводок в какой-то мере является резервом материнского муравейника. Процесс разделения общины в определенных условиях обратим. Это еще одно важное отличие почкования от гесмозиса у пчел и некоторых муравьев, о чем говорилось в предыдущей главе.
При благоприятных условиях соотношение сил муравейников в колонии постепенно меняется в пользу дочернего. В период интенсивного выделения отводков материнская семья растет незначительно, поэтому изменения соотношения целиком зависят от роста дочернего муравейника. А он обычно растет довольно быстро, несмотря на невыгодный для него обмен с материнским муравейником. Конечно, при условии, что на кормовом участке достаточно пищи и имеется свободная территория, которую можно освоить.
С ростом отводка интенсивность обменов л колонии начинает падать. Для колонии это имеет очень серьезные последствия. Смысл постоянных обменов между муравейниками заключается в том, что обмены позволяют сглаживать качественные различия между обменивающимися семьями, поддерживать дружественные отношения и избегать столкновений. А сглаживать есть что. Обитание дочерней семьи в новом гнезде и пища, добываемая в определенном участке, приводят к изменению семейного запаха отводка, вначале совпадавшего с запахом материнского гнезда. Чем сильнее отличаются условия обитания и кормовые участки материнской и дочерней семей, тем быстрее накапливаются различия - причина отчуждения муравейников. Пока отводок молод и обмены интенсивны, различия минимальны: С ростом дочерней семьи нивелирующая роль обменов даже при постоянной интенсивности снижается, что ведет к снижению интенсивности обменов, а это, в свою очередь, - к ускоренному накоплению фототипических различий. И чем энергичнее растет отводок, тем это происходит быстрее.
Обмены только пищей и нерегулярные весенне-осенние обмены наблюдаются на последнем этапе существования колонии, предшествующем полному отделению дочернего муравейника. Колонии на этом этапе описаны П.И.Мариковским и мирмекологом из ФРГ Г.Кнайтпем.
Наиболее устойчивым оказывается обмен пищей. Впрочем, строго говоря, поступление добычи в материнское гнездо из отводка нельзя назвать обменом по двум причинам. Во-первых, это процесс односторонний, только в материнское гнездо. Как только отводок подрастает настолько, что начинаются взаимные переносы особей, подкармливайте его со стороны материнской семьи прекращается. Далее пищевой поток направлен только к материнскому гнезду. Получается еще одна форма «дани».
Правда, дань эта собирается не столько в самом отводке, сколько на его кормовом участке. Здесь отношения фуражиров из двух гнезд явно неравноправны. Фуражиры из материнской семьи почти до конца существования колонии могут проникать на кормовой участок дочерней, хотя он уже давно надежно охраняется от всех других соседей. Постепенно случаев такого проникновения становится все меньше, но все-таки они имеются. А вот фуражирам отводка путь на кормовой участок материнского муравейника всегда закрыт. Отсюда и возникает вторая неточность термина. Фактически это не обмен и не «дань» с муравейника, а скорее «концессия», по которой фуражиры одного муравейника охотятся на территории другого.
В тех случаях, когда соотношение сил материнской и дочерней семей изменяется очень быстро, обмен особями между ними иссякает в течение одного сезона. На следующий год еще может быть небольшой весенний обмен и поступление из отводка пищи, но часто не происходит и этого. Все заканчивается в один сезон. События развиваются тем быстрее, чем меньше материнский муравейник, т. е. дочерней семье легче с ним сравняться. Колонии становятся сезонными. Такие колонии имеются, например, у муравьеа-фаэтончиков, численность семьи которых ограничивается несколькими тысячами индивидов.
Можно считать, что в течение всего срока существования колонии обмены между материнским и дочерним муравейниками подчинены закономерным изменениям. И если принять весь период, от возникновения отводка до его отделения, за единицу, графики обменов совпадут для колоний с различными сроками существования.
Вообще при оценке подобных явлений абсолютное время характеризует процесс, но не определяет его. Даже сезонность не в состоянии регламентировать срок существования колонии. Она лишь предлагает наиболее подходящие моменты для решающего проявления назревающих в колонии процессов.
На обмен поступает определенный процент иовых рабочих-имаго и молоди, появившихся в гнезде. Но каков процент, сколь он велик? Чтобы проверить, какая часть обменного материала, имеющегося в материнском гнезде и в отводке, проходит на обмен, мы поставили опыты в колонии волосистого лесного муравья. Схема обменов в колонии дана на рис. 12. Крупный материнский муравейник (Л-10) с куполом 2,2 м в диаметре и высотой гнезда 1,1 м имел один отводок (Л-11) с диаметром купола 1,2 м и несколько кормовых почек на разных дорогах. Всего у муравейника Л-10 было шесть дорог, из которых на рисунке изображены только две, использованные в опытах.
Опыты проводились в августе, когда обе подопытные семьи уже вывели последнее поколение рабочих и обменивались только имаго, во сам обмен производился еще в соотношении 1:1. Как видно из схемы, дочерний муравейник и его кормовая почка ПК-12 находятся на одной дороге материнского гнезда. Кормовая почка материнского муравейника (ПК-13) расположена на ответвлении той же дороги. Вторая кормовая почка материнского муравейника (ПК-22) находится на самостоятельной дороге, т. е. принадлежит другой колонне.
В качестве обменного материала использовались куколки, взятые из постороннего муравейника того же вида. Куколки были сосчитаны и окрашены разными цветами. В первом опыте по 2000 куколок выложили на ПК-12 и ПК-13, т. е. на ответвления одной дороги материнского муравейника. После этого подсчитывали поступление меченых куколок из кормовых почек в материнское гнездо и отводок, а на обменной дороге регистрировали взаимную передачу этих куколок жилыми муравейниками.
Обмен начался через 40-45 минут после поступления первых куколок в муравейники и продолжался до конца дня. При этом отводок передал в материнский муравейник 5,6% куколок, а материнская семья-лишь 1,8%, или а 3,1 раза меньше. Причем все время общий обмен между семьями (с учетом продолжавшихся весь день взаимных переносов внутригнездовых рабочих) оставался равным. Этот опыт, повторенный по той же схеме, дал одинаковые результаты. Фактически мы анализировали обмен в пределах колонны, так как и отводок, и обе использованные в опытах ПК находятся на ответвлениях одной дороги материнского гнезда.
Интересно знать, как и в какой мере участвуют в подобных обменах другие колонны материнского муравейника. Для этого в следующих опытах куколок выкладывали на ПК-13 и ПК-22. На обменной дороге этих куколок учитывали по отдельности.
Специфика опыта заключалась в следующем. Куколки из ПК-13 сразу же попадали в место, откуда носильщики из отводка забирали объекты обмена, а куколки из ПК-22 должны были сначала попасть в эту зону из другого сектора, другой колонны. Последнее обстоятельство сразу же сказалось на результатах. Носильщики из отводка, как и в предыдущих случаях, забрали причитающееся им количество куколок. Но доли куколок, поступивших из ПК-13 и ПК-22, отличались в 5 раз: 1,6% - из ПК-13 и 0,32% - из ПК-22. В следующем опыте отличие было шестикратным, т. е. даже в ходе довольно острого эксперимента лишь 15-20% новых куколок из соседней колонны оказалось вовлеченным в обмен с отводком. Из общего числа куколок менее трети процента пошло на обмен. Все это показывает, что связи между отводком и материнским муравейником - это прежде всего связи между отводком и колонной, из состава которой выделилась дочерняя семья. Для остальных колонн материнского муравейника отводок существует как часть этой колоины. Но для отводка наличие и мощность других колонн в материнском муравейнике значимы.
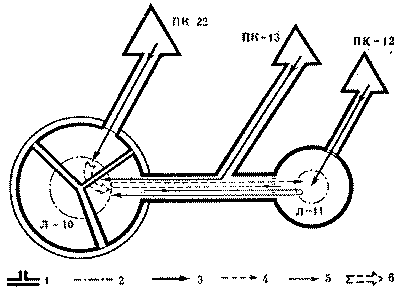
Pис. 12. Фрагмент схемы межсекторного и внутрисекторного обменов куколками в колонии волосистого лесного муравья
Л-10 - материнское гнездо, Л-11 - отводок, ПК-22, ПК-13, ПК-12 - кормовые
почки;
1 - границы секторов материнского гнезда, 2 - границы
внутреннего конуса, 3 - пути поступления куколок из кормовых почек, 4
- маршруты муравьев-иосильщиков из отводка, 5 - маршруты
муравьев-носильщиков из материнского гнезда, 6 - передача куколок из сектора в
сектор
Так или иначе процент обмениваемых муравейниками новых особей невелик, хотя существен для отводка. Чтобы выдержать равный обмен, он должен жертвовать относительио большей частью своего населения. Данная ситуация важна для отводка, особенно если он еще мал по размерам. Ведь можно предполагать, что обменные возможности каждой семьи не беспредельны. Община, разменявшая налево и направо слишком большую часть своего населения, просто утратит свою индивидуальность, целостность. Какой-то предел должен быть. То максимальное число особей, которое может единовременно обменять семья, сохраняя индивидуальность, будет обменным фондом семьи*. Получив новое качество - в виде прошедших линьку личинок, куколок или вышедших из куколок рабочих, семья передает партнеру по обмену определенный процент, соответствующий степени родства между данными муравейниками и отношению их размеров.
Обменный фонд, видимо, составляет небольшую часть населения муравейника. В муравейник северного лесного муравья с населением около 15 тыс, особей была внесена тысяча помеченных групповой меткой рабочих из отдаленной колонии. Пришельцы были приняты враждебно, часть из них была убита, но они все-таки остались в гнезде, влившись в различные функциональные группы местных рабочих. Через день в то же гнездо была выпущена еще тысяча рабочих из третьего, не связанного с первыми двумя, гнезда. Эти рабочие имели новую групповую метку. Их приняли возбужденно, ио схваток со смертельным исходом не было. Новые рабочие также влились в состав семьи. И, наконец, еще через день на купол подопытного гнезда выпустили очередную группу рабочих из четвертого, чужого всем предыдущим, гнезда, помеченных своей групповой меткой. На этот раз новички были приняты почти равнодушно. Предыдущие массовые введения новых качеств настолько расшатали монолитность небольшой муравьиной семьи, что она оказалась не в состоянии отличить чужих от своих. Число введенных особей оказалось большим, чем обменные возможности семьи, и она на какое-то время утратила индивидуальность.
Для контроля рабочих из четвертого гнезда выпускали на муравейник из одной колонии с подопытным. Здесь их приняли враждебно. Временная утрата индивидуальности происходит и при формировании искусственной колонии.
Формирование группировок отводков связано здесь с такими массовыми переходами и обменами, что некоторое время после этого уходит на стабилизацию семей. Нестабилизированные семьи легко, без посредства промежуточных почек, объединяются в колонии. В утрате индивидуальности, видимо, еще одна из основных причин поглощения слишком слабых дочерних семей материнскими.
Мы рассмотрели ряд сторон взаимодействия муравейников, входящих в одну колонию. И во всех случаях со всей олределеиностью проявляется неравноправность этих отношений. Материнская семья выступает как доминант. Доминирование материнской семьи вытекает из самого способа возникновения колонии - почкования. Дочерняя семья, поначалу несравнимо меньшая, может противопоставить этому только свой относительно быстрый рост.
Казалось бы, что с усилением отводка положение может измениться: он вырастет и в колонии установятся отношения, сходные с отношениями колонн в муравейнике. А поскольку отводок может и перегнать материнский муравейник по численности населения, не исключена возможность и смены доминанта. Но этого, как мы видели, не происходит. Обмены между семьями начинают угасать, и накануне выравнивания сил двух муравейников колония распадается. Семьи становятся друг другу чужими, между ними теперь могут начаться даже войны. Ни до координационной системы, ни тем более до смены доминаятов дело не доходит. Впрочем, в некоторых случаях, когда в результате разрушесния материнский муравейник резко слабеет, часть его населения может перебраться в сохранивщийся отводок. Тогда оставшаяся в материнском гнезде община может попасть в зависимое положепяие. Но такая ситуация и крайне редкая и неестественная. Сам же принцип возникновения колонии и характер отношеиий входящих в ее состав муравейников показывают: колония - система иерархическая, в которой обязательно доминирует материнская семья.
Основа ее доминирования - большая масса этой семьи. Именно соотношение масс и определяет в рамках вида исход любого события в жизни муравейников.
Но неужели разрыв выравнявшихся по силе муравейников неизбежен? Ведь в его основе, насколько сегодня можно судить, лежит все-таки не столько само равенство, сколько стремительно нарастающее в процессе выравнивания семей расхождение их по фенотипу. Муравьям становится все труднее признавать друг друга своими. И отчуждение происходит из-за того, что в каждом гнезде, прежде всего в отводке, слишком быстро накапливается новое качество. Столь быстро, что обмены не в состоянии сделать его общим достоянием, выравиять семьи по этому признаку до нужного уровня.
А если накопление различий идет так медленно, что обмены в состоянии полностью компенсировать их? Тогда колония сможет существовать неопределенно долго. Теоретически такой вариант возможен. А практически? Длительные наблюдения за многочисленными гнездами рыжих лесных муравьев показывают, что иногда данная тенденция проявляется, причем скорее в тех случаях, когда кормовые и территориальные возможности биотопов не допускают быстрого роста муравейников. Процесс отделения отводка может затянуться на 10 и более лет. Однако в конечном счете разрыв связей происходит, и дочерний муравейник становится самостоятельным. Выход отводка из колонии, по-видимому, правильно трактовать именно так, поскольку обмены объективно направлены на удержание отводка в составе колонии, противодействуют отделению дочерней семьи. Оно происходит не при достижении какого-то определенного, специфичного для вида уровня численности, а после приближения размеров отводка к размерам материнского муравейника.
Чем больше материнский муравейник, тем крупнее должен быть и отводок к моменту отделения. Это обстоятельство очень важно для оценки почкования как способа образования новых семей у муравьев. Естественный отводок в состоянии выделить только взрослый, развитый муравейник. И выйти из-под контроля такого муравейника может муравейник, ставший взрослым и развитым. Чем сильнее материнская семья, тем сильнее, а значит жизнеспособнее будет и отделившийся отводок. Если только позволяет территориальная обстановка, многие отделившиеся отводки в последующие один-два года уже становятся материнскими и выделяют отводки.
Пока что мы говорили о колониях, состоящих всего из двух жилых муравейников, материнского и одного отводка. У видов с малочисленными семьями обычно так и бывает: на формирование нескольких отводков одновременно у материнской семьи нет сил. Но для семей-миллионеров, таких, как у рыжих лесных муравьев, реально образование и удержание с помощью обменов сразу нескольких отводков.
СТРУКТУРА КОЛОНИИ
Если у материнского муравейника несколько отводков, они должны быть размещены по какому-то принципу. Этот принцип, как и само образование каждого отводка, должен вытекать из структуры материнской семьи, т. е. любое новое почкование - все тот же способ выравнивания ее колонн, но повторенный несколько раз. Очевидно, что в ряде случаев выделения одной дочерней семьи недостаточно для выравнивания колонн. Тогда отводки должны располагаться на одной дороге материнского муравейника.
Именно так обычно и обстоит дело. Вдоль наиболее сильной дороги материнского гнезда располагаются серии отводков. Помимо отводков в серию могут входить и кормовые почки. Однако у муравейника несоответствие в размерах колонн может складываться не только за счет одной из них. Например, из-за ограничения кормового участка с двух сторон у пятиколонного муравейника две колонны будут постоянны и минимальны по размерам. А три другие, ничем не ограниченные, имеют возможность быстро развиваться. В этих условиях выравнивание происходит за счет всех трех развивающихся колонн. Каждая из них в той или иной последовательности образует дочерние семьи. Тогда вдоль дорог муравейника возникает несколько серий отводков и вспомогательных муравейников. Функциональная схема такой колонии, расположенной в сосняке-зеленомошнике (Приокско-террасный заповедник), дается на рис. 13.
У муравейника Р-49 семь отводков и пять кормовых почек, распределенных по трем дорогам. Отводки разновозрастные и отличаются как но размерам, так и по характеру связи с материнским гнездом. Расположение отводков показывает последовательность их возникновения в пределах одной серии.
Сначала образовались более удаленные отводки 0-3 и 0-6. Они уже настолько крупны, что обменов особями между ними и материнской семьей нет, хотя связующие дороги еще сохраняются. Позже были сооружены гнезда 0-2, 0-4 и 0-7, Они мельче первых и еще обмениваются с Р-49 особями, хотя интенсивность обменов невелика - до 20 переносов в 10 минут.
Отводок 0-5 едва достиг размеров минимального автономного гнезда. Сюда ведет широкая дорога с интенсивными обменами. Наконец, самый маленький - 0-1 находится еще на этапе формирования.
Специфика серийной структуры колонии заключается в том, что каждый отводок, независимо от расстояния и количества гнезд в серии непосредственно связан с материнским муравейником. Это видно и из приведенной схемы колонии. Обмен между отводками одной серии в принципе возможен, особенно при ослаблении их связей с материнским муравейником.
Однажды в колонии волосистого лесного муравья удалось даже наблюдать редкий вариант: отводок, расположенный ближе к материнскому муравейнику, в силу различных причин обогнал в росте более старый и удаленный отводок, расположенный в той же серии. В течение двух лет он отделился от исходной колонии, «прихватив» с собой и отставшего в росте родственника. Отстающий оказался в роли отводка стремительно развившегося молодого муравейника.
А вот муравейники, расположенные на разных дорогах, не могут обмениваться непосредственно друг с другом. Это невозможно даже для гнезд, расположенных на различных ответвлениях одной дороги, например, для отводков 0-1 и 0-2. Такое положение вытекает из схемы работы муравьев-носильщиков, приведенной на рис.12. Обмен между сериями отводков производится только через метрополию, опосредованно. Мы застали колонию Р-49 в момент, предшествующий отделению первых отводков, но пока все еще «в сборе». Естественно, чтобы доминировать в такой многосемейной колонии, материнский муравейник должен быть очень сильным и иметь огромный обменный фонд. Ориентировочная численность семьи Р-49 - около 2,5 млн. особей. В день эта семья передавала на обмен в отводки около 9 тыс. особей, включая личинок, куколок и внутригнездовых рабочих. Данная цифра, по-видимому, максимальна, поскольку сюда входят и переносы во вновь создаваемый отводок 0-1. Но, с другой стороны, ясно, что в близком к зарегистрированному нами режиму муравейник находится уже несколько лет. В отводках 0-3 и 0-6 уже сформировались по три колонны; эти муравейники (1,1-1,2 м в диаметре купола) таковы, что могут сами образовывать отводки. Подобные случаи иногда наблюдаются в природе.
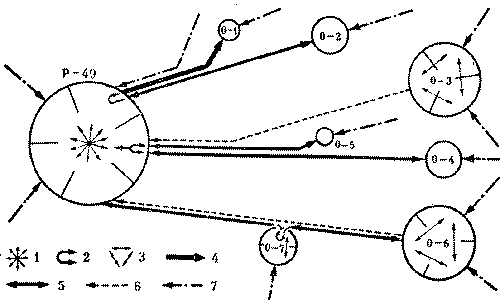
Рис. 13. Схема обменов в колонии обыкновенного муравья (фрагмент) Р-49 - материнский муравейник; 0-3, 0-6, 0-7 - дочерние муравейники с тремя и двумя колоннами; 0-1, 0-2, 0-4, 0-5 - дочерние муравейники с одной колонной; 1 - многосторонние обмены между колоннами материнского муравейника, 2 - эстафетный обмен особями между муравейниками одной серии, 3 - обмены особями между колоннами отводков, 4 - односторонний перенос особей из материнского гнезда в отводок, 5 - двухсторонний обмен особями, 6 - перенос пищи из отводка в материнский муравейник, 7 - поступление пищи с кормовых дорог
Наконец, логично предположить, что крупный материнский муравейник имеет меньшую пропускную способность при обменах между разными сериями в колонии, а следовательно, чем крупнее материнское гнездо, тем меньше родство дочерних семей, относящихся к разным сериям.
Степень родства не зависит от расстояния между гнездами. Размещение отводков и кормовых почек определяется рядом причин. Прежде всего - это расположение и плотность первичных, уже существующих гнезд и кормовых почек. Первые отводки, если нет иных ограничений, распределяются довольно произвольно, часто на самой границе или даже вне охраняемой территории материнского гнезда. Процесс почкования, таким образом, несет еще и функцию расселения, освоения муравьями новых земель. На размещении отводков сказывается наличие конкурирующих муравейников того же или других видов муравьев. В условиях острой территориальной конкуренции серии отводков, кормовые и обменные дороги располагаются параллельно друг другу, что показано на схеме колонии Р-49.
Если территориальной конкуренции нет, дочерняя семья может уйти так далеко от материнского гнезда, что кормовые участки двух муравейников оказываются пространственно разобщенными, т.е. здесь наблюдается прерывистость территории. В изображенной на рис. 14 колонии кормовые участки двух семей северного лесного муравья с самого начала отдалены друг от друга и связаны лишь полосой обменной дороги, которая не имеет других функций. Дочерний муравейник, созданный на небольшом плацдарме вне кормового участка материнского гнезда, в дальнейшем сам осваивает новую территорию. Следовательно, в этом случае образование отводка связано с расселением вида и не ущемляет в дальнейшем интересы материнской семьи. В то же время прерывистость территорий способствует, вероятно, более быстрому отчуждению дочерней семьи.
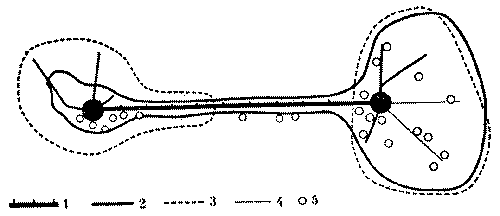
Рис. 14. Развитие колонии северного лесного муравья при отсутствии территориальной конкуренции
1 - обменно-кормовая дорога в 1966 г., 2 - границы охраняемых территорий в августе 1966 г., 3 - границы охраняемых территорий в августе 1967 г., 4 - кормовые дороги, 5 - деревья с колониями тлей
В тех случаях, когда формирование отводков происходит в условиях территориальных ограничений, материнская семья вынуждена создавать отводки на окраинах своего кормового участка. Пока дочерний муравейник мал и не имеет собственной охраняемой территории, его существование не сказывается на экономике материнской семьи. Однако в дальнейшем он начинает вытеснять материнский муравейник из кормовых угодий, прилегающих к отводку. В результате кормовая база ведущей ранее колонны резко сокращается, а наиболее сильной оказывается следующая по численности. Выделенные ею отводки тоже со временем отделяются, «прихватив» куски территории материнского гнезда. После этого наступает черед следующей колонии.
В классическом варианте подобная участь по очереди настигает все колонны, каждая из них теряет часть своего кормового участка. В итоге кормовой участок семьи выравнивается по наименьшей дороге. Процесс выравнивания продолжается до тех пор, пока в распоряжении материнского муравейника не остается совсем маленькая территория, пищевые ресурсы которой не позволяют формирования отводков вообще.
Примерно такую ситуацию мы имеем в колонии волосистого лесного муравья (рис. 15). За 10 лет (1966-1976) в результате поочередного изъятия все новых кусков исходной территории вырастающими отводками материнский муравейник оказался зажатым буквально на пятачке радиусом 15-18 м. Все возможности дальнейшего роста материнской семьи исчерпаны. Теперь, если ситуация радикально не изменится, например в результате случайной гибели одного из отводков, материнская семья обречена. Ограниченные ресурсы оставшегося в ее распоряжении участка недостаточны даже для поддержания населения на имеющемся уровне численности, а, неизбежное уменьшение населения повлечет за собой дальнейшее сокращение участка. Семья оказывается в тисках, которые постепенно сжимаются.
Что же происходит? Продуцируя отводки, муравейник в конечном счете обрекает себя на вырождение, гибель?
Именно так. При многолетних наблюдениях за одними и теми же комплексами гнезд приходится регистрировать этот неумолимый процесс. Процветавшие десятилетие назад муравейники на глазах утрачивают свою былую силу. Вокруг некоторых из них уже сомкнулись границы новых, ими же выделенных когда-то отводков.
Впрочем, положение преемников весьма сложно: они сразу же попали в условия жесткой территориальной конкуренции. Им обычно негде развернуться так, как когда-то это делали обитатели первых гнезд комплекса. Такая ситуация характерна для центральной части комплекса, а по периферии вырастают все новые мощные муравейники, и продолжается освоение еще не захваченных этим видом свободных территорий. До тех пор, пока все пригодные для существования угодья не будут заняты или на пути не появятся владения конкурирующих муравьев, непреодолимые преграды и т. п.
В центральной же части комплекса положение постепенно стабилизируется. Муравейники выравниваются по размерам гнезд и площади кормовых участков. Между соседними семьями устанавливается напряженное динамическое равновесие. В этих условиях всякое ослабление семьи вызывает быстрый натиск соседей, передел территории. С уменьшением одного муравейника увеличивается другой. Переделы территорий нередко сопровождаются войнами, истощающими конкурентов. Радикально решая насущные, сиюминутные задачи по сохранению семьи, муравьи не в состоянии предвидеть и оценить отдаленные последствия почкования для собственной семьи. Да собственно это чисто риторическая постановка вопроса, ибо представить ситуацию в целом некому: ни один из муравьев на это не способен, а специальный координирующий действия колонн центр в семье отсутствует. Согласование, достигаемое почти механическим взаимодействием многотысячных масс муравьев, не позволяет подняться до уровня прогнозирования событий. Перспектива для материнского муравейника оказывается неутешительной. Для отдельного муравейника, но не для популяции, которой обеспечены не только быстрое и эффективное распространение по территории, но и стабильный уровень численности при максимальном использовании ресурсов занимаемой территории.
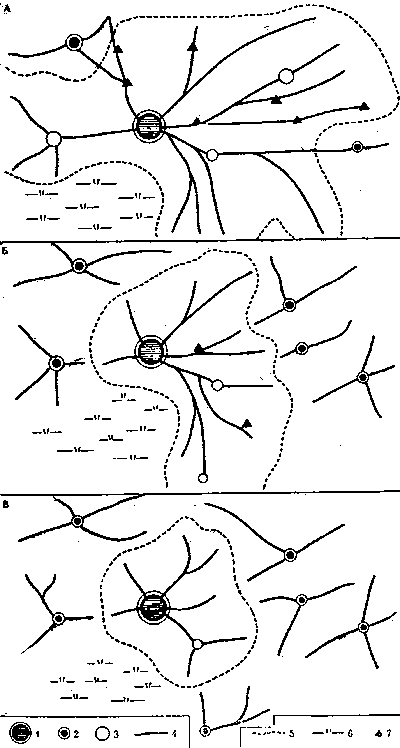
Рис. 15. Сокращение кормового участка материнского муравейника в результате образования и отделения отводков
Положение в колонии в различные годы: А - 1966 г.; Б -1970 г.; В - 1976 г. 1 -материнский муравейник, 2 - отводки, отделившиеся или имеющие охраняемую территорию, 3 - отводки без охраняемой территории, 4 - дороги, 5 - границы кормового участка материнского муравейника, 6 - участок, непригодный для обитания муравьев, 7 - кормовые почки
Все это ожидает материнскую семью впереди. А пока она доминирует в колонии, представляющей из себя семью муравейников: мать и дети. Материнская семья, продуцируя дочерние, сохраняет свою целостность, свою индивидуальность. Каждая дочерняя со временем приобретает свою индивидуальность, в чем-то неповторимую так же, как неповторимо гнездо, в котором она обитает.
Если мы сравним гнезда рыжих лесных муравьев разного возраста, то заметим интересную закономерность. Гнезда-первогодки одного вида удивительно похожи друг на друга. Примерно так же, как похожи суточные цыплята. Форма этих муравейничков фактически еще не отражает ни особенностей организации каждого из них, ни условий обитания, в которые попала юная семья. В архитектонике такого гнезда в зачаточной форме заложены все необходимые для последующего развития муравейника элементы, специфичные для этого вида. Конечно, некоторые факторы среды сказываются еще в момент закладки муравейника. Скажем, если гнездо строится на большом пне, то габариты последнего неизбежно отразятся на объеме и габитусе муравейника. Но это особый случай, не отвергающий самого принципа начального стереотипа гнездостроения, проявляющегося на первых порах существования муравейника. В дальнейшем в зависимости от особенностей организации и состояния семьи и условий ее обитания изменяется форма гнезда и соотношение его отдельных элементов. В результате, обследовав тысячи взрослых муравейников, мы не сможем найти двух одинаковых.
Еще оригинальнее по составу, организации и другим показателям обитающие в этих гнездах семьи. Конечно, изучение индивидуальных особенностей муравейников дело многотрудное, да и острой необходимости в этом нет. Но даже сравнив некоторые, лежащие на поверхности характеристики, например структуру кормовых участков различных семей, мы придем к выводу о значительных отличиях в составе каждой из них.
Вопрос индивидуальности и целостности - один из центральных вопросов биологии. Говоря о муравьях с их сложной и развитой «социальностью», мы сталкиваемся не только с выраженной индивидуальностью каждой отдельной особи, но и целых сообществ. Причем индивидуальность семьи или входящей в состав семьи колонны не имеет ничего общего с индивидуальностью организма, ибо базируется целиком на поведенческом взаимодействии составляющих общину индивидов. Сообщество муравьев или других общественных насекомых не теряет специфики как сообщества даже тогда, когда в результате развития внутрисемейных структур оно выходит на уровень, допускающий его деление.
В принципе образование колонии представляет собой растянутый во времени процесс деления муравейника, сохраняющего при этом целостность и индивидуальность. Все это очень наглядно проявляется в упрощенном варианте, когда колония состоит из двух семей - материнской и одного отводка, как, например, показано на рис. 14. Однако мы знаем, что если отпочкование одной дочерней семьи не приводит систему колонн в устойчивое (равновесное) состояние, образуется вторая, третья и последующие дочерние семьи. Время замедляет свой бег, зато отчетливо слышны внутренние часы колонии: спешащие стрелки отводков и сдерживающий их маятник материнского муравейника. В этом тоже проявляется индивидуальность семей. У каждой своя задача: дочерние семьи стремятся подняться к оптимуму, материнская - удержаться в оптимальном режиме.
Здесь уместно вспомнить о диффузных муравейниках. Почему применительно к ним мы не употребляем термин «колония»? Ведь и там несколько гнезд, хоть и маленьких, связаны обменными дорогами. Бывает, что связь между одним из таких гнезд и остальными обрывается. И тогда, что очень важно, этот обособившийся муравейничек не погибнет, а начнет развиваться как самостоятельный: в нем, как и в остальных, есть яйцекладущие самки. Чем не почкование?
Да, действительно, внешнее сходство имеется, но только внешнее. Ведь в микрогнездах диффузного муравейника нет главного - целостности и индивидуальности. Население этих гнездышек случайно и непостоянно. Оно, как живая ртуть, перетекает из убежища в убежище. Единственным проявлением целостности здесь является целостность всей семьи. В этих условиях речь идет не о выделении части семьи - части, обеспеченной полным составом и достаточной для самостоятельного существования численностью, а о потере, случайном отщеплении группы особей. Правда, эта группа живуча и начинает вести себя как целая семья. Но и произвольно взятая часть любой колонны, отделенная от муравейника и перенесенная на место, где нет муравьев, способна реорганизоваться и имеет шанс развиваться во взрослый муравейник. Именно эта возможность муравьев к подобной реорганизации и лежит в основе всех методов переселения полезных видов муравьев искусственными отводками. Были бы в таком отводке самки и достаточное число рабочих.
Иногда поговаривают о способности семьи к регенерации при гибели части особей, проводя при этом аналогию с некоторыми растительными или животными организмами. Однако ни о какой регенерации здесь не может быть и речи. С одинаковым успехом можно обсуждать регенерацию средневекового города после чумы. В действительности семья, потерявшая по какой-либо причине (например, взят искусственный отводок) значительную часть особей, со временем восстанавливает частично или полностью свою численность, как восстанавливает ее популяция после депрессии. Искусственно переселенный отводок начинает развиваться на новом месте как совершенно новое образование. И не стоит думать, что если он взят из пятиколонного муравейника, то вырастет именно в пятяколонный.
Итак, колония - образование временное и наделенное определенными функциями. Основной функцией колонии является выращивание новых муравейников. Серийная структура колонии выросла из организационной структуры материнского муравейника. Реальная структура каждой колонии на любом этапе ее существования продолжает отражать прежде всего состояние и структуру материнского муравейника. Значение колонии как эффективного пути размножения семей и расселения муравьев трудно переоценивать. Именно благодаря колониальности ряд видов получил возможность заселять огромные территории и поддерживать стабильную высокую плотность поселения.
КОЛОНИЯ И ФЕДЕРАЦИЯ
Казалось бы, образование колоний, тем более существующих длительное время, - это последняя вершина организационной структуры муравьиного сообщества. Однако, как уже говорилось в предыдущем разделе, колония, как и все прочие организационные достижения муравьев, небезупречна. Длительный процесс образования отводков ведет в конечном счете к перенаселению, а это очень опасно, так как влечет за собой вырождение.
Возможность перенаселения - косвенный результат одного из существенных недостатков иерархического принципа организации колонии. В самой колонии, так же как и в семье, численность особей регулируется в соответствии с пищевыми ресурсами участка. В семьях рабочие муравьи поедают лишних личинок при недостатке пищи. Избыточное число муравьев выселяется в отводки. Рост отводков сдерживается неравными обменами, а рост материнской семьи - последовательным сокращением ее кормового участка. Пока все семьи в колонии, перенаселения не будет.
Но все-таки тенденции к росту отводков столь сильны, что они постепенно высвобождаются из-под регулирующего влияния колонии. Вот тут-то и возникает опасность перенаселения. Инициатива принадлежит одиночным, вышедшим из колоний муравейникам. Каналов регуляции для них стало меньше, а возможностей использования кормового участка больше. Теперь вся без исключения добыча остается в таком гнезде. И если общая численность гнезд в комплексе достаточно велика, продолжающий свой рост отводок быстро достигает предела, обеспечиваемого ресурсами его кормового участка.
Одиночные муравейники являются стихийными элементами, они начинают теснить соседей, захватывая часть их кормовых участков. Начинаются истребительные войны, основным итогом которых, как правило, бывает лишь гибель множества муравьев. Это тоже регуляция численности, но производится она явно нерациональным путем.
Происходит все потому, что выросший отводок должен отделиться от колонии, чтобы не нарушать незыблемого принципа доминирования материнского муравейника. Он должен выпасть из системы надсемейной регуляции численности. Да и сама система обменов в колонии довольно несовершенна. Именно в тот момент, когда регуляция важнее всего, интенсивность обменов стремительно падает. Масштабы колонии, даже самой крупной, тоже недостаточны. Впрочем большего от колонии требовать трудно, ведь ее регуляторные возможности сводятся к возможностям лишь одного материнского муравейника. Таким образом, сам принцип иерархии семей в колонии ограничивает размеры такой системы муравейников и во времени, и по числу входящих в систему семей. Следовательно, для дальнейшего расширения границ системы регуляции численности принцип не пригоден. Но отказаться от него невозможно, ибо колония уже есть, и с ней у ряда видов связана сама возможность появления новых муравейников.
Тем не менее выход есть. Он заключается в новом этапе развития надсемейных структур у муравьев. На этом этапе возникают объединения колоний. Наиболее ярко проявляются такие объединения у северного лесного муравья, ставшего благодаря этому одним из наиболее многочисленных видов наших лесов. Система взаимосвязанных колоний - федерация* возникает именно в условиях перенаселения, нехватки охотничьих угодий.
В федерации каждая колония сохраняет свою структуру с доминированием материнского муравейника. Более того, обмены внутри каждой колонии становятся интенсивнее, при этом интенсивность в меньшей степени, чем в отдельной колонии, зависит от соотношения сил муравейников. Это означает, что родственные связи внутри колоний, входящих в федерацию, более прочные и стабильные, а возможности отделения отводков соответственно меньше. Но самое важное - это принцип объединения колоний. Оно должно быть таким, чтобы не затрагивало внутренней структуры составляющих единиц, не разрушало принципа колониальности. Иначе вместо нескольких колоний возникнет одна, состоящая из многих муравейников. А чем больше гнезд в колонии, тем она эфемернее.
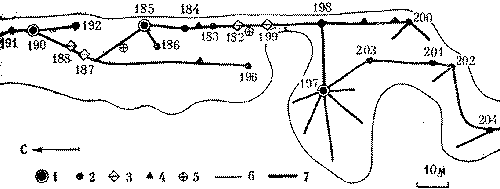
Рис. 16. Федерация муравейников северного лесного муравья (фрагмент)
182-204 - номера гнезд; 1 - гнезда-доминанты, 2 - жилые муравейники, 3 - буферные гнезда, 4 - кормовые почки, 5 - брошенные гнезда, 6 - границы пригодного для использования муравьями участка, 7 - муравьиные дороги
Основная угроза устойчивости федерации заключается в том, что среди материнских семей, вступивших в контакт, выделится сильнейшая, которая и станет доминантой по отношению ко всем, входящим в федерацию семьям. Поэтому прямого противостояния материнских семей надо избегать. Это достигается весьма простым способом: обменная связь материнских муравейников устанавливается только через отводки или специально сооружаемые промежуточные почки (как при породнении чужих гнезд в колонии). Но здесь промежуточные почки действуют постоянно. На рис. 16 показан фрагмент федерации северного лесного муравья, состоящий из 19 жилых муравейников. Видно, как из трех ранее обособленных колоний образовалась федерация. Первая колония включала до объединения четыре муравейника: 190 (материнский), 191, 192, 196 (отводки). Вторая колония состояла из четырех муравейников: 185 (материнский), 183, 184 и 186 (отводки). Третья колония наиболее мощная. Здесь имеется первичный материнский муравейник 197, один вторичный материнский муравейник 198 с отводком 200 и отводки муравейника 197-201, 202, 203 и 204, расположенные на одной дороге этого гнезда. Дорога изгибается, обходя не пригодные для муравьев слишком сырые участки. Принцип соединения колоний внутри федерации показан на рис. 17.
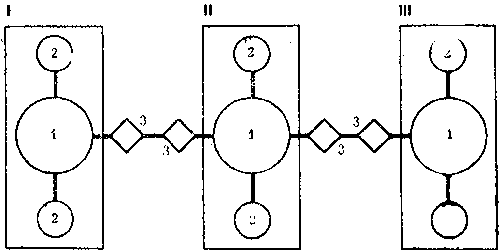
Рис. 17. Блок-схема федерации северного лесного муравья
I, II, III - колонии, входящие в состав федерации. 1 - материнские муравейники, 2 - отводки, 3 - буферные гнезда
Появление вторичных материнских муравейников симптоматично для федерации. Оно показывает, что даже очень крупные отводки, которые в состоянии сами образовывать дочерние муравейники, при интенсивных обменах удерживаются в составе колоний. В нашем примере муравейник 197 имеет диаметр купола 2,2 м, а его отводок 198 - 1,9 м. Обе величины предельны для данного вида. В сравнении с этими гнездами два других материнских муравейника выглядят карликами (диаметр купола гнезда 190 равен 1,2 м, гнезда 185 - 1,3 м). Тем не менее никакой иерархии колоний нет, так как при объединении между ними были сооружены буферные гнезда* - постоянные промежуточные почки. Буфером между колониями 1 и 2 служат муравейники 187-188, между колониями 2 и 3 - 182 и 199.
Объединение колоний, по-видимому, не обошлось без предварительных столкновений на территории. Об этом свидетельствуют имеющиеся в пограничных зонах колоний следы старых брошенных небольших гнезд, что обычно указывает на острую территориальную конкуренцию (см. рисунок). Однако теперь сражения за территорию в прошлом, все муравейники оказались связанными общей цепью обменов. Их интенсивность очень высока во всех звеньях федерации. Интенсивность переноски особей на основных дорогах, связывающих гнезда-доминанты, достигают 40 и белее в минуту (до 200). На остальных дорогах интенсивность - 15-30 переносов в минуту, что соответствует максимальным величинам в отдельных колониях. В колонии обмены в 40 и более переносов в минуту отмечены только в период формирования отводка.
Все обмены осуществляются через промежуточные почки.
Называть эти гнезда промежуточными почками можно только по аналогии. Фактически буферные гнезда - это особый тип летних гнезд, выполняющих «федеральные функции». Здесь имеется постоянное, частично смешанное население, среди которого высока доля муравьев-носильщиков. Благодаря этим муравейникам становится возможным эстафетный обмен между материнскими семьями. В обменных операциях участвуют десятки тысяч особей. Часть из них занята поддержанием в порядке буферных гнезд, другие осуществляют переносы.
Каким образом осуществляется запуск этого мощного механизма интеграции, неизвестно. С точки зрения экономики отдельных семей столь интенсивные обмены, по-видимому, мало рациональны. Но тем не менее это происходит, в виде федерации панмуравейник получает свое приближенное воплощение. Если в отдельной семье или колонии социальная регуляция колонн или входящих в колонию муравейников возникает как механизм, обеспечивающий общий рост населения, то в федерации направленность регуляции меняется. Здесь единственная функция обменных операций - ограничение общей численности муравьев в системе. Результатом действий регуляторных механизмов в семье и колонии является увеличение числа подсистем (колонн, семей), расселение вида; результатом регуляции в федерации - сохранение основных структурных единиц. О расселении речь не идет, поскольку федерации возникают в условиях, когда расселяться муравьям уже некуда.
И все это достигается одними и теми же средствами - обменами. Чем жестче территориальная конкуренция, тем интенсивнее обмены.
Данный принцип оправдан не только в федерациях. В эксперименте, проведенном на колонии муравьев-фаэтончиков в Туркмении в 1972 г., искусственное сокращение кормовых участков материнского и дочернего муравейников вызвало увеличение интенсивности обменов в 10-15 раз. Одновременно у подопытных муравейников появилась охраняемая территория, не свойственная фаэтончикам в обычных условиях.
Очень четкое разделение охраняемых территорий всех муравейников наблюдается в федерации. Такая взаимосвязь двух весьма различных по сути явлений понятна, так как оба они вытекают из объективной необходимости более эффективной регуляции в условиях жестких территориальных ограничений.
Сочетание четкого разграничения кормовых участков с интенсивными обменами обеспечивает стабильность федерации как системы муравейников. Устойчивость федерации значительно выше, чем обособленной колонии. Хотя разрыв отдельных связей между колониями возможен, тут же возникают новые, выполняющие ту же функцию. Сами же муравейники почти не меняются. За пять лет наблюдений в рассматриваемой федерации северного лесного муравья не погибло ни одного желтого муравейника и не возникло ни одного нового. Соотношение гнезд по размерам сохранилось приблизительно одинаковое, чего не бывает столь продолжительное время в самостоятельной колонии. Фактически неизменными по интенсивности остались и обмены, производимые муравьями летом.
Наименее устойчивыми элементами в федерации оказываются межколониальные обменные дороги и связанные с ними буферные гнезда. Они могут сдвигаться на местности, но сохраняют при этом полную функциональную преемственность. Таким образом в федерации реализуется стратегическая задача: обеспечение регулирования исключительно поведенческими средствами плотности поселения вида. Регуляция и устойчивость при максимальной для данных условий плотности поселения популяции. Такая задача в мире насекомых оказалась под силу только муравьям, точнее - лишь некоторым их видам. Все муравейники, объединенные в находящиеся под наблюдением федерации, активны, а гнезда - в хорошем состоянии. Почему? Ведь вся система функционирует в напряженном режиме (об этом можно судить хотя бы по интенсивности обменов). Может быть, следует сделать допущение, что именно такой режим и является лучшим средством, предохраняющим муравейники от старения и вырождения. Точнее, единственным шансом сохранить активное состояние в условиях перенаселения. Мы можем проверить это предположение, ознакомившись с развитием муравейников тех видов, у которых есть колонии, но не отмечены федерации. Например, волосистого лесного муравья. Гнезда у этого вида, как правило, крупнее гнезд северного лесного муравья, обычно обилие кормовых почек и отводков, но федерации нет. Собственно и не важно, может ли волосистый муравей создать федерацию в принципе. Достаточно взять какой-либо крупный комплекс, в котором федераций нет, а плотность поселения угрожающе высока. Ведь всякая возможность реализуется не всегда. В частности, старый муравейник, стабилизировавшийся в своем одиночном существовании, не дает отводков и не войдет в федерацию. Даже если это будет муравейник столь склонного к федерациям северного лесного муравья.
Весьма обычен комплекс муравейников, «законсервировавшихся» раньше, чем сложились условия перенасе ления, подталкивающие муравьев к объединению в федерации. Но за счет отдельных, разбросанных по комплексу колоний плотность поселения все-таки возрастает и достигает критического уровня. Территориальная конкуренция обостряется, разыгрываются ожесточенные сражения между соседними муравейниками, но за этими сражениями не следует объединение. Периодические сражения - плохой регулятор численности. Вскоре из-за нехватки белковой пищи изменяется пищевой баланс семей (возрастает доля пади), сокращается количество выкармливаемой молоди. Теперь уже не до отводков.
Семьи становятся все нетерпимее к чужим особям, в частности к чужим самкам. Замена старых самок молодыми затрудняется. Семьи приходят в упадок, погибают. От комплекса остаются отдельные, едва влачащие существование муравейники или периферийное кольцо муравейников, из которых со временем начнется повторное заселение центральной части комплекса.
Значит, одной системы охраняемых территорий еще недостаточно, чтобы поддерживать комплекс в активном состоянии долгое время. В комплексе остается стихийное начало, ведущее к подъемам и депрессиям, подобным зигзагам движения численности у одиночных насекомых. Конечно, амплитуды колебаний здесь не столь значительны, да и катастрофические спады случаются очень редко. Но все же время от времени они происходят.
А где-то рядом существуют вполне благополучные федерации.
Следовательно, тенденция к стабилизации, проявляющаяся у муравьев на всех уровнях организации, становится, несомненно, положительной только в тех случаях, когда она связана и со стабильной активностью муравейника. Лишь тогда она в состоянии отодвигать границы допустимой плотности поселения без опасности последующего вырождения.
Надо сказать, что для абсолютного большинства видов насекомых эта задача, по-видимому, вообще невыполнима. Поэтому мы и наблюдаем непрерывные колебания численности различных массовых вредителей и менее массовых видов, не причисленных по этой причине к вредным. Только появление достаточно эффективных способов социальной регуляции допускают стабильность. И чем совершеннее социальная организация вида, тем выше уровень, на котором стабильность сохраняется. Сколь же высок такой уровень для муравьев, добравшихся в ходе эволюции до уровня колоний и федераций?
В комплексах муравейников северного лесного муравья, еще далеких от верхнего предела численности и образования федераций, в среднем на 1 га обитает 5-7 млн. муравьев. Плотность поселения того же вида рассмотренной федерации превышает 15 млн. взрослых особей на 1 га. Весит население такой федерации более 120-130 кг. Это вес стационарный, т. е. постоянно поддерживаемый прп имеющемся довольно высоком уровне смертности на участке.
Эти цифры не идут ни в какое сравнение с соответствующими среднегодовыми величинами для любых других насекомых.
Федерация, пожалуй, последнее, высшее достижение социальной организации у муравьев. Прослеживая восхождение муравьев от одиночной (одинарной) семьи к федерации, мы можем наблюдать и параллельное развитие основного средства, обеспечивающего существование организационных структур как внутрисемейного, так и надсемейного ранга. Перенос особей, изначально связанный только с уходом за молодью в примитивных гнездах, а позднее - с переселением при смене гнезд, в дальнейшем вылился в мощное средство, без которого развитие социальности у муравьев трудно представить.
Сначала обмены открыли возможность возникновения в семье колонн. Затем, преодолев зачаровавший натуралистов отрицательный фототаксис, потоки муравьев-носильщиков вынесли молодь из гнезда и проложили путь к колонии. И наконец, интенсификация обменов позволила решить проблему «несовместимости» колоний и создания федерации.
В заключение данной главы следует отметить, что колониальность у муравьев изучается пока что на весьма ограниченном числе видов, относящихся в основном к различным подродам нескольких родов. Столь же ограничен круг поликалических видов, в той или иной мере изученных со структурных позиций. Степень изученности отдельных вопросов организации семьи, ее структуры, различна и не позволяет представить в настоящее время картины целиком даже у наиболее изученных в этом отношении муравьев. Отрывочные данные по структуре поселений других видов позволяют лишь предполагать степень сходства подобных явлений у других групп муравьев.
4. РАЗВИТИЕ ОБЩИННОГО ОБРАЗА ЖИЗНИ У МУРАВЬЕВ
Что вообще подразумевается под прогрессивным развитием той пли иной группы животных? Изучая этот вопрос, выдающийся советский биолог А.Н.Северцов сформировал два основных критерия биологического прогресса: рост общей численности вида и его широкое географическое распространение. Следовательно, все морфологические, физиологические, поведенческие и другие приспособления, ведущие к увеличению численности и расширению ареала вида, могут быть отнесены к прогрессивным.
Данные критерии в принципе справедливы для любых животных, однако использование их в общем виде не позволяет оценить значение отдельных факторов в эволюции ряда групп. И это в полной мере относится к муравьям. Ведь муравья ни в одном опыте, ни в одном теоретичееком построении практически невозможно оторвать от семьи, рассматривать вне ее. Семья, муравейник - вот основная среда для индивида. В борьбе за существование община выступает как единое целое. Независимо от способа основания общины она сразу же проявляется как целостная система, обладающая определенной степенью индивидуальности. Поэтому развитие муравьев как группы является процессом становления и развития семьи, а основным критерием биологического прогресса - рост численности поселения отдельной целостной системы. Но, как мы уже видели из предыдущих глав, увеличение поселения такой системы невозможно без усложнения ее организации. Оба процесса неотделимы друг от друга. Неотделим от них и биологический прогресс муравьев. Впрочем, не только муравьев. Во всех случаях, когда община становится единственной формой существования вида, биологический прогресс будет определяться этими же критериями, т.е. это положение в равной мере справедливо для термитов, общественных пчел и ос.
РОСТ СЕМЬИ И СОВЕРШЕНСТВОВАНИЕ ЕЕ ОРГАНИЗАЦИИ
Структура в известной мере является как бы результирующей различных аспектов жизни, характеристик семьи муравьев. В структуре находят достаточно полное выражение состав общины и ее численность, видовые особенности индивидуального поведения, стереотипы гнездостроения и способы использования кормового участка и т. д. Изменение любой из взаимосвязанных составляющих приводит к соответствующим перестроениям организации общины. Можно сказать, что ни одно из новоприобретений муравьев, не нашедшее отражения в структуре семьи, не в состоянии закрепиться и оказать сколько-нибудь серьезное воздействие на ее дальнейшую судьбу.
Возьмем, например, такой первоочередной показатель, как обеспеченность муравейника пищей. Одним из важнейших эволюционных достижений муравьев является трофобиоз - постоянные тесные взаимовыгодные отношения муравьев с выделяющими падь насекомыми (тлями, щитовками, пикадками и т. д.). Муравьи охраняют и даже расселяют этих насекомых, получая от них огромное количество углеводной пищи. Кормовая база муравьев стабилизировалась за счет стад тлей и цикадок, появилась возможность значительного роста численности.
Падь хорошо известна муравьям и большинству насекомых вообще. Падью периодически питаются наездники, различные мухи, пчелы, некоторые жуки и т. д.; известны сотни видов насекомых, собирающихся на падь в наших лесах. Но все это периодически, от случая к случаю. Эпизодическая разовая подкормка играет немалую роль в жизни одиночных насекомых, но для муравьев подобный подход несерьезен. Семья непрерывно функционирует в течение нескольких месяцев, существует годами и десятилетиями. Здесь значим только стабильный источник пищи. И выделяющие падь насекомые могут служить таким источником. Все дело за муравьями: они должны организовать службу по доставке пади и охране выделяющих ее насекомых.
Но постоянная служба связана с формированием постоянной функциональной группы - сборщиков пади. Нужно не просто сформировать, но и закрепить сборщиков пади за определенными колониями тлей, щитовок и т. д. Часть видов муравьев решила эту задачу и продвинулась благодаря этому вперед. Другим она оказалась не по плечу, например мирмикам. Они тоже пользуются тлями, но настоящих сборщиков пади у них не появилось, и тли не помогли мирмикам перейти к обитанию крупными семьями. Как не помогла и полигиния, в которой заложены тенденции к полицентрическим семьям, созданию колонн и т. д. Вместо этого - скоротечное деление семьи пополам и снова общины из 500-2000 особей.
Что же помешало мирмикам? Отсутствие развитой семейной структуры и вытекающая отсюда зыбкость общины как целого. А ведь среди достаточно близких этому роду муравьев есть виды, для которых трофобиоз стал ступенью для подъема на более высокий уровень численности. Но взойти на эту ступень смогли лишь те, структура семьи которых оказалась уже достаточно развитой для выделения особой функциональной группы - сборщиков пади.
Показательно, что стабилизация сборщиков пади как постоянной функциональной группы происходила параллельно с их территориальным закреплением за определенными колониями тлей, червецов и т. д. Эта тесная связь структуры семьи с пространственным распределением особей на кормовом участке примечательна для муравьев вообще.
По-видимому, задачи, возникающие при многоцелевом использовании кормовых угодий, всегда играли роль существенных факторов, формирующих структуру семьи. Взять к примеру тех же сборщиков пади. Для закрепления их территориальной привязанности определяющее значение имело исходное рассредоточение колоний выделяющих падь насекомых. Постоянство этих колоний привело к образованию устойчивых дорог к ним. Этими же дорогами пользуются и охотники. А связь дорог с возникновением колонн в семьях некоторых муравьев уже прослеживалась нами в разделе 2. Вспомним хотя бы, что разделение колонны на две связано с двумя основными обстоятельствами: несоразмерным увеличением населения одной из колонн и наличием у исходной дороги мощных ответвлений, без обособления которых раздвоение колонны невозможно.
Можно сделать вывод, что с развитием общинного образа жизни, усложнением организации семьи связь между структурами общины и кормового участка не ослабевает. Это означает, в частности, что для полной характеристики муравейников многих видов недостаточно одного, пусть даже очень точного описания гнезда. Необходима комплексная характеристика муравейника и используемой им территории, ее структуры. Глубина их взаимосвязи настолько велика, что некоторые мирмекологи, например Я.Добжанская, вообще склонны считать дороги частью гнезда.
Рост численности семьи - необходимое условие и для такого важного способа обеспечения общины пищей, как разведение грибных садов. Опять же многие виды более или менее регулярно питаются грибами. Поэтому и здесь задача сводится не к открытию грибов как типа пищи, а к переходу на их постоянное разведение. Но для этого потребовалось сооружение специализированных крупных гнезд с четкой дифференциацией их частей, а также поддержание строго определенного гигротермического режима в муравейнике. Ни одна из этих задач не могла быть успешно решена без многотысячных контингентов рабочих. Среди муравьев-листорезов, культивирующих грибницы, имеются виды, стоящие на различных ступенях использования грибов, и - соответственно - уровнях численности и сложности организации общины. Высокая «культура производства» у муравьев обязательно связана с большой численностью семьи и узкой специализацией индивидов. Отсюда вытекает важное условие для жизнеспособности общины: ее численность должна превышать определенный минимум, иначе муравейнику грозит гибель. Все это приходится постоянно учитывать и при исследовании жизни муравьев, и при расселении полезных видов, и при борьбе с вредными.
Вот пример из практики борьбы с муравьями-листорезами рода атта в штате Луизиана в США, где эти муравьи приносят огромный ущерб лесному хозяйству, уничтожая листву деревьев. При проведении химической борьбы было истреблено большинство особей во всех крупных гнездах. Небольшие, расположенные по периферии участка семьи пострадали меньше. Но фактически в каждом гнезде осталось столь малое число рабочих, что последующая гибель муравейников не вызывала сомнения. Однако именно в этих условиях у муравьев начинается концентрация особей для достижения хотя бы минимальных размеров жизнеспособной семьи. Листорезы поступили так же, как вели себя рыжие лесные муравьи в рассмотренных ранее примерах. Остатки семей собрались в наименее пострадавших муравейниках и оттуда спустя некоторое время начали повторное расселение по лесному массиву.
Подобная картина наблюдается и при борьбе с фараоновым муравьем. Из-за этого оказывается практически невозможным уничтожение вредных муравьев разовой, пусть даже весьма эффективной обработкой. Необходим контроль за концентрацией уцелевших особей с последующими обработками мест их сосредоточения. Знание особенностей организации общины и поведения муравьев в экстремальных условиях оказывается необходимым для успешной борьбы с вредными видами.
Возможности развития структуры муравьиной семьи связаны с рядом факторов. Среди них очень важное значение имеет количество самок (т. е. является ли вид моногинным или полигинным). Моногиния и полигиния оказались двумя принципиальными путями, берущими свое начало из одного исходного типа семьи с ограниченным числом индивидов и невыраженными поведенческой и организационной структурами. Такие семьи характерны для многих видов понерин и мирмециин - наиболее древних подсемейств муравьев. Число самок во многом определило возможности дальнейшего развития «социальности» у муравьев. Например, для моногинных видов невозможен выход в колонию, хотя все предшествующие этапы организации ими пройдены (образование колонн, поликалия).
Как полигинные, так и монотонные муравьи развивались по пути увеличения численности семьи и усложнения ее структуры. Но для моногинных имелся барьер - ограниченная плодовитость единственной самки. Барьер, которого для полигинных видов не существовало: они могли просто увеличить число самок. Радикальным выходом из этой ситуации оказалась высокая продуктивность самок у моногинных видов - физогастрия. Физогастрическая самка имеет гипертрофированные яичники, благодаря чему отличается огромной продуктивностью. Виды с физогастрическими самками могут образовывать многочисленные (десятки и сотни тысяч особей) семьи при сохранении обязательной моногинии. К видам с физогастрическими самками относится, например, пахучий муравей-древоточец (до 300 тыс. особей в семье).
Физогастрия имеется не только у муравьев. У высших термитов она получила еще большее развитие. В отдельных случаях брюшко самки термита увеличивается до таких размеров, что та даже не в состоянии самостоятельно передвигаться.
Таким образом моногиния не стала непреодолимым препятствием на пути роста семьи и усложнения ее организации, хотя и воспрепятствовала появлению надсемейных структур.
Но есть еще одна принципиальная разница между семьями с одной и многими самками. И те, и другие семьи многолетние, но для большинства моногинных видов срок существования семьи определяется продолжительностью жизни самки. Погибает самка, и через некоторое время вымирает семья. И хотя самки муравьев живут очень долго (до 15-20 лет), время существования семьи достаточно жестко ограничено. Правда, как уже говорилось, некоторые моногинные виды все же могут оставить в семье одну из выращенных молодых самок. Но такие сведения имеются по очень небольшому числу видов с обязательной моногинией. Если же в семье несколько самок, муравьи могут постепенно обновлять их состав. Продолжительность жизни самки перестает быть мерилом существования муравейника. Так свойственная насекомым стратегия, направленная на максимальную плодовитость в ущерб индивиду, сочетается у муравьев со стратегией повышения устойчивости общины. Но не только общины.
Продолжительность жизни отдельной особи тоже возросла, не говоря уже о самках, которых по этому показателю сравнить можно только с самками термитов. Рабочие муравьи живут по несколько лет, в лаборатории, где смертность от случайных факторов ниже, прослежены индивиды до семилетнего возраста. Из устойчивости системы следует увеличение продолжительности жизни составляющих ее индивидов. Данная взаимосвязь имеет значение не только непосредственно для численности семьи, но для ее успешной жизнедеятельности, что связано с обучением. А поскольку в любом муравейнике одновременно присутствуют представители нескольких поколений рабочих, складывается благоприятная ситуация для накопления в семье опыта многих поколений, т.е. сигнальной преемственности. И в этом плане любая форма образования муравейника делением очень привлекательна, так как вновь образовавшиеся семьи оказываются преемниками не только генетической информации, но и передаваемого сигнальным путем опыта материнской общины.
С ростом семьи муравьи так или иначе приближаются к верхнему пределу численности особей, входящих в одну систему. Чем же все-таки обусловлен критический уровень для того или иного вида? Ведь этот уровень может многократно отличаться даже у весьма таксономически близких видов. Изученность данного вопроса пока что оставляет желать лучшего. Можно лишь предположительно очертить круг значащих здесь факторов.
Несомненно, что причина различий кроется в особенностях действия механизмов, объединяющих членов общины. Первое место, по-видимому, принадлежит характеристикам пищевого потока как реальной материализующей основе сообщества насекомых. Пищевой поток не может быть слишком большим, так как в этом случае он не в состоянии выполнять одну из основных функций - регуляцию состава общины. После какого-то предела пищевой поток просто утратит чувствительность к частным изменениям в составе сообщества. Если в группе из нескольких десятков особей муравьи способны заметить исчезновение одной особи, то в стотысячном муравейнике такая потеря остается незамеченной. В миллионной семье чувствительность общины к частным изменениям в составе будет еще ниже. Это общий принцип для любых муравьев. Несомненно, важное значение имеют пищевая специализация и форма пищи, предлагаемой личинкам. Известно, что, например, у муравьев феидолия часть личинок переводится на твердую пищу, с чем, кстати, связано развитие этих личинок в солдат.
Можно допустить, что у зерноядных муравьев пищевой поток должен быть иным, нежели у хищных или же у видов со смешанным питанием. Но и в пределах каждой из этих трофических групп есть виды с самой различной численностью одинарной семьи, т. е. с пищевой цепью разной длины.
А как обстоит дело со скоростью распространения пищи, например белковой и углеводной, а также в семьях различной численности? Уже имеются некоторые обнадеживающие данные. Энтомолог из ФРГ П.Шнейдер показал в 1966 г., что в небольших группах (10 рабочих) рыжих лесных муравьев распределение белков происходит так же, как и углеводов. Но уже при 450 особях в группе белки распределяются значительно медленнее. Значит, переход на углеводное питание связан с ускорением пищевого потока в семье. Чем быстрее распределяется пища, тем прочнее должны быть связи в общине, тем чувствительнее система контроля состава и жизнедеятельности самок, основанная на трофаллаксисе. Тогда у видов с тенденцией к гигантизму семьи скорость распространения пищи должна быть выше. Это подтверждено экспериментами, поставленными известным английским мирмекологом М.Брайеном и другими исследователями: у муравьев формика эта скорость в несколько раз выше, чем у мирмика. Особенно важно быстрое распространение углеводной пищи, так как ее основными потребителями являются взрослые рабочие особи - наиболее подверженная случайным колебаниям часть населения гнезда. Изменение соотношения возрастных и функциональшлх групп рабочих особей вызывает соответствующие изменения в составе гормональных веществ, передаваемых вместе с пищей и определяющих ход развития личинок и молодых рабочих. Чем быстрее распространяется такая информация, тем скорее реакция семьи и больше ее возможности компенсировать происходящие изменения перестройкой организации. Наконец, тем труднее обособление каких-то групп муравьев, ведущих к подразделению семьи на колонны.
Рис. 18. Транспортировка внутригнездового муравья муравьем-носильщиком в семье мирмика
Вторым, но не менее важным моментом, ограничивающим численность одинарной семьи муравьев, являются коммуникабельность отдельной особи. Известно, что у видов с небольшими семьями особи менее контактабельны, чем у муравьев с семьями-гигантами. У первых практически отсутствует взаимодействие фуражиров на участке. Даже если мобилизация и производится, то только внутри гнезда или на гнездовом валу (кратере). Совместные действия на территории при этом ограничиваются следованием пассивного фуражира за разведчиком к месту нахождения добычи. Обменов тактильными сигналами и трофаллаксиса на кормовом участке нет. Все это сказывается и на мобильности фуражиров. Чем ниже уровень контактов на территории, тем меньше возможность изменить направление движения фуражировочной экспедиции, т. е. произвести перемобилизацию или организовать несколько экспедиций одновременно. Сборщики семян аралокаспийского муравья-жнеца не в состоянии изменить выбранного заранее направления фуражировки, если идущие впереди разведчики прошли возможную точку поворота. Можно насыпать прямо на дороге любое количество семян, но сборщики семян будут перешагивать через них, не замечая и не реагируя. Они следуют за лидерами в потоке, следуют к цели, еще не известной им, но реальной, к цели, которая известна лишь нескольким разведчикам, идущим впереди. Дорожка из семян, уводящая в сторону от избранного разведчиками направления, сейчас не для сборщиков. Это, конечно, не значит, что она вообще осталась незамеченной. Другие разведчики уже нашли неожиданную добычу и деловито обследуют ее, видимо, оценивая запасы. Но им не остановить потока. Только в следующую фуражировку поток окажется в состоянии сменить свое направление. Но семян к этому времени может не оказаться. Их могут собрать муравьи из соседнего гнезда, так как кормовые участки семей муравьев этого вида перекрываются (рис. 19). Неподалеку от жнецов находятся гнезда пустынного дернового муравья. Муравьи здесь помельче, но гораздо мобильнее. Это не удивительно: активных фуражиров у них в несколько раз больше и мобилизацию они производят несколько раз в день. Пройдут минуты, и к рассыпанным семенам будет проложена дорога. Были бы только семена под силу сборщикам. Но и для этих муравьев предел возможностей лежит недалеко. Обитатели одной секции могут лишь один-два раза в день сменить направление фуражировки или образовать две-три дороги.
Но есть муравьи еще более мобильные. Например, семья феидолия может одновременно организовать более 10 фуражировочных потоков, легко переключается с одного объекта на другой. А если добыча велика для этих двух-трех миллиметровых муравьишек, то на дороге появятся головастые солдаты - они и побольше и могут мощными челюстями расчленить добычу на месте, чтобы сделать ношу посильной. Не хватит сил одного муравья - добычу понесут, поволокут несколько. Коллективная транспортировка добычи, так же как и коллективная охота, прямое следствие повышенной контактабельности фуражиров ряда видов. Само по себе это дает значительный выигрыш, так как расширяет размерный диапазон добычи. Совместно муравьи могут овладеть более крупными жертвами, и не только овладеть, но и доставить их в гнездо. При этом не ускользают и мелкие насекомые; ведь индивидуальная охота тоже сохраняется. Но этот выигрыш еще не означает роста численности семьи как единой системы. Семья может просто быстрее достичь верхнего предела и разделиться. Но в том-то и дело, что контакты на территории, совместные охоты и транспортировка добычи - это надежные, достоверные проявления присущей муравьям этого вида общительности.
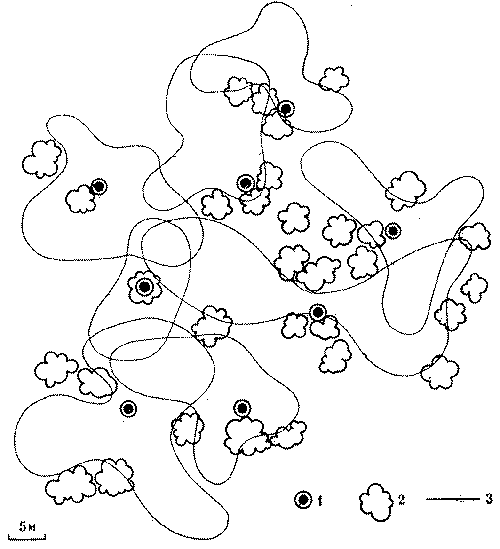
Рис. 19. Кормовые участки группы муравейников аралокаспийского муравья-жнеца 1 - гнезда. 2 - кусты и деревья, 3 - границы кормовых участков
Самые общительные среди наших муравьев - рыжие лесные; они же имеют самые многочисленные семьи. Контакты, обмен тактильными сигналами, обмен пищей, групповые охота и транспортировка добычи в любой части охраняемой территории - вот что характеризует в этом отношении рыжих лесных муравьев. А размер охраняемой территории у наиболее мощных муравейников может превышать 2 га. Почти столь же организован на участке и пахучий муравей-древоточец - второй по размерам семьи муравей наших лесов.
По мере снижения численности семьи из арсенала взаимодействия фуражиров на участке одно за другим выпадают совместная транспортировка добычи и групповая охота, прекращается трофаллаксис на территории, сокращаются зоны мобилизации и обмена тактильными сигналами. Наконец, фуражир становится одиночным охотником, ведущим охоту на свой страх и риск и избегающим любых контактов с кем бы то ни было. Только в гнезде в нем проснется тяга к общению с другими муравьями.
Прослеженный нами путь ретроспективен. Первые муравьи были одиночными охотниками и имели очень небольшие семьи. Численность общины и коммуникативные возможности индивидов нарастали параллельно и взаимосвязанно. Соответственно раздвигались и границы постоянного взаимодействия индивидов на участке. Отодвигался верхний предел численности муравьев, объединяемых в единую систему.
РОСТ ПРИСПОСОБИТЕЛЬНЫХ ВОЗМОЖНОСТЕЙ СЕМЬИ
Большие размеры семьи сами по себе - это важное новое свойство. (Примерно в той же степени, как и размеры отдельного индивида). Многочисленная семья всегда более конкурентоспособна, ей легче отстоять свой кормовой участок от посягательств соседних муравейников и расширить собственную территорию за их счет. Чем больше муравейник, тем меньше зависит он от колебаний погоды и других воздействий внешней среды, тем устойчивее сообщество во всех его проявлениях. О необходимости определенной численности семьи для поддержания гиг-ротермического режима в гнезде мы уже говорили. Добавим только, что при одинаковых прочих условиях более крупные муравейники раньше начинают активизироваться весной и позднее прекращают активность на участке осенью. В них строже выдерживается оптимальная для развития молоди температура, а значит сроки развития молоди минимальны. Это обстоятельство имеет в ряде случаев важное значение для сохранения и перспектив развития семьи. Например, холодным летом 1976 г. в небольших семьях северного лесного муравья развитие молоди так затянулось, что общее число поколений рабочих оказалось меньше обычного на одно (три вместо четырех). Семья недополучила четвертую часть пополнения. В крупных муравейниках и в этот неблагополучный год выросли все четыре поколения рабочих особей. В следующем году при весеннем переделе территорий более крупные семьи получают дополнительное преимущество.
При этом кроме абсолютной численности поселения семей существен и его возрастной состав. Очевидно, что для муравьев имеет значение не только возможность достичь высокого уровня численности одной общины, но и скорость роста, время, необходимое для выхода на этот уровень. Затягивать данный процесс опасно, так как в этом случае желанный уровень может оставаться недостижимым для абсолютного большинства семей из-за регулярно повторяющихся неблагоприятных изменений среды. Поэтому высокая плодовитость самок - необходимое условие успешного развития социальности у муравьев. Эта плодовитость не обязательно должна быть единовременной, так как переход части видов к выращиванию нескольких поколений рабочих особей в год позволяет им ускорить рост общины без увеличения разовой плодовитости или числа самок. Эта возможность, как уже говорилось ранее, появилась также в результате увеличения численности семьи муравьев.
Выведение крылатых репродуктивных особей тоже связано с численностью семьи. Маленькие семьи их не выводят. С ростом муравейников выведение крылатых особей становится все более регулярным. Крупные, активные муравейники выводят их ежегодно. Это связано прежде всего с семейными запасами пищи и численностью молодых рабочих муравьев, выделениями лабиальных желез, которыми выкармливаются личинки самок. По данным энтомологов из ФРГ К.Гёссвальда и К.Вира, у малого лесного муравья выращивание крылатых особей удается Щи наличии не менее 3500 рабочих на одну личинку самки. У лугового муравья минимальное число молодых рабочих на одну личинку самки - 600.
Увеличение численности позволило осуществить чрезвычайно узкую специализацию рабочих муравьев. Число профессий рабочих в принципе не ограничено. Узкая специализация позволила муравьям обеспечить все жизненно важные позиции инициативными, владеющими ситуацией исполнителями. Прогресс социальности связан с увеличением численности инициативных особей, совершенствованием рабочих муравьев как функционеров, развитием разнообразных форм индивидуального и группового поведения.
Таким образом, путь увеличения размеров семьи прогрессивен не только в приложении к муравейнику как целостной системе, но и применительно к индивиду (рабочей особи).
Наиболее упорные сторонники теории «сверхорганизма», пытающиеся методом аналогии приравнять семью общественных насекомых к организму, рассматривают отдельных муравьев как некое подобие клеток. Основным доводом при этом служит якобы возрастающая по мере дифференциации функций зависимость особи от общины и неспособность изолированного муравья к длительному существованию. Однако здесь упускается из виду, что основной показатель зависимости особи от семьи - неспособность одиночки к длительному существованию - у муравьев и других общественных насекомых изначальны. Муравьи не могли утратить его в ходе дальнейшей эволюции, оставаясь социальными насекомыми. Но вот дальнейшего развития эта зависимость не получила. Все разделение функций рабочих муравьев совершалось на едином фоне субординации индивида и семьи. И у примитивных видов с десятком неспециализированных рабочих, и в миллионных семьях с узкоспециализированными группами особей и сложной организацией эта зависимость одна и та же. Показательно, что для индивида в принципе важно состоять в любой общине, а не обязательно в родном муравейнике. Какой-либо «тканевой несовместимости», естественно, здесь нет и быть не может. Даже рабочие разных видов, вышедшие из куколок в одном гнезде, успешно живут совместно. На этом основано, в частности, и явление временного социального паразитизма, и существование муравьев-рабовладельцев. Взаимную терпимость могут проявлять не только рабочие, но и оплодотворенные самки.
«Рабовладельцы» муравьи-амазонки нередко основывают свои муравейники на семьях серого песчаного муравья. В дальнейшем амазонки совершают регулярные набеги на окрестные гнезда серого песчаного муравья, похищая куколок, так как без «рабов» муравьи-амазонки существовать не могут. Из куколок, захваченных рабовладельцами, выходят рабочие, выполняющие в гнезде все работы по строительству гнезда и уходу за самкой и молодью. В семьях амазонок всегда одна самка, которая при внедрении в семью вида-хозяина также с одной самкой убивает последнюю и занимает ее место.
Но серый песчаный муравей - вид полигинный, у которого полигиния сочетается с секционным типом гнезд. Вот здесь и открывается возможность для сосуществования в составе одной семьи муравьев двух видов, да еще и с самками. Такая ситуация была описана польским мирмекологом А.Чеховским в 1976 г.
Амазонки, обитающие в одной из секций крупного муравейника серого песчаного муравья, поддерживают с остальными секциями родственные отношения и даже обмениваются с ними молодью. Набегов на обитателей этих секций они не совершают. Те, в свою очередь, лояльны к внедрившимся в их поселение муравьям-рабовладельцам. Среди обмениваемых особей отмечены даже личинки крылатых амазонок, которых серые песчаные муравьи выкармливают в своих секциях. А за куколками амазонки отправляются в дальние рейды, нападая на чужие муравейники серого песчаного муравья. В лабораторных экспериментах одновременным выведением из куколок создавались общины из рабочих четырех-пяти видов, относящихся к разным подродам и даже родам. Такие сообщества оказывались довольно устойчивыми, функциональные группы формировались тут по видовому принципу. Представители наиболее воинственного вида становились охотниками, более мирные, но крупные муравьи - сборщиками пади и строителями, остальные делили между собой обязанности внутри гнезда.
Запаховый тест «свой - чужой» играет важную роль и при конкурентных отношениях муравьев на участке, накладывая определенный отпечаток на территориальное поведение муравьев. У птиц как конкуренты обычно воспринимаются только особи своего вида. При этом важно внешнее сходство, так как опознание во всех случаях визуэльное. Птицы другого облика, даже занимающие ту же экологическую нишу, остаются без внимания со стороны самых зорких стражей кормового участка. У муравьев ситуация другая. Все особи, пахнущие иначе, чем члены своей семьи, чужие и заслуживают настороженного внимания. Если встреченные муравьи близки хозяевам участка по размерам, они подвергаются нападению. Особенно четко это проявляется у видов, имеющих охраняемые территории. Статус охраняемых территорий строго соблюдается при соседстве муравейников разных территориальных видов. Многими авторами описаны столкновения между такими соседями, если один из них нарушает установившиеся границы.
Правда, это случается не всегда. В сложных условиях обитания жизнь учит и самых агрессивных муравьев терпению. В его основе лежит одна из многочисленных специфических характеристик муравьев как общественных насекомых.
Дело в том, что свой кормовой участок муравьи защищают только сообща, только тогда, когда их много. Не обязательно на самом месте событий, но хотя бы где-то рядом. В тех случаях, когда численность гнезд с охраняемыми территориями низкая, невелика и плотность поселения, по периферии кормовых участков складываются обширные зоны с низкой численностью фуражиров. Если соседствуют гнезда одного вида, все равно границы будут достаточно четкие. А вот для разных видов появляется исключение. Встретившись на участке, одиночные охотники, зная, что поблизости никого из своих нет, предпочитают мирно разойтись. Добычи на участке хватает, а конфликт с другим, примерно одинаковым по размерам муравьем - достаточно опасное дело. В результате такой осмотрительности фуражиров-одиночек по обе стороны границы возникает зона взаимного проникновения* муравьев из семей двух видов, как это показано на рис. 20 для муравейников разных видов. Границы, как таковой, здесь нет, нет и нейтральных пространств между охраняемыми территориями. Но они появятся, если плотность поселения муравьев значительно повысится и соответственно увеличится численность фуражиров хотя бы одного вида на участке. А пока что на весьма обширной территории конкуренты демонстрируют лояльность. Причем в одном участке встречаются и охотятся на паритетных началах особи сразу трех видов. Но достаточно кому-нибудь из неосторожных фуражиров увлечься и оказаться слишком близко от чужого гнезда, где численность хозяев уже достаточно велика, его сразу же атакуют, и если он не спасется бегством, то будет уничтожен. Здесь зона взаимного проникновения кончается и начинаются монопольные владения каждой семьи (зона монопольного использования)*.
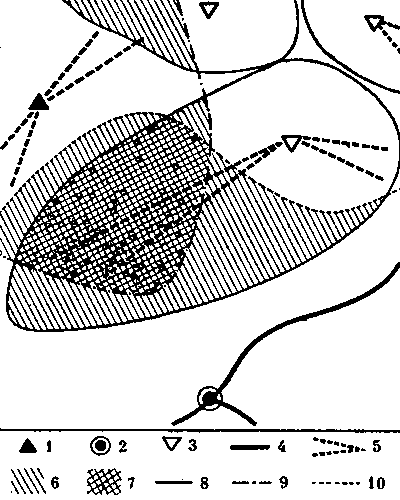
Рис. 20. Взаимно перекрывающиеся кормовые участки видов-доминантов в ассоциации муравейников
1 - гнездо красногрудого муравья-древоточца,
2 - гнездо
обыкновенного лесного муравья,
3 - гнездо кроваво-красного мур
авья-раб овладельца,
4 - дороги обыкновенного лесного муравья,
5 - основные направления фуражировки муравья-древоточца и
муравья-рабовладельца,
6 - зона взаимного проникновения фуражиров двух
видов,
7 - зона взаимного проникновения фуражиров трех видов-доминантов;
границы кормовых участков:
8 - кроваво-красного
муравья-рабовладельца,
9 - красногрудого муравья-древоточца,
10 -
обыкновенного лесного муравья
Все сказанное касается видов территориальных, доминирующих на своих кормовых участках. Но на этой же территории обитают и другие муравьи, послабее, с менее многочисленными семьями и не охраняющие своих кормовых угодий. Они подчиняются доминантам, что проявляется и во времени их фуражировок, в размещении гнезд, и в поведении при встрече с особями вида-доминанта. Отношения между семьями разных видов, обитающими на одной территории, строго регламентированы и построены по иерархическому принципу. В результате складываются весьма устойчивые многовидовые ассоциации муравейников*, жизнь которых регулируется исключительно поведенческими средствами. Исследования А. В. Демченко показали, что доминирование прямо связано с Соотношением численностей муравейников, входящих в ассоциацию. Ж.И.Резникова, проследившая в различных ландшафтных зонах Сибири состав многовидовых ассоциаций, установила, что если доминировал луговой муравей, то ассоциация была весьма стабильной. В этом случае можно говорить о глубоких взаимных адаптациях всех участвующих в многовидовой ассоциации муравейников и о развитии ее как достаточно целостной системы. Понятно, что сюда входят виды, близкие по экологическим требованиям к местам поселения и по типу питания. Однако этих условий еще недостаточно для стабильности таких систем. Необходим организующий центр - муравейник, характер и масштабы деятельности которого способны задать специфическую пространственную и временную структуру системы.
Здесь мы сталкиваемся с новым важным следствием прогрессивного роста численности муравьиной общины. Мощный муравейник становится именно таким центром, вокруг которого формируются поселения других видов муравьев. И не только муравьев. Воздействуя на почву и растительность, крупные муравейники оказывают заметное формирующее воздействие на среду и ее обитателей.
Рассмотрев различные аспекты роста численности и усложнения структуры муравьиной семьи, мы можем выделить основную адаптивную тенденцию этого процесса. По мере прогрессивного развития общинного образа жизни у муравьев происходило постепенное распространение сферы постоянно действующей регуляции плотности поселения и числа поселений с одиночной семьи на колонию, с колонии на федерацию и весьма обширные комплексы муравейников, которые в отдельных случаях можно считать популяциями. Параллельно возникли достаточно эффективные механизмы регуляции многовидовых ассоциаций муравейников, обеспечивающие максимальное использование всех доступных муравьям ресурсов каждого биотопа. Все это способствует достижению и стабилизации максимальной численности каждого вида, входящего в ассоциацию. Естественно, стабильность того максимума, который может обеспечить уровень развития социальности конкретного вида, допускает кормовая база. Пример муравьев достаточно наглядно показывает, что лишь стабильный рост численности вида может служить показателем его биологического прогресса. И все же, несмотря на то, что масштабы постоянной саморегуляции у ряда муравьев намного перекрыли размеры отдельной семьи, основным объектом регуляции и оптимизации остается именно она. Семья сохранила целостность и индивидуальность и в рамках колонии, и в составе федерации. Ее оптимизация и устойчивость - основная неизменная задача общего адаптивного процесса, что подтверждает высказанное еще Ч.Дарвином положение о воздействии отбора на семью общественных насекомых в целом. Здесь, по-видимому, следует подразумевать одинарную семью, не подразделяющуюся на колонны. При появлении в полигинном муравейнике нескольких колонн можно допустить смещение точки приложения отбора с целой семьи на составляющие ее колонны.
Данный раздел неспроста посвящен почти исключительно адаптации муравьиной общины к обитанию среди других муравейников. Если для особи основная среда обитания - семья, то для семьи, хотя и в несколько меньшей мере, - это окружающие ее другие семьи муравьев. Разумеется, есть ряд факторов, масштабы и степень воздействия которых непреодолимы. Но по мере развития социальности, роста численности населения семьи, совершенствования гнезд и стабилизации кормовой базы посредством трофобиоза и разведения грибниц зависимость муравейников от абиотических (погодных условий, состояния субстрата и т. д.) и ряда биотических факторов резко падает. Муравьи не только добиваются значительной автономности от внешних факторов, но и получают возможность активно воздействовать на среду обитания, создавая условия оптимума в гнезде и на используемой ими территории. Этим они обязаны преобладанию поведенческих приспособлений к условиям обитания и необычайной пластичности многих видов, обеспеченной развитием комплекса таких адаптации. По мере развития социального уклада жизни формы поведения и координации деятельности, активно используемые муравьями, становятся все разнообразнее, хотя в принципе большинство из них имеется в резерве у каждого вида. Очевидно, что у муравьев, достигших более высокой ступени социальной организации, шире возможности использования их биологического потенциала. Это, впрочем, не предполагает как обязательное следствие более высокую степень психологического развития особи. Преимущества просматриваются, как правило, на уровне общины и реализуются прежде всего на основе глубоких функциональных различий индивидов, составляющих общину.
Но какая участь постигла тогда те виды муравьев, которые пошли по пути морфологической и морфофизиологической специализации к определенным местам поселения или отдельным факторам среды? Ведь таких среди муравьев тоже немало. Они не только оказались в полной зависимости от состояния субстрата, но и не смогли расширить свои ареалы за пределы весьма ограниченных районов. Для всех видов, развивавшихся по пути морфологических и морфофизиологических специализаций, характерны небольшие семьи с относительно простой организацией.
Итак, только благодаря совершенствованию поведения и усложнению организации муравьи обеспечили рост общины, получили возможность активного противодействия окружающей среде, ее повсеместного освоения и оптимизации условий собственного существования. Они добились высокой степени автономности от многих факторов, целиком и полностью управляющих почти каждым шагом одиночного насекомого. Но вместе с тем вырос новый комплекс задач, связанный с взаимодействием муравейников друг с другом. Муравейник среди муравейников - одного и разных видов, больших и маленьких, агрессивных и миролюбивых, связанных друг с другом не только близостью требований к условиям обитания и пищевой специализацией, но и небходимостью жить рядом, как правило, на весьма ограниченной территории. Проблем стало не меньше, а скорее больше, и обеспечить процветание вида можно все тем же путем - дальнейшим совершенствованием поведения, усложнением организации, ростом численности семьи.
Что же дало муравьям развитие социальности? Какие преимущества получили они по сравнению с одиночными насекомыми? Поскольку многие вопросы достаточно подробно рассматривались в предыдущих разделах книги, перечислим основные моменты, комментируя лишь те из них, которые были слабее освещены ранее:
дифференциацию функций и специализацию особей, повысивших эффективность
деятельности индивида и семьи в целом;
создание оптимальных условий для
развития молоди и сокращение смертности на фазах яйца, личинки и куколки;
увеличение продолжительности жизни особи;
направленное воспитание потомства
и регуляцию этим путем соотношения каст и полиэтических групп рабочих особей;
расширение кормовой базы семьи за счет полиморфизма рабочих, групповых охоты
и транспортировки добычи, а также таких достижений, как трофобиоз и разведение
грибных садов;
накопление запасов пищи в телах муравьев или в виде исходного
продукта (семена у муравьев-жнецов);
уменьшение потребления пищи на одну
особь с ростом семьи;
экономичность поиска добычи и фуражировки при
появлении систем мобилизации и вторичного деления территории. Особенно
эффективен второй принцип, что подтверждается численностью фуражиров в семьях
видов с вторичным делением территории: фуражировкой занято около 13% взрослых
особей семьи, т. е. один фуражир в состоянии прокормить в среднем 7,7 взрослых
рабочих, не считая примерно такого же числа молоди в каждом поколении. У многих
видов численность молоди превышает таковую взрослых особей;
рост
количественных возможностей семьи, ее жизнеспособности, увеличение ее
автономности от внешней среды;
постоянную социальную саморегуляцию
численности и состава семьи, стабилизацию семей и комплексов муравейников на
высоких уровнях численности;
образование нового муравейника делением или
почкованием - избегание наиболее уязвимого этапа одиночных самок-расселительниц;
общий рост адаптивных возможностей вида.
Благодаря этим преимуществам муравьи смогли достичь вершин организации в мире насекомых и стать фактическими доминантами во многих биогеоценозах, компонентами ценозов, деятельность которых оказмвается наиболее заметна.
5. ПЕРСПЕКТИВЫ МУРАВЬЕВОДСТВА
Данный раздел можно начать по-разному. Показать, как по мере расширения и интенсификации хозяйственной деятельности человека приобретают угрожающие размеры последствия неосмотрительного хозяйствования. Для борьбы с ними необходимы не только совершенствование методов ведения хозяйства, но и привлечение на помощь наиболее активных живых компонентов естественных сообществ. Среди «наиболее активных» видное место занимают муравьи. Или перечислить биоценологические заслуги муравьев в условиях различных природных зон. Здесь большинство видов муравьев своей деятельностью способствуют повышению продуктивности земель. Наконец, общеизвестна роль муравьев как санитаров леса.
Конечно, муравей муравью рознь. Есть среди них и опасные вредители, и карантинные объекты, с которыми приходится вести длительную борьбу без надежд на быстрый успех. В предыдущих главах об этом уже говорилось. Желтый земляной муравей повинен в образовании кочек на лугах и сенокосах и, по-видимому, даже является одной из причин заболачивания этих угодий. Борьба с ним еще предстоит, что, естественно, потребует глубокого изучения его биологии. Но большинство муравьев несомненно полезны.
Надо сказать, что оценить характер деятельности того или иного вида можно только при специальном изучении. Ограниченность наших знаний нередко приводит к ошибочным выводам. При этом на основании случайных сведений или непроверенных историй и мнений нередко складывается предубеждение по отношению к тому или иному виду. Подобные примеры можно привести из разных областей биологии.
Есть незаслуженно обиженные и среди муравьев. Так, издавна считалось, что муравьи-жнецы рода мессор являются серьезными сельскохозяйственными вредителями, поскольку они собирают большое количество зерна. Поговаривали даже о необходимости решительной борьбы с этими муравьями. К счастью для муравьев-жнецов, в 1970-1972 гг. в Туркмении под руководством Г.М.Длусского было проведено детальное исследование биологии всех основных видов рода мессор. В том числе изучена и связь муравьев-жнецов с зерновыми культурами. Наблюдения показали, что в период до созревания зерновые муравьи занимаются сбором семян различных видов сорняков и не трогают колосков пшеницы или ячменя. При созревании зерновых муравьи также не собирают зерна с колосков. Они просто не в состоянии забраться по стеблю злака, чтобы добраться до растущих колосков. И лишь после уборки урожая муравьи-жнецы начинают собирать осыпавшееся, потерянное в ходе жатвы зерно. Добыча жнецов при этом оказывается весьма солидной - до 2 ц зерна с 1 га. Приведенный факт показывает, насколько ошибочными могут оказаться выводы, сделанные на основании отдельно взятого факта, например обнаруженных в муравейнике пшеничных зерен. Бороться, оказывается, надо не с муравьями-жнецами, а с потерями при уборке урожая.
Что же полезного для биоценоза и хозяйственных интересов человека несут с собой муравьи? Разберем некоторые аспекты воздействия муравьев на биогеоценоз.
ЧЕМ ПОЛЕЗНЫ МУРАВЬИ
Все живое связано с окружающей средой и оказывает на нее определенное воздействие. Малочисленные, с ничтожной плотностью поселения виды лишь в исключительных случаях могут сколько-нибудь заметно повлиять на биоценоз. Чем многочисленнее вид, выше численность и биомасса на единицу площади, тем значительнее его биоценотическая роль. Значимость муравьев обусловлена их многочисленностью. Но не только этим. Важны и особенности образа жизни муравьев, их питания и связей с другими животными, растениями, почвой.
Основные аспекты воздействия муравьев на биогеоценоз можно разделить на две группы: связанные с сооружением гнезда и постоянством места поселения (кочующие муравьи здесь не рассматриваются) и обусловленные трофическими связями муравьев с другими группами животных и растениями.
К первой группе относятся такие важные стороны деятельности муравьев, как концентрация и ускоренное разложение растительных остатков, аэрация и улучшение водного режима и структуры почв. Почва обогащается гумусом и важными для растений элементами (фосфор, азот, калий, магний и т. д.) в доступных для растений формах. Муравейники используются как места поселения другими животными. В них создаются особо благоприятные условия для развития ряда микроорганизмов.
Во вторую группу входят хищническая и собирательная деятельность муравьев, их трофобиоз с тлями и другими выделяющими падь насекомыми, а также связи муравьев с рядом позвоночных животных.
Почвообразующая деятельность муравьев.Сооружая гнездо, муравьи оказывают серьезное воздействие на почву, где строятся гнезда большинства видов этих насекомых. Численность гнезд, как правило, весьма высока: на одном гектаре может находиться до 7-8 тыс. муравейников, причем число гнезд менее тысячи на 1 га - явление редкое. Почвообразующая деятельность муравьев складывается из следующих факторов: перемешивания почвы, изменения механического состава, изменения химического состава почвы.
Перемешивание почвы происходит при рытье ходов, при котором муравьи поднимают частицы почвы из нижних горизонтов на поверхность. Одновременно улучшается доступ воздуха к корням растений. Перемешивание почвы производится муравьями постоянно, а не только в период сооружения гнезда, так как муравьи все время перестраивают свое жилище.
В гнездах и вокруг них накапливается большое количество экскрементов, удобряющих почву. Муравьи, поселяющиеся в древесине или включающие в гнезда старые пни, участвуют в механическом разрушении отмершей древесины, ускоряя тем самым процесс ее разложения. Рыжие лесные и тонкоголовые муравьи собирают с окружающей гнездо территории древесные и травянистые частицы, веточки, хвою, чешуйки, травинки и т. д., концентрируя их в муравейниках, где разложение этих частиц идет значительно быстрее, чем на поверхности почвы. Однако важное значение муравьев как фактора почвообразования объясняется не этим, а созданием в гнездах большинства видов специфической среды, значительно отличающейся от окружающей и способствующей более быстрому разложению и гумификации попадающих в гнездо растительных остатков, повышению биологической активности почв. Это связано с более высокой, чем в окружающей почве, температурой гнезда, с развитием в муравейнике микрофлоры - бактерий, грибов, актиномицетов. Разложение ускоряется в десятки раз.
Муравейники представляют собой очаги повышенного плодородия почв. Маленькие гнезда - микроочаги; крупные гнезда - макроочаги, уже ощутимые при стандартных методах учета.
Гнезда рыжих лесных муравьев старше 10 лет и диаметром более 1 м как «фабрики плодородия» настолько мощны, что в состоянии оказать серьезное влияние на рост не только произрастающих непосредственно у муравейников трав и кустарников, но и деревьев, достаточно удаленных от муравейника. Корни деревьев распространяются под поверхностью почвы на несколько метров. Попадая в зону влияния гнезда, они буйно разрастаются, используя накапливаемые под муравейником питательные вещества. Отмечен более быстрый рост сосны в местах, где имеется большое число муравейников. Гораздо выше и гуще вокруг муравейников травостой. Масса травостоя вокруг гнезд среднего муравья-жнеца в 5-8 раз выше, чем на контрольных участках.
Вокруг гнезд развивается растительность, требовательная к плодородию почв, что особенно заметно на бедных почвах. В сосняках Воронежского заповедника валы жилых и брошенных гнезд малого лесного муравья - места возобновления дуба. Муравейники оказываются форпостами более зрелых почв и растительных формаций, элементами, ускоряющими их смену (сукцессию). Детали и особенности этой роли муравейников в биогеоценозе еще предстоит выяснить, но сам факт важного участия муравьев, их активизирующей роли в сукцессионных процессах, протекающих в почвах, признан и не вызывает сомнения.
В муравейниках формируется и население для будущих фаз развития ценоза. Среди мирмекофилов - тяготеющих к муравейникам беспозвоночных (их насчитывается несколько сот видов) - многие избирают гнездо муравьев как место обитания именно потому, что здесь быстрее идет процесс гумификации, лучше структура и водный режим почв, т. е. достигнут тот уровень развития почв, который предстоит окружающей территории только в отдаленном будущем. Многие мирмекофилы также вносят свою лепту в почвообразующую деятельность муравейника.
Муравьи и вредные насекомые.Большинство муравьев охотятся на различных беспозвоночных, используя добываемую таким путем белковую пищу для выкармливания молоди. Многочисленная молодь требует огромных количеств пищи, что вынуждает фуражиров неустанно искать добычу. Значение хищничества муравьев для популяции насекомых-жертв зависит от размеров семей и плотности поселения различных видов муравьев. Некоторые муравьи как хищники могут подавлять размножившийся вид жертвы на контролируемых муравьями территориях. Этому способствует установленное Г.М.Длусским явление реактивности муравьев на пищу. Суть его сводится к переключению фуражиров на добычу, ставшую наиболее массовой. А ведь именно в случае массового размножения насекомое-фитофаг и становится вредителем. В этом случае оно приносит ущерб лесу или сельскохозяйственным растениям, а значит - хозяйственным интересам человека.
В добыче муравьев преобладают вредные насекомые. Во время массового размножения вредители в добыче муравьев составляют 90% и более.
Многочисленными исследователями на позвоночных животных показано, что хищники, как правило, способствуют оздоровлению популяции жертвы, поскольку изымают наиболее слабых, больных, малоподвижных животных. В некоторых выступлениях данный принцип механически переносится и на муравьев. Это ошибочная позиция, так как она не учитывает особенности поведения муравьев-охотников. Дело в том, что муравьи охотятся почти исключительно на подвижную добычу, неподвижных насекомых они обычно не замечают, т. е. больные (малоподвижные) и мертвые насекомые становятся добычей муравьев в последнюю очередь (обычно при недостатке здоровых). Это относится ко всем хищным муравьям.
Есть группа видов, в основном обитателей пустынь, относящихся к некрофагам, т.е. питающихся трупами насекомых. Но в лесах таких видов почти нет. Зато абсолютное большинство лесных муравьев - хищники, активно истребляющие вредителей леса. Рыжие лесные муравьи - эффективные защитники насаждений от хвоегрызущих и листогрызущих вредителей. Но кроме них существенна роль и других муравьев: красноголового, песчаного, прыткого степного, краснощекого и тонкоголовых муравьев - практически всех муравьев рода формика. Активно истребляют вредителей муравьи-древоточцы, желтый пахучий муравей, бегунки, фаэтончики и др. Даже среди муравьев-жнецов, специализированных на питании семенами, есть виды со значительным развитием хищничества. Например, в добыче среднего муравья-жнеца живые насекомые составляют (по числу принесенных в гнездо экземпляров добычи) до 30%.
Ряд важных аспектов деятельности муравьев как хищников мы разберем ниже. Хищничество муравьев - безусловно положительный фактор, действие которого направлено па стабилизацию биоценоза и сохранение его основных растительных компонентов (в лесу - древесного яруса).
Трофобиоз муравьев с тлями.Как известно, многие тли являются серьезными вредителями растений. Поэтому при оценке биоценотического и экономического значения муравьев необходимо тщательно взвесить влияние охраняемых муравьями тлей на растения. Исследования показали, что тли делятся на две экологические группы: в первую - входят виды, сосущие из паренхимы, во вторую - из проводящих тканей растений. Представители первой группы почти не выделяют пади и с муравьями не связаны. Не используют муравьи и тлей, покрытых восковым налетом. Именно к этим группам и относятся почти все основные виды тлей - вредителей леса.
Трофобиотические отношения имеются у муравьев с видами второй группы. Проводились длительные (в течение 10-15 лет) наблюдения, которые показали, что в хвойных лесах вреда от охраняемых муравьями тлей нет. Более того, присутствие муравьев избавляет лес от вредных последствий жизнедеятельности тлей. Дело в том, что основная угроза деревьям заключается не в потере части сока, а в сажистом грибке, который развивается на выделяемой тлями пади. Слизывая падь, муравьи предохраняют деревья от этого заболевания.
Но не во всех случаях трофобиоз безобиден. В садах тли наносят серьезный вред плодовым деревьям. Поэтому обычный в садах черный садовый муравей, разводящий тлей на яблонях, грушах и других садовых растениях, может рассматриваться как вредитель. Нецелесообразно по этой же причине и переселение в сады рыжих лесных муравьев. Между тем некоторые садоводы-любители, прослышавшие о пользе этих муравьев в лесу, поспешили заселить ими свои приусадебные участки. Размножившиеся под защитой переселенных муравьев тли причинили немалый вред садам. Не стоит переселять муравейники и в буковые леса, где тли также могут вредить деревьям. Но в остальных случаях значение трофобиоза следует оценивать положительно. Кстати, оно не ограничивается только непосредственным воздействием тлей на деревья, но имеет и другие стороны, о чем пойдет речь ниже, когда мы перейдем к перспективам использования муравьев.
Муравьи и позвоночные животные.Это важный аспект, без которого значение муравьев в жизни биоценоза будет выглядеть неполным. Муравьи служат пищей многим позвоночным, важным для биогеоценоза или имеющим хозяйственное и промысловое значение. Муравьями питаются многие певчие птицы, лесные куриные, многие земноводные и пресмыкающиеся, а также барсуки, лисы, медведи и другие звери. Гнезда рыжих лесных муравьев используются как места зимних ночевок кабанами. Учеными показано, например, что без муравьев невозможно нормальное развитие тетеревов и рябчиков. Поэтому в настоящее время при работах по восстановлению численности этих промысловых птиц планируется специальное расселение рыжих лесных муравьев.
Муравейники служат не только источником необходимой для нормального развития птенцов пищи, но и как «птичьи санпропускники», в которых птицы, «купаясь», очищаются от паразитов.
Характерно, что для ряда видов птиц муравьи являются резервным кормом, помогая пережить периоды бескормиц, что способствует увеличению численности птиц. По данным энтомолога из ФРГ Руппертшофа, искусственное заселение лесов рыжими лесными муравьями вызывает резкое (почти в 5 раз!) увеличение численности дятлов. Остается добавить, что поедаемые птицами муравьи составляют около 10% от общей величины естественной смертности муравьев, что не оказывает большого влияния на жизнь муравейника в целом. Правда, в отдельных случаях небольшие муравейники могут серьезно пострадать от дятлов.
В целом влияние муравьев на окружающую среду, несомненно, благоприятное. Это воздействие наиболее полно изучено в лесу на примере рыжих лесных муравьев. В результате многолетнего положительного влияния этих муравьев на лес увеличивается продуктивность основного яруса деревьев и всего насаждения в целом, а также повышается биологическая устойчивость лесов.
Результаты воздействия рыжих лесных муравьев на лес многообразны. Непосредственное влияние муравейника: обогащение почвы гумусом, К, N, P, Mg в доступных для растений формах; уничтожение вредителей хвои и листвы; увеличение численности насекомоядных птиц; увеличение численности промысловых птиц; возобновление требовательных к почве лесных пород. Опосредованное влияние муравейника: снижение численности стволовых вредителей; увеличение прироста древостоя; повышение продуктивности охотничьих угодий; сохранение прироста крон деревьев в очагах вредителей; повышение продуктивности леса; повышение биологической устойчивости насаждений.
Рыжих лесных муравьев уже в течение ряда лет используют в практике защиты леса. Пока что отрабатываются методы переселения муравейников и накапливаются данные об особенностях взаимодействия муравьев с отдельными вредителями в различных условиях. Без этого, так же как без знания биологии самих муравьев, невозможно их успешное использование. Одновременно выявляются другие перспективные для защиты леса виды, изучается их биология, ведется поиск путей расселения.
Все это лишь первый этап применения муравьев для хозяйственных целей человека. Муравьев можно использовать лучше, полнее, эффективнее. И не только как энтомофагов. Даже с позиции лесозащиты роль муравьев в насаждениях гораздо шире и значимее, чем трактуется в настоящее время. Кроме того ясно, что муравьи - это прежде всего фактор повышения продуктивности леса, т. е. использование их должно иметь не только лесозащитную, но и лесоводческую направленность. В общих мероприятиях по повышению продуктивности земель муравьи должны быть использованы не только в лесу, но и в других растительных зонах, в частности при зоологической мелиорации почв. В ряде случаев муравьи могут служить как индикаторы определенных условий среды и их изменений.
Наконец, муравьи дают и некоторые дополнительные продукты, без которых не обойтись в медицине (муравьиный спирт), а также при содержании певчих птиц (куколки). Возможно появление новых продуктов, которые смогут дать муравьи человеку. Добыча муравьиного спирта и куколок ведется до сих пор самыми примитивными и порою варварскими методами, что ослабляет муравейники и зачастую сводит на нет усилия лесоводов по искусственному их расселению.
Все это показывает, что необходимо более осознанно отнестись к этим полезным насекомым и разработать обоснованную и рациональную программу их комплексного использования на благо человека, разработать и довести эту программу до реального воплощения на деле.
Остановимся на одном из наиболее изученных частных вопросов использования полезных видов муравьев - применении рыжих лесных муравьев в защите леса.
РЫЖИЕ ЛЕСНЫЕ МУРАВЬИ КАК ОСНОВА БИОЛОГИЧЕСКОГО ЛЕСОЗАЩИТНОГО КОМПЛЕКСА
Проведение комплексных мероприятий по биологической защите лесных насаждений от насекомых-вредителей связано с повышением биологической устойчивости древостоев и созданием в лесных массивах стабильной плотности полезных видов, позволяющей контролировать численность основных вредителей леса и обеспечивать благополучное состояние насаждений.
Известно, что слабой стороной большинства используемых в настоящее время биологических средств борьбы с вредителями леса (паразитические насекомые, микробиологические препараты) является разовость их воздействия. Фактически они действуют как биоинсектициды. Кроме того, степень эффективности этих средств жестко связана с точным соблюдением фенологических сроков, что далёко не всегда осуществимо па практике. В крупных очагах и резервациях вредителей, где требуется постоянное регулирующее воздействие на плотность вредной энтомофауны, применение «биоинсектицидов» ограниченно и недостаточно эффективно. Только создание устойчивой и саморазвивающейся системы биологических регуляторов численности вредителей может обеспечить достаточную и экономически оправданную защиту насаждений. В центре такой системы должен находиться вид или группа видов полезных насекомых, которые отвечали бы ряду требований, из которых основными являются следующие:
а) территориальность;
б) эффективность воздействия этих насекомых на
основных вредителей в данном лесном массиве;
в) длительность воздействия на
вредителей в течение сезона;
г) стабильность плотности на протяжении ряда
лет независимо от колебаний численности вредных насекомых;
д) высокая
плотность особей на участке;
е) мультифункциональная положительная роль в
биоценозе;
ж) совместимость (или компенсируемость) действием других факторов
регуляции численности вредителей;
з) простота методов введения данных
насекомых в насаждения.
Всем этим требованиям отвечает только одна группа насекомых - рыжие лесные муравьи.
Безусловно, что создание действенных систем биологической защиты леса возможно только по территориальному принципу, а не применительно к популяции тех или иных вредителей. Поэтому важной характеристикой рыжих ласиых муравьев является их постоянная привязанность к месту поселения и территориальность. Локальные ассоциации муравейников занимают в оптимальных условиях сотни гектаров постоянно контролируемой территории, на которой -муравья способствуют сохранности древостоев.
Плотность поселения муравьев в развитых комплексах гнезд (15-20 м2 оснований куполов на 1 га) может достигать огромных величин - около 1,5•107 особей или свыше 100 кг биомассы на 1 га. Согласно имеющимся данным но энергетике муравьев при такой численности они должны ежедневно получать с гектара угодий не мемее 1 кг белковой пищи. Основную часть белковой пищи муравьи добывают, охотясь на насекомых - вредителей леса, сохраняя тем самым -кроны деревьев от повреждения.
Рыжие лесные муравьи обеспечивают сохранность насаждений от повреждения таких хозяйственно важных хвое- и листогрызущих вредителей, как сосновая совка, сосновая пяденица, зимняя пяденица, малый еловый пилильщик, рыжий сосновый пилильщик, обыкновенный сосновый пилильщик, ткач-пилильщик, березовый пилильщик, сосновый шелкопряд и др. Активно уничтожают рыжие лесные муравьи гусениц дубовой зеленой листовертки, монашенки, непарного шелкопряда, совки-лишайницы, лиственничной минирующей моли и многих других.
Обычно успешность охоты муравьев оценивают по числу приносимых в гнездо насекомых. Однако работа, которую недавно провела под Красноярском В.К.Дмитриенко, показала, что при таком методе учитывается лишь часть истребленных муравьями насекомых. Нападая на различных беспозвоночных, муравьи далеко не всегда тут же овладевают добычей и транспортируют ее в гнездо. Часть насекомых все-таки ускользает от охотников, или муравьи теряют к ним интерес. Но оказывается, что даже непродолжительного контакта гусеницы с фуражирами рыжих лесных муравьев или единственного укуса достаточно, чтобы у гусениц в дальнейшем пропадал аппетит, снижался вес, затягивалось развитие и повышалась смертность. Уже при трехминутном контакте с фуражирами малого лесного муравья погибают все гусеницы непарного шелкопряда. Из гусениц сибирского шелкопряда, контактировавших с муравьями, окукливается не более 10%. Бабочки выходят только из 50% куколок (5% от исходной численности). Небольшие гусеницы и личинки погибают после менее продолжительного воздействия муравьев. Таким образом число погибших от муравьев вредителей значительно превышает число приносимых в муравейник насекомых.
Период активной охотничьей деятельности продолжается у разных лесных муравьев четыре-пять месяцев (в условиях средней полосы Европейской части СССР), что позволяет им охотиться па большинство вредителей во всех доступных фазах развития.
Благодаря трофобиозу с тлями и другими сосущими насекомыми рыжие лесные муравьи получают важную с позиции лесозащиты возможность сохранять высокий уровень численности населения муравейников в периоды депрессии массовых видов насекомых. Плотность поселения тлей-лахнид возрастает вблизи муравейников в несколько раз, что способствует повышению плодовитости и продолжительности жизни более 200 видов паразитических насекомых, использующих падь в качестве дополнительного питания. Кроме того, падь собирают пчелы. Падевый взяток составляет от 30 до 50 % годового сбора меда в ряде европейских стран. С муравьями связана большая группа беспозвоночных-мирмекофилов, что обогащает фауну леса.
Даже при непосредственном взаимодействии с вредителями роль муравьев выходит за рамки прямого хищничества. Крупные насекомые изгоняются муравьями с охраняемой территории: отмечено, что муравьи нападают на бабочек, производящих яйцекладку; в результате количество яиц, отложенных вблизи муравейников, уменьшается. Кроме того, муравьи способствуют распространению среди вредителей различных заболеваний, так как ослабевшие после нападения муравьев гусеницы становятся более восприимчивыми к болезням.
Сами муравьи служат пищей для некоторых позвоночных животных (певчие птицы, пищухи, дятлы и др.), численность которых в лесах, заселенных муравьями, возрастает. Рост численности дятлов и других птиц способствует оздоровлению леса. Через дятлов муравьи опосредованно влияют на плотность стволовых вредителей. Рыжие лесные муравьи являются активными почвообразователями, а также участвуют в расселении ряда растений.
Важной особенностью рыжих лесных муравьев как основы биологического лесозащитного комплекса является характер их взаимодействия с другими полезными для леса животными. Оценка таких взаимодействий лишь по встречаемости тех или иных полезных беспозвоночных в добыче муравьев так же неверна, как и оценка подобным способом влияния муравьев на вредителей. Определение плотности популяций различных хищных и паразитических насекомых в сфере влияния муравейников и вне ее показали, что численность большинства полезных беспозвоночных (бракониды, хальциды, кокцинелиды и др.) вблизи муравейников имеет тенденцию к увеличению. На другие группы полезных беспозвоночных (хищные и паразитические двукрылые, пауки, сетчатокрылые и др.) муравьи не влияют. Численность крупных наездни-ков-ихневмонид и крупных жужелиц в присутствии муравьев начинает снижаться только вокруг самых больших муравейников. Это следствие территориальности воздействия муравьев существенно, так как данный принцип распространяется на все сферы влияния муравьев: при небольшой плотности поселения рыжих лесных муравьев они не в состоянии обеспечить защиту насаждений от вредителей, но и не снижают численности других полезных насекомых. При этом муравьи как фактор смертности вредителей дополняют другие факторы. Если плотность поселения муравьев достаточно высока, численность некоторых крупных паразитов и хищников снижается, но роль этих насекомых компенсируется муравьями. Важно также, что крупных жужелиц и наездников муравьи, как правило, не уничтожают, а лишь изгоняют в нейтральные зоны на стыках охраняемых территорий и за пределы муравьиных колоний. Точно так же воздействуют рыжие лесные муравьи на другие группы муравьев, имеющих значение как энтомофаги и почвообразователи. Эти виды обычно вытесняются на окраины кормовых участков семьи рыжих лесных муравьев. В результате за счет изменения времени охоты и расслоения по ярусам подстилки и растительности в пределах многовидовых ассоциаций муравейников происходит наиболее полное использование кормовых угодий.
При заселении лесов рыжими лесными муравьями сохраняется возможность использования в случае необходимости и микробиологических препаратов, поскольку последние не вызывают заболеваний муравьев.
Несомненное достоинство рыжих лесных муравьев как объекта биометода - простота способов использования их в лесозащите, дешевизна метода. Разработанные и опроби-рованные на практике методы искусственного переселения рыжих лесных муравьев позволяют использовать их как эффективное средство биологической защиты насаждений. Уже в настоящее время реально создание в достаточно короткие сроки (четыре-пять лет) путем искусственных переселений и усиления имеющихся комплексов гнезд необходимой плотности муравьиных поселений на площадях в 100 га и более. Для этого в ряде областей страны накоплен определенный опыт и имеется достаточно большое число муравейников, которые могут служить маточными*. Первые опыты по созданию биологических лесозащитных комплексов уже ведутся.
В Польше под руководством В.Келера были проведены работы но переселению муравьев в комплексе с другими средствами биологической защиты (создание условий для гнездования птиц, размножение паразитов и т. п.), позволившие обезвредить очаги опасного вредителя леса - ткача-пилильщика.
Рыжие лесные муравьи - наиболее реальная основа для биологического лесозащитного комплекса. Не претендуя на универсальность, они в то же время являются звеном, стабилизирующим на высоком уровне плотность большинства полезных насекомых и других полезных для леса животных. В результате прямого и опосредованного воздействия муравьев на лес сохраняются древостой от повреждения вредителей и обеспечивается более высокая продуктивность насаждений. Последнее приобретает все более важное значение в условиях участившихся вспышек массового размножения вредителей леса. Создание и поддержание саморегулирующихся биологических лесозащитных комплексов на основе использования рыжих лесных муравьев совершенно необходимо в ослабленных лесных массивах и резервациях вредителей.
ИСПОЛЬЗОВАНИЕ ПОЛЕЗНЫХ МУРАВЬЁВ
Муравьи начинают оказывать серьезное влияние на биоценоз при условии, что численность их достаточно велика. Однако в лесах гнезда рыжих лесных муравьев размещаются крайне неравномерно. Это происходит в силу различных причин. Прежде всего не каждый биотоп, тип леса благоприятен для жизни этих муравьев. Кроме того, на пути расселяющихся комплексов могут быть непреодолимые для муравьев преграды: реки, болота, разорванность лесных массивов и т. д. Все это имеет значение для рыжих лесных муравьев, ибо основным способом расселения является образование отводков. Мы можем помочь нашим шестиногим союзникам, перенеся искусственные отводки в недоступные им участки леса. Искусственное переселение рыжих лесных муравьев практикуется как в Советском Союзе, так и во многих европейских странах (Польша, ГДР, ФРГ, Италия, Австрия, Швейцария, Финляндия и др.). Но здесь мы сталкиваемся с весьма противоречивой ситуацией.
Дело в том, что количество муравейников повсеместно уменьшается. Основной причиной такого сокращения является человек. Огромный ущерб наносят муравьям сплошные рубки леса, при которых, как правило, уничтожаются все имевшиеся там муравейники. Даже при рубках ухода за лесом разрушается значительная часть гнезд. Муравейники уничтожают браконьеры ради куколок, разрушают туристы и другие отдыхающие. Катастрофически падает численность муравейников вокруг крупных городов и других населенных пунктов, вдоль дорог и туристических маршрутов. Для расселения нужна база - крупные гнезда, откуда можно взять искусственные отводки*, но эту базу мы сами подрываем. Разрушения муравейников, человеком в ряде областей сводит на нет все усилия лесоводов по их расселению.
Значит, использование муравьев невозможио без их охраны, без изменения отношения к ним со стороны хозяйственных органов и всего населения. Охрана муравьев должна стать эффективной уже сейчас, не дожидаясь занесения рыжих лесных муравьев в «Красную книгу». Кое-что в этом направлении уже сделано. Запрещея сбор куколок (так называемых муравьиных яиц), решеяием Совета Министров РСФСР введена материальная ответственность и утверждены таксы по возмещению ущерба лесному хозяйству за разорение муравейников. Постепенно входит в практику вывоз с лесосек главного пользоваяия всех имеющихся там муравейников. Спасенных муравьев переселяют в другие участки леса, ще их нет или численность гнезд слишком мала.
Оптимальная числеяность муравейников рассчитана. В сосияках это четыре активных муравейника диаметром 1,3-1,5 м на гектар леса, в дубравах - шесть-семь гнезд. Муравейников меньших размеров должно быть соответствеино больше.
Отношение общественности к проблеме охраны полезных муравьев изменилось под влиянием проводимой с 1971 г. Всероссийским обществом охраны природы (ВООП) и Министерством лесного хозяйства РСФСР операции «Муравей». В ней участвуют областные организации ВООП и лесхозы многих лесных областей России. Настоящими подвижниками в борьбе за сохранение муравьев стали школьные лесничества, руководимые учителями биологии школ и специалистами лесиого хозяйства. Повсеместно развернута массовая агитация: выпущены массовыми тиражами плакаты и листовки, выставлены тысячи аншлагов. Широко используются радио, телевидение, областная и местная печать.
В ходе операции уже проделана большая работа по учету и инвентаризации муравейников. Оценивается общая численность гнезд, выявляются комплексы, из которых можно брать отводки для промышленного расселения. Как базу переселения рационально использовать только крупные комплексы, так как при взятии отводков из одиночных муравейников в несколько раз возрастает стоимость работ.
Значительно улучшилась охрана муравейников. Большое число гнезд огорожено. Огораживаются муравейники в зеленых зонах городов, в местах массового отдыха, вдоль дорог и туристских троп, в массивах, где обитает большое количество кабанов, разоряющих гнезда. Зеленые патрули, школьные лесничества регулярно наблюдают за сохранностью гнезд, предотвращают их разорение, проводят разъяснительную работу. Активизировалась в этом направлении и деятельность лесной охраны.
Однако сделанное - это пока малая толика того, что необходимо сделать. Инвентаризация, проводимая в большинстве районов, еще не отвечает основной задаче - выявлению базовых комплексов муравейников, их паспортизации и взятию их под специальную охрану. Муравейники должны быть вписавы в книги лесных обходов. Планы комплексов, данные о числе, размерах муравейников и запасе отводков по каждому из них необходимы для всех лесничеств. Без этого невозможно планирование работ по защите, уходу и переселению. Необходимо и участие законодательных органов - пункт об охране муравьев должен быть включен в закон об охране природы. Лишь в таком случае все остальные мероприятия по сохранению и хозяйственному использованию этих полезнейших насекомых наших лесов получат реальную основу для их реализации.
Если мы сможем обеспечить действенную защиту муравьиным поселениям - это уже большое дело. Решение задачи сдерживается рядом обстоятельств, к которым относятся и такие первостепенные вопросы, как подготовка кадров, а также общая культура ведения лесного хозяйства. Без специалистов, знающих биологию муравьев, умеющих различать виды и освоивших технику переселения, рассчитывать на успех не приходится. Между тем специальный курс по использованию муравьев имеется лишь в некоторых лесных вузах и вообще отсутствует в техникумах и лесных школах. Не проводятся занятия по биологии и применению муравьев на курсах повышения квалификации лесопатологов и лесничих. Таким образом, на быстрое разрешение проблем, связанных с использованием муравьев, рассчитывать не приходится.
Правда, за последние годы в РСФСР, Белоруссии, Эстонии и на Украине выпущены рекомендации по переселению муравейников, что должно помочь проведению подобных работ. Но без соответствующих семинаров и хотя бы кратких курсов по практическому применению таких рекомендаций и руководств правильное использование информации не гарантировано. Всегда после выхода новых постановлений по какому-нибудь вопросу в соответствующей отрасли проводятся семинары и разъяснительная работа по их применению. Муравьи в этом плане не должны быть исключением. Тем более, что применение муравьев в лесозащите - дело новое, во многих местах еще только начинающееся. Пускать его на самотек недопустимо.
Сейчас накоплены значительные знания по всем основным вопросам, связанным с использованием муравьев в защите леса и для повышения его продуктивности. Большая работа в этом направлении проводится в ряде европейских стран. Значительный вклад в разработку биологических основ и методов использования полезных муравьев внесли советские энтомологи и лесоводы. Но внедрение этих знаний и практических разработок в лесное хозяйство зависит от того, насколько серьезно и последовательно будет проводиться соответствующая работа лесными ведомствами. Программа по охране и использованию муравьев должна принять форму постоянной, развивающейся и совершенствующейся сферы деятельности лесохозяйственных организаций.
Охрана муравьев - не частный вопрос: рыжие лесные муравьи - одно из важнейших звеньев, обеспечивающих жизнеспособность и продуктивность леса. Лишиться этого звена - значит оборвать, нарушить множество естественных связей, поддерживающих благополучие леса, обеднить и ослабить биогеоценоз, затормозить его развитие; значит ударить по популяциям полезных насекомых и птиц, открыть дорогу вредителям, потерять прирост деревьев, а в ряде случаев и весь древостой. Наконец, нельзя лишить человека радостной возможности понаблюдать за уникальными во многих отношениях созданиями, сумевшими преодолеть ряд самых жестоких ограничений, налагаемых малым ростом, твердым покровом, мимолетностью жизни индивида и другими следствиями принадлежности муравьев к классу насекомых. Присмотревшись к муравьям, человек еще раз удивится изобретательности и многообразию жизни, ее настойчивости и неодолимому стремлению к совершенству. Мы не можем и не должны лишиться всего этого.
Необходимы значительные усилия, чтобы помочь муравьям восстановить былую численность и вернуться в те леса, где они исчезли по нашей вине. Но мероприятия по охране и искусственному расселению рыжих лесных муравьев - не просто благотворительность, как это иногда пытаются представить. И не широкий жест хозяина, снисходительно высыпающего хлебные крошки воробьям. Муравьи приносили и приносят большую пользу лесу и человеку. Забота о них - забота о благополучии человека. Другое дело, что испокон веков повелось, что муравьи ничьи, что им числа нет и их не убудет, а если и убудет, так и беда невелика.
Когда-то приисковый метод господствовал в деятельности человека в лесу. Теперь на смену ему пришло настоящее хозяйствование. А вот в отношении лесных муравьев приисковый подход сохраняется, даже при работах по искусственному переселению. Ведь отводки берутся зачастую как попало, наблюдений за последующим состояиием муравейников, из которых взяты отводки, нет. Взяли, погрузили, перевезли, выполнили план. Даже те скромные правила использования комплексов маточных муравейников, что изложены в руководствах по переселению, не выполняются. Там, например, предусмотрено, что эксплуатация любого комплекса начинается с его инвентаризации, оценки запаса отводков, состояния гнезд и составления плана эксплуатации. Практически ни одна из предварительных работ не выполняется. Просто в места, где «много муравьиных куч», приезжает машина. Из тех гнезд, что поближе, берутся отводки и увозятся в другое место.
А ведь взятие отводков по той или иной схеме может по-разному сказаться на последующем состояния оставшихся. Правильным донорством можно стимулировать дальнейшее развитие комплекса, рост гнезд. В этом случае отводки можно брать из одних и тех же гнезд раз в четыре года, причем запас отводков от раза к разу не только не сокращается, но и увеличивается. Периодическое донорство активизирует семьи, дает толчок к их росту.
Неправильное взятие отводков нарушает структуру комплекса, подрывает перспективные гнезда и муравейники, являющиеся основными производителями естественных отводков. Это ведет к истощению запаса муравьев в исходном месте. Именно так проводится зачастую переселение. Подобиого отношения к этому делу пока что не в состоянии изменить многочисленные рекомендации, инструктивные письма и разъяснения.
В чем дело? Разве самому предприятию, проводящему переселение, не выгодно иметь дело с постоянными тремя-четырьмя комплексами, регулярно дающими материал для переселения? Несомненно, это наиболее рациональный путь. Дополнительные хлопоты по инвентаризации и оценке комплексов необходимы, ибо без них иельзя вести муравьиное хозяйство.
Может быть лесхоз не имеет других стимулов к разведению муравьев, кроме директив свыше? В ряде случаев это так и есть. Ведь муравейники все еще не имеют товарной стоимости. Куколки на рынке - имеют, муравьиный спирт в аптеке - имеет. Появляется стоимость и у сломанного браконьером или хулиганом гнезда. А вот процветающий, дающий реальную прибыль муравейник ничего не стоит. В расходах на переселение стоимость самого отводка не значится. Зарплата рабочим, автомашина, тара, сахар для подкармливания, сломанная в ходе работы лопата - это все оценено, а отводок - нет. Бесплатный, выросший сам по себе муравейник, бесплатный отводок. Может быть поэтому и не доходят руки до их инвентаризации и хозяйственного использования. Но ведь деревья, выросшие без помощи человека, все же имеют вполне определенную цену. Почему же нельзя оценить муравейник? Тогда отводки станут реальной продукцией, и лесхозы будут стремиться сохранять и использовать своих муравьев. Наравне с другими видами продукции леса материал для переселения станет давать прибыль этим предприятиям. Появится заинтересованность в ведеиии муравьиного дела. Обоснованным станет участие в инвентаризации муравейников лесоустроительных и лесопатологических экспедиций. Каждый комплекс муравейников обретает вполне определенное стоимостное выражение. Сохранение его превратится из дополнительной нагрузки в узаконенную обязанность. Да и отношение к результатам переселения тоже изменится.
Откроется возможность и для создания настоящих муравьиных хозяйств, специализирующихся на разведении муравьев, выращивании муравейников для получения искусственных отводков, куколок, муравьиного спирта. Методы получения этих продуктов не скажутся отрицательно на состоянии используемых гнезд и обеспечат рентабельное использование муравьев для нужд человека.
Основные теоретические предпосылки для организации муравьиных ферм уже имеются. Дело опять за ведомствами, заинтересованными в получении продукции, даваемой муравьями, но не желающими пока заниматься этим новым, хлопотным делом. Но иного пути нет. Необходимо преодолеть инерцию, недостаточность правовых и экономических основ использования рыжих лесных муравьев, а также снисходительное пренебрежение иных хозяйственных руководителей различных рангов. Такие смотрят на все, касающееся возможности использования муравьев, как на преходящую игру, лишь отрывающую солидных людей от неотложных дел. Да что он может ваш муравей, говорят они со снисходительной улыбкой, и показывают на кончике мизинца размеры муравьяшки. Размер муравья невелик - здесь все правильно. Но вот ведь в чем дело: муравей-то мал, да муравейник велик, и велика польза, приносимая лесными муравьями. Можно обсуждать детали, но лишь затем, чтобы научиться оберегать и наиболее эффективно использовать одних из самых надежных наших союзников и помощников в лесу.
КРАТКИЙ СЛОВАРЬ МИРМЕКОЛОГИЧЕСКИХ ТЕРМИНОВ
(Здесь употребляются термины как общейввестныё, так и предлагаемые автором.)
Активация семьи (фуражиров) - общая активизация фуражировочной деятельности семьи при обнаружении на участке добычи.
Буферные гнезда - постоянные гнезда, сооружаемые при объединении колоний в федерации для предотвращения прямого взаимодействия материнских муравейников.
Вид-хозяин - вид, семьи которого используются муравьями - социальными паразитами для основания семьи или постоянного обитания.
Внутренний конус - центральная часть наземного купола у рыжих лесных муравьев. Основное место выделения молоди и обменов между колоннами.
Временный социальный паразитиам - этап основания семей видов, самки которых неспособны к самостоятельному основанию семьи, на семьях видов-хозяев.
Вспомогательные гнезда - временные небольшие гнезда, служащие для использования кормового участка (станции, павильоны, кормовые почки) и породнения муравейников (промежуточные почки).
Вторичное деление территории - подразделение кормового участка семьи на элементарные поисковые участки, обслуживаемые одним фуражиром или небольшими группами.
Гнездовой двор - прилегающая к гнезду часть территории, на которой у крупных муравейников происходит формирование потоков фуражиров. Обычно лишен растительности.
Гнездовой запах (sanax семьи) - специфический запах, служащий основой для разделения особей одного вида по принципу «свой - чужой».
Гнездовая капсула - часть гнезда с высокой концентрацией камер, имеющая защитный слой и выраженную дифференциацию отделов.
Гнездо секционное - гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.).
Динамическая плотность особей - интенсивность посещения муравьями единицы площади в единицу времени (особь/дм2хмин.).
Диффузные гнезда - форма поликалии, при которой семья рассредоточена по готовым камерам и небольшим гнездам, не имеющим постоянного состава. Функциональной дифференциации таких гнезд и камер нет.
Дочерняя семья (дочерний муравейник) - семья, образованная путем почкования материнской семьи.
Зона взаимного проникновения-область перекрывания охраняемых территорий муравейников разных видов. Характеризуется низкой динамической плотностью особей и лояльностью фуражиров из этих муравейников.
Зона монопольного использования-центральная часть охраняемой территории муравейника (доминанта), которая охраняется от проникновения фуражиров из соседних территориальных муравейников других видов.
Иерархия функциональных групп (групповая иерархия) - система соподчинения особей, основанная на их принадлежности к той или иной полиэтичеокой или функциональной группе.
Искусственный отводок - часть муравейника (семьи), взятая для искусственного переселения.
Кинопсис - восприятие муравьями в качестве сигнала характерных движений других особей одной общины («язык поз»).
Колония - функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения.
Колонна - организационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
Комплекс муравейников - группа муравейников одного вида, выделяемая по территориальному признаку.
Кормовой участок - участок, используемый муравьями для добычи пищи.
Кормовая дорога - дорога, используемая муравьями для выхода в зону фуражировки и транспортировки добычи.
Кратер - земляной вал при выходе из жилого муравейника (секции), не имеющего наземного купола.
Материнский муравейник - муравейник, образовавшийся почкованием отводок.
Маточный муравейник - муравейник, из которого взят искусственный отводок.
Мобилизация - индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
Моногиния - наличие в семье одной яйцекладущей самки.
Многовидовая ассоциация муравейников - территориальная иерархическая группировка муравейников разных видов, обладающая устойчивой структурой, которая поддерживается поведенческими механизмами.
Нейтральное пространство (нейтральная зона) - пространство между охраняемыми территориями семей или колонн, не посещаемое фуражирами из конкурирующих семей (колонн).
Обменная дорога - дорога, связывающая родственные муравейники.
Обменный фонд муравейника - максимальное число особей, которое может единовременно обменять семья, сохраняя индивидуальность.
Отводок - дочерний муравейник: гнездо, в котором обитает дочерняя семья (в колонии).
Охраняемая территория - участок, охраняемый семьей (колонной) от посещения чужими особями своего или близких видов.
Полигиния - наличие в семье нескольких яйцекладущих самок.
Поликалия - обитание одной семьи в нескольких гнездах.
Полиэтизм - выполнение муравьем в течение длительного времени определенных функций в семье. Если функция связана только с возрастом - возрастной полиэтизм; если функция выполняется постоянно - постоянный, или «кастовый», полиэтизм.
Полиэтические группы - особи, объединяемые сходством психофизиологических данных и предрасположенные к выполнению соответствующих функций (например, активные и пассивные фуражиры).
Почкование - образование нового муравейника отделением небольшой части материнской семьи. Обычно ведет к возникновению колонии.
Семья - основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Сектор - часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
Тактильные сигналы (тактильный код) - сигналы, передаваемые муравьями прикосновением антенн, щупиков, лапок при непосредственном контакте особей.
Трофаллаксис - передача пищи и гормональных веществ от особи к особи путем кормления содержимым зобика, желудка или слизывания выпота с поверхности тела.
Трофобиоз - взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
Федерация - объединение нескольких колоний, поддерживающих родственные отношения посредством обменов особями.
Функциональные группы - устойчивые группы особей, занятых в семье выполнением определенных функций. Входят в состав полиэтических групп.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
ЛАТИНСКИЕ И РУССКИЕ НАЗВАНИЯ УПОМЯНУТЫХ В ТЕКСТЕ МУРАВЬЕВ
Acantholepis melanogaster - акантолепис
Aphaenogaster senilis - афеногастер
Atta - муравьи-листорезы
Camponotus - кампонотусы
- herculeanus - красногрудый муравей-древоточец
Cataglyphis aenescens - черный бегунок
Cataglyphis pallida - бледный бегунок
Cataglyphis setipes - муравей-фаэтончик
Coptoformica - тонкоголовые муравьи
Crematogaster - остробрюхие муравьи
Dolichoderus quadripunctatus - пятнистый муравей
Dorylinae - муравьи-кочевники
Formica - формика
- aquilonia - северный лесной муравей
- cinerea - серый песчаный муравей
- cunicularia - прыткий степной муравей
- fusca - бурый лесной муравей
- lugubris - волосистый лесной муравей
- polyctena - малый лесной муравей
- pratensis - луговой муравей
- rufa - обыкновенный лесной муравей (Группа Formica rufa (F. aquilonia, F. lugubris, F. rufa, F. polyctena) - рыжие лесные муравьи.)
- rufibarbis - краснощекий муравей
- sanguinea - кроваво-красный муравей-рабовладелец
- truncorum - красноголовый муравей
- uralensis - черноголовый муравей
Formicoxenus nitidulus - блестящий муравей-малютка
Lasius - лазиусы
- flavus - желтый земляной муравей
- fuliginosus - пахучий муравей-древоточец
- niger - черный садовый муравей
- umbratus - желтый пахучий муравей
Messor - муравьи-жнецы
- aralocaspius - аралокаспийский муравей-жнец
- intermedius - средний муравей-жнец
Мопошогшш gracillinum - изящный муравей
Monomorium pharaonis - фараонов муравей
Myrmecia - муравьи-бульдоги
Myrmica - мирмика
Oecophylla - муравьи-портные
Pheidole pallidula - феидолия
Polyergus rufescens - муравей-амазонка
Proformica epinotalis- степной медовый муравей
- Tetramarium - тетраморйумы
- caespitum - дерновый муравей
- schneideri - пустынный дерновый муравей
- turcomanica - степной дерновый муравей
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Длусский Г. М. Муравьи рода формика. М.: Наука, 1967.
Захаров А. А. Внутривидовые отношения у муравьев. М.: Наука, 1972.
Кастлер Г. Возникновение биологической организации. М.: Мир, 1967.
Сетров М. И. Организация биосистем. Л.: Наука, 1971.
Тинберген Н. Поведение животных. М.: Мир, 1969.
Фриш К. Из жизни пчел. М.: Мир, 1966.
Хайнд Р. Поведение животных. М.: Мир, 1975.
Халифман И. А. Муравьи. М.: Молодая гвардия, 1967.
Шовен Р. От пчелы до гориллы. М.: Мир, 1965.
СОДЕРЖАНИЕ
Введение
1. Муравей и муравейник .............. 5
Состав семьи
.................. 6
Профессии муравьев ............... 10
Взаимодействие муравьев в семье .......... 19
2. Структура муравейника
.............. 29
Обособленность колонн .............. 29
Факторы
сохранения целостности семьи ....... 37
Сравнимость колонн по численности
населения .... 43
Колонна - муравейник в муравейнике ........ 51
Семья
как система колонн ............. 56
Поликалия у муравьев .............. 58
3. Муравейник и колония .............. 65
Возникновение колонии
.............. 66
Материнская и дочерняя семьи ........... 70
Структура
колонии ................ 79
Колония и федерация ............... 89
4.
Развитие общинного образа жизни у муравьев .... 97
Рост семьи и
совершенствование ее организации . . . 98
Рост приспособительных
возможностей семьи ..... 108
5. Перспективы муравьеводства ............ 118
Чем полезны муравьи ............... 119
Рыжие лесные муравьи как основа
биологического лесозащитного комплекса ............. 126
Использование
полезных муравьев .......... 131
Краткий словарь мирмекологических терминов
...... 138
Латинские и русские названия упомянутых в тексте муравьев
...................... 141
Рекомендуемая литература .............. 142
Анатолии Александрович Захаром МУРАВЕЙ, СЕМЬЯ, КОЛОНИЯ
Утверждено к
печати редколлегией серии научно-популярных изданий Академии наук СССР
Редактор издательства В. Н. Вяземцеве Художник Б. Е. Захаров Художественный
редактор И. В. Разина Технический редактор Ф.М.Хенох Корректоры М.С.Бочарова, Б.
И Рывин
И Б № 7375
Сдано в набор 2,03.78. Подписано к печати 10.08.78
Т-14036. Формат 84х1б8 /„ Бумага типографская Ni 3 Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 6,3. Уч.-изд. л. 7,8 Тираж 50000 экз. Тип. зак.
208 Цена 45 коп.
Издательство «Наука»
117485, Москва, В-485, Профсоюзная
ул., 94а Ордена Трудового Красного Знамени Первая типография издательства
«Наука» 199034, Ленинград, В-34, 9 линия, д. 12