ГЛАВА 12. ПРОБЛЕМА ПРОИСХОЖДЕНИЯ И ЭВОЛЮЦИИ ЖИЗНИ. ВОЗНИКНОВЕНИЕ ПЕРВИЧНОЙ КЛЕТКИ
Согласно современным представлениям жизнь есть результат эволюции материи. Взгляды на происхождение жизни, ее развитие и сущность имеют длинную историю, но обсуждение этих вопросов до недавнего времени было предметом философских размышлений. Лишь в последние десятилетия решение этих вопросов было поставлено на экспериментальную основу и ответ на многие из них получен в лаборатории.
РАЗВИТИЕ ПРЕДСТАВЛЕНИИ О ПРОИСХОЖДЕНИИ ЖИЗНИ
Попытки ответить на вопрос, что такое жизнь, вероятно, следует отнести ко времени появления человека (Homo sapiens) . В самых ранних дошедших до нас памятниках культуры древнейших цивилизаций в художественной форме отразились существовавшие тогда представления о возникновении живых существ. При раскопках в Уруке, городе, существовавшем в середине IV тысячелетия до нашей эры, была обнаружена ваза, на которой изображено, как из морских волн появляются растения, над растениями располагаются животные, затем — люди, а над людьми — богиня жизни и плодородия.
Сведения о том, как различные живые существа возникают из воды и гниющих остатков, можно найти в древних китайских и индийских рукописях, об этом рассказывают египетские иероглифы и клинописи Древнего Вавилона. В Древнем Египте существовало убеждение, что лягушки, жабы, змеи и даже крокодилы рождаются из слоя ила, который остается после разливов Нила. В Древнем Китае считали, что тля возникает на молодых побегах бамбука. Большое значение при этом придавалось теплу, влаге и солнечному свету. Убеждение в спонтанном зарождении живых существ из неживых материалов было воспринято философами Древней Греции и Рима как нечто само собой разумеющееся. Первоначально вера в самозарождение не связывалась с определенным миропониманием. Самозарождение воспринимали как очевидный, постоянно наблюдаемый в природе факт. И только значительно позднее под самозарождение стали подводить определенную теоретическую основу, толкуя его с материалистических или идеалистических позиций.
Древнегреческий философ Фалес Милетский (конец VII — начало VI в. до н. э.) подходил к пониманию происхождения жизни со стихийно-материалистических позиций, считая, что жизнь есть свойство, присущее материи. Для Фалеса Милетского материальным первоначалом, из которого естественным путем возник мир, была вода. На позициях материалистического толкования самозарождения жизни стоял и другой древнегреческий философ Демокрит (460 — 370 гг. до н. э.). Согласно его теории, материя построена из атомов, мельчайших, неделимых, вечных и неизменных частиц, находящихся в движении, а жизнь возникла в результате взаимодействия сил природы, в особенности действия атомов огня на атомы влажной земли.
Противоположное идеалистическое толкование идеи самозарождения жизни связано с именем Платона (428/427 — 347 гг. до н. э.), считавшего, что сама по себе растительная и животная материя не является живой. Живой она становится только тогда, когда в нее вселяется бессмертная душа — "психея". Эта идея Платона оказалась очень жизнеспособной. Ее воспринял и Аристотель (384 — 322 гг. до н. э.), учение которого легло в основу всей средневековой научной культуры и господствовало около двух тысяч лет. В работах Аристотеля приводятся многочисленные "факты" самозарождения живых существ: растений, насекомых, червей, лягушек, мышей, некоторых морских животных. Необходимые условия для этого — наличие разлагающихся органических остатков, навоза, испорченного мяса, различных отбросов, грязи. Аристотель подвел под эти "факты" определенное теоретическое толкование, рассматривая внезапное появление живых существ как результат воздействия некоего духовного начала на безжизненную, косную материю.
В средние века идеи о возникновении живых существ из неживой материи подкреплялись новыми "фактами". Я. ван Гельмонт, голландский естествоиспытатель, известный своими исследованиями по питанию растений, предложил способ получения мышей, согласно которому, если открытый кувшин набить нижним бельем, загрязненным потом, и добавить туда некоторое количество пшеницы, то приблизительно через три недели появляется мышь, "поскольку закваска, находившаяся в белье, проникает через пшеничную шелуху и превращает пшеницу в мышь".
Развитие науки в эпоху Возрождения с ее экспериментальным подходом к изучению явлений природы поставило на повестку дня пересмотр с новых позиций идеи самозарождения Живых существ. Итальянский врач Ф. Реди (F. Redi, 1626 — 1698) решил проверить, действительно ли, как это всеми считалось, "черви" (личинки мух) зарождаются из гниющего мяса. Для этого он уложил мясо в три банки, одну из которых оставил открытой, вторую накрыл тонкой марлей, а третью — пергаментом. Все три куска мяса начали гнить, но "черви" появились только в открытой банке. Этим простым экспериментом Ф. Реди показал, что "черви" не возникли из гниющего мяса, а появились лишь там, где мухи могли откладывать яйца непосредственно на мясо. Опыты Ф. Реди впервые серьезно поколебали господствовавшую идею самозарождения макроскопических организмов.
После открытия А. ван Левенгуком микроорганизмов именно они стали основным объектом спора о зарождении жизни, поскольку логичным представлялось, что в первую очередь к самозарождению способны наиболее примитивно устроенные живые существа. Сам А. ван Левенгук отрицательно относился к возможности зарождения микроорганизмов из неживой материи.
Английский натуралист Дж. Нидхем (J. Needham, 1713 — 1781) попытался экспериментально ответить на этот вопрос. Дж. Нидхем поставил серию опытов, которые сводились к тому, что он готовил в стеклянных колбах разные настои, кипятил их в течение нескольких минут, затем закрывал обычными пробками. Через несколько дней в сосудах появлялись микроорганизмы. Это привело его к заключению о спонтанном возникновении микроорганизмов из неживого органического вещества, т. е. о возможности самопроизвольного зарождения на уровне низших живых существ.
Опыты Дж. Нидхема повторил итальянский естествоиспытатель Л. Спалланцани (L. Spallanzani, 1729 — 1799). Его опыты внешне не отличались от опытов Дж. Нидхема, за исключением того, что Л. Спалланцани закрывал сосуд пробкой не после, а до кипячения, а само кипячение длилось не несколько минут, как в опытах Дж. Нидхема, а значительно дольше — от 30 мин до 1 ч. В таких сосудах после выдерживания в течение нескольких дней не. было обнаружено никаких микроорганизмов. Л. Спалланцани сделал вывод, что в опытах Дж. Нидхема микроорганизмы в настоях появлялись, или попадая туда из воздуха (поскольку сосуды закрывали обычными пробками после кипячения), или погибали не все первоначально содержавшиеся в настоях клетки из-за недостаточно длительного кипячения. (В первую очередь это относится к наиболее термоустойчивым формам бактерий — спорам.) Л. Спалланцани под микроскопом удалось наблюдать деление микроба на две одинаковые дочерние клетки, каждая из которых также делилась на две клетки. Все сказанное позволило итальянскому ученому утверждать, что и микроорганизмы возникают не в результате самозарождения, а происходят от себе подобных. Выводы Л. Спалланцани, однако, не поколебали веры Дж. Нидхема и его сторонников в самозарождение. Дж. Нидхем объяснил отрицательные результаты, полученные Л. Спалланцани, тем, что тот подвергал свои настои слишком жесткой обработке, в результате которой разрушалась их "жизненная сила".
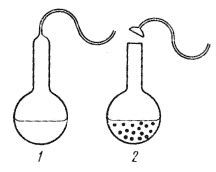
|
| Рис. 48. Опыт Л. Пастера в колбах с S-образными горлами: 1 — колба с подсахаренной дрожжевой водой; после стерилизации и охлаждения остается стерильной в течение длительного времени; 2 — та же колба через 48 ч после удаления изогнутого горла; наблюдается рост микроорганизмов (по Kenyon, Steinman, 1972) |
Окончательный конец спору о самозарождении микроорганизмов положил Л. Пастер. Серией четко поставленных опытов он доказал, что микроорганизмы не возникают самопроизвольно. Особенно изящными были его опыты, проведенные в колбах с S-образными горлами (рис. 48). В такие колбы наливали подсахаренную дрожжевую воду. Если колбы прокипятить, а затем осторожно охладить, то они остаются стерильными неопределенно долгое время, несмотря на то, что не закрыты пробками. Если же удалить S-образный участок горла, то спустя несколько дней в такой колбе будет наблюдаться бурное развитие микроорганизмов. Через S-образное горло непрогретый воздух может легко поступать в колбу, но содержащиеся в воздухе микроорганизмы задерживаются в изгибах горла, оседая в его нижнем колене. После удаления S- образной части горла микроорганизмы прямо попадают в колбу, начинается их быстрый рост. Этим простым опытом Л. Пастер опроверг возражение о разрушении при нагревании таинственной "жизненной силы", содержащейся в питательной среде и в обычном (непрогретом) воздухе. Он неопровержимо доказал, что "самозарождение" в большинстве опытов происходит в результате попадания в стерилизованные питательные среды микроорганизмов из воздуха.
Позднее идеи о самозарождении возникли уже в XX в. по отношению к субмикроскопическим живым частицам — вирусам. Однако и в этом случае было доказано, что вирусы не зарождаются из невирусного материала, а происходят только от себе подобных частиц, т. е. вирусов. Таким образом, хотя теория самозарождения была убедительно опровергнута на разных уровнях организации живых организмов, вопрос о происхождении жизни оставался открытым. Основной вывод, который можно сделать из рассмотренного выше материала, заключается в том, что в настоящее время (имеется в виду отрезок времени достаточной исторической протяженности) спонтанное возникновение жизни невозможно. Однако это не ответ на вопрос о происхождении жизни.
Точно так же не является ответом на вопрос и гипотеза о внеземном происхождении жизни и занесении ее на Землю в виде спор или зародышей с другой планеты34. Эта гипотеза не объясняет первоначального возникновения этих спор или зародышей, а просто истоки жизни выносит в просторы Вселенной. В настоящее время ни у кого не вызывает сомнения возможность существования жизни в других частях Вселенной, однако вероятность занесения на Землю живых организмов из космического пространства не имеет пока никаких подтверждений.
34 В конце XIX — начале XX в. большой популярностью пользовалась гипотеза панспермии, согласно которой живые организмы были занесены на Землю из космического пространства. Особенно привлекательно выглядела идея занесения их с метеоритами или космической пылью. Гипотеза панспермии была сформулирована в 1865 г. немецким исследователем Г. Рихтером (G. Richter) и поддержана С. Аррениусом (S. Arrhenius) и Г. Гельмгольцем (Н. Helmholtz). В наше время эту идею с учетом достижений науки и техники, и в первую очередь освоения человеком космического пространства, модернизировали Ф. Крик (F. Crick) и Л. Оргелл (L. Orgel), предположившие доставку зародышей жизни (микроорганизмов) на Землю из другой, более развитой цивилизации на космическом корабле.
Итак, на вопрос о возможности самозарождения в наше время живых существ из неживой материи был получен отрицательный ответ, и в этом огромная заслуга Л. Пастера. Однако многими современниками Л. Пастера его опыты, опровергавшие возникновение живых существ (микроорганизмов) из неживой материи, были восприняты как абсолютное доказательство полной невозможности зарождения живых организмов из неорганической природы35. Это поставило в тупик тех исследователей, которые видели в самозарождении единственный путь возникновения жизни.
В XX в. внимание к этой проблеме было привлечено советским биохимиком А. И. Опариным и английским исследователем Дж. Холдейном (J. Haldane), которые выдвинули предположение, что жизнь возникла в результате взаимодействия органических соединений, образовавшихся в бескислородных условиях на первобытной Земле. Согласно этой гипотезе, биологический синтез органических веществ происходит только на современном этапе существования Земли. На первобытной безжизненной Земле могли происходить химические (абиогенные) синтезы углеродистых соединений и их последующая предбиологическая эволюция. В результате этой эволюции имело место постепенное усложнение органических соединений, формирование из них пространственно обособленных систем и превращение последних в предшественников жизни, а затем и в первичные живые организмы. В последующие годы эти идеи получили широкое признание.
Конечно, вопрос о происхождении жизни — проблема общебиологическая. Более того, плодотворное его решение возможно только в комплексе с другими науками, такими как химия, геология, палеонтология, физика. Почему же этому вопросу так много внимания уделяется в курсе микробиологии? На это можно ответить словами К. ван Ниля: "...он (микробиолог) имеет дело с биологическим материалом, по-видимому, достаточно близким к "истокам жизни", и в то же время несет прямую ответственность за тупик, создавшийся вследствие того, что ему не удалось доказать самопроизвольное зарождение".
УСЛОВИЯ НА ДРЕВНЕЙ ЗЕМЛЕ
Возраст видимой нами Вселенной определяют как 10 — 15 млрд лет, а Земля возникла приблизительно 4,5 — 5,0 млрд лет назад. Согласно распространенным представлениям, образование Земли произошло путем аккумуляции холодных твердых тел. Первоначально Земля была довольно однородной и ее последующее изменение происходило в направлении дифференциации исходного гомогенного вещества на кору, мантию и ядро. Этот период, в течение которого происходило формирование Земли как единого твердого тела, завершился примерно 4,6 млрд лет назад. Для понимания процесса возникновения и эволюции жизни необходимо представлять, каковы были условия на Земле, в которых оказалось возможным "самозарождение" жизни. В последующий после сформирования Земли период на ней происходили активные геологические процессы, менявшие ее облик и приводившие к формированию земной коры, гидросферы и атмосферы.
35 Л. Пастер допускал возможность существования каких-то неизвестных условий, при которых могло произойти спонтанное зарождение жизни. В 1878 г. он писал, что не считает самозарождение в принципе невозможным.
На первобытной Земле основная масса воды находилась в связанном гидратированными породами состоянии, поэтому первоначально Мировой океан содержал меньше 10% того количества воды, которое содержат современные океаны. Остальные 90% образовались позднее за счет выделения паров воды из внутренних слоев Земли. Считается, что pH Мирового океана на протяжении всей истории Земли был довольно стабильным, в пределах 8 — 9. Формирование Мирового океана происходило, таким образом, постепенно, в тесной связи с формированием земной коры.
С формированием последней связано и образование атмосферы первобытной Земли, которая принципиально отличалась от современной атмосферы. По существующим представлениям атмосфера древней Земли, т. е. та атмосфера, в которой развивалась жизнь, имела восстановительный характер. Она содержала главным образом водород и его соединения (метан, аммиак, пары воды), в меньшем количестве — сероводород, азот, двуокись углерода и благородные газы. Эта атмосфера была лишена свободного кислорода. Возникновение атмосферы, содержащей O2, произошло значительно позднее и связано с жизнедеятельностью фотосинтезирующих организмов. Отсутствие свободного кислорода в первобытной атмосфере Земли имело принципиальное значение, поскольку органические вещества, образующиеся в этот период, не могли бы синтезироваться и сохраняться на протяжении геологических периодов в присутствии кислорода 36.
36 Получены данные о том, что УФ-излучение Солнца в первый миллиард лет его существования было в 100 000 раз интенсивнее, чем в наши дни. Поэтому первичный механизм образования O2 в предбиологической атмосфере Земли мог быть обусловлен разложением молекул водяного пара и CO2 из вулканических извержений под действием УФ-излучения молодого Солнца. А значит, предбиологическая атмосфера Земли могла содержать в миллион раз больше O2 чем это предполагается. Если эти данные получат подтверждение, возникнет необходимость пересмотреть некоторые из существующих идей о составе первобытной атмосферы Земли и условиях возникновения жизни.
Исходным материалом для синтеза органических веществ служили широко распространенные во Вселенной химические элементы: углерод, водород, кислород, азот, сера и фосфор. Однако синтез биологически важных молекул из этих элементов мог происходить только при условии обеспечения реакций свободной энергией, источником которой на первобытной Земле (как и на современной) были солнечное излучение, электрические разряды, тепловая энергия земных недр и радиоактивное излучение. Наиболее мощный из них — солнечное излучение. Поскольку молекулярный кислород в первобытной атмосфере Земли практически отсутствовал, не было и озонового экрана, существующего в современной атмосфере на высоте примерно 25 км от поверхности Земли и сильно поглощающего коротковолновую часть УФ-излучения. Можно представить, что значительная часть коротковолнового УФ проникала через атмосферу первобытной Земли и достигала ее поверхности, поэтому в условиях древней Земли длинноволновая часть солнечного излучения играла небольшую роль.
ВОЗМОЖНОСТЬ ОБРАЗОВАНИЯ ОРГАНИЧЕСКИХ ВЕЩЕСТВ НА ПЕРВОБЫТНОЙ ЗЕМЛЕ
Последовательность процессов возникновения органических веществ разной степени сложности можно представить следующим образом. В результате действия всех видов энергии из химических элементов синтезировались первичные соединения: углеводороды (в первую очередь метан), аммиак, цианистый водород, окись углерода, сероводород, простейшие альдегиды (и прежде всего формальдегид) и т. д. Эти соединения сами по себе не имели биохимического значения. Основным их свойством была высокая реакционная способность. Первичные соединения служили исходными веществами для образования биохимически важных органических соединений — мономеров. Из мономеров путем конденсации возникали полимеры — основные составные компоненты всех живых организмов.
В свое время А. И. Опарин и Дж. Холдейн высказали предположение о возможности моделирования процессов, происходивших на древней Земле. Это можно делать путем создания в лаборатории условий, имитирующих таковые, существовавшие на первобытной Земле. Выдвинутое положение стимулировало разработку экспериментальных подходов к изучаемой проблеме и оказалось весьма плодотворным. В настоящее время ряд процессов абиогенного синтеза сложных органических молекул, входящих в состав клеточных организмов, осуществлен в лабораторных условиях.
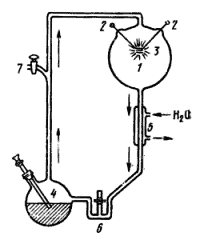
|
| Рис. 49. Схема прибора С. Миллера: 1 — реакционная колба; 2 — вольфрамовые электроды; 3 — искровой разряд; 4 — колба с кипящей водой; 5 — холодильник; 6 — ловушка; 7 — кран, через который в аппарат подается газовая смесь (по Kenyon, Steinman, 1972) |
Одним из первых в 1953 г. провел опыты по абиогенному синтезу биохимически важных соединений С. Миллер (S. Miller). Через газовую смесь, содержащую метан, аммиак, молекулярный водород и пары воды, т. е. имитирующую атмосферный состав первобытной Земли, он пропускал электрические разряды, а затем анализировал образующиеся продукты реакции. Схема прибора С. Миллера приведена на рис. 49. В реакционную колбу, содержащую смесь газов, были вмонтированы вольфрамовые электроды. В течение недели пропускали искровые разряды напряжением 60000 В. Содержащуюся в другой малой колбе воду поддерживали в состоянии кипения. Пары воды проходили через реакционную колбу и конденсировались в холодильнике. В процессе циркуляции они захватывали из реакционной колбы продукты реакции и переносили их в ловушку, где и осуществлялось их концентрирование. При идентификации продуктов реакции были обнаружены аминокислоты (глицин, a- и b-аланин, глутаминовая, аспарагиновая кислоты и др.) и органические кислоты (муравьиная, уксусная, пропионовая, гликолевая, молочная). По данным С. Миллера, основными первичными продуктами реакции в зоне разряда являются альдегиды и цианистый водород. Вторичные реакции, происходящие в водной фазе, приводят к образованию из них аминокислот и органических кислот.
В настоящее время в разных лабораториях осуществлен абиогенный синтез многих биологически важных мономеров. Большая информация получена относительно абиогенного синтеза аминокислот (табл. 14). Перечисленные в таблице аминокислоты образуются в простых по составу газовых или водных смесях в результате воздействия на них разными источниками энергии. При некотором усложнении реакционной смеси введением в нее C2-, C3-углеводородов, уксусного альдегида, гидроксиламина, гидразина и других соединений, образование которых легко происходит в условиях первобытной Земли, синтезируется значительно большее число аминокислот, в том числе и таких, которые не были обнаружены в качестве продуктов реакции в газообразных и водных смесях простого состава. Экспериментально доказано, что почти все аминокислоты, входящие в состав природных белков, можно получить в лаборатории при имитации условий первобытной Земли.
Таблица 14. Абиогенный синтез аминокислот
| Реагирующие вещества | Фаза | Источник энергии | Обнаруженные аминокислоты |
| CH4, NH3, H2, H2O | газовая | электрические разряды | аспарагиновая, аланин, глицин, диаминоянтарная, валин, гистидин, пролин, лизин, серии, аспарагин, аргинин, орнитин, глутаминовая, цистеин, таурин, цистамин |
| CO2, NH3, H2, H2O | газовая | электрические разряды | |
| CH4, CO2, NH3, H2, H2O, CO, N2 | газовая | рентгеновские лучи | |
| CH4, NH3, H2O | газовая | ультрафиолет | |
| NH3, HCN, H2O | водная | тепло (70°) | |
| CH4, NH3, H2O | газовая | b-лучи | |
| CH2O, N2, H2O | водная | солнечный свет | |
| H2S, NH3, H2O | водная | быстрые электроны |
В разных условиях и при воздействии разными источниками энергии из формальдегида абиогенным путем удалось синтезировать приблизительно 30 видов моносахаров (гексоз, пентоз, тетроз, триоз). Абиогенный синтез низших жирных кислот был обнаружен уже в опытах С. Миллера. Синтез жирных кислот, содержащих до 12 углеродных атомов, продемонстрирован после воздействия электрическими разрядами на смесь метана и воды. Абиогенное образование пуриновых оснований ввиду относительной сложности строения их молекулы представлялось весьма сомнительным. Однако испанский исследователь Дж. Оро (J. Oro) показал возможность синтеза аденина при нагревании водного раствора смеси HCN и NH3. Позднее были получены абиогенным путем и другие пуриновые основания. Удалось также синтезировать урацил из простых органических молекул.
Важный шаг на пути химической эволюции — синтез нуклеозидов и нуклеотидов, и в первую очередь адениновых. Американскому биохимику К. Поннамперума (К. Ponnamperuma) удалось показать, что при УФ-облучении смеси водных растворов аденина и рибозы при температуре 40° в присутствии фосфорной кислоты происходит реакция конденсации, приводящая к образованию аденозина. Если реакцию проводить при добавлении к реакционной смеси этилметафосфата, имеет место образование также и нуклеотидов: АМФ, АДФ, АТФ. Функция фосфорных соединений в этих химических синтезах двоякая: они играют каталитическую роль и могут непосредственно включаться в продукты реакции. Абиогенный синтез АТФ, представляющий собой результат нескольких относительно простых химических реакций, говорит о возможном раннем появлении этого соединения. Первые живые структуры могли получать АТФ из окружающей среды.
Следующий этап предбиологической эволюции — дальнейшее усложнение органических соединений, связанное с полимеризацией мономеров. Все живые клетки состоят из четырех основных типов макромолекул: белков, нуклеиновых кислот, липидов и полисахаридов. Из них белки и нуклеиновые кислоты являются самыми сложными веществами клетки.
С. Фоке (S. Fox) осуществил абиогенный синтез полипептидов, состоящих из 18 природных аминокислот, с молекулярной массой от 3000 до 10000 Да. Особенностью первичной структуры этих полимеров была обнаруженная у них определенная последовательность аминокислотных остатков в цепи, обусловленная, вероятно, структурными особенностями самих аминокислот. Полученные полимеры обладали многими свойствами, сближающими их с природными белками: служили источником питания для микроорганизмов, гидролизовались протеиназами, при кислотном гидролизе давали смесь аминокислот, обладали каталитической активностью и способностью к образованию микросистем, отграниченных от окружающей среды мембраноподобными поверхностными слоями. Из-за большого сходства с природными белками полипептиды, синтезированные С. Фоксом, были названы протеиноидами (белковоподобными веществами).
Принципиальная возможность образования полинуклеотидов. без участия ферментов была показана Г. Шраммом (G. Schramm). Образующиеся полимеры содержали от 60 до 200" нуклеотидов в цепи и имели молекулярную массу 15000 — 50000 Да. Так были получены полиадениловая, полиуридиловая, полицитидиловая кислоты и их сополимеры.
Таким образом, экспериментально показано, что в условиях первобытной Земли был возможен химический синтез биологически важных соединений (мономеров и полимеров), послуживших исходным материалом для построения всех организмов.
ВОЗНИКНОВЕНИЕ ПРОСТРАНСТВЕННО ОБОСОБЛЕННЫХ МИКРОСИСТЕМ
Химический синтез соединений углерода разной степени сложности мог привести только к накоплению органического вещества в гидросфере древней Земли. Для клеточной жизни характерно, что она всегда представлена в виде определенных структур, пространственно обособленных от внешней среды, но постоянно взаимодействующих с ней по типу открытых систем. Поэтому можно предполагать, что следующим этапом эволюции на пути возникновения жизни было формирование определенной структурной организации абиогенно синтезированных органических соединений. Этот этап эволюции также не является в настоящее время плодом умозрительных построений. Пространственно обособленные открытые системы можно получить экспериментальным путем из различных исходных компонентов.
С. Фоке, охлаждая растворенные в воде протеиноиды, получил микроскопические частицы, названные им микросферами, которые обладали определенной внутренней организацией и рядом интересных с биологической точки зрения, свойств. Смешивание раствора гуммиарабика и желатины приводит к формированию другого вида микроскопических структур, названных коацерватными каплями. Позднее было показано, что коацерваты возникают в результате объединения различных полимеров, например полипептидов и полинуклеотидов, при этом для получения коацерватов основное значение имеет не специфичность внутримолекулярного строения образующих их компонентов, а степень их полимеризации. Такие пространственно обособленные открытые системы, построенные из полимеров, были названы протоклетками37.
37 Термин "протоклетки", по-видимому, можно рассматривать в качестве аналога названия "прогеноты" (см. с. 160).

|
| Рис. 50. Протеиноидные микросферы С. Фокса (по Fox, 1965) |
Рассмотрим коротко некоторые свойства микросфер, взяв их в качестве модели протоклетки. Протеиноидные микросферы имеют сферическую форму, диаметр их в зависимости от условий получения колеблется от 0,5 до 7 мкм (рис. 50). По величине и форме они напоминают кокковые формы бактерий, иногда образуют цепочки, похожие на цепочки стрептококков. Каждая микросфера содержит около 1010 молекул протеиноида. Протеиноидные микросферы обладают определенной стабильностью: не разрушаются при центрифугировании, в солевых растворах устойчивее многих препаратов коацерватных капель. Их стабильность позволила приготовить препараты для электронной микроскопии, на которых удалось рассмотреть некоторые детали ультраструктуры. При изменении условий внешней среды наблюдали движение материала внутри частицы от центра к периферии, деление микрочастицы и образование двойного пограничного слоя. Окрашивание по Граму обнаружило, что микросферы, образованные из кислых протеиноидов, грамотрицательны; микросферы, в состав которых входят в достаточном количестве основные протеиноиды, грамположительны. Из других свойств, присущих микросферам и представляющих интерес с эволюционной точки зрения, можно указать на существование у них барьеров с избирательной проницаемостью; способность к делению и почкованию: подвижность, возрастающую после добавления к суспензии микросфер АТФ; способность к наращиванию массы микрочастицы; тенденцию к контактированию друг с другом. В протеиноидных микросферах найдена ферментоподобная активность, которой обладали образующие их протеиноиды. Однако этот вопрос для микросфер нуждается в дальнейшем исследовании, поэтому проблему каталитической активности в протоклетках мы разберем на модели коацерватных капель.
Интенсивные исследования по изучению коацерватных капель как модели доклеточной организации были проведены А. И. Опариным с сотрудниками. Обычно коацерватные капли получают, сливая растворы противоположно заряженных коллоидов, например желатины и гуммиарабика, гистона и РНК, гистона и желатины и т. д. При смешивании исходно гомогенных растворов каждого компонента образуются сферические частицы. Концентрация полимеров в частице на один-два порядка выше, чем в окружающем растворе. Коацерватные капли отделены от раствора четко выраженной поверхностью, способны избирательно поглощать из среды некоторые вещества (аминокислоты, сахара, мононуклеотиды) и выделять в среду продукты протекающих в них реакций.
Один из наиболее интересных опытов с коацерватными каплями состоял в том, что в коацерваты, образованные из гистона и гуммиарабика, вводили фермент фосфорилазу, а затем эти капли помещали в раствор глюкозо-1-фосфата. Коацерватные капли поглощали из раствора глюкозо-1-фосфат, и в них осуществлялось ферментативное превращение глюкозо-1-фосфата в крахмал, за счет скопления которого увеличивались размеры капли. Если в коацерватные капли вводить два фермента (фосфорилазу и b-амилазу), то в них имеет место последовательно ферментативное превращение глюкозо-1-фосфата в крахмал и крахмала в мальтозу, которая диффундирует из капли в раствор (рис. 51). Из приведенного примера видно, что коацерватные капли являются хорошей моделью открытой системы. Они способны поглощать из окружающей среды вещества и энергию, преобразовывать их в продукты синтеза или распада; продукты синтеза входят в состав капли, обеспечивая наращивание ее массы, а продукты распада выделяются в среду. Скорости ферментативных реакций в коацерватных каплях существенно выше, чем в гомогенных растворах. Особенно четко различие в скоростях проявляется при сочетании действия двух ферментов. Опыты с коацерватами показали важность надмолекулярной структурной организации и, в частности, ее значение для функционирования клеточных катализаторов.
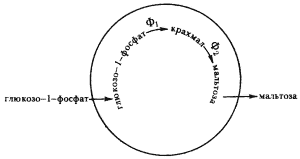
|
| Рис. 51. Синтез и гидролиз крахмала в коацерватной капле: Ф1 — фосфорилаза; Ф2 — (b-амилаза (по Опарину, 1976) |
На модели коацерватных капель была показана связь между уровнем внутренней организации капель и их способностью к наращиванию массы. Оказалось, что в одинаковых условиях капли, обладающие более совершенной экспериментально созданной внутренней организацией, наращивают массу быстрее,. чем капли, внутренняя организация которых менее совершенна. Для последних характерны также меньшая стабильность и более быстрый распад. Естественно, что дальнейшая судьба обоих типов коацерватных капель неодинакова. Очевидно преимущество коацерватов, обладающих большей стабильностью в условиях окружающей среды и более длительным временем существования.
Пространственно обособленные системы с определенным уровнем структурной организации приобрели новые свойства, отсутствующие у образующих их органических соединений. Эти свойства (зачатки метаболизма, способность к самоподдержанию структуры и наращиванию массы) присущи более высокому уровню организации материи, поэтому их можно рассматривать как зачатки тех свойств, дальнейшее развитие которых. в совокупности привело к возникновению живых клеток.
ЭВОЛЮЦИЯ ПРОТОКЛЕТКИ НА ПУТИ ВОЗНИКНОВЕНИЯ ПЕРВИЧНОЙ КЛЕТКИ
Как из гипотетической протоклетки возникла первичная клетка, способная к самовоспроизведению, до сих пор не известно. В лабораторных условиях не удалось получить самореплицирующуюся систему из простых предшественников. Поэтому мы можем остановиться только на некоторых процессах, имеющих определенное теоретическое или экспериментальное обоснование, необходимых для зарождения первичной клетки: появлении асимметрии живых организмов, возникновении и эволюции каталитической активности и матричного синтеза.
Возникновение оптической активности
В природе существует несколько видов пространственной изомерии (стереоизомерии). Один из них основан на присутствии в молекуле асимметрического атома углерода, связанного с четырьмя различными группами. Такие стереоизомеры характеризуются одинаковыми физическими и химическими свойствами, но обладают способностью вращать плоскость поляризованного света вправо или влево. В зависимости от этого различают право- и левовращающие (D- и L- соответственно) оптические стереоизомеры. Явление оптической изомерии химических соединений было открыто основоположником современной микробиологии Л. Пастером, который имел химическое образование.
Отличительная особенность всех живых организмов состоит в том, что органические соединения, из которых они построены, представлены одним из возможных оптически активных стереоизомеров. Например, за исключением глицина, самой простой аминокислоты, не обладающей оптической активностью, все аминокислоты, входящие в состав природных белков, состоят из L-, сахара, образующие полисахариды, — D- форм. В то же время при синтезе органических веществ в лабораторных условиях всегда получается равномерная смесь обеих форм стереоизомеров, называемая рацемической смесью. В этом проявляется принципиальное различие между синтезами органических веществ, происходящими в живой клетке и в лабораторных условиях. В первом случае продукт биосинтетической реакции представляет собой вещество в определенной оптической форме, во втором — продуктом реакции является смесь обеих оптических форм синтезированного вещества.
Асимметричный синтез клеткой органических веществ происходит на базе уже существующей в них асимметрии. Таким образом, вопрос сводится к тому, как впервые возник асимметричный синтез. В современной литературе можно найти значительное количество гипотез, объясняющих происхождение оптической активности. Согласно одной из них, возникновению жизни должно было предшествовать сильное нарушение зеркальной симметрии в виде скачкообразного перехода (как это имеет место при кристаллизации). По проведенным расчетам, в условиях первобытной Земли скачкообразный переход существовавших органических молекул из симметрического состояния в асимметрическое — событие весьма вероятное. Основные этапы процесса, по этим представлениям, следующие: первый этап — абиогенное образование и накопление органических молекул в виде рацемических смесей; следующий этап — нарушение зеркальной симметрии в рацемическом "бульоне" и формирование только одного типа асимметрических молекул: L-аминокислот и D-сахаров, из которых образуются короткие цепочки молекул — блоков будущих ДНК, РНК и белков. Принципиальное значение стереоизомерии в возникновении жизни заключается в том, что способностью к точной репликации (самовоспроизведению) и, следовательно, к передаче точной информации обладают только полимерные молекулы, построенные из асимметрических мономеров одного типа, т. е. только L-типа для аминокислот и D-типа для сахаров. Полинуклеотиды, синтезированные из мономеров разного типа, способностью к точной репликации не обладают.
Возникновение и эволюция каталитической активности
Уже пространственно обособленные открытые системы обладали примитивным метаболизмом в том смысле, что их структурная организация создавала благоприятные условия для протекания определенной последовательности биохимических реакций. В основе метаболизма современных клеток лежит совершенный каталитический аппарат, поэтому эволюционное развитие протоклеток связано также с развитием и совершенствованием их каталитических активностей. Первыми катализаторами, доступными для протоклеток, были относительно простые органические и неорганические соединения внешней среды. Хорошо известна способность солей ряда металлов ускорять реакции переноса водорода. Каталитическая активность этих неорганических соединений очень невысока. Оказалось,. что ее можно существенно повысить при сочетании неорганических соединений с некоторыми органическими молекулами. Например, ионы железа могут в незначительной степени ускорять реакции переноса водорода. Если железо ввести в порфириновое кольцо, каталитическая активность этого комплекса будет в 1000 раз выше, чем ионов железа. Можно представить, что аналогичный путь усовершенствования простых катализаторов имел место в процессе эволюции протоклеток.
Примером комплексов, возникших в результате сочетания различных молекул (органических и неорганических), могут быть современные коферменты. Число известных коферментов невелико, но они являются универсальными, присущими всем живым организмам катализаторами38. Универсальность современных коферментов говорит об их раннем возникновении в процессе формирования метаболического аппарата, а их стабильность на протяжении столь длительного процесса эволюции — о наилучшем из всех возможных вариантов соответствии выполняемым функциям. Протоклетки, будучи предельно гетеротрофными, вначале, вероятно, просто заимствовали сложные коферменты из внешней среды и только значительно позднее у них (или у более совершенных клеток) развилась способность , к самостоятельному синтезу коферментов.
38 Исключение составляют некоторые представители архебактерий.
Дальнейшее усложнение метаболизма потребовало более четкого согласования последовательностей составляющих его биохимических реакций. Коферменты, обладающие каталитической активностью, значительно более низкой, чем современные ферменты, и не обладающие свойством субстратной специфичности, на определенном уровне развития клеточного метаболизма не могли отвечать необходимым требованиям. Поэтому они были заменены или дополнены более мощными и совершенными катализаторами — ферментами. Скорости реакций, катализируемых ферментами, примерно в 1010 раз выше, чем скорости неферментативных реакций.
Вероятно, первым в процессе эволюции у предшественников современных ферментов появилось свойство каталитической активности, а свойство субстратной специфичности возникло значительно позднее. В качестве предшественников современных ферментов можно рассматривать простые пептиды, для которых показана способность ускорять определенные реакции, в частности, реакции гидролиза, аминирования различных соединений, а также реакции карбоксилирования a-кетокислот. Эволюция ферментных белков из предшественников — простых пептидов — прошла длительный путь в направлении наилучшего приспособления их первичной, вторичной и третичной структур к выполняемым функциям.
Возникновение матричного синтеза
Как известно, в современных клетках функции ДНК заключаются в получении, хранении и передаче информации последующим поколениям. Без ДНК и РНК невозможно точное воспроизведение всех свойств клетки, в основе которых лежит функционирование специфических белков. В модельных опытах была показана относительная простота и легкость возникновения пространственно обособленных систем, построенных из протеиноидов, характеризовавшихся определенным постоянством аминокислотных последовательностей. Это могло служить указанием на то, что информация о полипептидах типа протеиноидов была заключена в них самих, а следовательно, подводило к следующему выводу: на начальном этапе эволюции протоклетки могли воспроизводиться и передавать информацию потомству без участия нуклеиновых кислот.
Дальнейшее усложнение структуры и совершенствование функции полипептидов приводило к появлению в них определенных аминокислотных группировок, которым в какой-то степени была присуща полезная для протоклетки каталитическая активность. Однако возникновение более "совершенного" полипептида создавало преимущество для породившей его протоклетки только в том случае, если появившееся определенное сочетание аминокислотных остатков в полипептиде могло быть передано дочерним протоклеткам. При отсутствии такой способности возникшее "удачное" сочетание аминокислотных остатков в полипептиде терялось при последующем разрастании протоклеток. Таким образом, для дальнейшей эволюции протоклеток необходимо было создание специального аппарата, который обеспечивал бы в ряду их поколений точное воспроизведение полипептидов с определенно закрепленным расположением аминокислотных остатков. Это привело к формированию принципиально нового механизма синтеза — матричного синтеза, в основе которого лежит использование свойств нового класса органических соединений — полинуклеотидов.
Свойством полинуклеотидов, сформированных из одного типа асимметрических молекул, является способность к точному воспроизведению, основанная на принципе структурной комплементарности. В модельных опытах было показано, что поли- нуклеотидная цепь может служить матрицей, связывающей свободные нуклеотиды. При смешивании АМФ с полиуридиловой кислотой свободные молекулы АМФ связываются с остатками полиуридиловой кислоты при помощи водородных связей между комплементарными основаниями. В результате возникала спиральная структура. Точно так же наблюдали формирование устойчивой комплементарной спирали при смешивании полицитидиловой кислоты с гуанозинмонофосфатом. Для синтеза комплементарных полинуклеотидов необходимо было, чтобы между связанными с матрицей мононуклеотидами образовались межнуклеотидные связи. Экспериментально была показана принципиальная возможность возникновения таких связей без какого-либо участия ферментов. Таким образом, полинуклеотиды могли служить матрицей для неферментативного синтеза комплементарных полинуклеотидов.
Вопрос о том, каким путем в молекулах полинуклеотидов возникла и закрепилась информация о структуре белков, остается наиболее неясным. Имеются данные об избирательном взаимодействии между двумя типами полимеров — полиаминокислотами и полинуклеотидами — в зависимости от их аминокислотного и нуклеотидного составов, на основании чего высказывается предположение, что в принципе полиаминокислоты и полинуклеотиды могли "узнавать" друг друга в протоклетках. Образование специфических комплексов между этими полимерами можно рассматривать как первый необходимый шаг на пути установления между ними определенных "информативных" связей. Не исключено также, что на первых этапах поток информации шел в любом направлении (полинуклеотид протобелок) и, таким образом, устанавливались взаимные связи между определенными последовательностями аминокислот в протобелках и нуклеотидов в полинуклеотидах. Позднее поток информации стал однонаправленным (полинуклеотид протобелок).
Таким образом, дискуссионным остается вопрос о том, на каком этапе эволюционного процесса нуклеиновые кислоты сформировались как информационные молекулы. Согласно одним представлениям на начальном этапе эволюции роль последних выполняли белковоподобные молекулы, и первые примитивные клетки функционировали без нуклеиновых кислот. Другая гипотеза исходит из того, что первыми возникли нуклеиновые кислоты, а позднее, на базе содержащейся в них информации, возникли белки (гипотеза "генной жизни").
Эта гипотеза принадлежит американскому генетику Г. Мёллеру (Н. Muller), высказавшему предположение, что "жизнь" началась с абио-генного образования гена или группы генов. Появление мембран и белков, обладающих каталитическими свойствами, имело место на более поздних этапах. В пользу этой гипотезы приводятся соображения, первое из которых основано на современном представлении о молекулярной структуре и самовоспроизведении вирусов, а второе — на полифункциональных свойствах мононуклеотидов. Хорошо известно, что нуклеотиды, помимо того, что составляют генетический аппарат клетки, принимают участие в разнообразных метаболических реакциях: служат переносчиками энергии (АДФ, АТФ), электронов и атомов водорода (НАД, НАДФ, ФМН, ФАД), сахаров, ацильных групп и др.
Формы жизни, возникшие на "белковой основе", были неустойчивыми из-за отсутствия системы передачи информации, использующей свойства нуклеиновых кислот, а "генная жизнь" не могла прогрессивно эволюционировать без участия белков, обладающих каталитическими свойствами. Как произошло возникновение формы жизни, в основе которой лежат белки и нуклеиновые кислоты, пока не известно. Ясно только, что "встреча" обоих типов соединений положила начало пути эволюции, на котором произошло формирование механизмов синтеза белка и нуклеиновых кислот и кодовых взаимодействий между обоими механизмами.
ДАННЫЕ ПАЛЕОНТОЛОГИИ О ПРОИСХОЖДЕНИИ ЖИЗНИ НА ЗЕМЛЕ
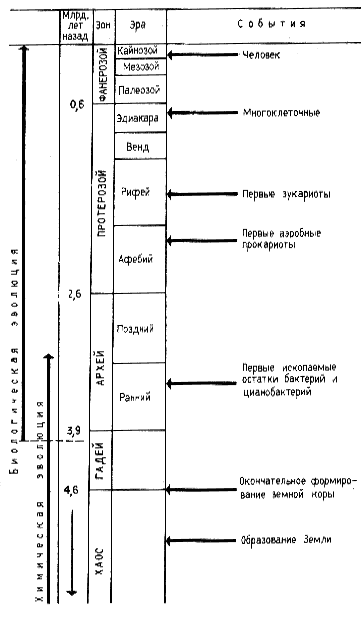
|
| Рис. 52. Этапы биологической эволюции (по Опарину, 1976; Fox, Dosel, 1975; Lehninger, 1974) |
Согласно современным представлениям окончательное формирование земной коры произошло около 4,6 млрд лет назад. Наши сведения об истории возникновения и развития жизни на Земле ограничены преимущественно последним периодом, длительность которого порядка 600 млн лет39. Остальной временной период, составляющий примерно 90% всей истории существования Земли, фактически является чистой страницей в изучении возникновения и развития жизни на Земле. Поэтому большой интерес представляют данные молекулярной палеонтологии изучающей органические вещества древнейших осадочных отложений. Трудность заключается в интерпретации полученных результатов, т. е. в отсутствии надежных критериев на основании которых можно было бы делать выводы о происхождении обнаруженных органических остатков: биогенном или абиогенном. В этой связи интересны находки, сделанные в Южной Африке в осадочных породах, возраст которых составляет около 3,5 млрд лет. В этих породах найдены заключенные в них окаменелые остатки палочковидных структур размером 05x025 мкм, напоминающих современные бактерии. При электронно-микроскопическом изучении у них выявлена двухслойная клеточная стенка, подобная клеточной стенке многих современных бактерий.
39 В геологии (см. рис. 52) приняты следующие названия временных интервалов: зон, эра, период, эпоха, век. Самое крупное геохронологическое образование — зон. Зоны подразделяются на эры и т. д. Фанерозой, самый последний и наиболее изученный зон, включает три эры: палеозой, мезозой и кайнозой Первый период палеозойской эры — кембрий (начало примерно 580 млн лет назад, длительность — 80 млн лет). Характеризуется появлением животных, имеющих скелетные элементы. Более ранние зоны часто объединяются под названием докембрия или дофанерозоя.
В породах, возраст которых также около 3, 5 млрд лет, обнаружены строматолиты, своеобразные известковые образования, являющиеся продуктами жизнедеятельности древних фотосинтезирующих организмов — цианобактерий, или синезеленых водорослей. Если принять, что найденные в породах ископаемые остатки действительно принадлежат древнейшим прокариотам или являются продуктами их жизнедеятельности, то следует признать, что к этому времени уже были сформированы некоторые типы жизни, которые дошли до нас в виде ее "следов" Отсюда приходится сделать вывод, что впервые земная жизнь должна была возникнуть в промежутке между 3,5 и 4,6 млрд лет тому назад, однако у нас нет никакой информации об этом периоде. Схематическое изображение во времени отдельных этапов эволюции представлено на рис. 52.
Цианобактериям мы обязаны появлением молекулярного кислорода в атмосфере Земли. Однако вначале весь выделяемый ими Оа поглощался земной корой, в которой происходили интенсивные процессы окисления. По имеющимся геологическим данным содержание кислорода в атмосфере достигло 1% от его содержания в современной атмосфере только в среднем протерозое, и к этому времени можно отнести возникновение первых аэробных прокариот. В пользу этого свидетельствуют обнаруженные в отложениях, возраст которых около 2 млрд лет, звездчатые образования, свойственные облигатно аэробной свободноживущей бактерии Metallogenium. Этот организм откладывает на поверхности клеток окислы железа. В природе встречается при разных концентрациях O2, но всегда в аэробных условиях, так что может служить индикатором молекулярного кислорода.
Первые эукариоты появились приблизительно 1,5 млрд лет назад. Таким образом, прокариоты были единственными обитателями нашей планеты в течение 2/3 времени эволюции биосферы. Жизнедеятельность прокариот привела к накоплению в атмосфере молекулярного кислорода и к обогащению лито- и гидросферы органическим веществом.
М.В.Гусев, Л.А.Минеева © 1992-2001
Кафедра клеточной физиологии и иммунологии биологического факультета МГУ им. М.В.Ломоносова © 2000-2001
Страница обновлена 09.09.01
Оформление: А.В.Киташов