ГЛАВА 11. СИСТЕМАТИКА ПРОКАРИОТ. ГРУППЫ ПРОКАРИОТНЫХ ОРГАНИЗМОВ
Систематика (таксономия) — наука о многообразии и взаимосвязях между организмами. Одна из задач систематики — распределение (классификация) множества организмов по группам (таксонам)27. Но прежде чем осуществлять такое распределение, необходимо достаточно полно охарактеризовать объекты и на основании отобранной информации идентифицировать их. Последнее может привести к выявлению организмов с неизвестными или известными признаками и соответственно помещению их в новый таксон на определенном уровне или же отнесению к известным таксонам.
27 Таксон — группа организмов, обладающих заданной степенью однородности.
Для характеристики организмов используют разнообразные признаки: морфологические, цитологические, культуральные, физиологические, биохимические, иммунологические и др. Если объем информации для характеристики объектов по существу беспределен, как бесконечен сам процесс познания природы, то для целей идентификации может быть использован ограниченный объем информации, достаточный для распределения организмов по таксономическим группам.
Специальный раздел таксономии — номенклатура — имеет дело с правилами присвоения наименований описанным объектам. В систематике бактерий для наименования объекта используют биномиальную номенклатуру К. Линнея (К. Linne, 1707—1778), согласно которой биологическому виду присваивают название, состоящее из двух слов: первое определяет принадлежность организма к определенному роду, второе — виду. Названия бактериям присваивают в соответствии с правилами Международного кодекса номенклатуры бактерий.
Основной таксономической категорией является вид. По современным представлениям, вид — это группа близких между собой организмов, имеющих общий корень происхождения и на данном этапе эволюции характеризующихся определенными морфологическими, биохимическими и физиологическими признаками, обособленных отбором от других видов и приспособленных к определенной среде обитания.
Важным признаком, определяющим принадлежность организмов к одному виду, является их способность скрещиваться и давать жизнеспособное потомство. Однако у прокариот размножение половым путем отсутствует, поэтому данный признак для определения видовой принадлежности к ним неприменим. Отнесение прокариотных организмов к одному или разным видам осуществляется в большой степени эмпирическим путем на основе анализа многих признаков, при этом генетическая информация, содержащаяся в нехромосомных генетических элементах, для определения видовой принадлежности не используется.
Виды объединяют в таксоны более высокого порядка — роды, роды — в семейства, далее следуют порядки, классы, отделы, царства. Для высших таксономических категорий пока нет удовлетворительного определения.
В микробиологии употребляются такие термины, как "штамм" и "клон". Под штаммом понимают бактериальные культуры одного вида, выделенные, например, из разных мест обитания. Различия между штаммами не выходят за пределы вида. Клон — еще более узкое понятие, это культура, выделенная из одной клетки.
Существуют 2 типа систематики биологических объектов: филогенетическая, или естественная, в основе которой лежит установление родственных (генетических, эволюционных) связей между организмами, и практическая, или искусственная, цель которой — выявление степени сходства между организмами для быстрой их идентификации и установления принадлежности к определенным таксонам. Если существующая систематика высших организмов отражает в определенной мере эволюционные связи между ними, т. е. признаки, используемые для выявления степени сходства, отражают и степень родства между этими организмами, то попытка создания на этой же основе систематики прокариот не была успешной.
ПРОБЛЕМЫ СИСТЕМАТИКИ ПРОКАРИОТ
В XX в. проблема систематики бактерий стала настоятельной в связи со стремительно увеличивавшимся как вширь (описание новых видов), так и вглубь (более детальное разностороннее изучение уже описанных видов) объемом знаний об этих организмах.
Хронологически первыми были попытки использовать накопленные сведения о фенотипических признаках бактерий для построения традиционной системы в виде некоего "генеалогического древа". В основу создания системы был положен и традиционный принцип. Все используемые признаки мысленно распределяли по степени их значимости, результатом чего являлось субъективное (основанное только на опыте и интуиции исследователя) создание иерархической системы признаков. Затем в зависимости от важности признаков объекты разбивали на таксономические группы. От того, в какой последовательности при классификации учитываются признаки, зависит путь, по которому осуществляют разделение микроорганизмов и в конечном счете полученные таксономические группы. Легко видеть, что структура иерархической системы определяется порядком, в котором располагаются признаки, а последний выбирается произвольно.
Вначале основное внимание уделяли морфологическим признакам бактерий. Так, в 1872 г. Ф. Кон (F. Cohn, 1828 — 1898) разделил бактерии на группы по морфологическим признакам: кокки, короткие палочки, удлиненные палочки, спирали. Однако вскоре стало ясно, что морфологические признаки недостаточны для удовлетворительного распределения бактерий по таксономическим группам. Для этой цели стали привлекать физиологические признаки. В 1909 г. С. Орла-Йенсен (S. Orla-Jensen) сделал попытку классифицировать бактерии на основе известных к тому времени физиологических признаков.
Важным шагом в развитии систематики прокариот явилось использование признаков, дающих информацию о химическом строении клетки: состав оснований ДНК, ДНК-ДНК- и ДНК-РНК-гомологии, аминокислотная последовательность белков, строение рибосом, компонентов клеточной стенки и т. д.
Первые предложенные схемы классификации бактерий были крайне субъективны. Это привело группу систематиков бактерий к использованию иного подхода для определения степени сходства между прокариотами — нумерической систематики. В основе ее лежат идеи, сформулированные в середине XVIII в. французским ботаником М. Адансоном (М. Adanson, 1727 — 1806): все признаки объекта считаются равноценными; при описании исследуемого объекта используется максимальное количество признаков, которые могут быть изучены и определены; степень сходства устанавливается на основании количества совпадающих признаков и выражается в виде коэффициента сходства. Последний для двух сравниваемых штаммов получают путем определения отношения числа одинаковых признаков к общему числу изученных признаков. Значение коэффициента сходства меняется в диапазоне от 1 (полная идентичность) до 0 (несовпадение ни по одному изученному признаку).
Классификация, построенная на принципах М. Адансона, — трудоемкий процесс, поэтому свое развитие и практическое применение она получила лишь в последнее время в связи с успехами в области вычислительной техники. Преимущества ее заключаются в формальном устранении элемента субъективности, поскольку все признаки объекта принимаются равноценными. Однако очевидны и ее слабые стороны. Как правило, для оценки сходства прокариот используют порядка 100 признаков, что составляет приблизительно 10% от количества признаков, определяющих бактериальный фенотип. Следовательно, учитывается только незначительная часть признаков классифицируемого объекта.
Кроме того, поскольку объем информации постоянно возрастает, значение коэффициента сходства может меняться в сторону увеличения (при обнаружении новых одинаковых признаков) или уменьшения (при увеличении числа несовпадающих признаков). Нумерическая таксономия может быть полезна при оценке степени сходства между таксонами невысокого ранга (виды, роды), но прямого отношения к созданию филогенетической системы прокариот не имеет.
Наиболее полно задача быстрой идентификации прокариотных организмов решается с помощью Определителя бактерий Берги, выпускаемого периодически Обществом американских бактериологов с привлечением крупных специалистов в области изучения тех или иных групп бактерий. Первое издание определителя было выпущено в 1923 г. группой американских бактериологов под руководством Д. X. Берги (D. H. Bergey, I860 — 1937); девятое издание в 4 томах вышло в 1984 — 1989 гг.
В девятом издании Определителя бактерий Берги все обнаруженные организмы, отнесенные в царство Prokaryotae, разделены на 33 группы. Признаки, по которым осуществляется разделение на группы, как правило, относятся к категории легко определяемых и вынесены в названия групп, например: грамотрицательные аэробные палочки и кокки (группа 4), анаэробные грамотрицательные кокки (группа 8), грамположительные палочки и кокки, образующие эндоспоры (группа 13), скользящие бактерии, образующие плодовые тела (группа 24). Основная идея классификации "по Берги" — легкость идентификации бактерий. Для осуществления этого используют совокупность признаков: морфологических (форма тела; наличие или отсутствие жгутиков; капсулы; способность к спорообразованию; особенности внутриклеточного строения; окрашивание по Граму), культуральных (признаки, выявляемые при культивировании в лаборатории чистой культуры), физиолого-биохимических (способы получения энергии; потребности в питательных веществах; отношение к факторам внешней среды; нуклеотидный состав и последовательность нуклеотидов в молекуле ДНК; наличие и характер минорных оснований в ДНК; нуклеотидный состав рибосомальной РНК; последовательность аминокислот в ферментных белках с аналогичными функциями).
Для девятого издания Определителя бактерий Берги характерен отказ от построения классической иерархической системы классификации. Она заменена списком упомянутых выше групп и сохранилась только фрагментами. Краткая характеристика этих групп дана в следующем разделе. Ценность определителя в том, что он представляет собой наиболее полную сводку известных бактериальных форм и самое современное пособие для идентификации бактерий.
В этом же руководстве предложена схема деления царства Prokaryotae на высшие таксоны (отделы, классы). В основу деления на отделы положено строение клеточной стенки. Название и краткая характеристика отделов и классов представлены в табл. 13. Представленная в Определителе бактерий Берги система классификации является строго идентификационной и не решает задачи выявления эволюционных связей между прокариотами. В то же время конечной целью является построение такой системы, в основе которой лежали бы родственные связи между прокариотными организмами. Первая попытка в этом направлении принадлежит С. Орла-Йенсену, предложившему филогенетическую систему бактерий, основанную на физиологических признаках. С. Орла-Йенсен исходил при этом из предположения, что, поскольку на первобытной Земле отсутствовало органическое вещество, первые бактерии должны были быть автотрофами. В качестве самых примитивных бактерий он рассматривал аэробные бактерии, окисляющие метан. Позднее такие попытки предпринимались крупнейшими микробиологами А. Клюйвером, К. ван Нилем, Р. Стейниером.
Важный шаг на пути создания естественной систематики прокариот связан с успехами молекулярной биологии. В 60-х гг. было установлено, что все свойства организма определяются уникальными химическими молекулами — ДНК, поэтому бактерии могут быть классифицированы путем сравнения их геномов. По такому признаку, как генетический материал, оказалось возможным на основании выявления степени сходства делать вывод о степени родства между организмами. Первоначально для таксономических целей сравнивали молярное содержание суммы гуанина и цитозина (ГЦ) в процентах от общего количества оснований ДНК у разных объектов. Этот показатель у прокариот колеблется от 25 до 75% 28. Однако ГЦ-показатель дает возможность только для грубого сравнения геномов. Если организмы имеют одинаковый нуклеотидный состав ДНК, возможно и сходство и различие между ними, поскольку генетическое кодирование основано не только на определенном содержании оснований в единице кодирования (триплете), но и на их взаимном расположении.
28 Колебания нуклеотидного состава ДНК у эукариотных микроорганизмов (молярная доля, %): грибы — 26 — 70, водоросли — 37 — 68, простейшие — 22 — 68; у высших растений и животных — 35 — 45. Колебания в составе оснований ДНК вирусов приблизительно такие же, как у прокариот.
Более тонкий метод оценки генетического сходства организмов — сравнение нуклеотидных последовательностей ДНК из разных источников методом ДНК-ДНК-гибридизации. Метод наиболее полезен для классификации на уровне вида, т. е. в случае высокой степени гомологии, и мало информативен для классификации объектов на уровне высоких таксонов29. В то же время часто несовпадение выводов, сделанных на основании фенотипических признаков и ДНК-гибридизации. В целом значение данных о строении ДНК для систематики прокариот огромно, так как позволяет перейти от установления степени сходства к выводам о степени родства между организмами.
29 Метод описан в книге Маниатиса Т., Фрича Э., Самбрука Дж Молекулярное клонирование. М.: Мир, 1984.
Помимо анализа молекул ДНК для установления степени родства между прокариотными организмами разработаны методические подходы, позволяющие сравнивать продукты отдельных генов, выполняющие в клетке одинаковые функции. Это могут быть белки (ферредоксины, цитохромы и др.) или рРНК. С использованием последних в качестве филогенетических маркеров связано крупное открытие, позволяющее по-новому взглянуть на систему живого мира.
Выбор рРНК для решения проблем эволюционной систематики прокариот оказался удачным по ряду причин: эти молекулы обнаружены у всех клеточных форм жизни, что указывает на их древнейшее происхождение; их функции всегда одинаковы; первичная структура в целом характеризуется высокой консервативностью. Особенностью рРНК является нахождение вне сферы действия отбора, поэтому данные молекулы эволюционируют в результате спонтанных мутаций, происходящих с постоянной скоростью, и накопление таких мутаций зависит только. от времени. Таким образом, мерой эволюционного расстояния между организмами служит количество нуклеотидных замен в молекулах сравниваемых рРНК.
Известно, что в рибосомах прокариот и эукариот присутствуют 3 типа рРНК, различающихся молекулярной массой и коэффициентом седиментации. Информационная емкость крупных молекул больше, но их труднее анализировать. Поэтому наиболее удобным оказался анализ молекул рРНК средней величины: 16S (у прокариот) и 18S (у эукариот), состоящих из 1600 и 2500 нуклеотидов соответственно. К настоящему времени последовательности 16S и 18S рРНК изучены более чем у 400 организмов, принадлежащих к разным царствам живой природы. На основании полученных данных рассчитаны коэффициенты сходства сравниваемых организмов, что привело к. неожиданным результатам: выявлены не две группы организмов, различающихся прокариотным и эукариотным типом клеточной организации, а три. Одну образуют все эукариоты: высшие растения, животные, дрожжи, водоросли и т. п. В эту группу не вошли органеллы эукариот (митохондрии, хлоропласты). Таким образом, первая группа представлена ядерно-цитоплазматическим компонентом эукариотных клеток. Ко второй группе, получившей название истинных бактерий, или эубактерий, относится подавляющее большинство прокариот. Сюда же попали на основании степени гомологии 16S рРНК митохондрии и хлоропласты эукариотных клеток. Наконец, в третью группу вошли некоторые малоизученные прокариоты, обитающие в экстремальных условиях: метанобразующие бактерии, экстремальные галофилы и термоацидофилы. Эта группа организмов получила название архебактерий.
Хотя клетки архебактерий структурно относятся к прокариотному типу, они построены из макромолекул (липидов, полисахаридов, белков), многие из которых являются уникальными и не синтезируются ни эукариотами, ни эубактериями. Архебактерии осуществляют ряд биохимических процессов, не свойственных остальным живым организмам. На основании этого был сделан вывод, что архебактерии, по-видимому, представляют собой одну из самых древних групп живых существ.
Обнаружение в недрах мира прокариот группы архебактерии поставило заново вопрос о путях клеточной эволюции с момента возникновения некоей гипотетической первичной клетки.
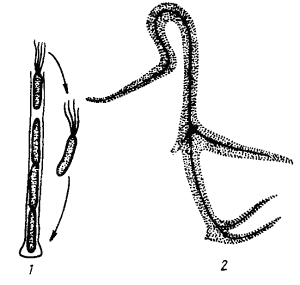
Традиционная общая схема клеточной эволюции основывается на следующих предположениях: из популяции первичных клеток в результате целого ряда событий, приведших к повышению уровня клеточной организации, под давлением естественного отбора возникла популяция предковых прокариотных клеток, из которых в конечном итоге произошли разные группы прокариот. Маловероятно, чтобы предковые прокариотные клетки все были "на одно лицо". Единственная их общая черта — прокариотная организация (см. табл. 1). Эукариотная клетка возникла в результате эндосимбиоза, в котором ядерно-цитоплазматическим компонентом, т. е. клеткой-хозяином, и эндосимбионтами, превратившимися впоследствии в митохонд-рии и хлоропласты, были существенно различающиеся между собой прокариотные клетки (рис. 41, А). Следствием такого взгляда на общий ход эволюции явилось признание двух основных царств живых организмов — Prokaryotae и Eukaryotae.
Новая схема клеточной эволюции исходит из признания существующих трех фундаментально различающихся типов живых организмов: эубактерий, архебактерии и эукариот. Согласно этой схеме от общего гипотетического предка, получившего название "прогенота", эволюционировали три различные ветви прокариот: эубактерий, архебактерии и уркариоты (рис. 41, Б). Уркариоты представлены ядерно-цитоплазматическим компонентом эукариотной клетки, включившим в себя в качестве эндосимбионтов представителей разных групп эубактерий, превратившихся в митохондрии и хлоропласты.
По целому ряду свойств на молекулярном уровне, и прежде всего по анализу 16 — 18S рРНК, выявленные три группы живых организмов значительно различаются между собой, что привело исследователей к выводу о приблизительно одинаковом "эволюционном" расстоянии между ними. На основании анализа 16S рРНК сделаны также первые попытки выяснить филогенетические взаимоотношения в группе эубактерий. Все изученные эубактерий в соответствии с вычисленными коэффициентами сходства оказались распределенными на 10 эволюционных ветвей. Наиболее неожиданный результат — обнаружение фотосинтезирующих эубактерий в 5 ветвях из 10, позволяющее сделать заключение о большем их родстве с определенными нефотосинтезирующими эубактериями, чем между собой. Это позволяет по-новому подходить к проблеме происхождения разных видов фотосинтеза и фотосинтезирующих эубактерий.

|
| Рис. 41. Пути клеточной эволюции. Тонкими стрелками обозначено эволюционирование разных групп прокариот, в том числе давших начало митохондриям (1), хлоропластам (2), эукариотному ядру и цитоплазме (3). Жирными стрелками обозначено эволюционирование разных групп эукариот |
Итак, в настоящее время отсутствует сколько-нибудь детализированная эволюционная система прокариот. Все описанные выше попытки подойти к ее созданию позволяют сделать вывод о том, что решение этой проблемы — дело неблизкого будущего. Особенности прокариот в области морфологической, физиолого- биохимической, генетической организации говорят о неприменимости к ним хорошо разработанных принципов, используемых при построении системы высших организмов. Это, естественно, значительно усложняет задачу, но не делает ее безнадежной. Уже сейчас в мире прокариот для наиболее изученной части этих организмов — эубактерий — можно проследить основные направления эволюционного развития. Одна из многообещающих идей заключается в том, что в основе прогрессивной эволюции эубактерий лежит совершенствование способов получения ими энергии.
Таблица 13. Деление царства Ргоkагуоtае на высшие таксоны (Murray, 1984)
| Отдел I. Gracilicutes | Отдел II. Firmicutes | Отдел III. Tenericutes | Отдел IV. Mendosicutes |
| Включает организмы с разной морфологией, имеющие грамотрицательную клеточную стенку. Размножение в основном бинарным делением, в некоторых группах – почкованием, в одной – множественным делением. Спор не образуют. Многие подвижны: с помощью жгутиков или скольжения. Аэробные, аназробные или факультативно аназробные формы. Отдел подразделяется на 3 класса, объединяющие нефотосинтезирующие (Scotobacteria) и фотосинтезирующие (Anoxyphotobacteria, Oxyphotobacteria) организмы | Входят организмы с грамположительной клеточной стенкой. Размножаются в основном бинарным делением. Некоторые образуют зндоспоры. У других споры на гифах или в спорангиях. Перемещаются с помощью жгутиков. Большинство неподвижны. В состав отдела входят азробные, анаэробные, факультативно анаэробные формы. В зависимости от морфологии предложено деление на 2 класса: Firmibacteria (кокки, палочки, неветвящиеся нити) и Thallobacteria (ветвящиеся формы) | Относятся прокариоты, у которых отсутствует клеточная стенка и не синтезируются предшественники пептидогликана. Клетки окружены ЦПМ, чрезвычайно плеоморфны. Размножение бинарным делением, почкованием, фрагментацией. Окрашивание по Граму отрицательное. Характерно образование мелких, врастающих в агар колоний. Могут быть сапрофитами, паразитами или патогенами. Представлены одним классом Mollicutes | Объединены прокариоты, по имеющимся данным, претендующие на более раннее происхождение, чем формы, включенные в 1 и 11 отделы. Клетки разной формы: кокки, палочки, нити. Многие плеоморфны. Большинство имеют клеточную стенку, но она не содержит типичного пептидогликана. Клеточная стенка может быть построена только из белковых макромолекул или гетерополисахаридов. Окрашивание по Граму отрицательное или положительное. Большинство – строгие аназробы. Многие имеют жгутики. Характеризуются экологическим и метаболическим разнообразием, способностью жить в экстремальных условиях. Объединены в класс Archaeobacteria |
Термины образованы от следующих латинских слов: cutes – кожа; gracilis – тонкий; firmus – крепкий, прочный; tener – мягкий, нежный; mendosus – ошибочный.
ГРУППЫ ПРОКАРИОТНЫХ ОРГАНИЗМОВ
В девятом издании Определителя бактерий Берги все прокариоты распределены по группам, не имеющим таксономического статуса. Ниже следует краткая характеристика этих групп. Группы бактерий, более подробно разбирающиеся в последующих главах, охарактеризованы намеренно предельно кратко. Наоборот, тем группам, которые в дальнейшем не обсуждаются, уделено несколько больше внимания.
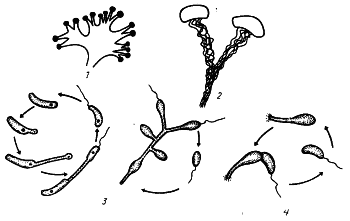
Группа 1. Спирохеты. Включает порядок Spirochaetales. Тонкие спиралевидные одноклеточные формы, обладающие своеобразной морфологией и способом движения (см. рис. 13). Длина клеток колеблется от 5 до 250 мкм. Склонны к образованию аномальных форм (гранул, цист). Размножаются поперечным делением. Клетки состоят из протоплазменного цилиндра, аксиальной нити и наружной оболочки. Оболочка тонкая и эластичная, что и обеспечивает спирохетам своеобразный способ передвижения. Грамотрицательны. Представители этой группы различно относятся к кислороду. Есть среди них облигатно аэробные, факультативно и облигатно анаэробные формы. Хемоорганогетеротрофы, существенно различающиеся по степени требовательности к субстрату. Среди них есть свободноживущие формы, основное место обитания которых — пресные и соленые озера, среда с высоким содержанием H2S; комменсалисты30, обитающие в желудочно-кишечном тракте пресноводных и морских моллюсков, и паразиты. Некоторые виды патогенны: Treponema pallidum — возбудитель сифилиса, Borrelia recurrentis — возбудитель возвратного тифа.
30 Комменсализм — вид симбиоза, когда один организм живет за счет другого, не причиняя ему, однако, какого- либо вреда.
Группа 2. Аэробные, подвижные спиралевидные или изогнутые грамотрицательные бактерии. Прокариоты, входящие в эту группу, имеют жесткую клеточную стенку, так что клетка свою форму не меняет. Движение осуществляется с помощью одного или множества полярно расположенных жгутиков. Хемоорганогетеротрофы, существенно различающиеся потребностями в питательных веществах. Некоторые виды могут расти на простой синтетической среде, для роста других необходимы сложные среды. К роду Spirillum относятся в основном сапрофиты, обитающие в стоячих и загрязненных водах, на гниющих растительных и животных остатках. Ряд свободноживущих форм — обитатели морских вод. Есть среди них и паразиты. Некоторые виды патогенны.
К этой же группе отнесен род Bdellovibrio. Обнаруженный в 1963 г. прокариотный организм, названный Bdellovibrio bacteriovorus31, вызвал огромный интерес, так как может паразитировать внутри клеток других бактерий. В. bacteriovorus — мелкая, слегка изогнутая палочка с одним полярно расположенным жгутиком. Последний несколько толще обычных бактериальных жгутиков.
31 Bdello — от латинского слова "пиявка", vorus — "пожирающий", в целом название, данное этим бактериям, можно перевести как "пиявковибрионы, пожирающие бактерии".
Внутри вида обнаружены штаммы, существующие только в клетках других бактерий, т. е. облигатные паразиты, и свободноживущие. Последние могут быть получены в лабораторных условиях из природных штаммов, ведущих паразитический образ жизни. Недавно описаны виды, являющиеся факультативными паразитами, т. е. способные существовать как внутри бактериальной клетки хозяина, так и сапрофитно на питательной среде сложного состава.
Особый интерес представляет способность Bdellovibrio паразитировать внутри клеток других бактерий. Спектр поражаемых бактерий очень широк: это различные виды грамположительных и грамотрицательных эубактерий. Bdellovibrio обладают способностью перемещаться с помощью своего развитого жгутика значительно быстрее многих других бактерий. Это дает им возможность активно искать и находить клетки хозяина. Настигнув бактериальную клетку, Bdellovibrio прочно к ней прикрепляется, пробуравливает клеточную стенку, проникает в пространство между клеточной стенкой и ЦПМ и начинает там размножаться. Как правило, в клетку проникают сразу несколько бактерий, поэтому целостность клеточной стенки нарушается в нескольких местах. Количество клеток-паразитов, образовавшихся в клетке-хозяине, может достигать 20 — 50 в зависимости от размеров хозяина. Клетка-хозяин напоминает в таких случаях "мешок", набитый Bdellovibrio. Цикл внутриклеточного развития В. bacteriovorus длится 3 — 5 ч, после чего паразиты освобождаются из разрушенных ими клеток-хозяев. Часть клеток Bdellovibrio переходит в покоящееся состояние, формируя цисты. В подходящих условиях (когда много бактериальных клеток) цисты быстро прорастают. Бактерии рода Bdellovibrio широко распространены в природе. Они обитают в почве, морских и пресных водах. Их используют для борьбы с возбудителями эпидемических заболеваний, например холеры.
Группа 3. Неподвижные грамотрицательные изогнутые бактерии. Включает семейство Spiromonaceae, объединяющее облигатно аэробные формы с характерной клеточной морфологией: от прямых палочек до колец, не полностью или полностью закрученных; при этом в культуре одновременно могут присутствовать клетки разной формы.
Группа 4. Грамотрицательные аэробные палочки и кокки. Группа представлена 8 семействами. К семейству Pseudomonadaceae относятся одиночные прямые или слегка изогнутые подвижные палочки. Движение осуществляется с помощью полярно расположенных жгутиков. Типичные представители семейства объединены в род Pseudomonas. Это в основном облигатно аэробные хемоорганогетеротрофы, потребляющие широкий набор органических соединений. Некоторые представители рода могут получать энергию также за счет окисления молекулярного водорода или окиси углерода, т. е. являются факультативными хемолитотрофами. Описаны виды, использующие в качестве конечного акцептора электронов нитраты, т. е. способные существовать факультативно аэробно.
Представители рода Pseudomonas повсеместно распространены в природе. Они постоянные обитатели воздуха, почв, морских и пресных вод, илов, сточных вод, где им принадлежит активная роль в минерализации органических веществ. Многие виды образуют водорастворимые и флюоресцирующие пигменты. Последние привлекли к себе внимание в связи с тем, что из них выделили вещества, обладающие антибиотической активностью против грибов, дрожжей, грамположительных и грамотрицательных эубактерий. Ряд видов Pseudomonas используют в микробиологической промышленности для получения различных органических соединений: кислот (пировиноградной, глюконовой, a- кетоглутаровой), аминокислот (глутаминовой, аспарагиновой, валина и др.), ферментов (аспарагиназы, перо-ксидазы). Некоторые виды вызывают заболевания растений и животных.
Семейство Azotobacteraceae объединяет виды, имеющие крупные клетки, склонные к изменению морфологии в зависимости от возраста культуры и условий культивирования. Среди представителей этого семейства встречаются подвижные и неподвижные формы. Бактерии рода Azotobacter образуют цисты. Хемоорганогетеротрофы. Способны активно фиксировать молекулярный азот. Облигатные аэробы. Обитают в почве, воде и на поверхности растений. Азотобактер — первый аэробный микроорганизм, для которого была показана способность фиксировать молекулярный азот.
В состав семейства Rhizobiaceae входят виды, во многих отношениях сходные с таковыми семейства Pseudomonadaceae, но отличающиеся от них плеоморфизмом, а также способностью вызывать разрастание тканей, приводящее к образованию клубеньков и галлов на корнях или стеблях различных видов растений. В род Rhizobium (клубеньковые бактерии) объединены бактерии, вызывающие образование клубеньков на корнях бобовых растений и способные фиксировать азот в условиях симбиоза с ними.
Клубеньковые бактерии проникают через корневые волоски в корневую систему растения и стимулируют деление тетраплоидных клеток корня, приводящее к образованию клубенька. В нем происходит интенсивное размножение бактерий. В молодых клубеньках большинство бактериальных клеток имеют форму палочек. В процессе последующего развития наблюдается образование клеток неправильной формы (бактероидов), в которых и происходит активная фиксация Na. Бактероиды можно рассматривать как дифференцированные формы, приспособленные для наилучшего осуществления определенной функции. В первую очередь это связано с контролированием поступления в бактероиды молекулярного кислорода, которое осуществляется с помощью находящегося в клубеньках леггемоглобина, цитоплазматического гемопротеина, в синтезе которого участвуют растение и бактерии. Апогемоглобин (белок без гема) синтезирует растение. За синтез гема ответственны бактероиды. Леггемоглобин, обладающий как и гемоглобин, высоким сродством к O2, обеспечивает перенос кислорода к бактероидам в количестве, необходимом для обеспечения их энергией, и в то же время защищает бактероиды от избытка O2 путем его связывания. К фиксации N2 способны только клубеньки, содержащие леггемоглобин.
Отношения между клубеньковыми бактериями и бобовыми растениями можно определить как мутуализм, т. е. такой вид симбиоза, при котором оба симбионта извлекают выгоду из сожительства: растение получает азот, клубеньковые бактерии — углеродсодержащие вещества и минеральные соли. Показана способность различных видов клубеньковых бактерий фиксировать N2 без какой-либо связи с растительными клетками. Для этого необходимо обеспечить клубеньковые бактерии подходящими источниками углерода (преимущественно пенто-зами), минимальным количеством фиксированного азота и промежуточными соединениями ЦТК. Свободноживущие клубеньковые бактерии синтезируют свой собственный гемоглобин, отличающийся структурно, но не функционально от леггемоглобина.
Все бактерии, принадлежащие к роду Agrobacterium, за исключением вида A. radiobacter, вызывают тканевые разрастания на стеблях различных растений, на основании чего они могут рассматриваться как внутриклеточные паразиты. Бактерии проникают в ткань растения-хозяина, используя для этого повреждения на его поверхности. Круг растений, поражаемых ими, очень широк. Например, типичный представитель рода A. tumefaciens вызывает образование галлов у растений, относящихся более чем к 40 семействам.
К семейству Methylococcaceae относятся бактерии, общим свойством которых является способность использовать метан в качестве единственного источника углерода и энергии в аэробных или микроаэробных условиях.
Группа 5. Факультативно анаэробные грамотрицательные палочки. Объединяет 3 семейства: Enterobacteriaceae, Vibrionaceae и Pasteurellaceae. В состав первого семейства входят подвижные или неподвижные бесспоровые палочки. Некоторые виды образуют капсулы. Хемоорганогетеротрофы. Отдельные представители характеризуются высокими пищевыми потребностями. Энергию получают за счет дыхания или брожения. Представители этого семейства широко распространены в природе. Среди них много патогенных и сапрофитных видов. Это обитатели почв, морских и пресных вод, разлагающихся остатков Животных и растений, постоянные обитатели кишечника человека и многих видов животных, птиц, рыб, рептилий.
Наиболее изученный представитель семейства — Escherichia coli. Эта бактерия всегда содержится в кишечнике человека и животных, поэтому о загрязнении воды и пищевых продуктов судят по наличию в них Е. coli. E. coli относится к числу условно патогенных бактерий, т. е. является постоянным компонентом микрофлоры кишечника человека, но при ослаблении защитных функций организма может проникать в другие органы и вызывать сильные воспалительные процессы. К числу возбудителей тяжелых заболеваний человека принадлежат бактерии из родов Salmonella и Shigella. Salmonella typhi — возбудитель брюшного тифа; разные виды Shigella — возбудители так называемой бактериальной дизентерии.
Группа 6. Анаэробные грамотрицательные прямые, изогнутые или спиралевидные палочки. Основная таксономическая единица группы — семейство Bacteroidaceae. Это палочки правильной формы или склонные к плеоморфизму, бесспоровые, неподвижные или подвижные. Облигатные анаэробы. Хемоорганогетеротрофы. При сбраживании глюкозы образуют смесь кислот. Основное место обитания — кишечник человека и животных, пищеварительный тракт насекомых. Некоторые виды патогенны и вызывают различные поражения кожных покровов, а также других органов и тканей тела.
Группа 7. Бактерии, характеризующиеся диссимиляционным восстановлением серы или сульфата. В составе группы эубактерии с разной морфологией и следующими одинаковыми свойствами: грамотрицательные строгие анаэробы, использующие в качестве акцептора электронов молекулярную серу или ее окисленные соединения, которые восстанавливаются при этом до H2S. Некоторые виды способны к брожению. Есть среди них азотфиксаторы.
Группа 8. Анаэробные грамотрицательные кокки. Представлена одним семейством Veillonellaceae. Кокки, как правило, соединены попарно, но могут образовывать цепочки или скопления клеток. Хемоорганогетеротрофы с высокими потребностями в питательных веществах. В эту группу входят только паразиты теплокровных животных: обитатели пищеварительного тракта, ротовой полости и дыхательных путей человека и животных.
Группа 9. Риккетсии и хламидии. В состав группы включены два порядка: Rickettsiales и Chlamydiales. Первый объединяет бактерии, характеризующиеся в большинстве случаев совокупностью следующих признаков: плеоморфные, неподвижные, грамотрицательные, с типичными для эубактерий клеточными стенками, размножающиеся делением внутри клеток-хозяев; культивировать можно на специальных средах, содержащих живые или переживающие ткани, такие как куриные эмбрионы или культуры клеток позвоночных. Однако перечисленные выше признаки свойственны не всем представителям этого порядка. Среди риккетсий имеются подвижные виды, красящиеся положительно по Граму, а также виды, которые можно выращивать на относительно простых искусственных питательных средах. Различны взаимоотношения между риккетсиями и организмами-хозяевами. Помимо паразитизма эти отношения в некоторых случаях можно определить даже как мутуалистические. Среди риккетсий-паразитов большая часть относится к непатогенным и только меньшая вызывает заболевания человека, позвоночных и беспозвоночных животных (так называемые риккетсиозы). Успехи последнего времени, связанные с изучением риккетсий, позволили многое понять в биологии этих организмов, но многое еще остается не выясненным.
Все исследованные риккетсий обладают определенной активностью энергетических и биосинтетических процессов. У них найдена цитохромная система и показано запасание энергии, освобождающейся в процессе дыхания, в виде АТФ. Риккетсии могут осуществлять некоторые биосинтетические процессы, например биосинтез белка и липидов.
Порядок Chlamydiales включает одно семейство Chlamydiacaaa и один род Chlamydia. Хламидии — облигатные внутриклеточные паразиты позвоночных и человека, характеризующиеся сложным циклом развития. Могут размножаться только в цитоплазме клеток. Вне клеток их культивировать пока не удается.
Облигатный внутриклеточный паразитизм хламидии наложил специфический отпечаток на их метаболизм. Прежде всего это коснулось их энергетического метаболизма. Обладая способностью осуществлять определенные реакции окислительного характера (например, при добавлении необходимых кофакторов окислять глюкозу, пировиноградную и глутаминовую кислоты), хламидии не могут синтезировать высокоэнергетические соединения, и в первую очередь АТФ, поэтому они получили название "энергетических паразитов". Хламидии паразитируют в организме различных позвоночных (птиц, человека и других млекопитающих), вызывают у человека ряд заболеваний, например трахому и воспаления дыхательных органов.
Группа 10. Микоплазмы. К ним относятся формы, у которых отсутствует клеточная стенка. Таксономическая значимость этого признака позволила все прокариоты, не имеющие клеточной стенки, выделить в группу, присвоив ей ранг отдела (см. табл. 13). В девятом издании Определителя бактерий Берги микоплазмы отнесены к отделу Tenericutes, классу Mollicutes, порядку Mycoplasmatales32.
32 От греческих слов: myce — гриб; plasma — плазма.
Отсутствие ригидной клеточной стенки повлекло за собой ряд морфологических, культуральных, цитологических особенностей, присущих этим микроорганизмам. Для них характерен ярко выраженный полиморфизм. В культуре одного вида можно одновременно обнаружить крупные шаровидные тела, мелкие зерна, клетки эллипсовидной, дискообразной, палочковидной и нитевидной формы. Последние могут ветвиться, образуя структуры, подобные мицелиальным. Для микоплазм описаны различные способы размножения: бинарное деление, фрагментация крупных тел и нитей, процесс, сходный с почкованием.
В культурах микоплазм обнаружены формы с наименьшими из всех известных клеточных микроорганизмов размерами. Поэтому, вероятно, именно микоплазмы можно считать наиболее простыми самостоятельно воспроизводящимися системами. По проведенным подсчетам теоретически наименьшая структурная единица, способная к самостоятельному воспроизведению на искусственной среде, не может иметь размеры меньше, чем сферическое тело диаметром 0,15 — 0,20 мкм или нить длиной приблизительно 13 мкм и диаметром примерно 20 нм. Все эти структуры встречаются в культурах микоплазм и, вероятно, могут рассматриваться, как жизнеспособные репродуцирующиеся формы. По объему генетической информации, содержащейся в геноме, микоплазмы занимают промежуточное положение между Е. coli и Т-фагами.
Отсутствие клеточной стенки привело к развитию у микоплазм более стабильной и эластичной ЦПМ по сравнению с ЦПМ бактериальных протопластов. Важная роль в обеспечении этих свойств принадлежит, по-видимому, холестерину — основному компоненту мембранных липидов паразитических микоплазм. Большая часть известных микоплазм для роста нуждается в экзогенном холестерине и других стеринах. Относительно недавно были обнаружены виды, не требующие для роста экзогенных стеринов. Это различие положено в основу деления порядка Mycoplasmatales на семейства Mycoplasmataceae и Spiroplasmataceae, в которых объединены стеринзависимые микоплазмы, и Acholeplasmataceae, куда вошли виды, не требующие для роста экзогенных стеринов. Отсутствие клеточной стенки обусловливает еще одну отличительную особенность микоплазм — их нечувствительность к антибиотикам, специфически действующим на эубактериальную клеточную стенку, и в первую очередь, к пенициллину и его аналогам.
Микоплазмы (особенно после обнаружения новых свободноживущих видов) представляют собой группу, чрезвычайно разнообразную с точки зрения физиолого-биохимических особенностей. Эти прокариоты могут расти на искусственных средах разной степени сложности (от простых минеральных сред до сложных органических) или только внутри организма-хозяина, из чего можно заключить, что диапазон их биосинтетических способностей весьма широк. Разнообразны и способы получения микоплазмами энергии. Среди них описаны виды, получающие энергию за счет окисления или сбраживания органических соединений (моно- и полисахаридов), а также, возможно, окисления неорганических соединений железа. Описаны микоплазмы, являющиеся строгими аэробами и облигатными анаэробами.
Если раньше считали, что микоплазмы — в основном формы, паразитирующие на человеке и высших животных, то теперь представление о способах существования и распространения этой группы прокариот в природе значительно расширено. Микоплазмы находят в почве и сточных водах, они выделены из каменного угля и горячих источников. Помимо свободножи-вущих форм, способных расти как на чисто минеральных средах, так и сапрофитно, описаны микоплазмы, существующие в различных симбиотических ассоциациях с бактериями, низшими грибами, растениями, птицами, высшими животными и человеком. Формы симбиоза также разнообразны. Иногда это, вероятно, комменсализм, в большинстве случаев — типичный паразитизм. Многие паразитические формы микоплазм патогенны. Они являются возбудителями заболеваний растений, животных и человека, например, М. pneumoniae — возбудитель острых респираторных заболеваний и пневмоний у человека.
Представители семейства Mycoplasmataceae — хемооргано-гетеротрофы, характеризующиеся высокими потребностями в питательных веществах. Энергетический метаболизм ферментативного или окислительного типа. Использование глюкозы происходит по гликолитическому пути. У микоплазм, осуществляющих полное окисление энергетического субстрата, обнаружен функционирующий ЦТК и цепь переносчиков электронов.
В состав семейства Acholeplasmataceae входит один род Acholeplasma, насчитывающий 8 видов стериннезависимых микоплазм. Наиболее хорошо изучена A. laidlawii — первая сапрофитная микоплазма, выделенная в 1936 г. из сточных вод Лондона. Сейчас в составе рода объединены свободноживущие сапрофитные микоплазмы, микоплазмы — паразиты млекопитающих и птиц; некоторые из них, возможно, патогенны.
В третье семейство Spiroplasmataceae выделены микоплазмы, схожие с таковыми семейства Mycoplasmataceae, но отличающиеся своеобразной морфологией: в стадии роста среди разнообразных форм преобладают спиралевидные нити. Из листьев цитрусовых растений выделена Spiroplasma citri. Особенностью ее строения является часто обнаруживаемый на мембране наружный слой, который, возможно, представляет собой модифицированную клеточную стенку или структуру, весьма напоминающую последнюю.
Группа 11. Эндосимбионты. В эту группу выделены прокариоты — эндосимбионты простейших, насекомых, грибов и беспозвоночных. Для большинства представителей эндосимбиоз об- лигатен и их не удалось культивировать в лаборатории в чистой культуре. Основные методы изучения эндосимбионтов — цитологические, с применением световой и электронной микроскопии. Важным признаком служит характеристика взаимоотношений с хозяевами, а также морфология эндосимбионтов, циклы развития, специфичность локализации в клетке хозяина. Спектр отношений эндосимбионтов с хозяевами очень широк: от мутуализма до паразитизма с проявлениями патогенности. Некоторым эндосимбионтам присвоены биномиальные названия. Так, среди эндосимбионтов простейших описано 5 родов и 14 видов.
Группа 12. Грамположительные кокки. В состав группы входят представители 15 родов, значительно различающихся филогенетически и фенотипически. Это облигатные аэробы, анаэробы или факультативные формы. Энергию получают за счет дыхания и/или брожения. Хемоорганогетеротрофы с различными потребностями в питательных веществах.
Бактерии, объединяемые в семейство Micrococcaceae — кокки, делящиеся более чем в одной плоскости, склонные не расходиться после деления и поэтому образующие скопления сферической или неправильной формы. В основном сапрофиты. Разрушая многие сложные органические вещества, выполняют функцию "мусорщиков".. К этой же группе отнесен род Streptococcus, представители которого получают энергию, осуществляя гомоферментативное молочнокислое брожение, и род Leuconostoc, бактерии, входящие в его состав, осуществляют гетероферментативное молочнокислое брожение. Представители этой группы обнаружены в почве, на поверхности злаков, в ротовой полости, желудочном тракте и дыхательных путях человека и животных. Некоторые виды, преимущественно относящиеся к роду Staphylococcus, патогенны.
Группа 13. Грамположительные палочки и кокки, образующие эндоспоры. В составе группы представители 6 родов. Два из них (Bacillus и Clostridium) наиболее многочисленны и интересны. Род Bacillus объединяет подвижные палочковидные клетки, размеры которых колеблются в довольно широких пределах. Жгутики расположены перитрихиально. Окрашивание по Граму различно: положительно или положительно только в молодой культуре. Облигатные или факультативные аэробы. Бактерии рода Bacillus синтезируют различные литические ферменты, расщепляющие полисахариды, белки, жиры и другие макромолекулы. Некоторые виды образуют антибиотики, такие как бацитрацин, субтилизин. Большинство бацилл — сапрофиты. Основное место их обитания — почва. Есть среди них и патогенные для животных и человека формы, например B. anthracis — возбудитель сибирской язвы, а также виды, вызывающие различные заболевания членистоногих.
В состав рода Clostridium входят палочки, отличающиеся от предыдущего рода формой спорообразования и облигатно анаэробным способом существования. Источник энергии в большинстве случаев — маслянокислое брожение. Большинство бактерий рода Clostridium — сапрофиты, обитатели почвы. Некоторые виды живут в кишечнике человека и животных. К этому роду относятся весьма опасные патогенные формы: С. tetani — возбудитель столбняка, С. perfringens и некоторые другие виды клостридиев — возбудители газовой гангрены, С. botulinum — продуцент экзотоксина, одного из самых сильных биологических ядов.
Группа 14. Грамположительные, не образующие спор палочки правильной формы. Группа — конгломерат, состоящий из 7 родов, объединенных несколькими общими морфологическими и физиологическими признаками: клетки палочковидной формы (от кокковидных до удлиненных, одиночных или образующих цепочки), мезофилы; строгие или факультативные аэробы, есть микроаэрофилы и аэротолерантные анаэробы; хемоорганогетеротрофы, растущие только на сложных средах.
Представители рода Lactobacillus, в составе которого около 50 видов, получают энергию в процессе гомоферментативного или гетероферментативного молочнокислого брожения. Широко распространены в природе: их можно обнаружить в почве, на разлагающихся остатках животного и растительного происхождения, в молоке и молочных продуктах, в кишечнике позвоночных; лишь единичные представители рода Lactobacillus обладают патогенными свойствами.
Группа 15. Грамположительные, не образующие спор палочки неправильной формы. Группа разнообразна. Большинство — Грамположительные палочки неправильной формы, растущие в присутствии воздуха и не образующие эндоспор, но есть в группе бактерии, имеющие форму кокков или палочек правильной формы, окрашивающиеся отрицательно по Граму и являющиеся строгими анаэробами.
Так, к роду Corynebacterium относятся формы, склонные к морфологической изменчивости. Кроме коротких палочек в культуре можно обнаружить кокковидные формы, клетки, имеющие булавовидные выпячивания, слабоветвящиеся формы. Для представителей этого рода характерно образование фигур, состоящих из расположенных под углом или примыкающих друг к другу дочерних клеток. Неподвижны. Хемоорганогетеротрофы. Энергию получают за счет дыхания или брожения. Преимущественно факультативные анаэробы, но некоторые — аэробы.
В состав рода входят свободноживущие виды, а также паразиты человека и животных. Некоторые из них патогенные, например, С. diphteriae — возбудитель дифтерии. Большая группа коринебактерий — возбудители болезней растений.
К группе отнесены и бактерии рода Arthrobacter, для которых характерна большая по сравнению с предыдущим родом тенденция к ветвлению и образованию кокковидных клеток. Культуры, находящиеся в экспоненциальной фазе роста, неправильной палочковидной формы; культуры же, перешедшие в стационарную фазу, состоят в основном или исключительно из кокковидных форм. При перенесении последних на свежую питательную среду происходит "удлинение" кокковидных клеток путем образования выпячиваний; у одной клетки может быть от двух до четырех таких выпячиваний, приводящих к появлению палочек неправильной формы или клеток с рудиментарным ветвлением. Виды в молодой культуре (на стадии палочек) неподвижны или подвижны, окраска по Граму в этот период может быть нечеткой, но у кокковых форм она положительна. Все виды — облигатно аэробные хемоорганогетеротрофы. Бактерии рода Arthrobacter — основные представители микрофлоры почвы, активно участвующие в разложении органических веществ.
Группа 16. Микобактерии. Объединены в семейство Micobacteriaceae и представлены одним родом Mycobacterium. Микобактерии — грамположительные, неподвижные палочки, прямые или неправильных очертаний (рис. 42, А). В процессе развития палочковидные формы превращаются в кокковидные. Характерным морфологическим признаком микобактерий является образование ветвящихся форм. Степень ветвления зависит от вида бактерий и условий выращивания, в первую очередь от состава питательной среды. Ветвление можно наблюдать только в молодых активно размножающихся культурах. У микобактерий мицелий не образуется. На некоторых стадиях роста характерна повышенная устойчивость к кислотам и спиртам. Большинство микобактерий — сапрофиты, живущие в почве и использующие различные органические соединения (белки, углеводы, жиры, воска, парафины). Некоторые виды патогенны, например, М. tuberculosis — возбудитель туберкулеза, М. leprae — возбудитель проказы.

|
| Рис. 42. Микобактерии (А) и нокардии (Б):1 — воздушный; 2 — субстратный мицелий |
Группа 17. Нокардиоформы. В этой группе объединены бактерии, в цикле развития которых существует мицелиальная стадия. В старых культурах мицелий распадается на палочковидные или коккоидные элементы (рис. 42, 5). Настоящих спор нет. Группа достаточно разнообразна по морфологическим и физиолого-биохимическим признакам: большинство представителей (но не все) обнаруживают в старых культурах фрагментацию мицелия, образование воздушного мицелия, формирование конидий.
Все представители группы грамположительные аэробы. Молярная доля ГЦ в составе ДНК — 63 — 79%. В основу деления на роды положены такие признаки, как химический состав клеточной стенки, набор липидов.
Группа 18. Фототрофные бактерии, осуществляющие бескислородный фотосинтез. В эту группу отнесены фотосинтезирующие эубактерии, характеризующиеся специфическим набором пигментов и особым типом фотосинтеза: пигменты представлены различными видами бактериохлорофилла и каротиноидов; фотосинтез не сопровождается выделением кислорода.
Группа 19. Фототрофные бактерии, осуществляющие кислородный фотосинтез. Группа представлена эубактериями, содержащими разные наборы фотосинтетических пигментов, но обязательно — хлорофилл a; фотосинтез сопровождается выделением молекулярного кислорода.
Группа 20. Аэробные хемолитотрофные бактерии и близкие к ним организмы. К этой группе относятся прокариоты, получающие энергию за счет окисления восстановленных неорганических соединений азота, серы, железа, а также молекулярного водорода. Группа разделена на 4 подгруппы в зависимости от химической природы окисляемых неорганических соединений.
В первую подгруппу включены грамотрицательные бактерии, объединенные в семейство Nitrobacteraceae, источником энергии для которых являются процессы окисления аммонийного азота или нитритов. Во второй подгруппе объединены бактерии, способные окислять неорганические восстановленные соединения серы. У большинства из них доказана способность использовать этот процесс для получения клеточной энергии. Облигатно хемолитотрофные водородные бактерии, представленные одним родом Hydrogenobacter, выделены в третью подгруппу. В четвертую подгруппу отнесены бактерии, способные окислять и/или откладывать вне клетки окислы железа и марганца. Последние накапливаются в капсулах или во внеклеточном материале, редко — внутри клетки. Поскольку большинство бактерий этой подгруппы не получено до сих пор в чистой культуре, многие стороны их метаболизма остаются неясными.
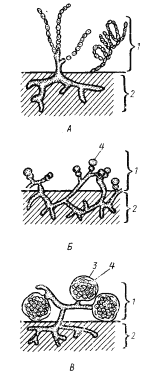
Группа 21. Почкующиеся и/или стебельковые бактерии. В эту группу входят бактерии, образующие состоящие из слизи отростки (стебельки), не связанные с цитоплазмой клетки, или нитевидные клеточные выросты — простеки (рис. 43). Бактерии рода Nevskia образуют слизистые отростки, не связанные с цитоплазмой клетки. Слизь, выделяющаяся с одной стороны клетки, имеет вид стебелька, на конце которого располагается клетка. Стебельки обнаруживают дихотомическое ветвление, повторяющее деление зрелых клеток (рис. 43, 1). Своеобразный вид имеют стебельки, образуемые бактериями рода Gallionella: клетки, имеющие бобовидную форму, на вогнутой стороне выделяют гидроокись железа в коллоидной форме в виде многочисленных тонких фибрилл, образующих спирально извитую ленту (рис. 43, 2). Изучение Gallionella в чистой культуре показало, что энергия, освобождающаяся при окислении железа, не используется клеткой.
|
| Рис. 43. Почкующиеся и/или стебельковые бактерии: 1 — Nevskia; 2 — Gallionella; 3 — Hyphomicrobium; 4 — Caulobacter (по Brock, 1970; Lechevalier, Pramer, 1971; Schlegel, 1972) |
Выросты представляют собой выпячивание клеточного содержимого, не отделенного от цитоплазмы клетки. Окружены клеточной стенкой. В них можно различить цитоплазматическую мембрану, цитоплазму с рибосомами, иногда ядерный материал и мезосомы. Выросты приводят к увеличению клеточной поверхности и ЦПМ и служат для обеспечения повышенного транспорта веществ в клетку. Для простекобактерий это имеет первостепенное значение, так как многие из них обитают в условиях низкой концентрации органических веществ в среде. Общим свойством простекобактерий является способность расти с сохранением типичной морфологии только при незначительном содержании органического субстрата в среде. При дефиците питательных веществ выросты удлиняются. С увеличением концентрации необходимых для роста питательных компонентов выросты сильно сокращаются или исчезают совсем.
У некоторых бактерий выросты имеют отношение к функции размножения. Бактерии, принадлежащие к роду Hyphomicrobium, — палочки с заостренными концами, но могут быть овальной, яйцеобразной или бобовидной формы. Для них характерен своеобразный цикл развития (рис. 43, 3). Прикрепленная к субстрату материнская клетка образует нитевидный вырост, куда переходит один из поделившихся нуклеоидов. Вырост, удлиняясь, формирует гифоподобную структуру, на конце которой появляется почка. В процессе созревания почки образуется жгутик. Дочерняя клетка (созревшая почка) отделяется от материнской и в течение некоторого времени подвижна. Затем она прикрепляется к субстрату или другим клеткам, теряет жгутик и формирует вырост и почку. Нитевидные выросты клетки могут ветвиться, и на концах каждой ветви формируются почки. В некоторых случаях созревшие почки не отделяются от материнской клетки и в свою очередь формируют выросты и почки. В результате имеет место скопление гиф и клеток. Выросты могут появляться на обоих полюсах клетки.
У бактерий рода Caulobacter клеточные выросты не имеют отношения к репродуктивной функции (рис. 43, 4). У представителей этого рода клетки большей частью палочковидной формы с одним полярно расположенным жгутиком. Прикрепившись к какой-нибудь поверхности тем концом, на котором расположен жгутик, они формируют вырост, имеющий такое же происхождение и внутреннее строение, как у бактерий рода Hyphomicrobium. На конце выроста выделяется небольшое количество клейкого вещества, с помощью которого клетка прикрепляется к субстрату. Размножение осуществляется поперечным делением, при этом к концу деления одна клетка несет стебелек, другая — жгутик. После разделения клетка, снабженная жгутиком, формирует стебелек с клейким веществом, прикрепляется к субстрату и переходит в неподвижную фазу. В эту же группу отнесены и бактерии, размножающиеся почкованием, но не образующие выросты.
Большинство рассмотренных выше бактерий — хемоорганогетеротрофы; некоторые — олигокарбофилы. Как правило, облигатные аэробы, есть и факультативные аэробы. Описаны виды, тяготеющие к низким концентрациям кислорода в среде (микроаэрофилы).
|
| Рис. 44. Бактерии, образующие слизистую оболочку (влагалище): 1 — Sphaerotilus; 2 — Leptothrix (no Lechevalier, Pramer, 1971) |
Группа 22. Бактерии, образующие слизистую оболочку (влагалище). В состав группы входят нитевидные бактерии, окруженные общим влагалищем (рис. 44). Нити могут быть свободно плавающими или прикрепленными к различным находящимся в воде предметам. Влагалище состоит из гетерополисахарида, часто инкрустированного окислами железа или марганца. Клетки размножаются внутри влагалища поперечным делением. Выходящие из влагалища одиночные клетки могут быть снабжены жгутиками, с помощью которых они перемещаются, или же жгутики отсутствуют, и одиночные клетки не способны к активному движению. Все бактерии этой группы — аэробы и хемоорганогетеротрофы. Наиболее распространены представители родов Sphaerotilus и Leptothrix. Бактерии рода Sphaerotilus — типичные обитатели сточных вод. Они хорошо растут в проточной воде, богатой органическими веществами, а представители рода Leptothrix — в бедных органическими веществами местах с высоким содержанием железа.
Группа 23. Нефотосинтезирующие скользящие бактерии, не образующие плодовых тел. К этой группе отнесены морфологически и физиологически разнообразные бактерии. Большинство объединяет способность передвигаться по твердому субстрату без помощи жгутиков. Внутри группы выделены 3 порядка. Основной по числу представителей — порядок Cytophagales. В него помещены грамотрицательные бактерии, имеющие палочковидную форму, часто плеоморфные. Способны использовать различные полисахариды (агар, целлюлозу, хитин, крахмал, пектин и др.). Источником энергии служит дыхание, но некоторые могут получать энергию за счет брожения.

|
| Рис. 45. Представители группы нефотосинтезирующих скользящих бактерий, не образующих плодовых тел: 1 — Beggiatoa; 2 — Vitreoscilla; 3 — Leucothrix (no Lechevalier, Pramer, 1971) |
В порядок Beggiatoales объединены нитчатые формы. Нити эластичны и способны к скользящему движению. Разделение на роды осуществляется в зависимости от способности откладывать или нет в клетке гранулы серы при росте в присутствии сульфида (рис. 45, 1, 2). Сходной морфологией обладают бактерии рода Leucothrix. Они образуют длинные нити, состоящие из овальных или цилиндрических клеток. Нити обычно прикреплены к субстрату и неподвижны (рис. 45, 3). Размножаются с помощью одиночных подвижных клеток, выходящих из нити. Во многих отношениях напоминают нитчатые цианобактерии, отличаясь отсутствием фотосинтетических пигментов.
Группа 24. Скользящие бактерии, образующие плодовые тела: миксобактерии. Включает один порядок Myxococcales, подразделяющийся на 4 семейства. Это палочковидные грамотрицательные бактерии, имеющие тонкие эластичные клеточные стенки. Для них характерно образование слоя слизи, окружающего клетку. Бактерии могут передвигаться по твердому субстрату скользящими движениями. Локомоторные структуры (жгутики) отсутствуют. Миксобактерии образуют так называемые плодовые тела, внутри которых клетки переходят в покоящееся состояние (см. рис. 21). Представители порядка — облигатно аэробные хемоорганогетеротрофы. Энергию получают только за счет дыхания. Синтезируют активные литические ферменты, способные гидролизовать такие макромолекулы, как полисахариды (целлюлоза, клетчатка, хитин), белки, нуклеиновые кислоты, эфиры жирных кислот. С этим связана роль миксобактерии в природе: они активно разрушают мертвые растительные остатки. Многие миксобактерии способны лизировать. клетки прокариотных и эукариотных микроорганизмов. Хорошо растут на поверхности твердых сред. Основное место обитания — почва.
Группа 25. Архебактерии. В соответствии с современными представлениями в эту группу выделены прокариоты, представляющие одну из трех линий эволюции жизни (см. рис. 41, Б).
В IX издании Определителя бактерий Берги впервые сделана попытка классифицировать известные архебактерии. Они разделены на 5 подгрупп. В I, самую большую, подгруппу включены метаногенные бактерии, главным и характерным признаком которых является способность образовывать метан в качестве конечного продукта энергетического метаболизма.
Во II подгруппу отнесены экстремально термофильные, строго анаэробные формы, образующие H2S из сульфата в процессе диссимиляционной сульфатредукции.
Экстремально галофильные архебактерии, составляющие III подгруппу, представлены грамположительными или грамотрицательными формами, аэробными или факультативно анаэробными хемоорганотрофами. Характерна потребность в высоких концентрациях NaCl. Некоторые виды содержат бактериородопсин и способны использовать энергию света для синтеза АТФ. В природе распространены в местах с высокой концентрацией соли: в соленых озерах, белковых продуктах, законсервированных с помощью соли, например в соленой рыбе.
IV группа представлена архебактериями без клеточной стенки. В ее составе один род Thermoplasma, вид Т. acidophilum.
К V подгруппе отнесены архебактерии, характеризующиеся совокупностью следующих признаков: облигатные термофилы; ацидофилы или нейтрофилы; аэробы, факультативные или строгие анаэробы; автотрофы или гетеротрофы. Метаболизм большинства из них связан с молекулярной серой (S0). Представители порядка Thermoproteales в процессе хемолитоавтотрофного. роста получают энергию в реакции:
H2 + S0 « H2S.
При использовании органических субстратов S0 служит конечным акцептором электронов, конечные продукты энергетического метаболизма — CO2 и H2S.
Представители порядка Sulfolobales могут в аэробных условиях окислять H2S до S0 и далее до SO42 – , а в анаэробных — восстанавливать S0 до H2S с участием H2.
Группы. 26 — 33: актиномицеты. Сравнительный анализ 16S рРНК привел к заключению, что все грамположительные эубактерии образуют одну из 10 основных филогенетических ветвей, выявленных среди изученных прокариот. В свою очередь грамположительные эубактерии на основании данных по нуклеотидному составу ДНК распадаются на 2 основные ветви: к одной относятся организмы, молярное содержание ГЦ в ДНК которых составляет больше 55%, к другой — группы Bacillus — Clostridium — Streptococcus с содержанием в ДНК ГЦ меньше 50%. В соответствии с проведенными исследованиями грамположительные эубактерии с молярным содержанием ГЦ-оснований больше 55%, обнаруживающие родство на основании данных анализа 16S рРНК и гибридизации ДНК, относят к актиномицетам.
При последующей их систематике большое значение придают морфологическим признакам. Эта группа объединяет организмы с разной морфологией: от кокков и палочек до форм, образующих ветвящиеся нити или формирующих развитый мицелий. В последнем случае при выращивании актиномицетов на твердых питательных средах различают субстратный и воздушный мицелий. Субстратный мицелий развивается в толще агаризованной среды, над поверхностью которой разрастаются гифы воздушного мицелия. Актиномицеты характеризуются разными способами размножения. Большинство размножаются с помощью спор, образующихся в специальных органах спороношения — спорангиях. Последние различаются строением (длинные или короткие, прямые или спиралевидные с разным числом завитков) и расположением (последовательное, супротивное, мутовчатое и др.).
В последнее время все больший удельный вес в таксономических целях занимают сведения о химическом составе и структуре отдельных клеточных компонентов: генетическом материале, клеточной стенке, мембранах и др. ГЦ-показатель ДНК актиномицетов колеблется в пределах от 58 до 75%. На основании присутствия характерных аминокислот пептидного хвоста пептидогликана и Сахаров, входящих в состав полисахаридов, у актиномицетов выделено несколько типов клеточной стенки. Кроме того, в систематике актиномицетов используют культуральные, физиолого-биохимические, экологические признаки. Только результаты анализа 16S рРНК у актиномицетов, относящихся к разным родам, позволили разделить их на 7 групп. В большинстве случаев эти группы совпадают с предложенными в IX издании Определителя бактерий Берги, хотя при классификации последних наряду с данными анализа 16S рРНК и нуклеотидного содержания ДНК учитывались также морфологические, химические и физиологические признаки. Вопрос о присвоении даже некоторым выделенным группам таксономического ранга пока не ясен.
В дальнейшем при характеристике выделенных в определителе групп актиномицетов мы остановимся в основном на их краткой морфологической характеристике.

|
| Рис. 46. Актиномицеты рода Dermatophilus (А), Actinoplanes (Б), Micromonospora (В): 1 — воздушный; 2 — субстратный мицелий; 3 — спорангий; 4 — споры; 5 — капсула |
Группа 26. Нокардиоподобные актиномицеты. В этой группе дана характеристика организмов, уже описанных в группе 17 (см. рис. 42, 5), но несколько более детализированная и дополненная отнесением к ней представителей еще двух родов.
Группа 27. Актиномицеты. с многоклеточными спорангиями. В эту группу выделены актиномицеты, характеризующиеся своеобразным строением вегетативного тела (таллома): мицелиальные нити (гифы) делятся в продольном и поперечном направлениях, в результате чего образуется паренхиматозная масса клеток, представляющая собой спорангии (рис. 46, А). У представителей родов Geodermatophilus и Dermatophilus стадия в виде нитевидного мицелия в цикле развития отсутствует или рудиментарна. В состав рода Frankia входят виды с хорошо развитым нитевидным мицелием, спорангии формируются только из части клеток нити в виде интеркалярных или терминальных "опухолей". При распаде спорангиев из них освобождаются подвижные или неподвижные споры. Воздушный мицелий отсутствует.
Все представители группы — хемоорганогетеротрофы с высокими пищевыми потребностями, аэробы (главным образом микроаэрофилы), мезофилы. Представители рода Frankia развиваются в качестве эндосимбионтов в корневых клубеньках небобовых растений. Для клубеньков показана способность фиксировать молекулярный азот. Бактерии этой группы распространены в почве, воде, обитают на кожных покровах млекопитающих.
Группа 28. Актинопланеты. В группу объединены актиномицеты, характеризующиеся приспособленностью к обитанию в водной среде, имеющие подвижную стадию в течение жизненного цикла. В процессе роста образуют развитый, разделенный на перегородки, устойчивый субстратный мицелий, иногда также и воздушный. Для представителей этой группы характерно формирование спорангиев разной формы, возвышающихся над поверхностью субстрата, внутри которых образуются споры (рис. 46, В). Форма спорангиев, количество и расположение в них спор различны; споры также неодинаковой формы. Эти признаки легли в основу классификации актинопланет на роды.
У отнесенных к этой же группе представителей рода Micromonospora спорангии отсутствуют (рис. 46, В). Неподвижные одиночные споры располагаются непосредственно на гифах мицелия или на очень коротких спороносных гифах (спорофорах). Все актиномицеты, входящие в состав группы, — аэробные хе-моорганогетеротрофы, сапрофиты или факультативные паразиты. Основные места обитания: пресная вода, почва, мертвые растительные и животные остатки.
|
| Рис. 47. Актиномицеты рода Streptomyces (А), Microbispora (Б) и Streptosporangium (В). Обозначения см. рис. 46 (по Lechevalier, Pramer, 1971) |
Группа 29. Стрептомицеты и родственные формы. Актиномицеты образуют хорошо развитый воздушный мицелий, который в процессе последующего цикла развития не распадается на фрагменты (рис. 47, А). Размножение спорами, формирующимися на концах гиф, или кусочками вегетативного мицелия. Актиномицеты, объединяемые в эту группу — облигатно аэробные хемоорганогетеротрофы. Основной род Streptomyces насчитывает около 500 видов, для которых характерно образование на воздушном мицелии прямых или спирально закрученных цепочек, состоящих из трех или более неподвижных спор. Многие стрептомицеты синтезируют антибиотики, активные против бактерий, грибов, водорослей, простейших, фагов, обладающие также противоопухолевым действием.
Группа 30. Мадуромицеты33. Аэробные актиномицеты, формирующие развитый субстратный мицелий, на котором споры никогда не формируются. Они образуются только на воздушных гифах, дифференцирующихся или в короткие цепочки спор, или в спорангии, содержащие одну или множество спор (рис. 47, Б, В). Группа недостаточно изучена и, по мнению специалистов, нуждается в значительной ревизии.
33 Madura — название провинции в Индии, где впервые был описан один из представителей этой группы.
Группа 31. Термомоноспоры и родственные формы. Представители этой группы формируют воздушный мицелий, на гифах которого образуются подвижные или неподвижные споры, одиночные или в виде цепочек. Спорангии у большинства представителей отсутствуют. Для актиномицетов типового рода Thermomonospora характерна способность расти в температурном диапазоне от 40 до 48°.
Группа 32. Термоактиномицеты. Объединяет 1 род термофильных актиномицетов. Недавние исследования обнаружили, что споры Thermoactinomyces относятся к типичным эндоспорам, и по этому признаку организм следует отнести к бациллам. На близость к последним указывают также данные анализа 16S рРНК. Однако подобно истинным актиномицетам бактерии этой группы образуют хорошо развитый мицелий и по морфологии напоминают представителей рода Thermomonospora, что в данное время позволяет рассматривать их вместе с другими актиномицетами. Все представители группы формируют хорошо развитый субстратный и воздушный мицелий. Аэробные хемоорганогетеротрофы. Основное место обитания — почвы, воды, разлагающиеся растительные остатки.
Группа 33. Другие формы актиномицетов. В последние годы описано несколько новых актиномицетов, выделенных в отдельные роды, которые помещены в эту группу, так как еще недостаточно изучены и отсутствует информация, необходимая для выявления степени их сходства с другими актиномицетами.
М.В.Гусев, Л.А.Минеева © 1992-2001
Кафедра клеточной физиологии и иммунологии биологического факультета МГУ им. М.В.Ломоносова © 2000-2001
Страница обновлена 09.09.01
Оформление: А.В.Киташов