ГЛАВА 10. ГЕНЕТИЧЕСКИЕ МЕХАНИЗМЫ ЭВОЛЮЦИИ ПРОКАРИОТ
Вся информация о признаках, присущих организму, сосредоточена в его генетическом аппарате. Он обеспечивает сохранение и воспроизведение этих признаков в процессе размножения организма, так как возникающие дочерние особи обнаруживают в большинстве случаев полное сходство с родительскими формами. Это говорит о том, что генетический аппарат обладает высокой стабильностью и точностью механизмов, обеспечивающих его функционирование. Однако стабильность генетического аппарата не абсолютна, так как это исключало бы всякую возможность его изменений и, следовательно, эволюционных преобразований, приведших в конечном итоге к возникновению разнообразных форм жизни, свидетелями (и представителями) которых мы являемся. Таким образом, генетический аппарат должен быть организован так, чтобы, с одной стороны, обеспечивать свою стабильность, с другой — быть достаточно пластичным, т. е. обладать способностью к изменчивости.
В этой главе мы обсудим применительно к прокариотам вопросы, касающиеся организации и функционирования генетического аппарата, определяющего свойства наследственности и изменчивости, а также изменения в генетическом материале и процессы, приводящие к этим изменениям. Постараемся также оценить вклад каждого из процессов в эволюцию прокариотных организмов.
ГЕНЕТИЧЕСКИЙ АППАРАТ ПРОКАРИОТ
Как это ни кажется в настоящее время парадоксальным, но до 40-х гг. нашего столетия немногие микробиологи думали, что бактерии обладают наследственностью, основанной на тех же принципах, которые установлены для высших организмов. Прокариоты не имеют ни оформленного ядра, ни хромосом, аналогичных таковым эукариотных клеток, поэтому считали, что бактерии в генетическом отношении представляют собой неупорядоченную форму жизни. Одним из первых к пониманию того, что бактерии и высшие организмы подчиняются общим генетическим законам, подошел М. Бейеринк, описавший у прокариот стабильные, легко распознаваемые и наследуемые изменения.
Генетический материал любой клетки представлен ДНК, информационные свойства которой определяются специфической последовательностью четырех нуклеотидов в полинуклеотидной цепи. Полуконсервативный механизм репликации ДНК, в результате которого из одной родительской двухцепочной молекулы образуются две дочерние молекулы, содержащие по одной родительской и одной вновь синтезированной комплементарной полинуклеотидной цепи, наилучшим образом обеспечивает идентичность исходной и синтезированных молекул и, следовательно, сохранность видоспецифической наследственной информации в ряду поколений клеток и организмов (см. гл. 4). Частота ошибок, возникающих в процессе репликации, порядка 10-7.
Реализация наследственной информации в процессе жизненного цикла (онтогенеза) организма — двухступенчатый процесс. Сначала с определенных участков ДНК информация подписывается (транскрибируется) в виде комплементарных нулеотидных последовательностей молекул иРНК, которая перемещается в цитоплазму, связывается с рибосомами и в рибосоме с иРНК осуществляется перевод (трансляция) генетической информации в определенную последовательность аминокислотных остатков молекулы белка.
Процесс транскрипции находится в клетке под строгим контролем, поэтому имеет место как неодинаковое транскрибирование во времени разных участков ДНК (генов), так и неодинаковая скорость, с которой гены могут транскрибироваться. В результате количество молекул иРНК в клетке, комплементарных разным генам, сильно различается. Хотя в целом механизмы синтеза ДНК и РНК сходны, процесс транскрипции не обладает той степенью точности, которая характерна для репликации ДНК. Однако поскольку иРНК не способна к самовоспроизведению, возникающие при ее синтезе ошибки в последующих клеточных генерациях не воспроизводятся и, следовательно, не могут наследоваться.
Информационные РНК служат матрицами для синтеза различных белковых молекул. Перевод генетической информации с "языка" нуклеотидов на "язык" аминокислот—сложный многостадийный процесс, включающий активацию аминокислот, образование ими комплексов с особым видом РНК (транспортными РНК, или тРНК), взаимодействие этих комплексов с иРНК, связанной с рибосомой, приводящее в конечном итоге к формированию полипептидной цепи, аминокислотный состав которой изначально запрограммирован в определенном участке ДНК. В осуществлении каждой из стадий, ведущих к синтезу молекулы белка, участвует несколько различных ферментов.
Хотя механизм трансляции отличается высокой точностью, вероятность ошибки в целом выше, чем в случае синтеза молекул ДНК и РНК. Наиболее уязвимый этап — "узнавание" с помощью фермента аминокислоты соответствующей молекулой тРНК. По имеющимся данным, частота возникновения ошибок на этом этапе порядка 10-4, что и определяет, возможно, уровень точности процесса синтеза белка в целом. Однако, как и в случае синтеза РНК, ошибки в процессе трансляции, приводящие к синтезу измененной молекулы белка, не воспроизводятся, если они не закодированы исходно в генетическом материале.
Таким образом, процессы транскрипции и трансляции, служащие для выражения в онтогенезе генетической информации, не приводят к наследованию изменений, возникающих при их функционировании. Только изменения, происходящие в молекулах ДНК, могут сохраняться в ряду поколений, поскольку они воспроизводятся в процессе репликации. Следовательно, в основе эволюции прокариот лежит способность к изменению только их генетического материала. У прокариот весь генетический материал, необходимый для жизнедеятельности, локализован в одной хромосоме, т. е. бактериальная клетка гаплоидна. В определенных условиях в клетках бактерий может содержаться по нескольку копий хромосомы.
У многих бактерий обнаружены нехромосомные генетические элементы: плазмиды, умеренные фаги и мигрирующие элементы (транспозоны и IS-элементы)26. Для плазмид характерно стабильное существование в нехромосомном состоянии. Транспозоны и IS-элементы входят, как правило, в состав хромосом, но способны переходить из хромосомы в плазмиду, поэтому также могут быть отнесены к нехромосомным генетическим элементам.
26 От англ. insertion sequences — вставные последовательности.
Изучение нехромосомных генетических элементов обнаружило, что общий объем ДНК, входящий в их состав, превышает объем генома каждой особи. Таким образом, у прокариот большой объем генетической информации оказывается рассредоточенным в нехромосомных элементах. Это заставляет по-новому подходить к вопросу об организации генетической информации в мире прокариот. В бактериальной хромосоме локализована генетическая информация, необходимая для существования конкретного вида бактерий в определенном диапазоне условий внешней среды: при наличии используемых источников углерода, азота, доступности или отсутствии молекулярного кислорода и т. д. Особенностью генетической информации, содержащейся в нехромосомных элементах, является ее необязательность для жизнедеятельности бактерий, т. е. в ее отсутствие бактериальная клетка жизнеспособна, но, как видно из дальнейшего материала, важная роль нехромосомных генетических элементов заключается в том, что они расширяют возможности существования бактериального вида, обеспечивают обмен генетическим материалом на большие расстояния по горизонтали и играют определенную роль в эволюции прокариот.
Плазмиды обнаружены у многих бактерий, принадлежащих к разным таксономическим группам. Количество плазмидной ДНК в клетке составляет обычно не более нескольких процентов от клеточного генома, а число плазмид колеблется от 1 до 38. Плазмиды — это линейные или кольцевые ковалентно замкнутые молекулы ДНК, содержащие от 1500 до 40 000 пар нуклеотидов. Большинство плазмид состоит из трех групп генов: участка ДНК, ответственного за автономную репликацию плазмиды в клетке; системы генов, обеспечивающих возможность переноса плазмид из одной клетки в другую; генов, определяющих свойства, полезные для клетки-хозяина. Отличительная особенность плазмид — способность к автономной репликации, поэтому минимальное количество ДНК, которое может быть названо плазмидой, — это фрагмент, обеспечивающий автономную репликацию плазмидной ДНК в клетке как единого целого.
Обычно о присутствии плазмид в бактериальной клетке судят по проявлению определенных признаков, к которым относится устойчивость к отдельным лекарственным препаратам, способность к переносу генов при конъюгации, синтез веществ антибиотической природы, способность использовать некоторые сахара или обеспечивать деградацию ряда веществ. Из перечисленного выше видно, что плазмиды делают возможным существование организмов в более широком диапазоне условий внешней среды, т. е. действуют как факторы адаптации. Большую группу составляют плазмиды с нерасшифрованными функциями; такие плазмиды выявляют с использованием физико-химических методов.
Мигрирующие элементы, представленные транспозонами и IS-элементами, — это линейные молекулы двухнитевой ДНК, размеры которых колеблются от 200 до 6000 пар нуклеотидов. Отличительная особенность мигрирующих элементов — их неспособность к автономной репликации. Мигрирующие элементы могут встраиваться в разные участки бактериальной хромосомы или мигрировать с бактериальной хромосомы на плазмиду; их репликация осуществляется под контролем тех же механизмов, что и у соответствующей хромосомы или плазмиды. Частота переносов (транспозиции) мигрирующих элементов колеблется от 10-4 до 10-7. IS-элементы содержат информацию, необходимую только для их переноса внутри клетки, никаких выявляемых признаков в них не закодировано. Транспозоны устроены более сложно: в них включены некоторые гены, не имеющие отношения к процессу транспозиции. Известны транспозоны, содержащие гены устойчивости к антибиотикам, ионам тяжелых металлов и другим ингибиторам.
Для переноса мигрирующих элементов между клетками нужен переносчик, которым могут быть определенные плазмиды или фаги. Встраивание мигрирующих элементов в бактериальную хромосому оказывает мутагенное действие, так как при этом происходит включение фрагмента ДНК, приводящее к изменению порядка расположения нуклеотидов в триплете и, как следствие этого, нарушению процесса транскрипции.
ИЗМЕНЕНИЕ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
Первое, что поразило исследователей, когда они поближе познакомились с миром прокариот, — огромное разнообразие присущих им признаков. Это послужило в свое время основанием для дискуссии о том, существуют ли у прокариот реально очерченные виды, между которыми нет переходов, или же имеет место почти бесконечная вариабельность одного вида, так что в конечном итоге само понятие "вид" применительно к этим организмам теряет тот смысл, который в него вкладывается. Спор был разрешен после разработки Р. Кохом метода чистых культур. Было доказано, что и у бактерий вид — это реальность. Представления о простоте переходов между родами и видами оказались неверными.
В процессе экспериментальной работы с прокариотами исследователи часто наблюдали, что популяция одного вида при культивировании в течение длительного времени или в разных условиях подвержена изменениям. Накопилось огромное количество фактов, иллюстрирующих эти изменения, однако механизмы, лежащие в основе наблюдаемых явлений, были непонятны. И только успехи, достигнутые за последние десятилетия в области изучения строения и функционирования генетического аппарата прокариот, позволили разобраться в этом вопросе.
Прежде чем переходить к дальнейшему изложению материала, целесообразно ввести некоторые понятия. Генотипом, или геномом, называют совокупность всех генов, присущих данному организму, т. е. его генетическую конституцию. Под фенотипом понимают совокупность признаков, присущих данному организму. Оказалось, что все наблюдаемые изменения можно разделить на два типа. К первому относят те из них, которые, как правило, проявляются у подавляющего большинства особей в популяции при изменении внешних условий и наблюдаются до тех пор, пока действует фактор, вызвавший эти изменения. Такой тип изменчивости получил название не наследственного, или модификационного, а само явление названо модификацией.
Ко второму типу относятся изменения признаков, которые первоначально возникают как редкие события в популяции особей (с частотой 1 на 104—1011 клеток). Если измененные особи имеют некоторое преимущество перед неизмененными, выражающееся в повышенной скорости роста или жизнеспособности, они постепенно накапливаются в популяции и вытесняют исходные особи. Изучение особенностей второго типа изменений привело к заключению, что последние возникают случайно. И наконец, эти изменения постоянны, т. е. передаются из поколения в поколение при размножении организма. Такой тип изменчивости был назван наследственным.
Долгое время не было ясно, каков механизм модификационных изменений, могут ли они наследоваться и какова их роль в эволюции организмов. В настоящее время показано, что модификация — изменение, происходящее на уровне фенотипа и не затрагивающее клеточный генотип. Все признаки клетки определяются ее генотипом, но в определенных условиях она пользуется не всей заложенной в ней генетической информацией, количество которой гораздо больше, чем необходимо клетке для существования в конкретных условиях. Реакция клетки на изменение внешних условий приводит к проявлению какихто новых признаков, свойств, которые не обнаруживались в исходной культуре. Однако информация, необходимая для проявления этих признаков, обязательно содержится в клеточном Геноме. Модификация есть результат пластичности клеточного метаболизма, приводящего к фенотипическому проявлению "молчащих" генов в конкретных условиях. Таким образом, модификационные изменения имеют место в рамках неизменного клеточного генотипа.
Существует несколько типов модификационных изменений. Наиболее известны адаптивные модификации, т. е. ненаследственные изменения, полезные для организма и способствующие его выживанию в изменившихся условиях. Причины адаптивных. модификаций кроются в механизмах регуляции действия генов. Адаптивной модификацией является адаптация клеток Е. coli к лактозе как новому субстрату (см. гл. 8). У ряда бактерий обнаружена универсальная адаптивная реакция в ответ на различные стрессовые воздействия (высокие и низкие температуры, резкий сдвиг pH и др.), проявляющаяся в интенсивном синтезе небольшой группы сходных белков. Такие белки получили название белков теплового шока, а само явление — синдром теплового шока. Стрессовое воздействие на бактериальную" клетку вызывает ингибирование синтеза обычных белков, но индуцирует синтез небольшой группы белков, функция которых предположительно заключается в противодействии стрессовому воздействию путем защиты важнейших клеточных структур, в первую очередь нуклеоида и мембран. Еще не ясны те регуляторные механизмы, которые запускаются в клетке при воздействиях, вызывающих синдром теплового шока, но очевидно, что это универсальный механизм неспецифических адаптивных модификаций. Не все модификации обязательно адаптивны. При интенсивном действии многих агентов наблюдаются ненаследуемые изменения, случайные по отношению к вызвавшему их воздействию. Они проявляются только в условиях, которые их вызывают. Причины появления таких фенотипически измененных клеток связаны с ошибками процесса трансляции, вызванными этими агентами.
Таким образом, модификационная изменчивость не затрагивает генетической конституции организма, т. е. не является наследственной. В то же время она вносит определенный вклад в процесс эволюции. Адаптивные модификации расширяют возможности организма к выживанию и размножению в более широком диапазоне условий внешней среды. Возникающие в этих условиях наследственные изменения подхватываются естественным отбором и таким путем происходит более активное освоение новых экологических ниш и достигается более эффективная приспособляемость к ним.
Наследственные изменения можно подразделить на изменения, возникающие в результате мутаций и рекомбинаций генетического материала. Скачкообразные изменения в генетическом материале клетки, приводящие к появлению новых признаков, получили название мутаций. Мутации возникают в популяции особей всегда, часто без видимых воздействий на популяцию. Такие мутации, причины возникновения которых нам неизвестны, называются спонтанными. Повышать частоту мутаций по сравнению со спонтанным фоном, т. е. индуцировать их, могут физические, химические и биологические факторы, действующие на генетический материал клетки. Физические факторы — это прежде всего коротковолновое излучение (ультрафиолетовые и рентгеновские лучи). К химическим мутагенам относятся аналоги оснований, производные акридина, алкилирующие и дезаминирующие агенты. Биологические факторы — это в первую очередь мигрирующие элементы (транспозоны и IS-элементы).
Мутации, независимо от того, имеют ли они спонтанное происхождение или индуцированы какимлибо мутагеном, по характеру перестроек, происшедших в ДНК, можно разделить на мутации, состоящие в изменении одного нуклеотидного остатка молекулы ДНК, так называемые точковые мутации, и мутации, при которых наблюдается изменение участка молекулы ДНК размером больше одного нуклеотида. Точковые мутации в свою очередь могут быть разделены на несколько классов в зависимости от того, какие конкретно химические перестройки происходят в молекуле ДНК в рамках одного нуклеотидного остатка: замена, вставка или выпадение. К мутациям, затрагивающим сегмент бактериальной хромосомы, ведут выпадение нескольких оснований или даже генов, перемещение их в пределах одной хромосомы, умножение или удвоение части хромосомы.
Частым типом структурных повреждений ДНК, вызываемых УФизлучением, является образование пиримидиновых димеров в результате ковалентного связывания соседних пиримидиновых оснований. Реже УФ вызывает разрыв водородных связей, образование межцепочечных поперечных сшивок и поперечных сшивок между ДНК и белком. Ионизирующие излучения всех видов вызывают главным образом одноцепочечные разрывы в ДНК; разрывов, поражающих обе цепи, обычно на порядок меньше. Различные химические мутагены индуцируют образование внутрицепочечных и межцепочечных поперечных сшивок и одноцепочечные разрывы ДНК.
В процессе эволюции прокариоты выработали способы защиты генетического материала от повреждающего воздействия облучения и различных химических факторов. В клетках прокариот обнаружены эффективные системы репарации мутационных повреждений.
Наиболее изученными механизмами восстановления повреждений ДНК являются фотореактивация, вырезание повреждений и пострепликационное, или рекомбинационное, восстановление. Фотореактивация — наиболее простой механизм, восстанавливающий лишь индуцированные УФизлучением повреждения ДНК, сопровождающиеся образованием пиримидиновых димеров. Особенность фотореактивации в том, что ее действие распространяется только на одну цепь ДНК и не зависит от того, является ли молекул" В ДНК одно или двухцепочечной. Осуществляется фотореактивация светозависимым фотореактивирующим ферментом, обеспечивающим специфическое расщепление пиримидиновых димеров (рис. 38, А).

|
| Рис. 38. Механизмы восстановления повреждений ДНК. А — фотореактивация пиримидиновых димеров; 1 — фотореактивирующий фермент + видимый свет. Б — вырезание одноцепочечных повреждений: 1 — сегмент интактной ДНК; 2 — повреждение в одной из цепей ДНК; 3 — вырезание короткого сегмента, содержащего поврежденный участок; 4 — заполнение образовавшейся бреши нуклеотидами, комплементарными к интактной цепи, функционирующей в качестве матрицы; сшивание их с помощью ДНКполимеразы и ДНКлигазы; В — пострепликационное восстановление ДНК; — сегмент двухцепочечной молекулы ДНК, содержащей повреждение; 2 — репликация молекулы, приводящая к образованию двух молекул, одна из которых содержит повреждение и брешь в разных цепях; 3 — обмен генетическим материалом между идентичными цепями сестринских молекул; 4 — образование молекул, каждая из которых содержит одну интактную цепь, а в другой — повреждение или брешь. Крестиком обозначено повреждение; точками — восстановительный синтез; волнистой линией — синтезированные цепи дочерних молекул ДНК |
Вырезание повреждений — основной темновой механизм восстановления различных одноцепочечных повреждений ДНК, в том числе и пиримидиновых димеров. Особенность этого механизма репарации в том, что восстановление одноцепочечных повреждений происходит только тогда, когда неповреждена комплементарная цепь молекулы ДНК. В процессе темновой репарации происходит вырезание в одной из цепей молекулы ДНК коротких сегментов (длиной около 30 нуклеотидов), содержащих поврежденный участок, и последующее заполнение образовавшейся бреши комплементарными нуклеотидами с использованием неповрежденной цепи ДНК в качестве матрицы (рис. 38, Б).
Механизмы, обеспечивающие восстановление повреждений в обеих цепях молекулы ДНК, зависят от характера повреждений. Принципиальная схема заключается в следующем (рис. 38, В). ДНКполимераза, катализирующая репликацию ДНК, "встретив" на своем пути повреждение, "перескакивает" через него, и процесс репликации продолжается. Образуются две дочерние молекулы, одна из которых содержит в одной цепи первичное повреждение, в другой — брешь, возникшую при репликации и располагающуюся напротив повреждения.
Заделывание бреши происходит путем генетического обмена между идентичными цепями сестринских двухцепочечных молекул. В результате каждая из них имеет теперь по одной неповрежденной цепи, которая может служить матрицей в процессе репарации повреждений разного типа, как это изображено на схеме Б того же рисунка.
Фенотипическое проявление мутаций. Поскольку мутация — это стабильное изменение наследственного материала клетки, она реализуется по тем же каналам, что и любая другая генетическая информация. На этом пути судьба мутаций различна. Некоторые из них не влияют на признаки организма, оставаясь "молчащими". Такие мутации могут не проявляться в процессе трансляции, т. е. не приводить к изменению аминокислотной последовательности синтезируемого белка. В другом случае изменение может происходить вдали от активного центра фермента и потому не сказываться на его функции. Если же мутация приводит к изменению в активном центре или резко влияет на его структуру, это сразу сказывается на функциях фермента. Диапазон изменения функциональной активности фермента в этом случае велик: от незначительного понижения активности до полной ее потери. В последнем случае это часто приводит к гибели организма.
Для проявления мутации необходимо, чтобы прошел по крайней мере один цикл репликации ДНК, в которой исходно имело место изменение нуклеотидной последовательности (премутация). Только если это исходное изменение закрепится после репликации в дочерней молекуле ДНК, оно становится стабильным, а отсюда и наследственным. Для выражения мутации в фенотипе необходимо прохождение этапов транскрипции и трансляции. Иногда для проявления мутационно измененного признака, т. е. фенотипического выражения мутации, необходимо несколько клеточных делений. Так, если мутация привела к нарушению способности синтезировать какойлибо витамин, например тиамин, то в течение нескольких генераций потребность в тиамине у мутантных клеток не обнаруживается. В этот период мутантные клетки доиспользуют тиамин, содержащийся в исходной немутантной клетке. Когда же запасы витамина иссякнут, мутанты смогут размножаться только при добавлении экзогенного тиамина.
На проявление мутантных признаков влияет также количество копий хромосомы, содержащихся в клетке. Все прокариоты гаплоидны, имеют набор генов, локализованных в одной хромосоме. В определенных условиях в клетке можно обнаружить несколько копий одной хромосомы. Если в такой клетке произошла мутация, приведшая к нарушению синтеза определенного метаболита, то она сразу (после одного цикла репликации—транскрипции—трансляции) не проявится, поскольку синтез необходимого клетке метаболита будет осуществляться в результате функционирования неповрежденных генов, содержащихся в остальных хромосомных копиях. Для фенотипического выражения мутантного гена необходимо, чтобы он содержался в клетке в "чистом" виде, т. е. клетка имела одну копию хромосомы с мутантным геном, или чтобы все копии хромосомы в клетке имели одинаковый генотип. Это происходит через несколько клеточных делений (рис. 39).

|
| Рис. 39. Проявление мутаций в прокариотной клетке, имеющей четыре копии хромосомы (по Schlegel, 1972) |
Ко второму типу наследственной изменчивости относятся изменения, возникающие у прокариот в результате рекомбинации генетического материала, при которой происходит частичное объединение геномов двух клеток. Известны три основных способа, приводящих к рекомбинации генетического материала прокариот (конъюгация, трансформация и трансдукция), различающихся механизмами передачи хромосомной ДНК.
При конъюгации, для которой необходим непосредственный контакт между бактериальными клетками, осуществляется направленный перенос генетического материала от клеткидонора
в клеткуреципиент. Как правило, в клеткуреципиент переносится только часть генетического материала клеткидонора, в результате чего образуется неполная зигота, или мерозигота, содержащая часть генома донора и полный геном клеткиреципиента. Участки перенесенной от донора ДНК находят гомологичные участки в молекуле ДНК реципиента, между которыми происходит генетический обмен (рис. 40, А). В результате часть донорной ДНК встраивается (интегрируется) в геном реципиента, а соответствующая часть реципиентной ДНК из него исключается.
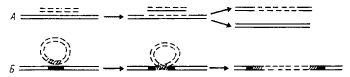
|
| Рис. 40. Рекомбинация между гомологичными (А) и негомологичными (Б) фрагментами ДНК. Объяснения см. в тексте |
Трансформация бактерий заключается в переносе ДНК, выделенной из одних клеток, в другие. Для трансформации не требуется непосредственного контакта между двумя клетками. Способность ДНК проникать в клеткуреципиент зависит как от природы самой ДНК, так и от физиологического состояния клеткиреципиента. Трансформирующей ДНК могут быть только высокомолекулярные двухцепочечные фрагменты, при этом проникать в бактериальную клетку может ДНК, выделенная из разных биологических источников, но включаться в геном — только ДНК с определенной степенью гомологичности. После того как экзогенный фрагмент ДНК, проникший в клетку, нашел гомологичный фрагмент ДНК клеткиреципиента, между ними происходит генетический обмен аналогично тому, как это имеет место на последнем этапе конъюгации (рис. 40, А).
Конъюгация и трансформация — не единственные способы передачи генетического материала. Гены могут переноситься из одной бактериальной клетки в другую с помощью умеренных фагов. Такой перенос бактериальных генов получил название трансдукции. Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц случайно захватит фрагмент бактериальной хромосомы, как правило, содержащий очень небольшое число генов. Когда такая фаговая частица заражает бактериюреципиент, бактериальная ДНК проникает в клетку таким же путем, как фаговая. Между трансдуцированной бактериальной ДНК и гомологичным участком бактериальной хромосомы может произойти обмен, и как следствие его возникают рекомбинанты, несущие небольшую часть генетического материала клеткидонора (рис. 40, А). Передача признаков с помощью фагов показана для бактерий, принадлежащих к разным родам.
Наконец, еще один путь переноса генетического материала у прокариот осуществляется с помощью плазмид определенного типа, обладающих генами, обеспечивающими эту возможность. Такие плазмиды помимо переноса собственного генетического материала могут обеспечивать перенос хромосомных генов, плазмид, не обладающих способностью к самостоятельному переносу, а также осуществлять передачу транспозонов из плазмиды в хромосому или другую плазмиду.
Все известные способы передачи генетической информации с помощью плазмид создают огромные возможности для интенсивных генетических обменов между клетками различных бактерий. Плазмидам и другим нехромосомным генетическим элементам принадлежит основная роль в передаче генетической информации "по горизонтали". Можно предположить, что в природе любая генетическая информация может быть перенесена в любую клетку прокариот, если не прямо, то через посредников. Подтверждением этого могут служить данные по введению с помощью сконструированной плазмиды в бактериальную клетку эукариотной ДНК и ее репродукции там.
Как редкое событие, происходящее с частотой 10–4—10–7, плазмиды или отдельные гены, входящие в их состав, могут включаться в бактериальную хромосому. Поскольку ДНК плазмиды и бактериальной клетки не имеют одинаковых нуклеотидных последовательностей, т. е. не являются гомологичными, рекомбинация между ними происходит не по механизму обмена, а по механизму встраивания (рис. 40, Б). Рекомбинации такого типа происходят также с участием транспозонов и IS-элементов при их перемещении (транспозиции) в пределах хромосомы. Встраивание плазмид и мигрирующих элементов помимо того, что приводит к введению в хромосому дополнительного генетического материала, может вызывать перестройку бактериального генома: нарушать целостность генов или регуляцию их функционирования, т. е. вызывать мутации.
ВКЛАД ОТДЕЛЬНЫХ ГЕНЕТИЧЕСКИХ МЕХАНИЗМОВ В ЭВОЛЮЦИЮ ПРОКАРИОТ
Выше мы рассмотрели организацию генетического аппарата прокариот, осуществляющего передачу генетической информации от одного поколения к следующему, т. е. по "вертикали", обратив внимание на такие его черты, как стабильность и точность функционирования. Однако стабильность генетического аппарата не абсолютна и при всей надежности изменения являются его неотъемлемым свойством. Для прокариот характерна большая способность к генетическим изменениям, являющимся результатом мутаций, а также развития путей "горизонтального" переноса генов между бактериальными клетками.
Закономерности и вклад различных процессов в эволюцию прокариот проще представить, если рассматривать их применительно к некой достаточно генетически однородной популяции организмов. Генетический состав такой популяции называется генофондом и по отношению к ней эволюцию можно определить как изменения в генофонде, происходящие в результате действия определенных генетических механизмов и отбора.
Новые наследственные признаки возникают в генофонде в результате генных мутаций. Последние создают фонд наследственных изменений, служащих исходным материалом (сырьем) для эволюции. Вероятно, мутации являются и самым первым видом наследственной изменчивости, возникшим одновременно с началом функционирования ДНК как информационной молекулы, поскольку для них не нужно никаких дополнительных структур и механизмов. Способность к мутированию заложена в химическом строении молекулы ДНК, а проявление мутационных изменений идет по тем же каналам, что и обычная генетическая информация клетки. Возможно, в течение длительного времени мутационные изменения были единственной формой изменчивости. На протяжении миллионов лет мутации в сочетании с естественным отбором сыграли решающую роль в появлении тех видов бактерий, которые известны сейчас.
Скорость эволюции определялась частотой возникновения мутаций. Можно только предполагать, что в начале биологической эволюции частота мутаций была значительно выше, чем в настоящее время, а соотношение "полезных" для организма мутаций к "вредным" сдвинуто в сторону первых. В пользу повышенной частоты мутирования на раннем этапе эволюции говорит тот факт, что в тот период значительно интенсивнее было действие на прокариотную клетку коротковолнового излучения при отсутствии у нее защитных приспособлений в виде соответствующих репарационных механизмов.
Следующее важное приобретение — сформирование механизмов обмена генетическим материалом, принадлежащим разным особям, "по горизонтали", и возникновение на базе этого особей с рекомбинантным геномом. При генетических рекомбинациях новых генов в генофонде, как правило, не появляется. В этом их принципиальное отличие от мутаций. Значение генетических рекомбинаций в том, что в результате этого обеспечивается возможность объединения разных генов и создание разных вариантов генных сочетаний в геноме прокариотной клетки. Поскольку отбор действует на всю совокупность признаков организма, генетическая рекомбинация поставляет дополнительный материал для действия отбора, ускоряя таким образом процесс эволюции.
Особенность организации генетической информации в мире прокариот — рассредоточение большого ее объема в нехромосомных элементах. Из этого следуют две существенно различающиеся возможности "горизонтального" обмена генетической информацией: первая связана с хромосомной, вторая — нехромосомной ДНК. Из трех основных процессов, приводящих у прокариот к обмену хромосомной ДНК, наиболее совершенным является процесс конъюгации, так как он обеспечивает возможность более полного обмена генетическим материалом двух клеток. (При благоприятных условиях возможно вхождение в реципиентную клетку всей донорной ДНК.) Однако эффективность механизмов генетической рекомбинации в этих процессах высока для близкородственных прокариотных организмов. Обмен участками хромосомной ДНК у бактерий в большинстве случаев ограничен пределами одного вида. Возможность "горизонтальной" передачи генетической информации на большие таксономические расстояния реализуется при переносе нехромосомных молекул ДНК, способных к автономной репликации.
М.В.Гусев, Л.А.Минеева © 1992-2001
Кафедра клеточной физиологии и иммунологии биологического факультета МГУ им. М.В.Ломоносова © 2000-2001
Страница обновлена 09.09.01
Оформление: А.В.Киташов