
СУЖДЕНИЕ ПАЛЕОНТОЛОГА ОБ ЭВОЛЮЦИИ
Л.Татаринов, академик
…Лучшие свои
доказательства Дарвин должен
был взять из неисчислимых собраний
рассыпанных
в земле остатков животных…
БюллетеньМосковского
общества испытателей
природы за 1860 год.
К концу ХIХ века практически все палеонтологи приняли идею эволюции, хотя далеко не все были согласны с Дарвином в оценке ее движущих факторов. Прежде всего это касается приверженцев сальтационизма - теория «скачков», идеи о мгновенном, в течение одного-двух поколений превращение видов, не связанных переходными факторами.
Сальтационизм родился задолго до создания научной теории эволюции; Элементы этих взглядов можно встретить у мыслителей Древней Греции. Казалось, сальтационизм отошел в прошлое, однако в последнее время многие исследователи в той или иной форме вернулись к идее о скачкообразных превращениях видов.
Против Дарвина выступили и сторонники автогенетических и эктогенетических взглядов. Первые полагают, что ход эволюции определен внутренними, заложенными в организме силами. Эктогенетики, напротив, считают, что ход эволюции целиком определяет среда.
Напомним, что по Дарвину основными факторами эволюции является естественный отбор в процессе внутри и межвидовой конкуренциии наследственная изменчивость, которая имеет неопределенный, ненаправленный характер. После смерти Дарвина прошло уже больше ста лет, а дискуссия по вопросу о движущих силах эволюции продолжаются, причем критики дарвинизма пытаются обосновать старые идеи новыми данными, которые черпают из современной экспериментальной биологии и палеонтологии.
И хотя материал палеонтолога обычно не позволяет изучать наследственность, отбор и борьбу за существование, он имеет первостепенное значение для суждений, например, о направленности или темпе эволюции.
Направленность. Что ее определяет?
Палеонтология знает много примеров того, как эволюция целых групп организмов на протяжении десятков миллионов лет шла в определенном направлении. Пример такой эволюции лошадей стал уже хрестоматийным. Их предок, гиракотерий, который жил около 50 миллионов лет назад, был размером с лисицу и имел четыре пальца на передних и три – на задних конечностях. Но чем ближе к современности, тем ископаемые представители этой группы больше становятся похожи на нынешних рослых «однопалых» лошадей.
Факты такого рода приводили многих палеонтологов к выводу, что изменчивость организмов идет в определенном направлении. Но ведь по Дарвину наследственная изменчивость, которая есть материал для эволюции, не имеет направленности. Как разрешить противоречие?
Ответ состоит в том, что эволюция в таких рядах (биологи называют их ортогенетическими) становится направленной не оттого, что направлена наследственная изменчивость, а оттого, что ее ограничивает естественный отбор. Если на протяжении миллионов лет отбор действует примерно в одном направлении (такой отбор называется ортоселекцией), то конечный результат будетпочти таким же, как если бы направлена была исходная наследственная изменчивость. В примере с лошадиными ортоселекция на протяжении десятков миллионов лет действовала в сторону их приспособления к жизни в степных ландшафтах (они развивались в то же геологическое время) – к быстрому бегу и травоядности.
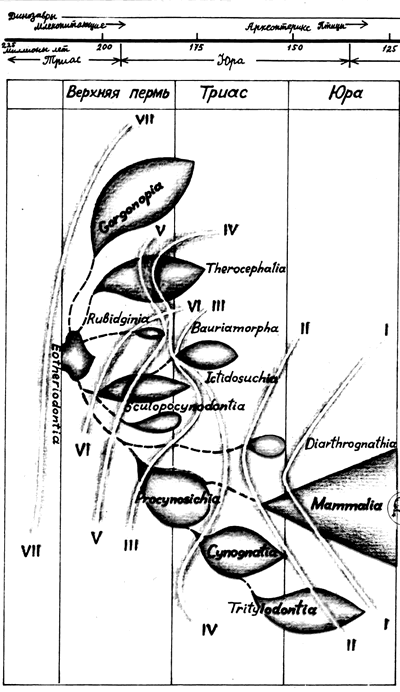 |
Генеалогическое дерево млекопитающих, составленное академиком Татариновым. Как видно, они - современники динозавров. Есть даже точка зрения, что первый примат успел увидеть последнего динозавра |
То, что в этом случае мы имеем дело с направленным отбором, а не направленной изменчивостью, подтверждают факты постоянного ответвления от магистрального ствола развития лошадиных разнообразных боковых ветвей. Вымирание – отпадение этих боковых линий – придало картине эволюции лошадиных направленный характер.
И хотя анализ ископаемого материала на индивидуальную изменчивость веститрудно, потому что в одном слое могут оказаться останки нескольких поколений, изменчивость которых уже прошла ворота отбора, но примеры разнообразных и разнонаправленных индивидуальных отклонений все же подтверждают, что в эволюции вектор отбора значит неизмеримо больше, чем вектор наследуемых изменений.
Можно ли прогнозировать эволюцию?
О направленности эволюции в известной мере свидетельствуют явления конвергенции и параллелизма – сходства признаков.
Яркий пример конвергенции – веретенообразная форма тела у разделенных миллионами лет по времени происхождения акулы, ихтиозавра и дельфина. Но. При конвергенции ясно выходит на первый план зависимость приспособления от функции организма (в нашем примере речь идет о приспособлении к быстрому плаванию в воде). Значит, конвергенция – всегда следствие отбора, следствие приспособления.
Сложнее ситуация с параллелизмами. При них, кроме зависимости от функции, отчетливо проявляется зависимость приобретаемого признака от внутренних особенностей организма. Самая частая форма параллелизма – многократное появление сходных особенностей строения в группах родственных организмов. Параллелизмы могут обнаруживаться на очень глубоких уровнях, - вплоть до уровня генотипа. Генетические параллелизмы, например, являются важной составляющей в знаменитой "Периодической таблице признаков" - гомологических рядах Николая Ивановича Вавилова. Особенно впечатляют параллелизмы в группах, осваивающихновую среду обитания.Например, у всех членистоногих,когда и где бы они ни переходили к наземному образу жизни, независимо становится жесткой хитиновая оболочка. Похожие превращения в покровах происходят и при выходе на сушу позвоночных животных. Но если в этих разделенных десятками миллионов лет примерах параллелизма приспособительное значение параллельных внутренних изменений более или менее ясно, то в других случаях оно может быть неопределенным. На такой пример указывает Валентин Абрамович Красилов, когда анализирует независимое свертывание чашелистиков в трубку в самыхразличных линиях предков цветковых растений.
Итак, распространенность вызванных «изнутри» параллелизмов действительно подтверждает «внутреннюю» направленность филогенезма - процесса образования видов.
И все-таки факт направленности не позволяет делать вывод о возможности прогнозирования эволюции конкретных организмов на отдаленное будущее. Ведь даже в таком направленном ряду форм, как развитие лошадиных, вряд ли можно сказать, двинется ли ряд в будущем по «магистральному пути» или «отклонится в сторону». Ход эволюции зависит от множества факторов, связанных с процессами, протекающими в биосфере, и подчас случайно влияющих на организм в ходе развитияи вид в ходе эволюции. Поэтому прогнозирование эволюции на основе простой экстраполяции пути изменений, пройденного в прошлом, может оказатьсяошибочным.
Советский анатом и палеонтолог А.П.Быстров сделал в свое время такую экстраполяцию для человека будущего, который, по его прогнозу, наделен огромным мозгом, беззубыми челюстями, укороченными шеей и поясницей и трехпалыми руками. Правда, А.П.Быстров оговорил, что таков был бы облик человека будущего, если бы эволюция человека контролировалась естественным отбором. Мне кажется, что и для животных и для растений прогнозирование даже ближайших шагов эволюции возможно лишь в очень приближенной степени. Прогнозирование отдаленных результатов эволюции, которые наступят через сотнимиллионов лет,практически невозможно.
Темп. Плавно или скачками движется эволюция?
Дарвин придерживался мнения, что эволюция происходит весьмапостепенно, путем накопления мелких изменений. «Природа не делает скачков», - любилон повторять вслед за Лейбницем. Но сторонники точки зрения, что эволюция совершается сальтациями (скачками), в последнее время стали заметно активнее. Почему? На каких фактах они основываютсвою позицию?
На уровне мутаций трудно провести грань между небольшими скачками и постепенными (градационными) изменениями. Однако известны и случаи, когда относительно крупные перестройки происходят скачком. Такие сальтации встречаются у птиц при переносе сосудов с одного ствола на другой. У большинства птицтаким путем исчезаетправая сонная артерия. Это скачок. Путем своеобразных сальтаций изменяется число позвонков, образуется новыеанастомозы-перемычки между сосудами.
Конечно, наибольший интерес вызывает вопрос, могут ли скачком не только меняться признаки, но и возникать виды даи группы организмов более высокого ранга? По концепции прерывистого равновесия, виды остаются неизменными миллионы лет, а затем за несколько десятков или сотен лет формируются новые. Хотя создатели концепции подчеркивают, что переход от вида к виду происходит «по-дарвинистски» - путем накопления мутаций и отбора, но придают чрезмерное значение в образовании видов случайным, неприспособительным изменениям и отдельным мутациям с крупным эффектом.
Одним из хорошо исследованных примеров, на которые опирается концепция прерывистого равновесия,является эволюция озерных моллюсков. Турканы, области на севере Кении.Их развитие в течение почти пяти миллионов лет было изучено на массовом материале. Оказалось, что новые виды или появляются внезапно, или преобразование видов происходит за пять – пятьдесят тысяч лет. Длительность стабильного стазиса, во время которого не происходит крупных изменений, составляет не меньше миллионов лет.
Но. Во-первых, практически во всех случаях новые виды моллюсков возникали после перерывов в накоплении озерных осадков, вслед за прослоями вулканических туфов. Значит, эволюционные события в перерывах постепенно развития моллюсков не документированы, а происходили за пределами исследованной территории. То есть речь не об отсутствии эволюции, а об отсутствии фактов о ней. Во-вторых, в некоторых случаях старые и новые виды моллюсков соединены рядом переходных форм (видообразование шло плавно).
Все это говорит о неполноте документации хода эволюции на уровне видов даже в классическомслучае с моллюсками Турканы. И в других примерах «прерывистой эволюции» перерывы чаще всего отмечаются, когда есть основания говорить о перерывах времени наблюдения за последовательностью сменяющих друг друга поколений.
Зато, например, для остатков планктонных организмов, добытых из кернов скважин при океаническом бурении показано очень постепенное превращение одних видов в другие.
К сторонникам скачкообразной эволюции – сальтационистам – примыкают современные катастрофисты, которые полагают, что биота Земли периодически изменяется после массовых вымираний, вызванных глобальными катастрофами возможно, космического происхождения. При этом нередко звучит утверждение, что массовые вымирания освобождают экологические ниши и поэтому являются необходимым условием для вымирания новых групп организмов, заселяющих «пустые» ниши.
Особенно часто глобальной катастрофой объясняют преобразования в органическом мире Земли на границе мела – последнего периода мезозойской эры, и палеогена, первого периода кайнозоя, 66,4 миллиона лет назад, когда окончательно вымерли динозавры. При этом утверждают, что именно вымирание динозавров создало предпосылки для экспансии млекопитающих.
Кризис, который длился миллионы лет
По традиционным представлениям поэтапная смена флоры и фауны происходит из-за постепенного изменения жизненных условий и конкуренции вновь возникающих групп животных и растений с древними. Таким образом В.А.Красилов, который пытается проследить этапы изменений в мезозойских биоценозах, вызванные экспансией новой группы растений -покрытосеменных, пока он обсуждает факты, придерживается некатастрофистской точки зрения.
Надо отметить, что в последние годы традиционные представления стали менее популярны. Наиболее распространенная гипотеза катастрофистов связывает массовые вымирания в геологической истории со столкновениями Земли с крупными астероидами. При этом основная роль отводится непрямым последствиям соударения. Предполагают, что вещество астероида, который взрывается при столкновении с Землей, распыляется в атмосфере и буквально заслоняет Солнце. Начинается «астероидная зима». Температура Земли резкоснижается на многие месяцы, а фотосинтез приостанавливается. Все это, по мнению катастрофистов, и приводит к массовому вымиранию организмов и запустению экологических ниш.
Действительно, геологами открыты воронки, оставшиеся после падения астероидов. Диаметр воронок составляет подчас десятки километров. Особенно впечатляет, что в прослое глин, который отделяет известняки верхнемеловых слоев от слоев нижнего палеогена, повышено содержание иридия и некоторых других металлов платиновой группы. Катастрофисты считают, что этот иридий имеет астероидное происхождение. Кроме того, обнаружилось, что промежуточный слой глин обогащен гранулами графита. По мнению катастрофистов, графит – след глобального лесного пожара, вызванного падением астероида. Правда, не исключено, что иридий в пограничном прослое имеет вулканическое происхождение, но, как бы то ни было,действительно есть много доказательств тому, что Земля время от времени сталкивается с крупными астероидами.
Однако последствия столкновенийне могли быть столь грандиозны, как это считают крайне катастрофисты. Во всяком случае, различные группы мезозойских организмов вымирали не одновременно и вымерли подчас задолго до образования иридиевого прослоя. Так, вымирание аммонитов завершилось, по-видимому, за сто тысяч лет до конца мелового периода. Другая группа меловых моллюсков – иноцерамы – вымерла даже за миллион лет до конца мела. Задолго до конца мелового периода начались процессы вымирания мезозойских рептилий: ихтиозавров, панцирных динозавров, гигантских завроподов, таких, как бронтозавры и диплодоки. И хотя хищные динозавры, гидрозавры и рогатые динозавры действительно вымерли в самом конце мела, в целом можно утверждать, что процесс смены мезозойской биоты на кайнозойскую был не одномоментным, а многоэтапным. Группы животных и растений кайнозойского облика начали появляться за десятки миллионов лет до конца мезозоя (в частности, покрытосеменные растения, насекомые-опылители, ящерицы, змеи, млекопитающие). Понятно, что невозможно все этапы этой смены объяснить одной катастрофой, приуроченной к самому концу мела. Не объясняют гипотезы катастрофистов и избирательного характера вымирания. Ведь многие группы животных пересекли границу мезозоя, как бы не заметив ее. Так обстояло дело с ящерицами, птицами и млекопитающими. Наконец, климатические последствия возможного падения астероидов в конце мела не были такими уж значительными, если на Земле не исчезали пояса тропического климата с характерной для них флорой и фауной. Ведь даже кратковременного глубокого похолодания в тропиках хватило бы для гибели благополучно перешедших в кайнозой коралловых рифов, крокодилов и гигантских черепах.
Неверны и представления катастрофистов, которые считают, что экспансия млекопитающих началась только после вымирания динозавров. Хотя по размеру мезозойские млекопитающие и были невелики, число их видов и родов к концу мезозоя вполне можно сопоставить с числом видов и родов динозавров. Поданным Д.Рассела, в самых верхних мезозойских слоях в Северной Америке насчитывается 34 рода динозавров и 29 родов млекопитающих.
И все же – отчего?
Вымирание организмов при смене эр геологической истории, безусловно, имело множество причин. Его нельзя свести к последствию возможныхкатастроф. Избирательность вымирания и распространения различных групп есть косвенное доказательство того, что существенную роль в процессе смены сыграла конкуренция старых и новых групп и общая перестройка биологических межгрупповых отношений – биоценозов. Перестройка биоценозов была связана с экспансией новых групп и флоры, в частности, цветковых растений. И в этом отношении концепция В.А.Красилова мне кажется более правильной, чем взгляды крайних катастрофистов.
В замещении динозавров и птерозавров млекопитающими и птицами большую роль могло сыграть то, что эти современные нам группы явно обгоняли рептилий напути эволюционного прогресса еще в мезозое. Быстрее росла их активность, улучшался аппарат питания, они стали теплокровными, а млекопитающие переходили к живорождению. Особое значение имело совершенствование головного мозга и усложнение поведения. Млекопитающие уже могли успешно соперничать если не со взрослыми, то с молодыми динозаврами, избирательное истребление которых должно было резко уменьшитьчисленность ящеров. Отметим, что гигантизм некоторых их видов мог вести к побочным отрицательным последствиям. Поскольку координация ощущений, которые приходят из разных частей огромного тела, требует определенного времени, то ограничивается скорость реакции, а это – проигрыш в межвидовой борьбе.
Некоторую роль могло сыграть и то, что в самом конце мезозоя, как раз на переходе в кайнозойскую эру, климат стал холоднее и суше, что, конечно, ухудшило условия существования гигантских ящеров.
«Естественный отбор, писал Дарвин, - никогда не может делать внезапных больших скачков, а всегда подвигается короткими, но верными, хотя и медленными шагами». И хотя теперь ясно, что сальтационные изменения составляют часть в общей сумме эволюционных изменений, по-прежнему нет серьезных свидетельств тому, что в результате скачков, без постоянного контроля со стороны естественного отбора могут возникать новые группы организмов более сложных, чем вирусы и бактерии.
Что касается направленности, которая подтверждает большую роль сложившейся организации в определении путей дальнейших эволюционных изменений, то она, видимо, недооценивается современной версией дарвинизма - синтетической теорией эволюции. Вместе с тем создается впечатление, что случайные процессы могут играть более существенную роль, чем это предполагали классики дарвинизма.
Видимо, значение для хода эволюции как внутренних организменных факторов, так идействующих извне случайных процессов постоянно колеблется. Основной силой, определяющей ход развития жизни, как гениально показал Дарвин, остается естественный отбор.
Хотя во времена Дарвина не было научной генетики и молекулярной биологии, но его теория эволюции сохранила жизнеспособность в наши дни. Кажущаяся простота основных построений Дарвина лишь увеличивает восхищение, которое все мы или большинство из нас чувствуем по отношению к этому мыслителю.