В.А. Красилов, Е.В.
Бугдаева,
В.С. Маркевич, Н.П. Маслова
Проангиоспермы и происхождение цветковых растений

Цветковые – наиболее многочисленная и разнообразная группа высших растений, доминирующая в большинстве наземных экосистем. Именно к цветковым относятся основные культивируемые растения, от которых, в конечном счете, зависит наше существование. Поэтому разработка системы, отражающей историческое развитие цветковых и позволяющей предсказать свойства многих тысяч малоизученных и вновь открываемых видов, имеет не только теоретическое, но и практическое значение.
Еще Ч.Дарвин говорил о “проклятой тайне” происхождения цветковых, имея в виду их кажущееся внезапным появление в геологической летописи и невозможность распознать предковые формы. Хотелось бы лучше понять события, изменившие облик планеты, но существующая теория эволюции не дает ключа к их расшифровке.
Нами были получены факты, позволяющие приблизиться к разгадке
“проклятой тайны”. Собранные материалы, относящиеся к древнейшим цветковым Дальнего
Востока, Забайкалья, Монголии, Ближнего Востока и Австралии, были изучены с
помощью современных методов электронной микроскопии и разработанных нами
методов, в частности электронно-микроскопического изучения субкрустаций –
минеральных реплик, сохранивших клеточное строение растительных тканей, а также
извлечения растительных остатков из желудков ископаемых насекомых. Для каждого
образца изучены данные о геологическом возрасте, условиях захоронения и
сопутствующих остатках животных и растений.
материалы, относящиеся к древнейшим цветковым Дальнего
Востока, Забайкалья, Монголии, Ближнего Востока и Австралии, были изучены с
помощью современных методов электронной микроскопии и разработанных нами
методов, в частности электронно-микроскопического изучения субкрустаций –
минеральных реплик, сохранивших клеточное строение растительных тканей, а также
извлечения растительных остатков из желудков ископаемых насекомых. Для каждого
образца изучены данные о геологическом возрасте, условиях захоронения и
сопутствующих остатках животных и растений.
Полученные результаты позволили сформулировать ряд эволюционных гипотез, относящихся к факторам и механизмам становления цветковых растений.
Древнейшие покрытосеменные появляются в средних широтах Северного и Южного полушарий 120–115 млн лет назад. В древнейших слоях, относящихся к этому периоду, найдены пыльцевые зерна с дырчатым покровом и другими признаками репродуктивной самонесовместимости, характерными для покрытосеменных. Немного позднее к ним присоединяются крупномерные остатки: листья, цветки, плоды, причем первые листья, как правило, очень мелкие, с “дезорганизованным” сетчатым жилкованием, цветки невзрачные, собранные в соцветия, плоды также мелкие, нередко снабженные хохолками или зацепками для распространения животными.
Местонахождения древнейших покрытосеменных встречаются главным образом в переходной полосе между тропической и умеренной зонами, которая в меловом периоде находилась приблизительно вдоль параллелей 50° с.ш. и 40° ю.ш. При этом сходные события происходили одновременно в нескольких регионах, т.е. существовало несколько крупных центров параллельного развития цветковых растений. Среди них стоит особо выделить байкальско-гобийский центр. О его существовании можно было догадываться по известным ранее единичным находкам пыльцевых зерен и листьев в нижнемеловых отложениях верховьев реки Витим в Забайкалье. Нам удалось значительно пополнить подобные находки в Забайкалье и обнаружить новые местонахождения древнейших покрытосеменных в Западной Монголии. Здесь впервые появляются травянистые формы, а также, судя по остаткам плодов, растения с признаками семейства ореховых – одного из наиболее древних среди современных покрытосеменных. Подобные плоды недавно обнаружены также в нижнемеловых отложениях Махтеш-Рамона в Южном Израиле. Там они встречаются в вулканических отложениях, что позволяет достаточно точно определить их возраст. Таким образом, можно считать доказанным существование и ближневосточного центра происхождения древнейших цветковых растений, причем, несмотря на географическую удаленность, эволюционные события в обоих центрах развивались одновременно.
Еще более удивителен параллелизм ранней истории цветковых растений на Азиатском и Австралийском континентах. На юге последнего (в штате Виктория) был крупный независимый центр происхождения цветковых растений – Кунварра. Наиболее примечательными оказались находки плодов с признаками цератофилловых – обособленного по морфологическим и биохимическим признакам современного семейства водных цветковых. При этом в одновозрастных отложениях Забайкалья были найдены остатки цветковых, очень близкие морфологически австралийским.
Примечательная особенность всех этих местонахождений состоит в
том, что они приурочены к отложениям древних рифтовых долин (рифт – зона растяжения земной коры,
где между двумя расходящимися плитами появляется трещина, из которой изливается
магма), расположенных как на территории Забайкалья и Монголии, так и в пустыне
Негев и на юге Австралии (бассейн Джиппсленд). По-видимому, древнейшие
покрытосеменные возникли в рифтовых долинах, по которым и происходило их
первоначальное расселение. Экологические условия в древних рифтах, вероятно,
характеризовались исключительно высоким биологическим разнообразием и
продуктивностью наземных и водных экосистем. Можно предположить, что вулканизм,
неоднородность геологического субстрата, геохимических и микроклиматических
условий создавали тенденцию к ускорению развития, которая отчетливо выражена у
примитивных покрытосеменных, и способствовали эволюционным новообразованиям.
древних рифтовых долин (рифт – зона растяжения земной коры,
где между двумя расходящимися плитами появляется трещина, из которой изливается
магма), расположенных как на территории Забайкалья и Монголии, так и в пустыне
Негев и на юге Австралии (бассейн Джиппсленд). По-видимому, древнейшие
покрытосеменные возникли в рифтовых долинах, по которым и происходило их
первоначальное расселение. Экологические условия в древних рифтах, вероятно,
характеризовались исключительно высоким биологическим разнообразием и
продуктивностью наземных и водных экосистем. Можно предположить, что вулканизм,
неоднородность геологического субстрата, геохимических и микроклиматических
условий создавали тенденцию к ускорению развития, которая отчетливо выражена у
примитивных покрытосеменных, и способствовали эволюционным новообразованиям.
Еще одним важным эволюционным фактором становления цветковых растений было взаимодействие с животными, которые способствовали распространению плодов и участвовали в опылении цветковых растений. Уже упоминавшиеся зацепки на древнейших плодах свидетельствуют о распространении их млекопитающими, птицами, а в случае водных форм, возможно, и личинками стрекоз и водных жуков. Нам удалось обнаружить хорошо сохранившуюся пыльцу в желудках ископаемых насекомых различного геологического возраста. Юрские кузнечиковые и близкие к ним формы – крупные насекомые, достигавшие 15 см в размахе крыльев, – питались преимущественно пыльцой Classopollis, у которой впервые развиваются сложные проростковые структуры, характерные для цветковых. Это лишь первые конкретные доказательства взаимодействия растений и насекомых в геологическом прошлом. Дальнейшее накопление подобных данных может пролить свет на роль сопряженной эволюции в становлении крупных групп растительного и животного мира.
 |
|
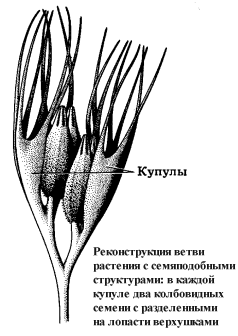
Во всех изученных нами центрах происхождения древнейшим покрытосеменным сопутствуют растения, которые формально не относятся к этой группе, но обладают важнейшими характерными для нее признаками – листьями с многопорядковым сетчатым жилкованием, пыльцевыми зернами с разнообразными проростковыми щелями и порами, цветкоподобными и плодообразными структурами, семенами, заключенными в купулы, которые аналогичны, а в ряде случаев, может быть, и гомологичны завязи. Мы назвали такие растения проангиоспермами. Признаки, свойственные покрытосеменным – настоящим ангиоспермам, – у них еще не получили полного развития и к тому же проявляются разрозненно, не образуя устойчивых сочетаний.
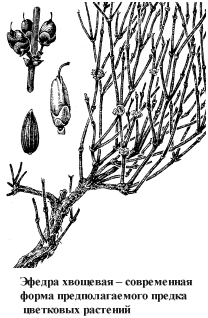
К проангиоспермам относятся недавно открытые вымершие формы гнетовых. Сходство гнетовых с покрытосеменными (проявляющееся не только в морфологических признаках, но и в развитии зародышевого мешка и оплодотворении, предваряющем двойное оплодотворение покрытосеменных), разумеется, не ускользнуло от внимания морфологов и в свое время даже послужило основой для “эфедровой” теории происхождения цветковых. Однако высокая специализация современных форм гнетовых, представленных тремя морфологически далеко разошедшимися родами, и отсутствие каких-либо сведений о геологическом прошлом этой группы растений не позволяли считать их возможными предками покрытосеменных. Для серьезного исследования эволюционной роли гнетовых просто не было достаточных данных.
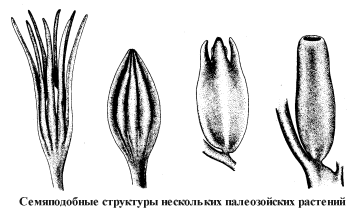
Изучение ископаемых гнетовых началось с открытия нового рода – Eoantha – в нижнемеловых отложениях Забайкалья, где он встречается вместе с древнейшими покрытосеменными. За этой находкой последовали новые, свидетельствующие о заметной роли гнетовых в мезозойской растительности и их значительном морфологическом разнообразии. Недавно нами изучен еще один новый род гнетовых, обнаруженный в уже упоминавшемся австралийском нижнемеловом местонахождении Кунварра, который близок современной эфедре. В тех же растительных сообществах заметную роль играли и другие проангиоспермы. Так, в Байсинском местонахождении Забайкалья кроме эоанты встречены своеобразные околоводные растения Baisia, произошедшие от мезозойских бенниттитов. Их органы размножения – купулы с пучками длинных волосков – напоминали семянки пушицы и других осоковых. В местонахождении Махтеш-Рамон (Израиль) первым покрытосеменным сопутствуют кейтониевые – еще одна широко распространенная группа проангиоспермов.
Разнообразные проангиоспермы входили в состав одних и тех же растительных сообществ и подвергались давлению одних и тех же факторов среды. Некоторые из них оказались в эволюционном плане более перспективными, чем другие. Однако нельзя утверждать, что лишь какая-то одна линия проангиоспермов дала начало всем покрытосеменным. Мы можем констатировать лишь то, что признаки различных групп покрытосеменных появляются не в одной, а в нескольких линиях проангиоспермов. Так, у изученных нами чекановскиевых семена развивались в двустворчатых купулах с краевыми рыльцевыми гребнями, характерными для наиболее примитивных магнолиецветных. В то же время кейтониевые демонстрируют возможность параллельного возникновения иного типа завязи с многочисленными базальными семяпочками. Уже упоминавшаяся байсия имела купулы с единственной прямой семяпочкой, развивавшейся на верхушке цветоложа, как у злаков и близких к ним однодольных.
Одним из существенных достижений двух последних лет было детальное морфологическое изучение Eoantha и родственных ей гнетофитов, листья которых по внешней морфологии и анатомическим признакам соответствуют листьям злаков. Принимая во внимание также существенное сходство репродуктивных структур, можно теперь с известной долей уверенности говорить о филогенетической близости гнетовых и злаков – самой важной для человека группой цветковых растений.
Учитывая большое морфологическое разнообразие проангиоспермов и то, что вегетативные и продуктивные структуры, которые могли быть прототипами соответствующих структур разных групп цветковых растений, появились в разных эволюционных линиях, нельзя утверждать, например, что злаки произошли от магнолиевых или другой стволовой группы двудольных. Палеонтологическая летопись свидетельствует о независимом происхождении основных эволюционных линий покрытосеменных от разных проангиоспермов, которые, в свою очередь, происходят от птеридоспермов, беннеттитов и других древних голосеменных. В генетическом плане цветковые, таким образом, унаследовали значительную часть развивавшегося в течение многих миллионов лет генофонда семенных растений. В этом основа их исключительно высокого биологического разнообразия и больших эволюционных потенций.
Словарь терминов
Беннеттиты – вымершая группа семенных растений с однополыми или двуполыми шишками, характеризующимися циклическим расположением частей и напоминающими цветок.
Гнетовые – группа семенных растений, представленная в современной флоре тремя родами, для которых характерны сосуды в древесине и свободноядерное развитие зародышевого мешка. По этим признакам гнетовые близки к покрытосеменным.
Кейтониевые – вымершие мезозойские растения с сетчатым жилкованием листьев и семенами, развивающимися в сферических купулах.
Купула – открытое или сомкнутое вместилище семян (семяпочек).
Чекановскиевые – вымершие древесные растения с игольчатыми или лентовидными листьями и семяпочками в двустворчатых купулах.