ГЛАВА 7. ЭНЕРГЕТИЧЕСКИЙ МЕТАБОЛИЗМ ПРОКАРИОТ
Энергетические процессы прокариот по своему объему (масштабности) значительно превосходят процессы биосинтетические, и протекание их приводит к существенным изменениям в окружающей среде. Разнообразны и необычны в этом отношении возможности прокариот, способы их энергетического существования. Все это вместе взятое сосредоточило внимание исследователей в первую очередь на изучении энергетического метаболизма прокариот.
ЭНЕРГЕТИЧЕСКИЕ РЕСУРСЫ
Организмы могут использовать не все виды энергии, существующей в природе. Недоступными для них являются ядерная, механическая, тепловая виды энергии. Чтобы теплота могла служить источником энергии, необходим большой перепад температур, который в живых организмах невозможен. Доступными для живых систем внешними источниками энергии (энергетическими ресурсами) являются электромагнитная (физическая) энергия (свет определенной длины волны) и химическая (восстановленные химические соединения). Способностью использовать энергию света обладает большая группа фотосинтезирующих организмов, в том числе и прокариот, имеющих фоторецепторные молекулы нескольких типов (хлорофиллы, каротиноиды, фикобилипротеины). Для всех остальных организмов источниками энергии служат процессы окисления химических соединений.
Часто энергетическими ресурсами служат биополимеры, находящиеся в окружающей среде (полисахариды, белки, нуклеиновые кислоты), а также липиды. Прежде чем быть использованными, биополимеры должны быть гидролизованы до составляющих их мономерных единиц. Этот этап весьма важен по следующим причинам. Белки и нуклеиновые кислоты отличаются исключительным разнообразием. Количество видов белков исчисляется тысячами, после гидролиза же образуется только 20 аминокислот. Все разнообразие нуклеиновых кислот (ДНК и РНК) после гидролиза сводится к 5 видам нуклеотидов. Таким образом, расщепление полимеров до мономерных единиц резко сокращает набор химических молекул, которые могут быть использованы организмом.
Полимерные молекулы расщепляются до мономеров с помощью ферментов, синтезируемых и выделяемых прокариотами в окружающую среду (экзоферментов). Крахмал и гликоген гидролизуются амилазами, гликозидные связи целлюлозы расщепляются целлюлазой. Многие бактерии образуют пектиназу, хитиназу, агаразу и другие ферменты, гидролизующие соответствующие полисахариды и их производные. Белки расщепляются внеклеточными протеазами, воздействующими на пептидные связи. Нуклеиновые кислоты гидролизуются рибо- и дезоксирибонуклеазами. Образующиеся небольшие молекулы легко транспортируются в клетку через мембрану.
Процесс распада жирных кислот локализован в клетке и включает несколько этапов. На первом из них жирная кислота с помощью соответствующего фермента превращается в КоА-производное, которое окисляется в b-положении с последующим отщеплением ацетил-КоА. Другим продуктом реакции является КоА-производное жирной кислоты, укороченное на два углеродных атома. Ацетил-КоА по катаболическим каналам используется для получения клеткой энергии.
Процесс расщепления биополимеров не связан с образованием свободной, т. е. доступной клетке энергии19. Происходящее при этом рассеивание энергии также невелико. Образовавшиеся мономеры подвергаются в клетке дальнейшим ферментативным превращениям, которые сводятся к тому, чтобы путем перестройки химической структуры получить молекулы, которые могли бы включиться на каком-либо этапе в качестве метаболитов в функционирующие клеточные катаболические системы. Таких в прокариотной клетке несколько. Основные из них: путь Эмбдена — Мейергофа — Парнаса (гликолиз), окислительный пентозофосфатный путь, путь Энтнера — Дудорова и цикл трикарбоновых кислот (ЦТК). Общее для всех катаболических путей — многоступенчатость процесса окисления исходного субстрата. На некоторых этапах окисление субстрата сопряжено с образованием энергии в определенной форме, в которой эта энергия может использоваться в самых разнообразных энергозависимых процессах.
19 Свободная энергия (DG) — это та часть энергии, которая может быть превращена в работу. При протекании химических реакций в живом организме самопроизвольно идут те процессы, в которых изменение свободной энергии будет отрицательным (–DG). Такие процессы называются экзергоническими. Процессы, для которых DG является величиной положительной, называются эндергоническими. Эти процессы не могут происходить самопроизвольно. При протекании эндергонических процессов необходим приток энергии извне.
Для каждой химической реакции характерно определенное изменение стандартной свободной энергии (DG0'), т. е. изменение свободной энергии при стандартных значениях температуры и давления, 1М концентрации исходных веществ и продуктов реакции и pH 7,0. Например, DG0' гидролиза АТФ до АДФ и ФН равняется 31,8 кДж/моль (при 1М концентрации исходных веществ и продуктов реакции, температуре 37°, pH 7,0 в присутствии избытка ионов Mg2+). Для биолога важно, что по такому параметру, как изменение свободной энергии, он может осуществлять анализ биологических процессов.
Таким образом, внешние доступные организмам источники энергии (свет, химические соединения) должны быть трансформированы в клетке в определенную форму, чтобы обеспечить внутриклеточные потребности в энергии.
ОБЩАЯ ХАРАКТЕРИСТИКА ЭНЕРГЕТИЧЕСКИХ ПРОЦЕССОВ
В самом общем виде процессы, способные служить источником энергии для прокариот, можно представить следующим образом:
А ® В + е– .
Например,
Fe2+ ® Fe3+ + е– ; (1)
-CH2-CH2- ® -CH2=CH2- + 2e– ; (2)
СН4 + 1/2 O2 ® СН3ОН . (3)
В первой реакции окисление иона двухвалентного железа — это потеря электрона. Во втором примере окисление углеродного субстрата можно в равной мере рассматривать как отрыв от него водорода (дегидрирование) или независимое удаление двух протонов (Н+) и электронов (e–). В биохимических процессах, как правило, перенос водорода осуществляется путем раздельного транспорта протонов и электронов: протоны выделяется в среду и при необходимости поглощаются из нее, электроны обязательно должны быть переданы на соответствующие молекулы Поэтому все окислительно-восстановительные превращения определяются по существу "перемещениями" электронов. В последнем примере имеет место присоединение атома кислорода к молекуле субстрата. Окислительно-восстановительный характер реакции в этом случае не столь очевиден, как в предыдущих, поскольку не происходит отрыва электрона (водорода) от молекулы метана. В данном случае в результате окисления метана происходит замена связи C-H на связь C-OH, кислород оттягивает электроны от атома углерода, подвергшегося окислению, сам при этом восстанавливаясь. Таким образом, внутри молекулы происходит "деление" электронной пары между атомами углерода и кислорода, т. е. окислительно-восстановительные перестройки. Реакции, в которых имеется возможность отрыва электронов, могут быть использованы прокариотами для получения энергии.
Разнообразные соединения, способные окисляться, т. е. являющиеся источниками отрываемых электронов, называются донорами электронов. Поскольку электроны не могут существовать самостоятельно, они обязательно должны быть перенесены на молекулы, способные их воспринимать и, таким образом, восстанавливаться. Такие молекулы называются акцепторами электронов. Какие ограничения здесь возможны? Донором электронов не может быть предельно окисленное вещество, а их акцептором — предельно восстановленное. Таким образом, должен существовать внешний энергетический ресурс — исходный субстрат. С помощью ферментных систем организм извлекает энергию из этого субстрата в реакциях его ступенчатого окисления, приводящего к освобождению энергии небольшими порциями.
У прокариот известны три способа получения энергии: разные виды брожения, дыхания и фотосинтеза. В процессах брожения в определенных окислительно-восстановительных реакциях образуются нестабильные молекулы, фосфатная группа которых содержит много свободной энергии. Эта группа с помощью соответствующего фермента переносится на молекулу АДФ, что приводит к образованию АТФ. Реакции, в которых энергия, освобождающаяся на определенных окислительных этапах брожения запасается в молекулах АТФ, получили название субстратного фосфорилирования. Их особенностью является катализирование растворимыми ферментами. Образующийся в восстановительной части окислительно-восстановительных преобразований сбраживаемого субстрата восстановитель (НАД·H2, восстановленный ферредоксин) переносит электроны на подходящий эндогенный акцептор электрона (пируват, ацетальдегид, ацетон и др.) или освобождается в виде газообразного водорода (H2).
Нередко в процессах брожения окислительные и восстановительные преобразования могут происходить внутримолекулярно, т. е. одна часть образуемой молекулы подвергается восстановлению, другая — окислению. В ряде брожений восстановительное и окислительное превращение связано с разными образующимися продуктами брожения, т. е. происходит межмолекулярно.
Многие прокариоты получают энергию в процессе дыхания. Они окисляют восстановленные вещества с относительно низким окислительно-восстановительным потенциалом (E0), возникающие в реакциях промежуточного метаболизма или являющиеся исходными субстратами, например НАД·H2, сукцинат, лактат, NH3, H2S и др. (табл. 11).
Таблица 11. Окислительно-восстановительные потенциалы (E0', мВ) веществ, участвующих в энергетических процессах у прокариот
| Окислительно-восстановительная система | E0', мВ |
| Пируват/ацетат + CO2 | –700 |
| H+ + 1/2H2 | –420 |
| Ферредоксин окисл/восст (из Clostridium pasteurianum)* | –420 |
| НАД(Ф)+/НАД(Ф)·H2 | –320 |
| S0/HS– | –270 |
| SO42–/HS– | –220 |
| ФАД/ФАД·H2 | –220 |
| ФМН/ФМН·H2 | –190 |
| Менахинон окисл/восст | –74 |
| Рубедоксин окисл/восст | –57 |
| Фумарат/сукцинат | +30 |
| Цитохром b окисл/восст | +70 |
| Убихинон окисл/восст | +100 |
| Цитохром c окисл/восст | +220 |
| Цитохром a окисл/восст | +290 |
| NO3–/NO2– | +433 |
| Fe3+/Fe2+ | +772 |
| 1/2 O2/H2O | +820 |
* E0' ферредоксинов Fe2S2-типа у разных прокариот имеет значения от –225 до –455 мВ, а Fe4S4-типа — от + 350 до –480 мВ.
E0 характеризует способность определенных веществ быть донорами или акцепторами электронов. Он может быть измерен экспериментально Для любой окислительно-восстановительной системы. В соответствии с полученными значениями различные вещества образуют определенную шкалу окислительно-восстановительных потенциалов, которые принято отсчитывать относительно окислительно-восстановительного потенциала реакции H2«2H+ + 2e– . Стандартное значение его при pH 7 (E0') равно –420 мВ.
Высокая отрицательная величина E0 водорода говорит о его активной восстановительной способности, т. е. способности отдавать электроны. Чем больше отрицательная величина E0 определенной окислительно-восстановительной системы, тем выше ее восстановительная способность и наоборот. ео системы H2O«1/2O2 + 2H+ + 2e– равен+ 820 мВ. Большая положительная величина потенциала объясняет слабую способность воды отдавать электроны и одновременно высокую способность молекулярного кислорода акцептировать электроны. Таким образом, в соответствии со значениями окислительно-восстановительных потенциалов для двух или нескольких окислительно-восстановительных систем электроны без подведения энергии извне будут перемещаться в направлении от более электроотрицательных систем к более электроположительным.
Окисление происходит в результате переноса электронов через локализованную в мембране дыхательную электронтранс-портную цепь, состоящую из набора переносчиков, и приводит в большинстве случаев к восстановлению молекулярного кислорода до H2O. Таким образом, в процессе дыхания молекулы одних веществ окисляются, других — восстанавливаются, т. е. окислительно-восстановительные процессы в этом случае всегда межмолекулярны.
Наиболее широко распространена среди прокариот способность окислять органические субстраты. Обнаружены также весьма специализированные группы прокариот, способные окислять различные неорганические субстраты (H2, NH4+, NO2–, H2S, S0, S2O32–, Fe2+ и др.) с соответствующим восстановлением O2. Наконец, прокариоты могут окислять органические и неорганические вещества с использованием в качестве конечного акцептора электронов не молекулярного кислорода, а целого ряда органических и неорганических соединений (фумарат, CO2, NO3–, S0, S042–, S0, S032– и др.). Количество освобождающейся энергии определяется градиентом окислительно-восстановительных потенциалов при переносе электронов от донора к акцептору. Так, окисление H+ молекулярным кислородом сопровождается освобождением значительно большего количества свободной энергии (DG0' = –238 кДж/моль), чем окисление НАД·H+ фумаратом (DG0' = –68 кДж/моль).
У прокариот известны три типа фотосинтеза: I — зависимый от бактериохлорофилла бескислородный фотосинтез, осуществляемый группами зеленых, пурпурных бактерий и гелиобактерий; II — зависимый от хлорофилла кислородный фотосинтез, свойственный цианобактериям и прохлорофитам; III — зависимый от бактериородопсина бескислородный фотосинтез, найденный у экстремально галофильных архебактерий. В основе фотосинтеза I и II типа лежит поглощение солнечной энергии различными пигментами, приводящее к разделению электрических зарядов, возникновению восстановителя с низким и окислителя с высоким окислительно-восстановительным потенциалом. Перенос электронов между этими двумя компонентами приводит к выделению свободной энергии. В фотосинтезе III типа окислительно-восстановительные переносчики отсутствуют. В этом случае энергия в доступной для организма форме возникает в результате светозависимого перемещения H+ через мембрану.
Изучение у прокариот электронтранспортных цепей, функционирующих в процессах дыхания и фотосинтеза I и II типа, выявило принципиальное сходство между ними. В обеих системах электронного транспорта есть флавопротеины, хиноны, цитохромы и белки, содержащие негемовое железо, позволяющие переносить электроны вниз по термодинамической лестнице. Таким образом, по существу обе электронтранспортные цепи являются окислительными. Разнообразие в их организации обнаружено при более детальном изучении и выражается как в широком наборе доноров и акцепторов электронов, так и в конкретной организации самих цепей: химическом строении переносчиков, принадлежащих к одному типу, их наборе, расположении и т. д.
В процессах дыхания и фотосинтеза освобождающаяся при переносе электронов энергия запасается первоначально в форме электрохимического трансмембранного градиента ионов водорода (DmH+), т. е. имеет место превращение химической и электромагнитной энергии в электрохимическую. Последняя затем может быть использована для синтеза АТФ. Поскольку в обоих процессах синтез АТФ обязательно связан с мембранами, реакции, приводящие к его образованию, получили название мембранзависимого фосфорилирования. Последнее подразделяется на два вида: окислительное (АТФ образуется в процессе электронного переноса при окислении химических соединений) и фотосинтетическое (синтез АТФ связан с фотосинтетическим электронным транспортом) фосфорилирование. Следует подчеркнуть, что принципы генерации АТФ при фотосинтезе и дыхании, т. е. механизмы мембранзависимого фосфорилирования, одинаковы. Таким образом, энергия, получаемая в процессах брожения, дыхания или фотосинтеза, запасается в определенных формах.
Существуют две универсальные формы энергии, которые могут быть использованы в клетке для выполнения разного рода работы: энергия высокоэнергетических химических соединений (химическая) и энергия трансмембранного потенциала ионов водорода (электрохимическая).
ВЫСОКОЭНЕРГЕТИЧЕСКИЕ СОЕДИНЕНИЯ. АТФ — УНИВЕРСАЛЬНАЯ ФОРМА ХИМИЧЕСКОЙ ЭНЕРГИИ В КЛЕТКЕ
У прокариот существует несколько типов богатых энергией химических соединений. Самую большую группу составляют соединения с высокоэнергетической фосфатной связью: ацилфосфаты, фосфорные эфиры енолов (фосфоенолпируват), нуклеотидди- и трифосфаты, аденозинфосфосульфат. Другая распространенная группа — соединения с высокоэнергетической тиоэфирной связью — ацилтиоэфиры.
Эти соединения характеризуются тем, что по крайней мере одна из входящих в состав молекулы групп имеет высокий энергетический потенциал. При переносе этой группы происходит разрыв связи, соединяющей ее с молекулой, что приводит к резкому уменьшению свободной энергии, заключенной в молекуле химического соединения. Такие связи называются высоко-энергетическими, или макроэргическими. Присоединение группы с высоким энергетическим потенциалом к молекуле-акцептору повышает уровень ее свободной энергии, переводя, таким образом молекулу в активированную форму, в которой это соединение может участвовать в биосинтетических реакциях.
Центральное место в процессах переноса химической энергии принадлежит системе АТФ. АТФ образуется в реакциях субстратного и мембранзависимого фосфорилирования. При субстратном фосфорилировании источником образования АТФ служат реакции двух типов:
I. Субстрат ~ Ф20 + АДФ « субстрат + АТФ;
II. Субстрат ~ X + АДФ + ФцН « субстрат + Х + АТФ.
20 Символ "~", введенный американским биохимиком Ф. Липманом (F. Lipmann), служит для обозначения макроэргической связи.
В реакциях первого типа осуществляется перенос высокоэнергетической фосфатной группы от молекулы-донора на АДФ катализируемый соответствующими киназами. Реакциями такого типа являются реакции субстратного фосфорилирования на пути анаэробного превращения Сахаров. У прокариот, имеющих ЦТК, реакция превращения сукцинил-КоА в янтарную кис лоту сопровождается запасанием энергии в фосфатной связь ГТФ, который затем отдает фосфатную группу АДФ. Эту реакцию можно рассматривать как реакцию субстратного фосфорилирования второго типа.
АТФ образуется также за счет энергии Dm в процессе мембранзависимого фосфорилирования. В общих чертах этот механизм фосфорилирования изложен в следующем разделе.
Молекула АТФ содержит две макроэргические фосфатные связи, при гидролизе которых высвобождается значительное количество свободной энергии:
АТФ + H2 ® АДФ + ФН; DG0' = –31,8 кДж/моль;
АДФ + H2 ® АМФ + ФН; DG0' = –31,8 кДж/моль;
Отщепление последней фосфатной группы от молекулы АМФ приводит к значительно меньшему высвобождению свободной энергии:
АМФ + H2 ® аденозин + ФН; DG0' = –14,3 кДж/моль;
Молекула АТФ обладает определенными свойствами, которые и привели к тому, что в процессе эволюции ей была отведена столь важная роль в энергетическом метаболизме клеток. Термодинамически молекула АТФ нестабильна, что вытекает из большой отрицательной величины DG ее гидролиза. В то же время скорость неферментативного гидролиза АТФ в нормальных условиях очень мала, т. е. химически молекула АТФ высокостабильна. Последнее свойство обеспечивает эффективное сохранение энергии в молекуле АТФ, поскольку химическая стабильность молекулы препятствует тому, чтобы запасенная в ней энергия бесполезно рассеивалась в виде тепла. Малые размеры молекулы АТФ позволяют ей легко диффундировать в различные участки клетки, где необходим подвод энергии извне для выполнения химической, осмотической, механической работы.
И наконец, еще одно свойство молекулы АТФ, обеспечившее ей центральное место в энергетическом метаболизме клетки. Изменение свободной энергии при гидролизе АТФ составляет — 31,8 кДж/моль. Если сравнить эту величину с аналогичными величинами для ряда других фосфорилированных соединений, то мы получим определенную шкалу. На одном из ее полюсов будут расположены фосфорилированные соединения, гидролиз которых приводит к высвобождению значительного количества свободной энергии (высокие отрицательные значения DG). Это так называемые "высокоэнергетические соединения". На другом полюсе будут располагаться фосфорилированные соединения, DG гидролиза которых имеет невысокое отрицательное значение ("низкоэнергетические" соединения). Пример высокоэнергетического соединения — фосфоенолпировиноградная кислота (DG0' = –58,2 кДж/моль), низкоэнергетического — глицеро-1-фосфат (DG0' =—9,2 кДж/моль). АТФ на этой шкале занимает промежуточное положение, что и дает ему возможность наилучшим образом выполнять энергетические функции: переносить энергию от высокоэнергетических к низкоэнергетическим соединениям.
Если часто АТФ называют "энергетической валютой клетки, то, продолжая эту аналогию, можно сказать, что "валютная единица" выбрана клеткой в процессе эволюции весьма рационально. Порция свободной энергии в макроэргической фосфатной связи АТФ — это как раз та энергетическая порция, использование которой в биохимических реакциях делает клетку высокоэффективным энергетическим механизмом.
DmH+ — ВТОРАЯ УНИВЕРСАЛЬНАЯ ФОРМА КЛЕТОЧНОЙ ЭНЕРГИИ
В течение длительного времени считали, что АТФ и другие высокоэнергетические соединения, находящиеся в равновесии с ним, представляют собой единственную форму энергии, которая может использоваться живыми клетками во всех энергозависимых процессах. Вопрос о характере связи между транспортом электронов, с одной стороны, и превращением фосфорных соединений, с другой, долгое время оставался неясным. Было установлено, что использование энергетических ресурсов (органических или неорганических соединений при дыхании, света при фотосинтезе) связано с переносом электронов по цепи, состоящей из белковых и небелковых компонентов, способных к обратимому окислению — восстановлению. В результате этого переноса освобождающаяся на отдельных участках дыхательной или фотосинтетической цепи энергия трансформируется в химическую энергию фосфатных связей АТФ. Молекулярный механизм фосфорилирования, сопряженный с электронным транспортом, был неизвестен.
Позднее были получены экспериментальные данные о существовании еще одной формы энергии, также используемой клеткой для совершения разного рода работы. Открытие этой формы энергии принадлежит английскому биохимику Питеру Митчеллу (Р. Mitchell), разработавшему в 60-х гг. хемиосмотическую теорию энергетического сопряжения, объясняющую превращение (трансформацию) энергии, освобождающейся при электронном транспорте, в энергию фосфатной связи АТФ. П. Митчелл постулировал, что при переносе электронов по окислительно-восстановительной цепи, локализованной в мембранах определенного типа, называемых энергопреобразующими, или сопрягающими, происходит неравномерное распределение H+ в пространстве по обе стороны мембраны (рис. 25). Предложенная им модель предусматривает определенное расположение переносчиков электронов в сопрягающей мембране, например ЦПМ, которые могут быть погружены в глубь мембраны или локализованы у наружной и внутренней ее поверхностей так, что образуют "петли" в цепи переноса электронов. В каждой "петле" (у прокариот электронтранспортные цепи в сопрягающих мембранах могут формировать разное число "петель") два атома водорода движутся от внутренней стороны ЦПМ к наружной с помощью переносчика водорода (например. хинона). Затем два электрона возвращаются к внутренней стороне мембраны с помощью соответствующего электронного переносчика (например, цитохрома), а два протона освобождаются во внешнюю среду.
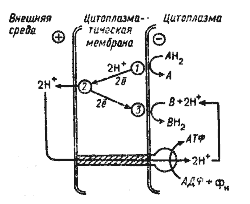
|
| Рис. 25. Схема переноса электронов и протонов по электронтранспортной цепи и протонной АТФ-синтазы: АНг и В — донор и акцептор электронов соответственно; 1, 2, 3 — компоненты электронтранспортной цепи. Объяснения см. в тексте |
Таким образом, в каждой окислительно-восстановительной "петле" два H+ переносятся из цитоплазмы клетки во внешнюю среду. Общее число протонов, перенесенных через ЦПМ и выделенных во внешнюю среду, при переносе двух электронов по электронтранспортной цепи зависит от числа образуемых ею окислительно-восстановительных "петель". Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой электронтранспортной цепи (фотосинтетической или дыхательной) во внешней среде происходит накопление ионов водорода (протонов), приводящее к подкислению среды, а в клеточной цитоплазме — их уменьшение, сопровождающееся ее подщелочением, т. е. на мембране возникает ориентированный поперек (трансмембранный) градиент ионов водорода.
Поскольку H+ — химические частицы, несущие положительный заряд, неравномерное их накопление по обе стороны мембраны приводит к возникновению не только химического (концентрационного) градиента этих частиц, но и ориентированного поперек мембраны электрического поля (суммарный положительный заряд, где происходит накопление H+, и отрицательный заряд по другую сторону мембраны). Таким образом, при переносе электронов на ЦПМ возникает трансмембранный электрохимический градиент ионов водорода, обозначаемый символом DmH+, и измеряемый в вольтах (В, мВ), который состоит из электрического (трансмембранная разность электрических потенциалов Dy) и химического (концентрационного) компонентов (градиент концентраций H+ — D pH). Измерения показали, что на сопрягающих мембранах прокариот при работе дыхательных и фотосинтетических электронтранспортных цепей DmH+ достигает 200–250 мВ, при этом вклад каждого компонента непостоянен. Он зависит от физиологических особенностей организма и условий его культивирования.
Итак, в соответствии с хемиосмотической теорией П. Митчелла, энергия, освобождаемая в результате работы; электронтранспортной цепи, первоначально накапливается в форме трансмембранного градиента ионов водорода. Разрядка образующегося DmH+ происходит с участием локализованного в той же мембране протонного АТФ-синтазного комплекса: Н+ возвращаются по градиенту DmH+ через Н+–АТФ-синтазу, при этом без возникновения каких-либо промежуточных высокоэнергетических соединений из АДФ и неорганического фосфата образуется АТФ. (Сами сопрягающие мембраны в интактном состоянии непроницаемы для ионов, особенно Н+ и ОН–.) Предположительно, для синтеза одной молекулы АТФ достаточен перенос двух протонов, т. е. Н+/АТФ=2. Однако не исключено, что Н+/АТФ может быть больше.
Локализованная в мембране H+–АТФ-синтаза катализирует реакции синтеза и гидролиза АТФ в соответствии с уравнением:
АДФ + ФН + nHНАР+ « АТФ + H2O + nHВНУТР+
Реакция, протекающая слева направо, сопряжена с транспортом H+ по градиенту DmH+, что приводит к его разрядке и синтезу АТФ. Протекающая в противоположном направлении реакция гидролиза АТФ, сопровождающаяся переносом Н+ против градиента, приводит к образованию (или возрастанию) DmH+ на мембране. Таким образом, АТФ-синтазный ферментный комплекс служит механизмом, обеспечивающим взаимное превращение двух форм клеточной энергии (DmH+ « АТФ), устройством, сопрягающим процессы окислительной природы с фосфорилированием.
Известно несколько реакций, генерирующих DmH+. У разных групп прокариот от 1 до 3 из них локализованы в дыхательной цепи. На 2 или 3 этапах DmH+ генерируется в темновых реакциях переноса электронов в фотосинтетической цепи. Образование DmH+ происходит при гидролизе АТФ в H+-зависимой АТФ-синтазной реакции. К числу устройств, генерирующих DmH+ посредством трансмембранного переноса H+, относится бактериородопсин галофильных архебактерий. У некоторых групп прокариот обнаружена локализованная в мембране неорганическая пирофосфатаза, катализирующая расщепление и синтез пирофосфата. Расщепление последнего приводит к генерированию DmH+. Наконец, источником DmH+ на ЦПМ прокариот могут быть процессы, связанные с выделением во внешнюю среду продуктов брожения, транспорт которых через мембрану происходит вместе с протонами.
Энергия в форме DmH+ может использоваться в различных энергозависимых процессах, локализованных на мембране. Синтез АТФ за счет DmH+ можно рассматривать как пример химической работы. С использованием энергии DmH+ могут осуществляться и другие виды химической работы в клетке: синтез пирофосфата, катализируемый связанным с мембраной ферментным комплексом; обратный перенос электронов, приводящий к восстановлению НАД(Ф)+.
Энергия в форме DmH+ используется для поглощения ДНК в процессе генетической трансформации и для переноса белков через мембрану. Движение многих прокариот обеспечивается энергией DmH+. Важная роль принадлежит DmH+ или одной из его составляющих в осуществлении процессов активного транспорта молекул и ионов через ЦПМ прокариот (рис. 26).
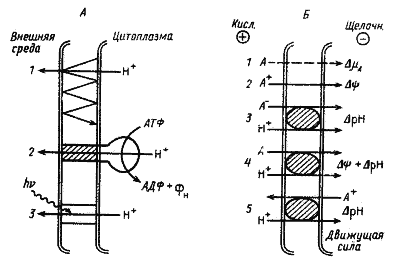
|
| Рис. 26. Транспортные системы в клетках прокариот. А — системы первичного транспорта: 1 — перенос электронов по окислительно-восстановительной цепи; 2 — протонная АТФ-синтаза; 3 — бактериородопсин. Б — системы вторичного транспорта: 1 — пассивный транспорт нейтральных молекул; 2 — активный перенос катионов (унипорт); 3 — симпорт анионов и протонов; 4 — симпорт нейтральных молекул и Н+; 5 — антипорт катионов и протонов (по Konings, Veldkamp, 1980) |
Все известные системы транспорта у прокариот можно разделить на два типа: первичные и вторичные. Разобранные выше примеры трансмембранного переноса H+ с участием окислительно-восстановительной "петли", бактериородопсина или в результате гидролиза АТФ, катализируемого H+—АТФ-синтазой, происходящие за счет химической энергии или электромагнитной энергии света, относятся к первичным транспортным системам (рис. 26, А). В результате их функционирования на мембране генерируется энергия в форме DmH+, которая в свою очередь может служить движущей силой, обеспечивающей с помощью индивидуальных белковых переносчиков поступление в клетку необходимых веществ разной химической природы и удаление из нее конечных продуктов метаболизма. Устройства, с помощью которых осуществляется трансмембранный перенос веществ по градиенту DmH+ или одной из его составляющих, относятся ко вторичным транспортным системам (рис. 26, Б).
Как известно, в случае пассивной диффузии вещества движущей силой служит только градиент его концентрации (Dm) вне и внутри клетки. Если подобный градиент существует и в процессе активного транспорта вещества, он может вносить определенный вклад в общую движущую силу процесса, однако этот вклад не является определяющим. В большинстве случаев перенос вещества по механизму активного транспорта происходит против концентрационного градиента этого вещества.
Вторичные транспортные системы могут быть также разделены на три группы. Перенос молекул вещества, не сопряженный с какими-либо встречными или сопутствующими перемещениями молекул других веществ, получил название унипорта. По механизму симпорта перенос молекул вещества сопряжен с переносом протонов в том же направлении и осуществляется при участии одного и того же белкового переносчика, В процессе антипорта перенос вещества сопряжен с переносом H+ в противоположном направлении. Поступление веществ в клетку по механизму симпорта и унипорта широко распространено у прокариот и служит для поглощения ими большинства необходимых органических и неорганических соединений.
Для понимания движущих сил, участвующих в активном транспорте разных типов молекул (электронейтральных, несущих положительный или отрицательный заряд), следует помнить, что в цитоплазме более щелочная среда и суммарный отрицательный заряд. Незаряженные молекулы (глюкоза, галактоза, нейтральные аминокислоты) переносятся в клетку вместе с протонами за счет обоих компонентов Dm — Dy и DpH
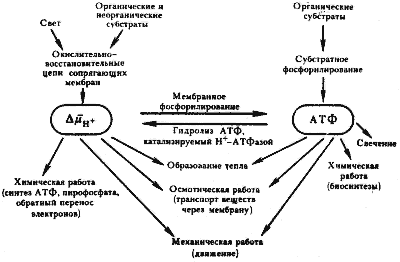
|
| Рис 27. Преобразование энергии в клетке прокариот (по Скулачеву, 1980) |
Молекулы, имеющие отрицательный заряд (глюконат, глутамат, H2PO4–), котранспортируются с H+ в электронейтральной форме за счет только DpH. Положительно заряженные молекулы и ионы переносятся через мембрану в цитоплазму по механизму унипорта с использованием в качестве движущей силы электрического компонента DmH+. Таким путем осуществляется перенос в клетку, например, ионов К+ или лизина. Примером антипортной системы может служить откачивание из цитоплазмы ионов Na+, происходящее в обмен на поступление в нее ионов H+. Движущей силой процесса является DpH.
Все это позволяет рассматривать энергию в форме DmH+ (наряду с АТФ) как широко используемую внутри клетки. Преобразование энергии в клетке прокариот схематически изображено на рис. 27. Как видно из этой схемы, АТФ и DmH+ можно считать двумя взаимопревращаемыми "энергетическими валютами" клетки, каждая из которых способна служить источником энергии для выполнения химической, осмотической, механической работ.
ДЛЯ ЧЕГО КЛЕТКЕ НЕОБХОДИМЫ ДВЕ ФОРМЫ ЭНЕРГИИ?
АТФ участвует в реакциях, протекающих в цитоплазме, т е. в форме АТФ энергией обеспечиваются все процессы, протекающие в водной среде. К числу последних относится большинство биосинтетических реакций. Помимо этого АТФ служит источником энергии для протекания ряда мембранзависимых процессов.
Энергия протонного градиента связана исключительно с мембранами, которые являются н необходимым компонентом Для его образования. Поэтому энергией в форме DmH+ могут обеспечиваться только процессы, локализованные на мембране. Таким образом, у DmH+ более узкая область приложения. В то же время использование клеткой энергии в форме DmH+ имеет определенные преимущества: DmH+ в форме его электрической составляющей — более удобная форма энергии для внутри- и межклеточной транспортировки. Скорость переноса энергии посредством диффузии АТФ в цитоплазме значительно медленнее, чем скорость передачи Dy по мембранам. Диффузия АТФ может быть сильно затруднена в клетках с развитой системой внутрицитоплазматических мембран. Наконец, перенос энергии посредством диффузии АТФ совсем неэффективен, если речь идет о межклеточном транспорте энергии, что важно для многоклеточных организмов. В этом случае эффективность передачи энергии по мембранам наиболее очевидна. Энергия в форме DmH+ не содержится в виде определенных порций, как это имеет место в молекуле АТФ. При гидролизе макроэргической фосфатной связи АТФ освобождается определенное количество энергии (DG0'=–31,8 кДж/моль). Если для сопряженного эндергонического процесса требуется меньшее количество энергии, остальная часть рассеивается в виде тепла. При использовании энергии в форме трансмембранного потенциала потерь, обусловленных запасанием энергии в виде порций, не происходит. С этим связано и еще одно преимущество при использовании клеткой энергии в форме DmH+ не существует нижнего порога для его образования. Для синтеза АТФ необходима разность окислительно-восстановительных потенциалов порядка 200 мВ. Ниже этого порогового значения АТФ не может быть синтезирован. Для образования энергии в форме трансмембранного потенциала подобных ограничений нет. Поэтому энергия DmH+ может образовываться и потребляться клеткой в условиях, когда синтез АТФ невозможен.
Таким образом, хотя каждая форма клеточной энергии может быть использована для осуществление химической, механической и осмотической работы, между ними существует определенное "разделение труда", например, большинство биосинтезов обеспечивается энергией АТФ, активный транспорт — энергией DmH+. Из этого следует, что клетке необходимо всегда иметь определенное количество энергии в той и другой легко мобилизуемой форме. Это может быть одной из причин существования в клетке двух взаимосвязанных энергетических пулов (резервуаров), между которыми при необходимости легко может осуществляться перекачка энергии (рис. 27). Емкость обоих энергетических пулов невелика. Например, величина DmH+ поддерживается на уровне 200–250 мВ. Внутриклеточная концентрация АТФ составляет около 2 мМ. Это также указывает на каталитическую роль АТФ в клетке. Подсчитано, что для удвоения клеточной массы молекула АТФ должна около 10000 раз участвовать в реакциях гидролиза и синтеза.
ЭНЕРГЕТИЧЕСКИЕ ЗАТРАТЫ КЛЕТКИ
В растущей бактериальной культуре потребление энергии в первую очередь связано с процессами биосинтеза веществ, из которых состоит клетка. Количество энергии, необходимое для биосинтетических целей, в большой степени зависит от состава среды культивирования. Теоретически рассчитано, что при выращивании культуры бактерий в среде с минеральными солями и глюкозой в качестве единственного источника углерода 1 моль АТФ затрачивается для синтеза 27 г вещества клеток. Если же единственным источником углерода служит СО2, тогда использование того же количества АТФ приведет к синтезу только 5 г вещества клеток.
Для Е. coli большая часть биосинтетических реакций в настоящее время известна. Это позволило определить количество АТФ, используемого для образования основных клеточных макромолекул. В культуре Е. coli, растущей в среде с глюкозой в качестве единственного источника углерода, для синтеза 1 г клеточного материала расходуется около 37 ммоль АТФ. Больше половины этого количества (около 20 ммоль) используется в реакциях, ведущих к синтезу белковых молекул. Следующими по энергетическим затратам биосинтетическими процессами являются ферментативные пути синтеза РНК и ДНК (около 7 ммоль АТФ), а также реакции полимеризации моносахаров (около 2 ммоль АТФ).
Помимо энергетических затрат на биосинтетические процессы, связанные с ростом, определенная часть клеточной энергии всегда тратится на процессы, не связанные непосредственно с ростом. Последние получили название процессов поддержания жизнедеятельности. К специфическим функциям поддержания жизнедеятельности относятся: обновление клеточного материала, осмотическая работа, обеспечивающая поддержание концентрационных градиентов между клеткой и внешней средой, подвижность клетки и др. Энергию, расходующуюся на осуществление перечисленных функций, обозначают как энергию поддержания жизнедеятельности,
Когда бактерии находятся в условиях, в которых у них по каким-либо причинам отсутствует рост и, следовательно, нет надобности в энергии для биосинтезов, у них и в этом случае не прекращаются определенные метаболические процессы. Примером крайнего проявления состояния покоя служат бактериальные эндоспоры, метаболическая активность которых находится на уровне, не всегда регистрируемом современными приборами. Однако и в этом случае она не равна нулю.
Здесь необходимо остановиться на одном принципиально важном моменте. Состояние покоя живых организмов — всегда динамическое, в отличие от статического состояния неживых систем. Это означает, что в покоящихся организмах концентрации большинства молекул поддерживаются не статически, а динамически, т. е. процессы распада органических соединений и компенсирующие их биосинтетические процессы продолжаются и в состоянии кажущегося покоя.
Активно обновляющимися клеточными веществами являются молекулы ферментных белков, иРНК, материал клеточных стенок и мембран. Динамическое состояние свойственно почти всем метаболитам и структурам клетки, различие наблюдается только в отношении скоростей процессов обновления различных клеточных компонентов. Исключение составляют молекулы ДНК и некоторых бактериальных белков. В отношении ДНК понятно, что динамическое состояние генетического материала повышает опасность возникновения ошибок при его обновлении и связанные с этим летальные последствия для организма.
В условиях активного роста энергия, используемая в процессах обновления, составляет лишь небольшую часть общих энергетических расходов клетки. В состоянии покоя общие энергетические затраты клетки значительно понижаются, и на этом фоне удельный вес расходуемой на обновление клеточных веществ энергии заметно возрастает. Значительная часть энергии поддержания жизнедеятельности расходуется на совершение осмотической работы. В приведенном выше расчете энергетических затрат E. coli для процессов активного транспорта используется примерно 5 ммоль АТФ.
Величина энергии поддержания жизнедеятельности в значительной степени зависит от условий роста. Так, для Azotobacter vinelandii, фиксирующего азот при низком (0,02 атм) и высоком (0,2 атм) уровне растворенного кислорода, она колеблется от 22 до 220 ммоль АТФ на 1 г биомассы, т. е. прямо пропорциональна концентрации растворенного O2. Клетка тратит много дополнительной энергии для защиты от избытка кислорода, ингибирующего ферментную систему, ответственную за фиксацию молекулярного азота. Энергия поддержания жизнедеятельности обычно составляет 10–20% всей энергии, расходуемой в энергозависимых процессах. Описаны, однако, условия, в которых бактерии расходуют на поддержание жизнедеятельности до 90%, вырабатываемой энергии.
КОНСЕРВИРОВАНИЕ ЭНЕРГИИ
Энергетический метаболизм в целом сопряжен с биосинтетическими и другими энергозависимыми процессами, происходящими в клетке, для протекания которых он поставляет энергию, восстановитель и необходимые промежуточные метаболиты. Сопряженность двух типов клеточного метаболизма не исключает некоторого изменения их относительных масштабов в зависимости от конкретных условий. Прокариоты, обладающие наиболее совершенными системами получения энергии (дыхание, фотосинтез), способны генерировать энергию в значительно большем количестве, чем необходимо для. роста и обеспечения всех энергозависимых клеточных функций.
Возможны такие условия, когда клетка запасает энергии больше, чем тратит. В этом случае она сталкивается с проблемой консервирования энергии. В молекулах АТФ энергия не хранится в течение длительного времени. Средняя продолжительность "жизни" молекул АТФ составляет около 1/3 с. Энергия в форме DmH+ также не может накапливаться. Движение H+ против градиента возможно только до достижения определенного уровня, после которого возникшая разность концентраций и электрических зарядов будет тормозить поступление ионов водорода против градиента. Таким образом, в молекулах АТФ и в виде DmH+ энергия находится в мобильной форме, призванной обеспечивать все идущие в настоящий момент энергозависимые процессы.
Проблема консервирования энергии решена прокариотами путем синтеза восстановленных высокополимерных молекул, главным образом полисахаридов, реже липидов или полипептидов. Молекулы запасных веществ плотно упакованы в гранулах и часто окружены белковой оболочкой (см. табл. 5). В таком виде они находятся в осмотически неактивном состоянии, что очень важно для клетки.
СПОСОБЫ СУЩЕСТВОВАНИЯ И ТИПЫ ЖИЗНИ У ПРОКАРИОТ
В зависимости от того, какой источник энергии могут использовать прокариоты, их делят на фототрофов (источник энергии — свет) и хемотрофов (источник энергии — окислительно-восстановительные реакции). Организмы, у которых источниками (донорами) электронов в энергетическом процессе являются неорганические вещества, предложено называть литотрофными, а те, у которых донорами электронов служат органические соединения, — органотрофными. Тогда в зависимости от источника энергии и природы донора электронов возможны четыре основных типа энергетического метаболизма: хемолитотрофия, хемоорганотрофия, фотолитотрофия и фотоорганотрофия.
У прокариот с хемотрофным типом энергетического метаболизма одно и то же соединение служит донором электронов, большая часть которых перемещается в соответствии с термодинамическим градиентом, что приводит к выделению свободной энергии, а меньшая — используется для образования восстановителя, потребляемого в конструктивном метаболизме. Это положение справедливо в отношении прокариот с энергетикой бродильного и дыхательного типов, при использовании в качестве энергетических ресурсов органических и неорганических соединений. У фототрофов использование света в качестве источника энергии требует дополнительного подключения химических соединений, служащих донорами электронов для образования восстановителя. Это связано со спецификой света как энергетического ресурса для живых систем.
Каждый тип энергетического метаболизма может осуществляться на базе различных биосинтетических способностей организма. Выше уже обсуждалось деление всех прокариот в зависимости от особенностей конструктивного метаболизма на две группы: авто- и гетеротрофов. Следовательно, можно выделить 8 сочетаний типов энергетического и конструктивного метаболизма, которые отражают возможности способов существования (питания) прокариот (табл. 12). Всем способам питания соответствуют реально существующие прокариотные организмы. Однако число видов прокариот, относящихся к группам, характеризующимся разными способами питания, далеко не одинаково. Подавляющее число прокариот сосредоточено в группе с хемоорганогетеротрофным типом питания. Такая же неравномерность в распределении по типам питания присуща и фотосинтезирующим прокариотам. Большинство (цианобактерии, пурпурные и зеленые серобактерии) относится к группе с фотолитоавтотрофным способом питания. Часто для характеристики способа питания прокариот пользуются более общими понятиями, чем приведенные в табл. 12, употребляя для этого сочетания только двух признаков: источник энергии (фото-, хемо-) + источник углерода для построения веществ тела (авто-, гетеро-).
Перечисленные в табл. 12 сочетания основных видов энергетического и конструктивного метаболизма характеризуют все возможные способы питания прокариотных организмов. Некоторые прокариоты могут существовать только на базе одного какого- нибудь способа питания. Например, одноклеточная цианобактерия Synechococcus elongatus может использовать в качестве источника энергии только свет, а как основной источник углерода в конструктивном метаболизме — углекислоту. Характеризуя способ существования (образ жизни, тип метаболизма) этого организма, мы говорим, что он облигатный фотолитоавтотроф. Многие бактерии, относящиеся к роду Thiobacillus, — облигатные хемолитоавтотрофы, т. е. источником энергии для них служат процессы окисления различных соединений серы, а источником углерода для построения веществ тела, — углерод углекислоты. Подавляющее большинство бактерий — облигатные хемоорганогетеротрофы, использующие в качестве источника углерода и энергии органические соединения.
Таблица 12. Способы существования прокариот
| Источник энергии | Донор электронов | Источник углерода | Способ существования | Представители прокариот |
| Окислительно-восстановительные реакции | неорганические соединения (H2, H2S, NH3, Fe2+ и др.) | CO2 | хемолито- автотрофия |
нитрифицирующие, тионовые, водородные бактерии; ацидофильные железобактерии |
| органические соединения | хемолито- гетеротрофия |
метанобразующие архебактерии, водородные бактерии | ||
| органические соединения | CO2 | хемооргано- автотрофия |
факультативные метилотрофы, окисляющие муравьиную кислоту | |
| органические соединения | хемооргано- гетеротрофия |
большинство прокариот* | ||
| Свет | неорганические соединения (H2O, H2S, S0, и др.) | CO2 | фотолито- автотрофия |
цианобактерии, пурпурные и зеленые бактерии** |
| органические соединения | фотолито- гетеротрофия |
некоторые цианобактерии, пурпурные и зеленые бактерии | ||
| органические соединения | CO2 | фотооргано- автотрофия |
некоторые пурпурные бактерии | |
| органические соединения | фотооргано- гетеротрофия |
пурпурные и некоторые зеленые бактерии, галобактерии, некоторые цианобактерии |
* Все животные, грибы.
** Высшие растения.
Для некоторых представителей группы цианобактерий наряду с фотолитоавтотрофней показана способность к фотолито- или хемоорганогетеротрофии. Ряд хемолитоавтотрофных видов Thiobacillus способы существовать за счет использования в качестве источников энергии и углерода органических соединений, т. е. хемоорганогетеротрофно.
В приведенных выше примерах метаболические возможности указанных цианобактерий и видов Thiobacillus оказались гораздо шире, чем необходимо для осуществления метаболизма по одному типу. В этом случае мы говорим о том, что данные организмы — факультативные фотолитоавтотрофы или хемолитоавтотрофы, т. е. осуществление данного типа метаболизма не является для них единственным и обязательным (облигатным). Организмы, способные одновременно использовать два источника углерода (CO2 + органические вещества) и/или энергии (например, энергию света — энергию окисления химического соединения), называются миксотрофами.
В рамках разобранных выше основных способов питания, определяющих возможности существования прокариотных организмов, в мире прокариот обнаружено множество типов (форм) жизни. Тип жизни — понятие, отражающее, с одной стороны, специфику процессов энергетического метаболизма, с другой — специфику процессов конструктивного метаболизма, присущую определенной группе организмов. Разберем это на примере прокариот, для которых обязателен хемоорганогетеро-трофный способ существования. Энергетические процессы этих организмов различаются исходными субстратами, специфичностью промежуточных окислительно-восстановительных превращений и природой конечных акцепторов электронов; конструктивные — разной степенью развития биосинтетических способностей, т. е. различными потребностями в готовых питательных веществах.
Наиболее примитивную и древнюю группу энергетических процессов составляют процессы брожения, когда органическое вещество служит донором и конечным акцептором электронов, а молекулярный кислород в реакциях окислительной природы участия не принимает. Известны молочнокислое, спиртовое, пропионовокислое, маслянокислое и некоторые другие виды брожения, каждое из которых является специфической формой решения "энергетической проблемы" и осуществляется группой прокариот, характеризующихся определенными биосинтетическими способностями. Известны прокариотные организмы, получающие энергию за счет процессов неглубокого или полного окисления органического субстрата молекулярным кислородом.
Таким образом, на базе хемоорганогетеротрофного способа питания можно выделить несколько типов жизни, представленных определенными группами прокариот, осуществляющими конкретный тип энергетического метаболизма в сочетании с присущими им особенностями метаболизма конструктивного.
М.В.Гусев, Л.А.Минеева © 1992-2001
Кафедра клеточной физиологии и иммунологии биологического факультета МГУ им. М.В.Ломоносова © 2000-2001
Страница обновлена 09.09.01
Оформление: А.В.Киташов