Историческая зоогеография наземных тетрапод.
Н.Н.Каландадзе, А.С.Раутиан
Палеонтологический институт, Российская Академия Наук.
Около двадцати лет авторы изучали наземных тетрапод и, используя некоторые несложные методы, получили серию зоогеографических реконструкций с карбона до плейстоцена (рис. 1.1-15).
Некоторые из этих результатов были опубликованы на русском языке и, мы полагаем, что было бы полезно сделать краткое резюме этой работы, которое будет переведено на английский язык.
Некоторые результаты выглядят очень необычно. Но наши реконструкции - не плод воображения. Они являются проекцией системы наземных тетрапод на географию (см. Материалы и методы). И, если наши реконструкции ошибочны, это значит, что неверна система наземных тетрапод.
Конечно, это не значит, что все наши реконструкции абсолютно верны. Все они могут и должны быть откорректированы. Но, следуя примеру А.Р.Уоллеса, мы исследовали все массивы суши Земли и использовали весь доступный нам материал. Мы получили некоторые новые результаты и сейчас их можно уточнять.
Приблизительно 1% наземных тетрапод “несогласно” с нашими реконструкциями. Часть из них обсуждена в разделе “Некоторые таксономические результаты”.
Одним из наиболее важных аспектов этой работы является методика, которой мы пользовались и некоторые заключения о систематическом положении ряда групп, которые трудно получить, используя обычные зоологические методы.
Материалы и методы.
Мы использовали данные о географическом и стратиграфическом распространении наземных тетрапод (не морских и не летающих). Каждый таксон рассматривался как монофилетичный, монотипичный и монохронный по происхождению. Фауны сравнивались по степени их таксономической изоморфности на уровне от семейства до рода включительно. На первой стадии работы все фауны сравнивались с каждой другой попарно, несмотря на их прежнее или настоящее положение на поверхности Земли. При анализе каждой пары фаун мы использовали две операции.
I - операция “проваливания”. Если данный таксон, указывающий на прямую связь между двумя фаунами, встречается в обеих этих фаунах в предшествующее время и эта связь может быть объяснена присутствием этого таксона в предшествующее время, мы исключали этот таксон из рассмотрения зоогеографии данного времени.
II - операция “редукции”. Если прямая связь между двумя фаунами двух регионов является слабой и может быть полностью сведена к связям между этими фаунами с третьей (или больше) фаунами (то есть если каждый таксон на прямой связи присутствует на непрямых, более сильных, связях), такая связь полагалась фальшивой. Ее можно объяснить другими, опосредованными, связями (см. Каландадзе, Раутиан, 1980-1991).
Результаты и дискуссия.
Поздний девон.
Наземные тетраподы позднего девона известны из разных территорий (Гренландии, России, Австралии), но их совершенно недостаточно для успешных зоогеографических исследований. Мы пробовали использовать данные по кистеперым и двоякодышащим рыбам, но не получили удовлетворительного результата скорее всего потому, что эти рыбы не являются строго пресноводными. То есть они могли пересекать морские преграды.
Карбон
Две известные фауны карбоновых наземных тетрапод С. Америки и З. и Ц. Европы настолько похожи, что можно рассматривать их как две части единой фауны. Этот факт хорошо известен и наш анализ подтвердил его (Каландадзе, Раутиан, 1981).
Зоогеографический метод глобально начинает работать фактически только с пермского периода. Для девона и карбона данных по наземным тетраподам недостаточно. В перми нет современных континентов и океанов. Из древних достоверно нам известен, (но известен очень хорошо), только океан Тетис.
Очевидно, что помимо Тетиса должны были быть еще большие площади, занятые океаном (или океанами), но надо сразу же честно сказать, что мы о них решительно ничего не знаем. Все, что было накоплено этими океанами в виде осадков, в настоящее время существует в виде горных систем Земли. Отдельные куски страниц этой гигантской летописи разбросаны в разных уголках планеты и так перепутаны, что .....
В дальнейшем для каждого времени дать наиболее характерных животных для каждой территории.
Ранняя пермь (рис. 1.1).
Высокий уровень эндемизма показывает, что в ранней перми мы имеем три разные фауны на трех территориях.
1. Северная Америка + северная часть Западной Европы.
Наиболее обычными являются Lepospondyli и архаичные
лабиринтодонты: Edopida и Eryopida. Некоторые крупные амфибии
показывают высокую степень адаптации к жизни вдали от воды.
Они имели хорошо окостеневший массивный скелет, и у них нет
следов органов боковой линии на черепе и нижней челюсти. Так
как такие амфибии известны с позднего карбона до середины
перми, можно сказать, что в позднем карбоне - ранней перми
амфибии пытались “организовать” первую наземную биоту
позвоночных - амфибий, не рептилий.
Со среднего карбона мы знаем несколько групп рептилий (Cotylosauria, Diapsida, Theromorpha), которые были способны изгнать амфибий из их новых экологических ниш. Поэтому, с начала поздней перми, большинство амфибий является водными животными.
Наиболее обычными наземными животными С. Америки были Pelycosauria, но не Therapsida (!). В 60-е годы некоторые фрагментарные остатки тероморф С. Америки были определены как остатки Therapsida. Основываясь на данных исторической зоогеографии, мы предположили, что все эти остатки могли принадлежать сфенакодонтным пеликозаврам (Каландадзе, Раутиан, 1983) и проф. Р.Рейсц (перс. сообщ.) в целом подтвердил, что все эти фрагменты принадлежат пеликозаврам.
Мы полагаем, что остатков терапсид не будет найдено в С. Америке до позднего триаса, когда все континенты соединились и терапсиды вместе с первыми млекопитающими вторглись в С. Америку.
До сих пор мы не знали достоверных парарептилий в С. Америке. Р.Рейсц, однако, полагает, что Acleistorhinus родственен Lanthanosuchia (Reisz, 1996). В этой же работе из С. Америки им указана форма, близкая или тождественная Macroleter (Reisz, 1996, fig. 3). Если это так, то Acleistorhinus и Macroleter - первые достоверные представители парарептилий в С. Америке. Однако, существование в перми С. Америки восточноевропейских родов представляется нам очень мало вероятным (см. рис. 1.1).
2. Западная Европа + Средняя Азия + Китай.
Наиболее
обычными лабиринтодонтами здесь были Archegosauroidea и
своеобразные небольшие амфибии - Discosauriscidae. Как и
М.Ф.Ивахненко, мы полагаем дискозаврисцид древнейшими
амфибийными предками парарептилий. Этот отдельный ствол
рептилий - Parareptilia - М.Ф.Ивахненко, развивая
представления проф. Э.Олсона, прослеживает от Discosauriscidae
до Pareiasauria, Procolophonia и Chelonia (Ивахненко,
1987).
3. Южная Америка + Африка + ?
Мы знаем очень немного
тетрапод с этих территорий. Но известный род Mesosaurus,
скелеты которых обычны Ю. Африке и Бразилии, подтверждает, что
эти континенты были соединены в ранней перми. Мы не знаем
хороших остатков раннепермских наземных тетрапод из Индии и
Австралии.
Поздняя пермь - средний триас (рис. 1.2).
В поздней перми и начале триаса существуют два больших материка: Северный – Лавразия, располагавшийся в теплой зоне и тропиках, и Южный – Гондвана, располагавшийся в Южном полушарии. Именно поэтому нам известны несомненные следы огромных площадных оледенений на юге: Южной Америки, Африки, Индии и Австралии. Конечно, оледенение тогда охватывало и Антарктиду. Но в конце перми и триасе оледенение пошло на убыль и в Южной Африке (Карру), Индии, Австралии и Антарктиде появились флоры и фауны, - очень сходные, – то, что мы называем Гондванской фауной.
Кстати сказать, Гондвана названа по небольшому индийскому племени мелких охотников – гондов. Киплинг прославил их как прекрасных охотников в Книге Джунглей (Маугли).
В начале поздней перми и раннем и среднем триасе некоторые части Лавразии (Шотландия, Россия и Китай) были связаны с Гондваной. Предварительные исследования показали, что в целом зоогеографическая картина была одинаковой с поздней перми до среднего триаса и что С. Америка, а так же З. и Ц. Европа в это время была изолированы.
Фауны этих территорий состоят из некоторых древних, архаичных групп. Различия между этими двумя фаунами невелики и мы полагаем, что они принадлежали к разным провинциям одной зоогеографической области - Лавразии. Некоторые группы примитивных амфибий и капториноморф известны из этого региона, особенно из С. Америки. Во всяком случае, обе эти фауны демонстрируют существенное отличие от фауны Гондваны.
Так как все остальные фауны показывают большое сходство, мы полагаем, что они принадлежат Гондванской зоогеографической области. Ю. Америка, Африка, Индия, Антарктида и Австралия были соединены в то время и все фауны, известные с этих территорий были очень похожи.
Ситуация с северными фаунами не так ясна. Известная позднепермская фауна Шотландии (Элгин) включает только три рода: Geikia, Gordonia (Anomodontia) и Elginia (Pareiasauria). Anomodontia и Pareiasauria типичны для Гондваны, но не Лавразии (Каландадзе, Раутиан, 1981).
После открытия проф. В.П.Амалицким Северодвинской фауны с парейазаврами, горгонопсами, тероцефалами, дицинодонтами и цинодонтами, родство пермских фаун России с гондванскими стало очевидным.
Сейчас мы можем сказать, что большой континентальный массив от Шотландии до Китая был соединен в это время. И некоторые провинциальные различия этих северных и южных фаун подтверждают наше заключение, что в поздней перми - среднем триасе северные континенты соединялись с Гондваной по крайней мере дважды: в начале поздней перми (низы Ecca zone или раньше) и в начале раннего триаса (Lystrosaurus zone).
Наиболее обычными животными Гондванской зоогеографической области были парарептилии (Pareiasauria и Procolophonia), Therapsida и пресноводные Labyrinthodontia.
Известна одна очень важная группа животных, “несогласная” с нашей реконструкцией - некоторые триасовые Labyrinthodontia. В начале триаса (в З. Европе - с поздней перми) некоторые гондванского типа лабиринтодонты внезапно появляются на изолированных территориях. На рис. 2а и 2б стрелками показаны моменты изменения фаун амфибий в С. Америке и в З. и Ц. Европе. Эти группы лабиринтодонтов дали единственный пик в обеих частях Гондванской зоогеографической области: на юге (рис. 3) и на севере (рис. 3б, в).
Должна быть причина, объясняющая, почему в С. Америке и З. Европе лабиринтодонты дали два пика разнообразия, а в Гондване и В. Европе - один (Каландадзе, Раутиан, 1983).
Так как никаких терапсид не появилось в С. Америке и З. Европе до позднего триаса (за исключением Procynosuchus sp. из Германии), мы рассмотрели этих лабиринтодонтов и нашли, что все они принадлежат к семействам, представители которых иногда встречаются в прибрежно-морских отложениях: Capitosauridae, Benthosuchidae, Trematosautidae и Plagiosauridae.
Можно предположить, что начиная с триаса, некоторые лабиринтодонты пытались освоить морскую воду и, значит, могли пересекать морские проливы. Но, начиная с раннего триаса уже известны Ichthyopterygia, Sauropterygia и Placodontia. Это может объяснить, почему после позднего триаса известны только единичные находки юрских и раннемеловых лабиринтодонтов в Китае, Монголии, Средней Азии, России и Австралии. Очевидно, они были вытеснены рептилиями.
В поздней перми мы знаем одно очень важное изменение в наземной биоте. В начале поздней перми, известна уникальная Очерская фауна. Все наземные животные из Очера - терапсиды: крупные и мелкие, хищные и растительноядные.
В более поздней, Северодвинской фауне, все хищники были терапсиды (горгонопсы и тероцефалы), а растительноядные принадлежали к двум группам: дицинодонтам (Therapsida) и парейазаврам (Parareptilia). Крупные формы растительноядных терапсид начали вымирать.
В последней позднепермской фауне России - Вязниковской было найдено два хищника: крупный тероцефал и первый архозавр (Archosaurus). Это - очень важный момент в истории наземных тетрапод: крупные хищные терапсиды начали замещаться архозаврами. С этого времени Theromorpha (а позднее и Mammalia) существовали “в тени архозавров”. Две трети истории млекопитающих прошло “в тени динозавров” (Lillegraven, Kielan-Jaworowska, Clemens, 1979).
С поздней перми до позднего триаса мы знаем очень мало крупных тероморф и все они - дицинодонты. Однако мелкие тероморфы существовали до палеоцена (Chronoperates из С. Америки). Произошедшие от терапсид и вытесняемые динозаврами из крупного размерного класса, все мезозойские млекопитающие были мелкого размера.
Мелкий размер, ночь, нора, лес и кустарник - вот удел млекопитающих вплоть до начала кайнозоя.
Поздний триас (рис. 1.3, рис. 4).
Поздний триас - время максимальной регрессии. Зоогеографическая Пангея. Все континенты соединены, и Северная Африка является центром мира.
Данные исторической географии показали, что в позднем триасе наряду с Тетисом (океаном предшествующей генерации), уже существуют закладки всех ныне существующих океанов: Атлантического, Индийского и Тихого.
Появляются: черепахи, крокодилы, динозавры, птерозавры и млекопитающие (Chelonia, Crocodylia, Dinosauria, Pterosauria и Mammalia).
Некоторые провинциальные различия фаун позднего триаса показывают, что современные океаны начали раскрываться в триасе. Мы провели специальное исследование позднего триаса используя материалы по Thecodontia, Dinosauria, Theromorpha и Mammalia (Каландадзе, Раутиан, 1991).
Все следы предшествующих зоогеографических подразделений исчезли в позднетриасовой зоогеографической Пангее. Таким образом, поздний триас - наиболее раннее время, до которого можно пытаться построить историко-зоогеографические реконструкции, используя только современный материал.
Ранняя - средняя юра (рис. 5а, б).
Величайшая катастрофа наземной биоты с девона до современности (Каландадзе, Раутиан, 1983-1993; Benton, 1985-1989).
Эта катастрофа и есть причина, по которой мы не смогли построить реконструкции, пользуясь нашим методом. Хотя наземные тетраподы были найдены практически везде (в С. Америке, Европе, Азии, Африке, на Мадагаскаре и в Австралии; см. рис. 4); объем материала слишком мал (рис. 5а, 5б; Табл. 1). Специальные статьи были посвящены анализу этой катастрофы (Каландадзе, Раутиан, 1993 а, б).
Ряд высокого ранга таксонов вымерло и появилось в окрестностях юрского кризиса. Вот некоторые из них:
| Lepospondyli | Apoda | Urodela | ||
| Batrachomorpha | Salientia | |||
| Batrachosauria | Chelonia | |||
| Eolacertilia | Lacertilia | |||
| Thecodontia | Crocodylia | Pterosauria | Dinosauria | Aves |
| Synapsida | Mamamlia |
Динозавры и птерозавры - единственные вымершие отряды среди вновь появившихся таксонов.
Таким образом, можно сказать, что поздняя юра - это время с которого начинается эволюция современной наземной биоты.
Поздняя юра (рис. 1.4).
Есть одно, но важное отличие между позднеюрской и позднетриасовой реконструкциями - отделение Южных континентов от Северных.
С этого времени мы знаем несколько групп млекопитающих: Kuehneotheriidae, Amphidontidae, Amphitheriidae и Peramuridae (см. Табл. 1). Все эти млекопитающие являются примитивными Theria, но не Metatheria или Eutheria.
Древнейшие Metatheria и Eutheria известны с конца раннего мела. Однако в коллекции ПИН РАН хранятся зубы Zalambdalestidae и Deltatheridiidae (Решетов, перс. сообщ.), найденные в Монголии в отложениях зоны Psittacosaurus. Все млекопитающие из этого местонахождения были опубликованы как апт-альбские. Однако фауна Psittacosaurus, как правило, относится к неокому.
А.К.Агаджанян, просматривая материалы, опубликованные из поздней юры Гумиаротта (Португалия), нашел, что Crusafontia и Henckelotherium (Dryolestia) напоминают Amblypoda и могут быть отнесены к этому отряду.
Во всяком случае, мы можем предположить, что юра, время разделения Северных и Южных континентов, должно быть временем начала дивергентной эволюции Сумчатых (Metatheria) на юге и Плацентарных (Eutheria) на севере.
Поздняя юра - ранний мел (начало) (рис. 1.5).
Некоторые ископаемые млекопитающие известны из С. Америки, Европы и Африки.
Здесь важно подчеркнуть, что в поздней юре (формация Моррисон) в С. Америке мы знаем Multituberculata. Все эти Multituberculata, очевидно, вымерли и новые мультитуберкуляты появились в С. Америке из Азии в конце раннего мела, в альбе. Этот факт очень важен, так как он поможет нам выяснить время контактов С. Америки с Азией и Ю. Америкой.
Мы покажем позднее, что С. Америка, в то время, когда она была изолирована от Азии в середине раннего мела, имела краткий контакт с Ю. Америкой.
Это был первый “Панамский” мост, которым воспользовались, с одной стороны, плацентарные, заселив Ю. Америку; с другой стороны, - дидельфиды, проникшие в С. Америку. Этот мост исчез прежде, чем возник первый “Берингийский” мост, связавший С. Америку с Азией.
Ранний мел (средняя часть) (рис. 1.6).
Плацентарные млекопитающие (некоторые группы Pantodonta, Condylarthra, Notoungulata, Xenarthra, Rodentia и Primates) достигли Ю. Америки. Сумчатые (Опоссумы) (Didelphidae) проникли в С. Америку, а затем - в Европу, С. Африку и Азию (Зайсан и Таиланд).
Очевидно, Австралия в это время уже была изолирована от Антарктиды и Ю. Америки. Мы полагаем, что если бы Австралия была соединена с Антарктидой и Ю. Америкой в это время, плацентарные млекопитающие должны были бы проникнуть из Ю. Америки через Антарктику в Австралию.
Таким образом, Multituberculata и плацентарные млекопитающие (за исключением Chiroptera) не будут найдены в Австралии до начала плиоцена, когда первые наземные плацентарные (Muridae) достигли Австралии (Archer, Clyton, 1984).
Много лет назад мы высказали предположение, что наземные тетраподы Антарктиды должны быть ближе к южноамериканским, чем к австралийским, так как, судя по современному положению континентов, Антарктида должна была изолироваться от Австралии раньше, чем от Ю. Америки. “Если связь между Ю. Америкой и Антарктидой уцелела до середины мела, в Антарктиде должны быть найдены все крупные таксоны древней фауны Ю. Америки” (Каландадзе, Раутиан, 1982).
Предположение подтвердилось. В настоящее время фауна наземных млекопитающих Антарктиды “включает два эндемичных вида полидолопид: Antarctodolops ... и Eurydolops ... и три плацентарных, определяемых как тардиградное неполнозубое, спарнотериодонтидный литоптерн, тригоностилопид и астрапотерий” (Hooker, 1992). В олигоцене Антарктиды был так же найден форорак.
Антарктида и Ю. Америка еще и сейчас связаны шельфом с цепью островов.
Как долго длилась “Блестящая изоляция” Южной Америки? Это - один из наиболее волнующих вопросов исторической зоогеографии.
Время изоляции Ю. Америки и проблема происхождения Caviomorpha и Platyrrhina уже обсуждалась нами, но мы должны вернуться к этой теме вновь.
Дж.Г.Симпсон суммировал все известные факты (Simpson, 1980). Он сделал вывод, что Ю. Америка была изолирована с середины мела по крайней мере до палеоцена и, может быть, до верхов (но не до конца) эоцена.
По Симпсону Caviomorpha и Platyrrhina проникли в Ю. Америку приблизительно на границе эоцена и олигоцена.
Мы взяли стратиграфическую схему распространения североамериканских и южноамериканских млекопитающих из работы Арчера и Клайтон (Archer, Clyton, 1984), продолжили ее до раннего мела и показали на ней основные события и группы млекопитающих (рис. 6). Рассмотрим полученный результат.
“Великий Американский обмен” - наиболее ясная часть этой истории (см. рис. 6). Специальная монография, посвященная этой теме, была опубликована недавно (Stehlin, Webb, 1985). На схеме показан весь временной интервал обмена и все семейства млекопитающих, которые прошли из Ю. Америки в Северную и наоборот в позднем миоцене - позднем плиоцене, плейстоцене и современности (рис. 7).
Стрелкой показан уровень, с которого первые древнейшие приматы и грызуны были найдены в Ю. Америке. Ниже приведен список этих первых южноамериканских приматов и грызунов.
Данные взяты из работы Симпсона (Simpson, 1980): ![]() Primates: Cebidae
Primates: Cebidae ![]() Rodentia: Cavioidea (Eocardiidae)
Rodentia: Cavioidea (Eocardiidae) ![]() Erethizontoidea (Erethizontidae)
Erethizontoidea (Erethizontidae) ![]() Chinchilloidea (Chinchillidae,
Dasyproctidae)
Chinchilloidea (Chinchillidae,
Dasyproctidae) ![]() Octodontoidea (Octodontidae, Echimyidae)
Следует подчеркнуть два факта:
Octodontoidea (Octodontidae, Echimyidae)
Следует подчеркнуть два факта:
- Приматы и грызуны появляются в Ю. Америке внезапно и, за исключением одного сомнительного семейства (Eocardiidae) все они - современные.
- Platyrrhina и Caviomorpha - абсолютные эндемики Ю. Америки вплоть до позднего миоцена, когда некоторые кавиоморфы достигли С. Америки.
Полагаем, что эти факты указывают, что не Platyrrhina и Caviomorpha, но их древние предки достигли Ю. Америки задолго до начала палеогена. И их раннеолигоценовые представители являются результатом долгой эволюции в изоляции.
Время их вторжения в Ю. Америку - один из труднейших вопросов в этой истории.
Три группы млекопитающих особо важны в этой истории:
- Eutheria. Когда они появились в Южной Америке?
- Metatheria. Когда они появились в Северной Америке?
- Multituberculata. Когда первые меловые Multituberculata появились в С. Америке и почему они не достигли Ю. Америки?
Первые Eutheria появились в Ю. Америке в позднем мелу (Vilquechican stage).
Первые Metatheria появились в С. Америке в позднем мелу (среднем сеномане).
Это значит, что “Первый Великий Американский обмен” начался в среднем сеномане или раньше. Вот почему ситуация с Multituberculata так важна. Multituberculata отсутствуют в Ю. Америке.
Первые меловые Multituberculata появились в С. Америке из Азии в альбе, около 100 млн. лет назад. Если это так, то мы знаем верхнюю границу окончания “Первого Великого Американского обмена”, потому что отсутствие в Ю. Америке типичных Multituberculata свидетельствует, что “Первый Великий Американский обмен” закончился до того, как они появились в С. Америке. В ином случае Multituberculata были бы найдены в фаунах Ю. Америки.
Но ситуация с южноамериканскими Multituberculata не так проста. Три или четыре южноамериканских рода в разное время определялись как Multituberculata: Gondwanatherium, Sudamerica, Ferugliotherium и (?) Vucetichia.
Доводом в пользу такого определения послужило “сходство общего строения эмали, микроструктура эмали, направления движения челюстей Gondwanatherium и ... мультитуберкулята Ferugliotherium (который, вероятно, включает Vucetichia), ... указывает, что Gondwanatherium и Sudamerica так же являются мультитуберкулятами. Gondwanatheria, включает два семейства - Ferugliotheriidae (Ferugliotherium) и Sudamericidae (Sudamerica и Gondwanatherium), поэтому признаются как новый высоко специализированный подотряд Multituberculata” (Krause, Bonaparte, 1990).
Все меловые мультитуберкуляты С. Америки являются абсолютно ординарными представителями отряда и близки к азиатским формам. Вот почему мы полагаем, что мультитуберкуляты действительно отсутствуют на южных континентах.
Выясняя время начала “Блестящей изоляции”, мы рассмотрели все таксоны млекопитающих Америк с раннего мела до современности (рис. 7) (Каландадзе, Раутиан, 1991). Две линии кругов показывают объем фаун млекопитающих Ю. Америки (внизу) и С. Америки (наверху) с раннего мела до современности. Наружные круги показывают число новых родов, а внутренние, черные - число новых семейств. Верхние цифры близ кружков показывают число новых семейств, нижние - новых родов в фаунах.
Полное число семейств и родов, которые не пересекли пролива между Южной и Северной Америкой (146 семейств и 1226 родов) показано на рисунке большими кругами.
Все таксоны от рода до отряда, с раннего мела до среднего миоцена включительно, известные из Ю. Америки и всех других территорий Земли указаны на рисунке внутри большого круга.
| South America | North America | Europe | Asia |
| Alphadon K2 | Alphadon K2 | ||
| Pediomyinae K2 | Pediomyinae P1 | ||
| Pantolambdinae K2 | Pantolambdinae P13-P21 | ||
| Mioclaeninae K2 | Mioclaeninae K2 | ||
| Periptychidae K2 | Periptychidae K2 | ||
| Didelphini P13-P21 | Didelphini P13-P21 | ||
| Notostylopoidea P13-P21 | Notostylopoidea P13-P21 | ||
| Glasbiinae P22-3 | Glasbiinae K2 | ||
| Tardigrada P22-3 | |||
| Tardigrada PP13-P21 | |||
| Primates P31 | Primates P13-P21 | ||
| Rodentia P31 | Rodentia P13-P21 | ||
| Myrmecophagidae N11 | Myrmecophagidae P22-3 |
И это весь список таксонов, общих для Ю. Америки и всем остальным миром за период около 100 000 000 лет. Полный список отрядов и семейств кайнозойских млекопитающих - с палеоцена до современности - известных из Ю. Америки и С. Америки так велик, что мы сделали специальный график, чтобы показать результаты этого анализа (рис. 8). Большая часть данных была взята из “Mammalian paleofaunas of the World” (Savage, Russell, 1983).
Только одно общее семейство было найдено в Южной и Северной Америке за время с палеоцена до среднего миоцена - Didelphidae. Но оно известно с обоих континентов по крайней мере с позднего мела (рис. 8).
Мы надеемся, что ситуация для кайнозоя прояснилась. 146 семейств и 1226 родов млекопитающих свидетельствуют о том, что Ю. Америка была изолирована по меньшей мере с верхов раннего мела до позднего миоцена.
Мы упоминали выше, что основной причиной дивергенции Metatheria и Eutheria была изоляция Южных и Северных континентов (рис. 1.4). Но смешанная метатериево-еутериевые фауны Южной и Северной Америк показывают, что эта изоляция прервалась. Мост возник в середине раннего мела (рис. 1.6) и сейчас мы знаем, когда он исчез. Мезозойская составляющая этой истории заслуживает тщательного исследования.
Для понимания мезозойской исторической зоогеографии наиболее важны не млекопитающие, а динозавры, так как наши знания об этих гигантских животных значительно более полны. Сравнение списка меловых динозавров Южной и Северной Америки показывает, что в этих фаунах есть только один общий род - Kritosaurus. Большая часть гадрозавров известна из позднего мела, но некоторые остатки были найдены и в раннем мелу. Следует отметить, что Kritosaurus - один из наиболее примитивных Hadrosauridae (Weishampel, Dobson, Osmolska, 1990).
Все динозавры Ю. Америки очень архаичны, особенно карнозавры. Большую часть из них относят к семейству Abelisauridae и выглядит они как специализированные Megalosauridae.
Поэтому мы полагаем, что контакт между Америками был в середине раннего мела (рис. 1.6). И с этого момента, но в любом случае до альба, началась “Блестящая изоляция” Ю. Америки.
Одним из наиболее впечатляющих результатов последних лет было открытие меловых млекопитающих Ю. Америки. Бонапарт писал: “Фауна Лос Аламитос показывает большое отличие от всех других позднемеловых комплексов млекопитающих” (Bonaparte, 1990, p. 63).
Бонапарт полагал, что “Gondwanatheria могут быть предками
Xenarthra”. В этом мы совершенно согласны с ним. Общая
структура отряда может выглядеть таким образом:
Order
Xenarthra ![]() Suborder Gondwanatheria (Gondwanatherium,
Sudamerica);
Suborder Gondwanatheria (Gondwanatherium,
Sudamerica); ![]() Suborder Cingulata;
Suborder Cingulata; ![]() Suborder Pilosa
Suborder Pilosa
Ferugliotherium и Vucetichia, определенные как Gongwanatheria - брахиодонтные формы. Они являются самыми интересными зверями фауны Лос Аламитос (Bonaparte, 1986; Krause, Bonaparte, 1990; Krause, Kielan-Jaworowsca, Bonaparte, 1992).
Звери фауны Лос Аламитос очень разнообразны. Не время и не место разбирать разные точки зрения, высказанные относительно этих зверей. Однако, А.Агаджанян (перс. сообщ.) полагает, и мы согласны с ним, что настало время поисков меловых грызунов. И звери фауны Лос Аламитос, особенно такие как Barberina, должны быть изучены с особой тщательностью. Прежде всего потому, что меловые Condylarthra, Pantodonta и Notoungulata впервые были найдены в Южной Америке, - признанном рефугиуме самых разных животных.
С раннего мела (или раньше, но не позже) Австралия была изолирована вплоть до раннего плиоцена. Это объясняет, почему все динозавры Австралии принадлежат к триасовым или юрским группам. Мы не знаем ни одного рода динозавров, близко родственного хорошо известным динозаврами С. Америки и Азии: Hadrosauridae, Tyrannosauridae, Protoceratopsidae, Ceratopsidae, Pachycephalosauridae, Psittacosauridae и другие. По мнению Т.Рич и П.Виккерс-Рич "Раннемеловая фауна позвоночных Виктории необычна тем, что в ней доминируют мелкие орнитоподы. Нигде в мире более нет такого, за исключением, быть может, позднего триаса - ранней юры Южной Африки. [“The Early Cretaceous vertebrate faunas of Victoria are unusual in being dominated by small ornithopod dinosaurs. Nowere else in the world is this the case, exept perhaps in the Late Triassic - Early Jurassic of Southern Africa”] (Rich, Vickers-Rich, 1989, p. 16).
Полный список австралийских динозавров очень интересен.
Единственный юрский динозавр, Rhoetosaurus (Cetiosauridae?), был найден в среднеюрских отложениях. На современной реконструкции его стопы показано четыре когтя. Все известные нам Sauropoda имеют три когтя. Если реконструкция верна, это должно обозначать, что Rhoetosaurus является наиболее примитивным из всех известных Sauropoda. Sauropoda известны начиная c ранней юры, а Prosauropoda - с позднего триаса.
Все остальные динозавры Австралии происходят из раннего мела (J.Long, 1993).
Austrosaurus (Cetiosauridae ?). "... напоминает среднеюрских динозавров" (p.55).
Brachiosauridae ? (Indet.) "... очень похож на ... Brachiosaurus"из поздней юры.
Allosaurus sp. (Allosauridae). "Allosaurus известен из С. Америки, вымер в конце юрского периода. Это значит, что Allosaurus из Виктории был “живым ископаемым”" (p. 57).
Raptor (Abelisauridae ?). "... очень сходен с целюрозавром Ornitholestes" (p.58). Ornitholestes известен из поздней юры С. Америки.
Kakuru (Coeluridae ?). Североамериканский род Coelurus известен из поздней юры.
Walgettosuchus. Molnar (1982) :"... возможно, принадлежит к одному из трех семейств теропод (Allosauridae, Coeluridae and Ornithomimidae". Без комментариев.
Hypsilophodontidae. Смотри выше цитату из работы Т.Рич и П.Виккерс-Рич (Rich, Vickers-Rich, 1989, p. 16).
Muttaburrasaurus (Iguanodontidae). "Ближайший родственник ... Camptosaurus из поздней юры Европы и Северной Америки" (p. 63).
Minmi (Ankylosauridae). Очень странный Ankylosauridae. Древнейший анкилозавр известен с неокома.
Очевидно, что в мелу Австралия была рефугиумом для наиболее древних - триасовых и юрских - групп динозавров.
Присутствие “последнего в мире лабиринтодонта” (Rich, Vickers-Rich, 1989, р. 16) подтверждает то, что Австралия была изолирована с начала юры. Но из-за того, что это было время “Юрской биоценотической катастрофы” (рис. 4, 5; табл. 1), недостаточно материала для зоогеографического анализа. Даже в поздней юре практически нет наземных тетрапод на южных континентах (рис. 1.4-1.5).
Мы уже отмечали, что маммальные фауны южных континентов исходно содержат древние, примитивные группы млекопитающих: только Prototheria и Metatheria. Австралия изолировалась до того, как Южная и Северная Америка соединились в середине раннего мела (рис. 1.6). Следовательно, Eutheria и Multituberculata не могли достичь Австралии и поэтому Австралия имеет уникальную фауну млекопитающих, состоящую исходно только из Prototheria и Metatheria.
Так как грызуны (Muridae) достигли Австралии лишь в раннем плиоцене, этот континент имеет наиболее полно сохранившуюся фауну млекопитающих, которая начала свою эволюцию в юре. И сейчас, изучая современную фауну, мы можем видеть результаты этой эволюции.
Именно поэтому возможно проследить современные семейства австралийских млекопитающих до древнейших известных из Австралии фаун - миоцена и олигоцена (Archer, Hand, Godthelp, 1994). И первое раннемеловое млекопитающее Австралии - Steropodon - принадлежит к современному семейству, - Ornithorhynchidae (Archer, Flannery, Ritchie, Molnar, 1985). То же самое следует ожидать и для современных семейств Marsupialia.
В 1994 году была найдена вторая опаловая челюсть раннемелового млекопитающего. Она была описана как новый род Monotremata - Kallicodon. А.Агаджанян уверен, что челюсть принадлежит новому высоко специализированному роду семейства Phalangeridae. Существует известное структурное сходство австралийской и мезозойских фаун. В Австралии нет хороших маммальных хищников. Наиболее крупным был Thylacinus. Еще в плейстоцене был хищный дипротодонтоид, - Thylacoleo. Крупнейшими же хищниками были: варан Megalania, питон Wonambi и наземные крокодилы (Hecht, Archer, 1977; Rich, Monaghan, Baird, Rich, 1991). Таким образом, общая структура современной наземной фауны Австралии выглядит чрезвычайно архаично.
Итак, мы действительно можем ожидать, что некоторые группы современных млекопитающих будут прослежены до мелового времени, как это уже произошло с линией Ornithorhynchus - Obdurodon - Steropodon. Monotrematum, близкий к Obdurodon, найденный в палеоцене Южной Америки, подтверждает, что это действительно возможно.
В настоящее время Австралия - наиболее интересный для палеонтологов континент. Особо важно найти здесь палеогеновых и мезозойских млекопитающих, так же как и новые материалы по динозаврам, особенно позднемеловым. Конечно, весьма важно найти так же терапсид, но австралийские терапсиды будут более или менее близки южноафриканским, российским и другим формам Гондванской зоогеографической области.
Обратившись вновь к рис. 1, мы можем вновь рассмотреть основные отличия между Южными и Северными континентами.
После позднего триаса (рис. 1.3) Ю. Америка имела короткий контакт с С. Америкой в раннем мелу. И потом она была изолирована от северных континентов до позднего миоцена.
Австралия была изолирована по крайней мере с конца раннего мела до раннего плиоцена. Начиная с плиоцена в Австралию проникли: Muridae, Homo и Canis (рис. 1.1-15).
Африка была изолирована большую часть мелового периода (рис. 1.6-8). В палеоцене Африка соединилась с Европой. Ее контакт с Евразией существует до сих пор.
В середине раннего мела (рис. 1.6) С. Америка соединилась с Ю. Америкой, а затем, когда этот контакт уже прервался - с Азией (рис. 1.7).
В конце позднего мела и начале палеоцена все континенты были изолированы (рис. 1.8). Но после этого времени все северные континенты так или иначе соединялись вновь, а с юга к ним присоединились Африка и Индия.
В кайнозое все маммальные фауны Северных континентов так или иначе перемешивались. И основным результатом каждого контакта и смешения фаун было вымирание и ускорение дивергентной эволюции.
Если мы сравним списки палеоценовых млекопитающих в Южной и Северной Америке, мы можем увидеть весьма важный факт. В Ю. Америке известно 16 семейств палеоценовых млекопитающих. Половина из них вымерла в эоцене и олигоцене; другие же - после начала “Великого Американского обмена”: Didolodontidae (поздний миоцен); Interatheriidae (плиоцен); Borhyaenidae, Glyptodontidae, Protherotheriidae, Macraucheniidae (плейстоцен); Didelphidae и Dasypodidae дожили до современности. Это значит что шесть (6!) семейств палеоценовых млекопитающих Ю. Америки вымерло во время вторжения млекопитающих из С. Америки в Южную.
Абсолютно другая ситуация - в С. Америке. Здесь известно 44 семейства палеоценовых млекопитающих. Лишь четыре из них дожили до “Великого Американского обмена”. Два вымерло в плейстоцене (Equidae and Erinaceidae). Didelphidae и Soricidae - дожили до современности.
И такова ситуация на всех северных континентах.
Ранний - поздний мел (рис. 1.7).
Первый прямой контакт между С. Америкой и Азией - “Первый Берингийский мост”. По крайней мере с этого времени Мадагаскар изолирован от Африки. Таким образом, современная фауна наземных животных Мадагаскара является сохранившейся до нашего времени частью древней, мезозойской, фауны Африки.
Прежде мы уже отмечали, что Мадагаскар изолирован с раннего мела (Каландадзе, Раутиан, 1983). Здесь уместно упомянуть рисунок из работы Кука (Cooke, 1972), на котором морские отложения показаны вдоль обоих берегов Макассарского пролива с юры до миоцена. И поставлен только один вопрос - в палеоцене.
На Мадагаскаре мы не знаем маммальных фаун древнее плейстоцена. Но мы знаем современную и плейстоценовую фауну Мадагаскара и знаем фауны млекопитающих Африки начиная с палеоцена.
Сэведж и Расселл (Savage, Russell, 1983) опубликовали первый список палеоценовых млекопитающих Африки: Cimolesta (Palaeoryctidae), Creodonta (Hyaenodontidae) и Carnivora (Miacidae ?). С палеоцена (рис. 1.9) Африка соединена с Европой, а позднее - с Евразией. С этого времени значительная часть древних африканских млекопитающих начала вытесняться евразиатами.
Во всяком случае на Мадагаскаре неизвестны такие типичные для Африки млекопитающие, как Perissodactyla, Artiodactyla, Carnivora (кроме Viverridae), Catarrhina, Hyracoidea, Proboscidea, и многие другие.
Таким образом, мы действительно можем сказать, что Мадагаскар сохранил для нас некоторые группы древних, мезозойских, млекопитающих Африки.
Поздний мел - средний палеоцен (рис. 1.8).
Максимальная трансгрессия. Только Ю. Америка и Антарктида, быть может, соединены. Все другие континенты, а так же Мадагаскар и Новая Зеландия изолированы.
Это означает, что динозавры вымирали и млекопитающие начинали свою кайнозойскую эволюцию по крайней мере на девяти (!) изолированных массивах земли.
Поздний палеоцен - ранний эоцен (рис. 1.9).
Индия соединилась с Азией, Африка - с Европой. Азия не имеет прямого контакта с Европой.
Средний - поздний эоцен (рис. 1.10).
Северная Атлантика соединилась с Северным Ледовитым океаном, отделив С. Америку от Европы. Индия вошла в контакт с Азией и началось образование Гималаев, которые растут и сейчас. Продолжает существовать Берингийский мост.
Это означает, что все таксоны, которые появились после разъединения С. Америки и Европы, но отсутствуют в Азии, либо должны быть древнее среднего эоцена, либо являются парафилетичными.
Engesser (1979), изучив некоторые такие таксоны, получил великолепный результат. Выяснилось, что ряд групп млекопитающих по обе стороны Атлантики претерпели удивительное параллельное развитие.
Polydolopidae, Xenarthra, Litopterna, и Astrapotheria подтверждают, что фауна Антарктиды была ближе к южноамериканской, а не Австралийской фауне. Однако, это не значит, что Антарктида была в контакте с Ю. Америкой: маммального материала недостаточно. Гляциальные и интергляциальные отложения прослеживаются в Антарктиде до среднего (?) - позднего эоцена (Webb, Harwood, 1993).
Ранний олигоцен (рис. 1.11).
Европа и Азия объединились вновь впервые с позднего триаса и остаются соединенными до сих пор.
Средний - поздний олигоцен (рис. 1.12).
Единственное время со среднего палеоцена, когда мы можем показать разрыв между С. Америкой и Азией.
Ранний - средний миоцен (рис. 1.13).
Берингийский мост возникает вновь и существует до плейстоцена включительно.
Поздний миоцен (рис. 1.14).
Существует Берингийский мост, которым воспользовались некоторые млекопитающие: мастодонты, ...
Впервые после раннего мела восстановился контакт Северной и Южной Америки – возник Панамский мост, существующий и сейчас.
Начало “Великого Американского обмена”.
Разные группы млекопитающих проходили по мосту: мастодонты, кошки, включая саблезубых, собаки, медведи, еноты – с севера на юг; наземные ленивцы, броненосцы, глиптодонты, олени, верблюды (ламы), ...
Одним из наиболее важных фактов, подтверждающий, что Панамский пролив невозможно было пересечь наземному животному: уровень Атлантического океана на 15-20 метров выше уровня Тихого океана. Это значит, что чем уже был пролив, тем сильнее в нем было течение.
Ранний плиоцен - плейстоцен (рис. 1.15).
Подвижки: Африка, северная Америка и Южная Америка немного сдвигаются на юг. В Америке это видно по форме Калифорнийского залива и морфологии дуги Южных Антильских островов. Сдвиг Африки образовал Средиземное море. Мадагаскар стоит на месте и так же свидетельствует о сдвиге Африки на юг приблизительно на 500 км.
В плиоцене и плейстоцене по широкому Берингийскому мосту – далеко на север от нынешнего пролива до Алеутских островов на юге – продолжался не столько обмен, сколько заселение Северной Америки азиатами.
Собственно, Америка подарила Старому Свету, фактически, только верблюдов.
В то время как слоны (разные формы мастодонтов и мамонты), олени, бизоны, овцебыки, бараны, козы, яки, сайгаки, ... А также львы, гепарды, волки, рыси, медведи. И, наконец, по крайней мере, много раньше 30 тысяч лет назад, в Северную Америку прошел человек. Заселенные человеком пещеры Южной Америки датируются 40 - 45 000 лет. Из чего следует, что в Северной Америка он появился не позже 50 000 лет назад.
В течение этого времени Австралия по крайней мере дважды имела краткие контакты с Азией.
Мост этот существовал несколько кратких (геологически кратких) мгновений. Сначала (в плиоцене) его прошли мыши и крысы (но не белки, которые буквально царствуют в лесах Индокитая). Скорее всего это значит, что на мосту не успел вырасти ни лес, ни даже кустарник.
А в плейстоцене, около 120 тысяч лет назад, его пересек человек, а потом и собака.
Только эти плацентарные млекопитающие проникли в Австралию сами (кроме летучих мышей, конечно). Всех остальных завез, вольно или невольно, человек.
Мыши и крысы (Muridae) в плиоцене, а затем – человек (Homo) и собака (Canis) проникли в Австралию в течение плейстоцена.
Многие ученые Австралии полагают, что аборигены достигли Австралии 40 - 45 000 лет назад. Однако, Сингх, Кершан и Кларк (1979) и Сингх (1982) писали о резком возрастании количества пеплов в отложения озера Джорж с уровня, определяемого возрастом 120 000 лет. Они полагали, что этот факт свидетельствует о прибытии человека на континент см. Арчер (Archer, 1984). Недавняя находка древнейших петроглифов (60 000 лет) и орудий (более 116 тысяч лет) показала, что аборигены действительно достигли Австралии около 120 тысяч лет назад (S.Menon, 1997).
Ледниковый щит Антарктиды не был сплошным еще в позднем плиоцене, так как датируемые этим возрастом остатки нотофаговых лесов были найдены в Трансантарктических горах, в 500 км от Южного полюса (Webb, Harwood, 1993).
Некоторые таксономические результаты.
Мы не знаем типичных меловых динозавров в Ю. Америке до вторжения плацентарных млекопитающих, вместе с которыми некоторые меловые североамериканские динозавры проникли в Ю. Америку.
Отсутствие в Австралии наземных тетрапод, характерных только для позднего мела, подтверждает это заключение. Это значит, что Северные и Южные континенты были разобщены с поздней юры или ранее.
Австралийские млекопитающие могли проникнуть в Австралию только из Ю. Америки через Антарктиду до поздней юры. Следовательно, сумчатые должны были достичь Австралии до конца юры.
Есть две группы неэндемичных млекопитающих в Австралии: Prototheria и Metatheria. Таким образом, эти млекопитающие должны были проникнуть в Ю. Америку не позднее средней юры.
Если мы полагаем, что Австралия была изолирована от Ю. Америки с Антарктикой позже раннего мела, Prototheria, и Metatheria должны были существовать в Австралии, Южной Америке и Антарктиде до этого времени.
Ю. Америка была изолирована от С. Америки с конца раннего мела до среднего миоцена. Это значит, что все таксоны, известные из Ю. Америки и всех других территорий в течение всего времени изоляции, должны быть, по крайней мере, раннемелового возраста. Эти таксоны: Alphadon; Didelphini; Glasbiinae; Pediomyinae (Didelphidae); Pantolambdinae (Coryphodontidae); Tarsiiformes (ancestor of Platyrrhina); Rodentia; Myrmecophagidae; Tardigrada; Mioclaeninae (Hyopsodontidae); Periptychidae; Notoungulata.
С. Америка была изолирована в позднем мелу. В это время мы знаем Plesiadapiformes в С. Америке. Lemuriformes многочисленны на Мадагаскаре. Следовательно, они должны были быть и в Африке. Primates, таким образом, должны были существовать в раннем мелу в Африке, Европе и С. Америке.
Мадагаскар был изолирован от Африки по крайней мере в мелу. Следовательно, все млекопитающие, известные из Мадагаскара и из Африки, должны были существовать на Мадагаскаре и в Африке по крайней мере с мела. Эти таксоны: Geogalinae (Tenrecidae); Hemigalinae, Haepestinae (Viverridae); Lemuriformes; Cricetidae; Tubulidentata.
Со второй половины позднего мела до среднего палеоцена Азия была изолирована от С. Америки. Значит, все таксоны, известные из Азии и С. Америки в это время, должны быть на обоих континентах не позднее, чем начало позднего мела. Эти таксоны: Coryphodontidae; Hyopsodontidae; Periptychidae; Mesonychidae.
Благодарности
Я весьма обязан многим друзьям и коллегам:
Prof.
Patricia Vickers - Rich, Dr. Thomas H.V. Rich, Dr. Alex
Ritchie, Dr.Timothy F.Flannery, Linda Gibbson, Tish Ennis,
Prof. Michael Archer, Prof. Colin Groves, Zerina Johanson,
Галине Сергеевне Раутиан, Jennifer Klein и особо - другу и
коллеге - А.К.Агаджаняну.
Литература
Archer M., 1984. Effects of Human on the Australian Vertebrate fauna. p.151-161. In: Archer M., Clyton G., 1984. (Ed.) Vertebrate zoogeography and Evolution in Australasia. Hasperian Press, 1203 p.
Archer M., Clyton G., 1984.(Ed.) Vertebrate zoogeography and Evolution in Australasia. (Animals in Space and Time). Hesperin Press, 1203 p.
Archer M., Flannery T.F., Ritchie A.,Molnar R.E., 1985. First Mesozoic mammal from Australia - an Early Cretaceous monotreme. Nature, 318: 363-366.
Archer M., Hand S.J., Godthelp H., 1994. Riversleigh. The Story of Animals in Ancient Reinforest of Inland Australia. Reed, 264 p.
Benton M.J., 1985. Mass Extinction among families of non tetrapods. Nature, 306: 811-814.
Benton M.J., 1987. Mass extinction among families of non marine tetrapods:the data. Mem. Soc. geol. France, No 150: 21-32.
Benton M.J., 1988. Mass extinction on the fossil record of reptiles paraphyly,patchness and periodicity. Extinction and survival in the fossil record. Oxford: Clarendon press.(Syst. Assoc. Spec., No 34) : 269-294.
Benton M.J., 1989. Patterns of evolution and extinction in vertebrates. Evolution and the fossil record. L.: Belhaven press: 218-241.
Bonaparte G.F., 1986. A new and unusual Late Cretaceous mammal from Patagonia. J. Vert. Paleont. 6: 264-270.
Bonaparte J.F., 1990. New Late Cretaceous mammals from the Los Alamitos formation, Northern Patagonia. Nat. Geograph. Research, 6 (1): 63-93.
Bond M., Pascual R., Reguero M., Santillana S., Marenssi S., 1989. Los primeros "ungulados" extinguidos sudamericanos de la Antarctida. Ameghiniana, 26: 240.
Case J.A., Woodburne M.O., Chaney D.S., 1988. A new genus of polydolopid marsupial from Antarctica. Mem. Geol. Soc. America. 169: 505-521.
Cooke H.B.S., 1972. The fossil mammal fauna of Africa. In: Keast A., Erk F.C., Glass B.(Ed.),Evolution of Mammals, and Southern Continents. Univ. New York Press, Albany, 543 p. Engesser B., 1979. Relationships of some insectivores and rodents from the Miocene of North America and Europe. Pittsburgh: Bull. Carnegie Mus. N. H. 14: 68 p.
Hecht M., Archer M.,1977. Presence of xiphodont (sic.) crocodylians in the Tertiary and Pleistocene of Australia. Archeringa 1: 383-385.
Hooker J.J., 1992. An additional record of a placental mammal (Order Astrapotheria) from the Eocene of West Antarctica. Antarctic Sci., 4: 107-108.
Ивахнкенко М.Ф., 1987. Пермские парарептилии СССР. Тр. Палеонтол. ин-та АН СССР, т. 223, изд-во “Наука”, М., 160 с.
Каландадзе Н.Н., Раутиан А.С., 1980. Историческая зоогеография наземных тетрапод конца палеозоя - начала мезозоя. Палеонтология. Стратиграфия., Мю, с. 93-102.
Каландадзе Н.Н., Раутиан А.С., 1981. Межконтинентальные связи наземных тетрапод и решение проблемы шотландской фауны Элгин. Тр. 23-й сессии ВПО. Л., с. 124-133.
Каландадзе Н.Н., Раутиан А.С., 1981. Историч. зоогеогр. поздн. триаса.......
Каландадзе Н.Н., Раутиан А.С., 1982. Историческая зоогеография млекопитающих. В: сб. Млекопитающие СССР. Реф. сообщ. на 3 Всесоюзной териологическом совещании. “Наука”, М., с. 38.
Каландадзе Н.Н., Раутиан А.С., 1983. Место Центральной Азии в зоогеографической истории мезозоя. Тр. ССМПЭ. Вып. 24. Ископаемые рептилии Монголии. М., с. 6-44.
Каландадзе Н.Н., Раутиан А.С., 1989. Историческая зоогеография млекопитающих. В: Сб. реф. 1-го Всесоюзной конференции палеонтологов, М., с. 28-30.
Каландадзе Н.Н., Раутиан А.С., 1991.Система млекопитающих и их историческая зоогеография. В: Сб. Териология. “Наука”, М., с. 132-164.
Kalandadze N.N., Rautian A.S., 1991 b. Late Triassic zoogeography and reconstruction of the terrestrial tetrapod fauna of North Africa. Paleont. Journ. 1: 1-12.
Kalandadze N.N., Rautian A.S., 1993. Jurassic ecological crisis of terrestrial tetrapoda society and euristic model of co-evolution society and biota. In: Problems of Pre antropogenic evolution of the Biosphere. Nauka, Moscow, p. 60-95.
Krause D.W., Bonaparte J.F., 1990. The Gondwanatheria, a new suborder of Multituberculata from South America. Journ. Vert. Paleont., 10 (3), p. 31 A.
Krause D.W., Kielan-Jaworowska Z., Bonaparte J.F., 1992. Ferugliotherium Bonaparte ,the first known multituberculata from South America. J. Vert. Paleontol., 12 (3): 351-376.
Long J., 1993. Dinosaurs of Australia and other animals of the Triassic, Jurassic and Cretaceous period. Reed., 87 p.
Marchall L.G., de Muizon C., 1988. The Dawn of the Age of Mammals in South America. Nat. Geogr. Research, 4 (1): 23-55.
Menon S. 1997. Art in Australia, 60. 000 Years Ago. Discover, Spec. Issue: The Year in Science. The World of Science. January, 1997, p. 33-34.
Reisz R. 1996. The Early Permian Reptile Acleistorhinus pteroticus and its phylogenetic position. J. Vert. Paleontol. 16 (3), p. 384-395.
Rich P.V., Monaghan J.M., Baird R.F., Rich T.H.V., (Ed.), 1991. Vertebrate Palaeontology of Australasia. Monash Univ. Publ. Comm., Melbourne, 1437 p.
Rich T.H.V., Rich P.V., 1989. Polar dinosaurs and Biotas of the Early Cretaceous Southestern Australia. Nat. Geograph. Research 5 (1): 15-53.
Rich, T. H., Vickers-Rich P., 1994. Neoceratopsians and Ornithomimosaurs: Dinosaurs of Gondwana Origin? Research and Exploration 10: 129-131.
Simpson G.G., 1980. Splendid Isolation. The curious history of South American Mammals. New Haven, London, Yale Univ. Press. 266 p.
Sinch G., 1982. Environmental upheaval. p. 90-108. In: Smith J.M.B., A history of Australasian vegetation. McGraw Hill Book Co.: Sydney.
Singh G., Kershaw A.P., Clark R., 1979. Quarternary vegetation and fire history in Australia. Ch. 1. In: Gill A.M., Growes R.A., Noble I.R. (Ed.). Fire and Australian biota. Australian Acad. Sci., Canberra.
Stehlin F.G., Webb S.D.(Ed.), 1985. The Great American Biotic Interchange. Plenum Press, New York. 520 p.
Webb P.N., Harwood D.M., 1993. Pliocene fossil Nothofagus (Southern beech) from Antarctica : Phytogeography, dispersal strategies, and survival in high latitude glacial-deglacial environments. In: Forest Development in Cold Climates, Alden J. et al. (Ed.), Plenum Press, New York, p. 135-165.
Weishampel D.B., Dobson P., Osmolska H. (Ed.), 1990. The Dinosauria. Univ. Calif. Press, 733 p.
White M.E., 1994. The Greening of Gondwana. The 40 million year story of Australia's Plants. Singapure, 256 p.
Woodburne M.O., Zinsmeister W.J., 1984. The first land mammal from Amtarctica and its biogeographic implication. Journ. Paleont. 58: 913-948.
Рисунки
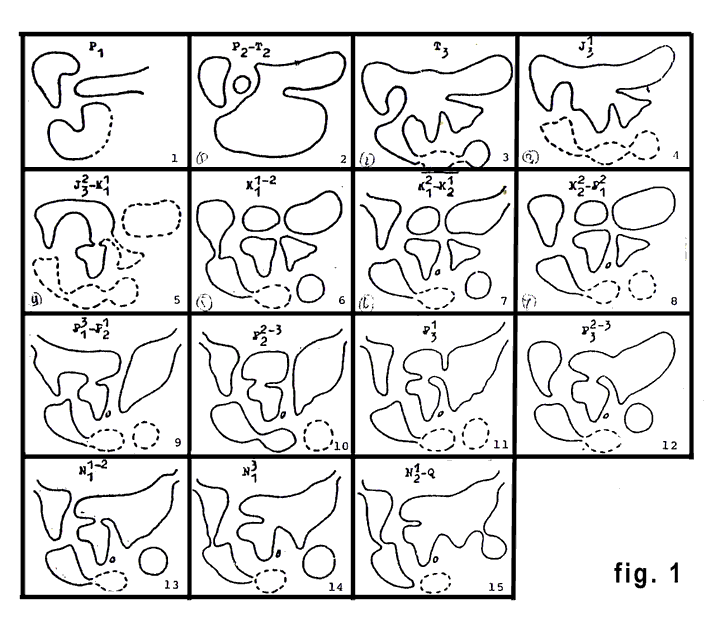
1. Серия зоогеографических реконструкций с ранней перми до плейстоцена включительно, сделанная на основе анализа строго наземных тетрапод. (Каландадзе Н.Н., Раутиан А.С., 1991, уточненная).
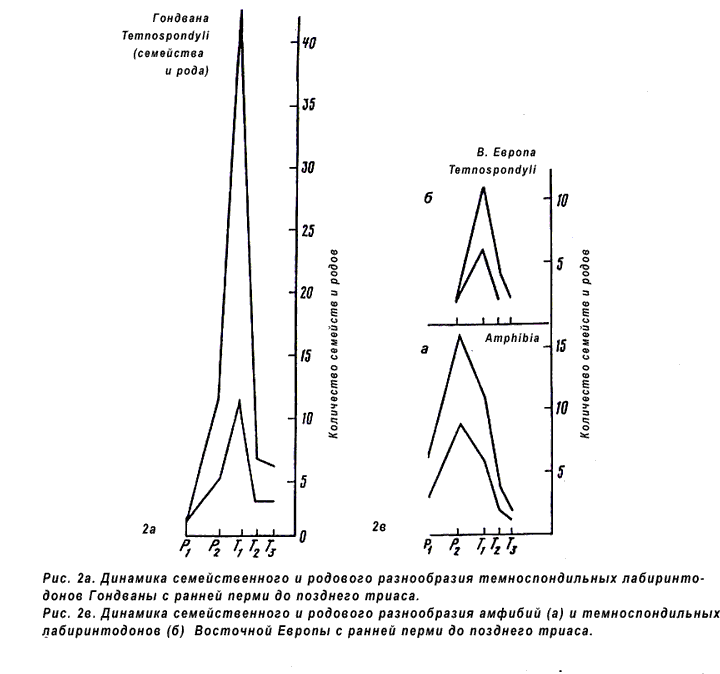
2 а. Динамика семейственного и родового разнообразия
темноспондильных лабиринтодонтов Гондваны с ранней перми до
позднего триаса (Каландадзе, Раутиан, 1991).
2 б. Динамика
семейственного и родового разнообразия амфибий (б) и
темноспондильных лабиринтодонтов Восточной Европы с ранней
перми до позднего триаса (Каландадзе, Раутиан, 1991).

3 а. Динамика семейственного и родового разнообразия
амфибий Северной Америки с карбона до позднего триаса
(Каландадзе, Раутиан, 1991).
3 б. Динамика семейственного и
родового разнообразия амфибий Западной Европы с карбона до
позднего триаса (Каландадзе, Раутиан, 1991).
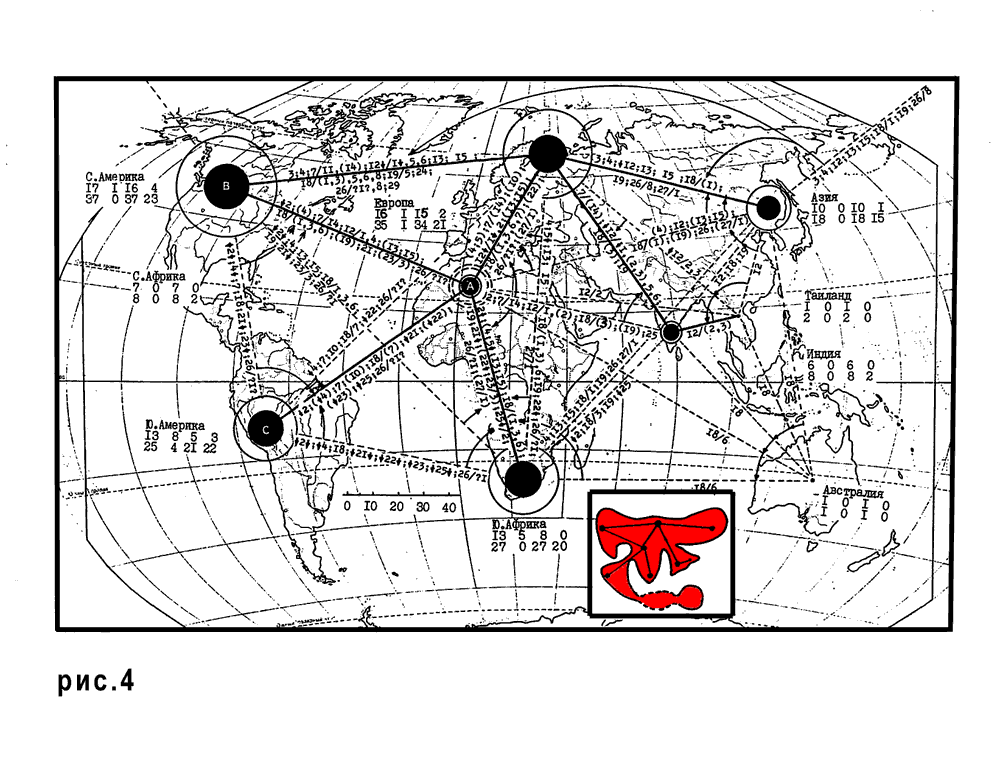
4. Реконструкция фаунистических связей наземных тетрапод и зоогеографическая картина позднего триаса. (Каландадзе Н.Н., Раутиан А.С., 1991 б).

5. Реконструкция фаунистических связей наземных тетрапод и зоогеографическая картина в ранней (5a) и средней (5b) юре (Каландадзе, Раутиан, 1983).
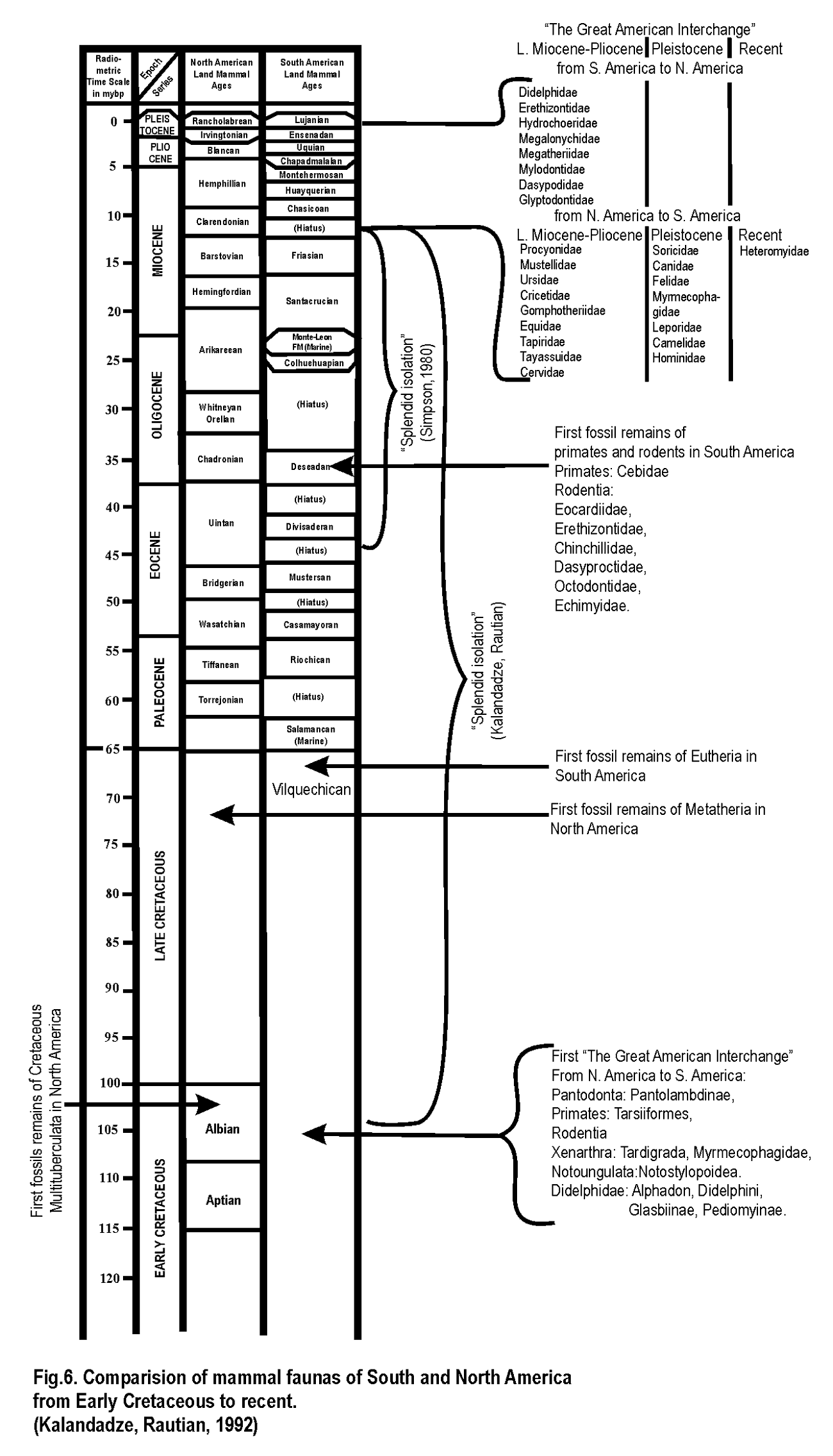
6. Рисунок, показывающий время предполагаемого Первого “Великого Американского обмена”(ранний мел) и “Великого Американского обмена” (поздний миоцен - современность) - “Блестящая изоляция” по Симпсону (Simpson,1980) и авторам. Пояснения в тексте.
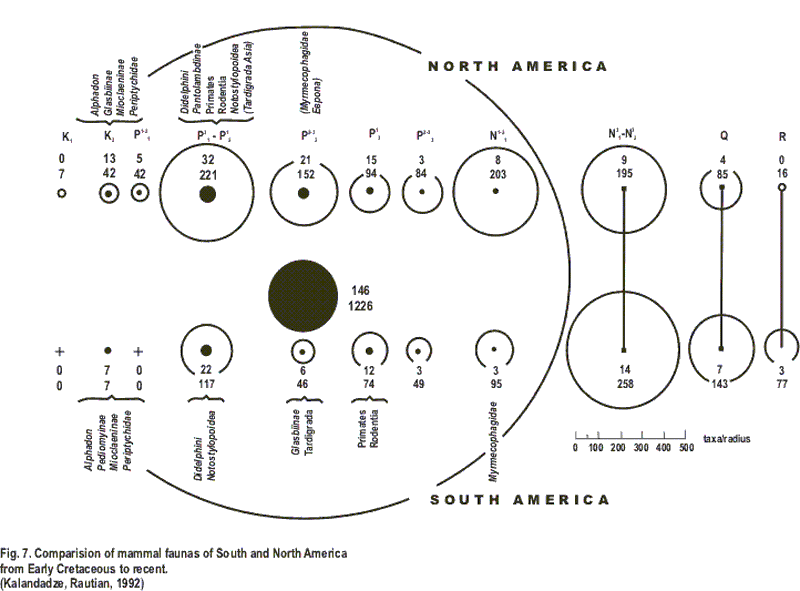
7. Сравнение фаун млекопитающих Южной и Северной Америки с раннего мела до современности. (Каландадзе, Раутиан, 1991, уточненно).
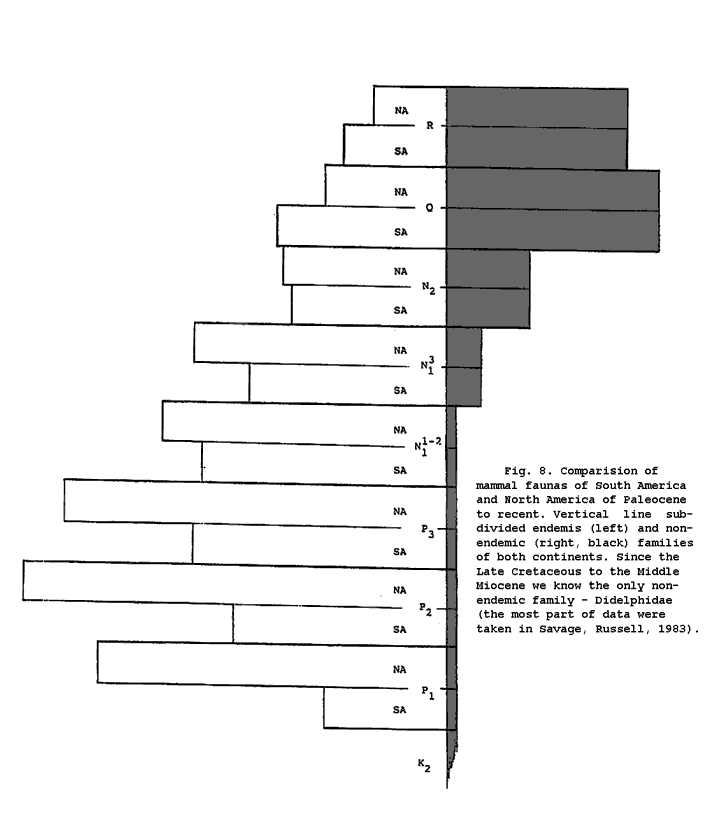
8. Сравнение фаун млекопитающих Южной и Северной Америки с палеоцена до современности. Вертикальная линия подразделяет эндемичные (слева) и не эндемичные (справа, зачернено) семейства обоих континентов. С позднего мела до среднего миоцена мы знаем только одно не эндемичное семейство - Didelphidae. Данные взяты в основном из работы Сэваджа и Расселла (Savage and Russell, 1983).