Переходные формы: как их нашли и не нашли.
Василий Томсинский
Copyright © 12 ноября 2012 г.
Последняя редакция 21 января 2013 г.
Содержание.
Часть 1. Переходы и переходные формы.
Археоптерикс и происхождение птиц.
Часть 2. Креационисты vs. переходные формы.
5. Трудности с эволюционными отношениями.
6. Где миллионы переходных форм?.
7. Их нет: признают даже эволюционисты.
Приложение 1. Эволюция перьев.
Приложение 2. Эволюция дыхательной системы.
Приложение 3. Сравнение анатомии дейнонихозавров, ранних и современных птиц.
Существование переходных форм – одно из самых захватывающих (если не самое захватывающее) свидетельств в пользу эволюции, в пользу вывода о том, что все живое связано одной общей родословной. Другие свидетельства вряд ли могут на равных тягаться с переходными формами. Атавизмы очень редки. Рудименты – части тела, существование которых не всегда очевидно, тем более у других животных. Гомология кажется просто сходством. В эмбрионах и компетентный биолог не всегда отыщет след эволюции, равно как и сами образцы. Молекулярное единство жизни – просто набор слов без точных знаний о внутриклеточных процессах. Понять значение генных деревьев трудно без знаний генетики, математики и систематики. Функции белков и их структуру в бытовых условиях не изучишь.
Иное дело переходные формы. Удивительно, что орел – это результат длительной модификации чего-то вроде ящерицы. Но вот окаменевшие кости животного, которое находится на промежуточной стадии этой модификации. Наша окаменелость – своего рода фотография процесса, который у нас нет никаких шансов наблюдать непосредственно. Преобразование ящерицы в птицу происходило более 200-80 миллионов лет назад. В то время как человечество изобрело письменность всего несколько тысяч лет назад, да и сам наш вид существует всего 200,000 лет. И тем не менее мы достаточно удачливы, чтобы иметь на руках реальную часть того эволюционного процесса. Как если бы мы могли отправиться в прошлое и поймать представителя промежуточной эволюционной группы. Мы можем изучить окаменелость по фотографиям, мы можем посмотреть на саму окаменелость через музейное стекло, мы можем сделать костям рентген, мы можем создать реконструкцию скелета, и даже можем попытаться реконструировать тело.
Переходные формы также не оставляют никаких сомнений насчет невозможности эволюции. Критики эволюционной теории говорят, что превращение ящерицы в птицу немыслимо, потому что оно потребовало бы кардинальной перестройки всех систем органов, появления сложнейшего механизма полета, возникновения удивительных инстинктов, что кажется невозможным. Однако переходные формы показывают, что это фактически произошло. Знаем мы, как это в точности случилось, или нет; кажется ли нам это невероятным или нет; можем ли мы в это поверить или нет – не имеет никакого значения. Эволюция произошла, как не крути.
Я подчеркнул ценность переходных форм лишь для того, чтобы теперь показать, что их иллюстративность слишком переоценивается. Они убедительны для специалистов. Для обычных людей существование переходных форм в большей мере вопрос веры, поскольку они не в состоянии оценить, почему те являются переходными. Это очень важное обстоятельство, которое популяризаторы науки почти всегда игнорируют. Есть несколько проблем с пониманием переходных форм.
Во-первых, по окаменелостям исследовать эволюцию беспозвоночных или растений существенно сложнее, чем эволюцию позвоночных. А небиологи главным образом не в состоянии сравнить строение беспозвоночных или растений. Следовательно, приходится ограничиться только позвоночными животными. Но в результате возникают подозрения такого рода:
"Если эволюционисты вообще ссылаются на свидетельства окаменелостей, то лишь для некоторых групп позвоночных организмов, так как рыбы, земноводные, пресмыкающиеся и млекопитающие образуют определенную последовательность в отложениях". |
| Уильям Гладсхауэр. "Как возник наш мир" [124]. |
Во-вторых, переходные формы представлены окаменевшими останками. Находки полностью сохранившихся скелетов - редкость. Отдельные кости сохраняются частично и могут накрывать друг друга. Требуется специальная подготовка и скрупулезная работа для анализа экземпляра. Поэтому фотографии окаменелостей не могут сказать почти ничего обычному человеку с улицы. Скорее всего, они произведут на него удручающее впечатление. Можно продемонстрировать реконструкцию скелета и надеяться, что читатель разберется в его строении сам. Другой вариант (самый распространенный) – нарисовать переходные формы такими, какими они могли бы быть при жизни. Правда, фантастичность таких картин довольно очевидна, и вряд ли они что-то демонстрируют. Вся очевидность их промежуточного эволюционного положения оказывается скрытой под нарисованными покровами.
В-третьих, – и это главная проблема, – неспециалист не знает анатомию и просто не в состоянии заметить, что перед ним переходная форма. Требуется несколько секунд, чтобы показать человеку окаменелость или ее реконструкцию, и затем нужно потратить не меньше часа на лекцию об анатомии скелета разных групп позвоночных животных. Причем необходимо, чтобы слушатель запомнил материал и мог в нем самостоятельно ориентироваться. Кто без такой лекции сможет сказать, сколько позвонков должно быть в шейном отделе рептилий и птиц, какая форма позвонков должна быть у каждой группы?
Именно поэтому Кен Миллер, блестящий полемист, решив доказать существование переходных форм, смог лишь показать несколько иллюстраций внешне похожих скелетов и описать работу палеонтологов [53]. Антрополог Джим Фоли, обсуждая ископаемых гоминид, сконцентрировался на двуногом передвижении и размере мозга. Палеонтолог Дональд Протеро, описывая многочисленные переходные формы, обычно ограничивается указанием двух-трех особенностей их анатомии. (Однако, на мой взгляд, он преуспел больше других в популяризации палеонтологии и эволюции, и я настоятельно рекомендую его замечательные книги всем, кто интересуется данной темой) [64, 65, 66].
Археоптерикс – пример переходной формы №1. "Берлинский экземпляр археоптерикса – самая важная находка палеонтологии", – сказал как-то палеонтолог Джон Остром. Но все же популярные издания описывают археоптерикса неизменно однобоко. Из птичьих особенностей называются оперение, а из рептилийных – когти, зубы и длинный хвост. Множество других анатомических черт, более важных, остаются неупомянутыми. Здесь нечему удивляться. Подробный анатомический обзор сделает описание нечитабельным. Но в то же время скупость популярных описаний производит ложное впечатление. Люди думают, что больше об археоптериксе и сказать-то нечего, и приходят к заключению, что палеонтологи делают поспешные выводы на основе столь скудных данных.
Подводя итог, можно сказать: не важно, существуют переходные формы или нет, так как даже если они существуют, вы не сможете это продемонстрировать без длительного курса палеонтологии и анатомии. И популярные описания эволюции, называя некоторых вымерших животных и объявляя их переходными, не достигают поставленной цели. Мало того, они еще и создают искаженное впечатление об эволюционной биологии.
В этой статье я попытаюсь закрыть пробел между популярными книгами и профессиональной палеонтологией, на материале скелетной анатомии рептилий и птиц.
Часть 1. Переходы и переходные формы.
Археоптерикс и происхождение птиц.
Существование переходных форм является необходимым следствием теории единой общей родословной всех форм жизни, которая составляет основу эволюционной биологии. В соответствии с этой теорией, какие бы формы жизни мы не взяли в качестве примера (при условии, что одна форма не является прародителем второй), они будут потомками общего предка, который существовал в некоторый момент в прошлом. Степень родства и соответственно древность общего предка может значительно изменяться. Так оценено, что человек разумный и шимпанзе связаны весьма близким родством. Общий предок этих видов жил примерно 6 миллионов лет назад. Человек разумный и неандерталец связаны еще более близким родством. Их прародитель существовал, согласно самой точной оценке, около 700,000 лет назад. По контрасту человек и сизый голубь имеют гораздо более дальнее родство. Их генеалогические линии разошлись около 300 миллионов лет назад. Графическим представлением теории единой общей родословной является всем известное эволюционное древо или в более современном варианте - кладограмма.
Теория единой общей родословной предполагает, что происхождение новых биологических видов, родов, семейств, отрядов и т. д. происходит путем преобразования (эволюции) биологии (в том числе строения) предка, которому они обязаны своим появлением. Например, птицы возникли в результате преобразования биологии наследственной для них группы рептилий. А значит, должны существовать формы, которые находились на различных стадиях этого преобразования. Однако среди ныне живущих организмов таких найти проблематично. В этом нет ничего удивительного. Мы знаем, что многие группы организмов существуют на Земле очень давно. Например, птицы живут на планете не менее 150 миллионов лет, с Юрского периода, если вы предпочитаете относительную хронологию. Стало быть, преобразование строения некой группы древних рептилий, которое в конечном итоге привело к появлению птиц, происходило в те далекие времена.
Учитывая сказанное, теперь мы можем определить, что же такое "переходные формы". Переходные формы - это такие формы жизни, которые демонстрируют преобразование одного уникального типа строения, характерного только для одной группы организмов (родительской), в другой уникальный тип, характерный для другой группы (потомковой). Таким образом, переходная форма должна обладать комплексом уникальных черт строения одновременно и предковой, и потомковой групп. Если теория единой общей родословной верна, переходные формы обязательно должны были существовать. И мы можем попытаться отыскать их в окаменелостях.
В этом разделе я опишу археоптерикса и покажу, что он занимает промежуточное положение между мезозойскими рептилиями и птицами, причем анатомически он даже ближе к рептилиям, чем к птицам. Но следует сразу оговориться, что это будет лишь частичное и упрощенное описание. Я буду использовать только наиболее простые признаки. Палеонтологи в своих исследованиях стараются учесть любую мелочь, поэтому их работа обычно включает длинные перечни и простых, и тончайших различий в анатомии. Однако включение подробностей такого рода здесь сделает текст нечитабельным. И хотя наиболее сложные различия останутся за скобками, следует помнить, что они есть, и все последующее описание не дает полной картины.
Для наглядности я предлагаю читателю преобразовать его собственный скелет в скелет археоптерикса. Мы можем это себе позволить благодаря гомологии. Широко распространено заблуждение, что гомология - это сходство в строении. В действительности гомология имеет более существенное основание. В гомологичных структурах составляющие их элементы расположены по типовому образцу и имеют типичную структуру. Рассмотрите в качестве иллюстрации гомологию костей черепа в трех группах динозавров.
Чтобы понять гомологию, стоит вспомнить Дарвина. Биологи задолго до Дарвина отмечали, что у разных и несхожих организмов образец строения органов и их расположения совпадает. Но почему это так, никто удовлетворительно объяснить не мог. Распространенность гомологии подталкивала разных исследователей к мысли о том, что в некоторых случаях она является результатом происхождения от общего предка. Однако именно Дарвин придал этому аргументу современную форму. Суть идеи в том, что новые биологические черты организмов возникают путем постепенного преобразования старых. С этой точки зрения, например, организм человека – это модифицированный организм рыбы. Даже когда появляются новые элементы, например, новые кости, они возникают из преобразования старых структур. Именно так возникает гомология. Скелет всех четвероногих животных – это модификация базового образца строения древних амфибий и рыб. То же верно для прочих систем организма – нервной, кровеносной, выделительной и т. д. И чем ближе родство, тем больше гомологическое сходство.
Здесь я поместил для сравнения рисунки скелетов археоптерикса и голубя. Если вы вернетесь к ним после сравнения анатомии археоптерикса, его ближайших родственников и птиц, уверен: схемы покажутся более информативными. Можно также рассмотреть интерактивную схему скелета голубя, а также подборку трехмерных моделей скелетов разных птиц.
|
|
| Археоптерикс | Голубь |
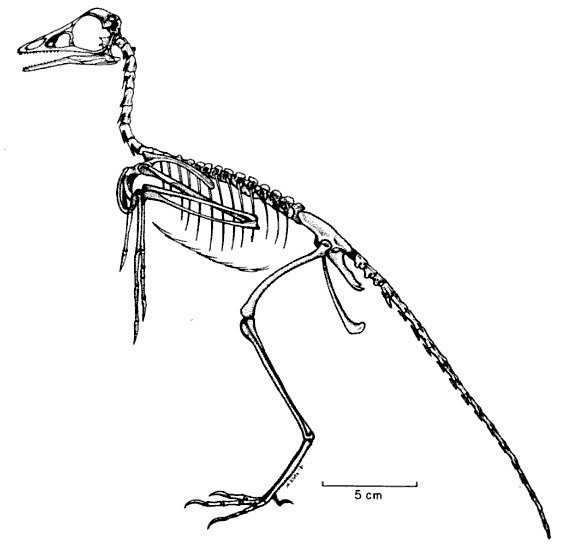
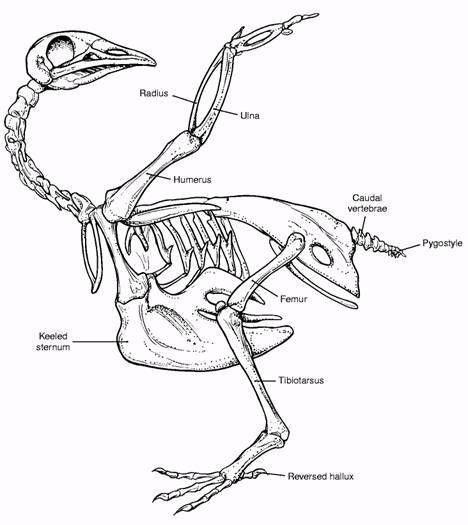
Итак, читатель, сделаем из Вас гордую мезозойскую птицу.
Чтобы стать ею для начала Вам придется перестать быть млекопитающим. У Вас три косточки в среднем ухе и одна кость в нижней челюсти, а нужно, чтобы в нижней челюсти было несколько костей и одна слуховая в среднем ухе. Форма черепа совершенно неудовлетворительна, он слишком овальный. Нельзя стать археоптериксом без грубо треугольного, вытянутого черепа. Но главное все же – проделать перед глазницей еще парочку отверстий. Особенно гордость всех архозавров – предглазничное окно и ближе к ноздрям скромное предглазничное отверстие. Чуть выше добавим утерянную предлобную кость. За глазницей тоже нужно поработать. Туда придется приладить еще одну потерю – заглазничную кость.
|
| Черепа голубя (A) и археоптерикса (B). Показаны не все кости. |
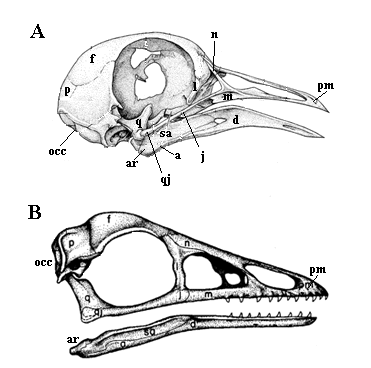
Небные кости явно не в порядке. Как минимум нужно добавить то, что называется наружной крыловидной костью.
У Вас есть зубы – это хорошо, но они слишком разнообразны, целых четыре типа. Археоптериксу такие излишества ни к чему. Отныне у Вас будут однообразные острые конические зубы. А между зубами расположатся межзубные пластины – маленькие костистые выросты на челюстях. И хотя Вам теперь будет сложнее пережевывать пищу, зато зубы могут сменять друг друга неограниченное число раз – на месте старого зуба неизменно будет вырастать новый.
Далее. Голова соединяется с шеей не в том месте. Чтобы получить право называться археоптериксом, придется оторвать голову, поместить большое затылочное отверстие сзади черепа, а не на дно, и снова соединить голову с шеей.
Далее. Суставные поверхности ваших шейных позвонков плоские. Это эстетика млекопитающих, в мезозое для нее еще не пришло время – придется поменять на двояковогнутые.
Далее. В крестцовом отделе позвоночника у Вас 5 позвонков. Именно столько и надо, чтобы быть археоптериксом. Но если Вам уже исполнилось 25 лет, то все ваши пять позвонков срослись в единое тело. Так не пойдет. Придется позвонки разъединить.
Далее. Кости таза. С такими лобковыми костями вам ничего не светит в мире археоптериксов. У вас они срастаются, но и только-то. Придется их вытянуть, отдаленные концы снабдить расширением в форме ботинка и уже тогда срастить отдаленные концы.
Далее. Смущает отсутствие хвоста. У вас есть несколько деградировавших позвонков, сросшихся в единое образование (копчик). Такого археоптерикса никто уважать не будет. Хвост должен быть длинным из двадцати цельных позвонков. Чем ближе к кончику хвоста, тем более вытянутыми должны становиться тела позвонков.
Далее. Передние конечности. С точки зрения археоптерикса, совершенно не ясно, зачем Вам столько пальцев. Оптимальное число – 3. Так что отбрасываем безымянный и мизинец. Придется также пересчитать количество фаланг пальцев. У вас по три фаланги в каждом из оставшихся пальцев. Это никуда не годится. В большом пальце две фаланге, в указательном – три, это правильно, а вот к безымянному одну нужно прибавить. Еще мы добавим вам изогнутые когти. Можете считать это компенсацией. Кроме того, ваше запястье довольно однообразно. Вам просто необходимо срастить первую и вторую кости запястья в единое образование и придать ему полулунную форму. Эта штука, называемая полулунной костью запястья, позволяет сворачивать кисть по-птичьи.
И не забудем про лопатки. У вас к лопатке прирастает еще одна кость – коракоид или вороновидная кость, образующая с лопаткой единое тело. Археоптериксу полагается иметь эту кость отдельной, самостоятельной и хорошо развитой, грубо квадратной формы. Причем этот коракоид должен иметь выступающий вперед отросток. Что же до лопаток, то они должны расширяться в отдаленном от плечевого сустава конце.
Далее. Ваши ключицы нуждаются в немедленном исправлении. Необходимо срастить их концы в единую кость – вилочку. И пусть грудина разделяется на отдельные части.
Далее. От бывшего млекопитающего мало что осталось. Не будем придираться к ребрам. Но добавим вам брюшные ребра – типичный признак пресмыкающихся. Теперь даже искушенный глаз не узнает в вас заурядного млекопитающего.
Далее. Ваши задние конечности практически не нуждаются в изменениях. Хотя одного пальца придется лишиться. А фаланги второго пальца придется растянуть так, чтобы этот палец стал самым длинным и загнутым кверху. Затем мы приделаем к нему хищный серповидный коготь. Кроме того, таранная кость должна иметь отличительный восходящий отросток.
Почти все. Осталось сделать несколько общих изменений.
Во-первых, у вас слишком тяжелые кости. Они явно нуждаются в пустотах, заполненных воздухом. От этого скелет станет только легче.
Во-вторых, вам пригодится оперение. Асимметричные перья на передних конечностях, пучок длинных перьев на конце хвоста и контурные перья тела.
Теперь вы очень похожи на археоптерикса, но в целом выглядите вы чудовищно. Чтобы стать археоптериксом, нужно еще уменьшиться в размерах. 50 сантиметров от кончика морды до кончика хвоста, 25 сантиметров в высоту. Это самое большее, что вы можете себе позволить.
Теперь, когда у нас есть археоптерикс, мы можем начать модифицировать его в птицу. Попутно я буду сравнивать археоптерикса с его ближайшими родственниками – дейнонихозаврами – группой звероногих динозавров. Вы сможете сами оценить, является ли строение археоптерикса в большей мере птичьим или рептилийным. Начнем быструю эволюцию, на которую у природы ушло около 80 миллионов лет.
Зубы и межзубные пластины. Все современные птицы являются беззубыми, межзубные пластины также отсутствуют. Зубы закладываются в эмбрионах птиц, но на более поздней стадии рассасываются. С помощью манипуляций с эмбриональными тканями ученые даже смогли получить цыплят с зубами. Эти данные показывают, что генетические схемы, ответственные за развитие зубов, все еще присутствуют в геноме птиц. У ближайших родственников археоптерикса зубы были. Как мы теперь знаем, потеря зубов у птиц происходила в разных эволюционных линиях независимо и в разное время. Некоторые птицы утратили зубы в начале мелового периода. Последние зубатые птицы вымерли 60 миллионов лет спустя. Роговой чехол, покрывающий челюсти – клюв – очевидная птичья особенность. Но ее придется добавить, так как археоптерикс и дейнонихозавры не имели такого чехла.
Череп нуждается во множественных модификациях. В отличие от археоптерикса и дейнонихозавров у птиц утрачены предлобная, заднеглазничная и наружная крыловидная кости, но предглазничное окно остается.
Характерная особенность черепа птиц - подвижность и верхней, и нижней челюсти, что контрастирует с типичным состоянием у большинства четвероногих животных (включая человека), у которых подвижна только нижняя челюсть. Механику челюстей археоптерикса установить непросто, но анатомия верхних челюстей несовместима с тем типом подвижности, который типичен для современных птиц.
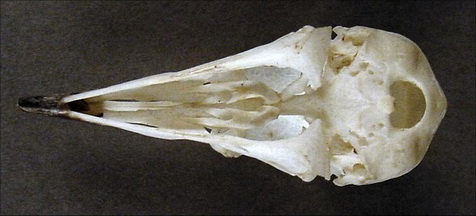
Большое затылочное отверстие на том же месте, что у дейнонихозавров, но не птиц. У птиц оно смещено на дно черепа.
|
| Череп голубя, вид снизу. Большое затылочное отверстие хорошо видимо. http://www.skullsite.com/completelist/index.htm |
Шейные позвонки археоптерикса с их двояковогнутыми суставными поверхностями такие же, как у ближайших родственников. По контрасту шейные позвонки птиц имеют седловидные суставные поверхности. Это предмет особой гордости птиц, они уникальны. Ни у каких других животных нет таких позвонков, и никогда не было. Исключение составляют цератозавры – группа триасовых звероногих динозавров. Однако у них эта особенность была не столь выражена, как у птиц. Благодаря этой уникальной адаптации птицы могут поворачивать голову на 180 градусов, а некоторые на 270.
|
| Шейные позвонки птиц. Позвонки с такими суставными поверхностями называются гетероцельными, то есть разновогнутыми. |
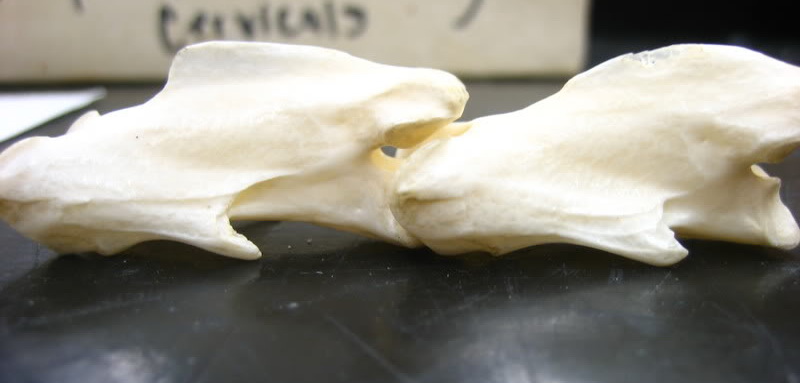
У археоптерикса и дейнонихозавров крестцовый отдел позвоночника насчитывает то же количество свободных позвонков. У птиц этот отдел сильно изменен. Число позвонков в нем увеличено (колеблется от 11 до 23), позвонки срастаются между собой в так называемый сложный крестец. Также у птиц тазовые кости срастаются между собой. Напротив, лобковые кости у птиц не срастаются, а их отдаленные концы лишены расширения в виде ботинка.
|
| Тазовые кости, вид сбоку. Слева направо ранний динозавр Целофиз - юрский динозавр аллозавр - дейнонихозавр велоцираптор - археоптерикс - голубь. Лобковая кость обозначена "Pubis" и выделена черным. Ischion - седалищная кость. Над лобковой и седалищной костями располагается подвздошная кость. Видимое отверстие, сформированное этими тремя костями, - это вертлужная впадина, в которую входит головка бедренной кости. |
|
| Часть скелета современной птицы. Две тонкие косточки, отходящие от места сочленения бедра с тазом - это лобковые кости. Можно ясно видеть, что их отдаленные концы не срастаются. Synsacrum - сросшийся крестец, pygostyle - пигостиль. |
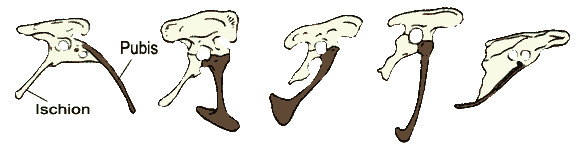

Хвостовой отдел у птиц редуцирован – хвостовые позвонки упрощены, срастаются между собой и в целом хвостовой отдел сливается со сложным крестцом. Заканчивается хвост пластинкой – пигостиль – это важнейшая адаптация. Пигостиль служит для прикрепления рулевых перьев, таким образом, он обеспечивает совершенство птичьего полета. Пресловутый длинный хвост археоптерикса – полная противоположность состоянию хвостового отдела у птиц. В популярной литературе он систематически упоминается неправильно. Дело не в длине. Длина сама по себе ничего не значит. В природе избыток хвостов любой длины. Но у археоптерикса хвостовые позвонки не срастаются и не уменьшаются, чем ближе к кончику хвоста, тем более вытянутыми, цилиндрическими становятся тела позвонков. Выглядит это так, как будто археоптерикс украл свой хвост у дейнонихозавров.
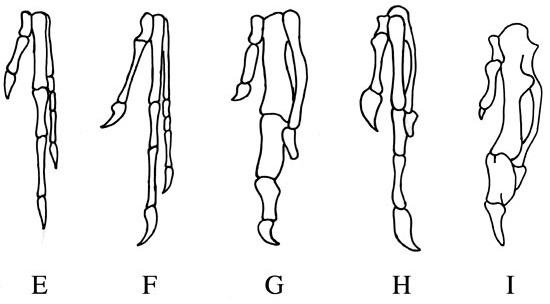
По строению передних конечностей дейнонихозавры приняли бы археоптерикса за своего. Те же три пальца, с такими же когтями, то же число фаланг. Но у птиц передняя конечность устроена иначе. Археоптерикс, дейнонихозавры и птицы, все трехпалы. Четвероногие животные (кроме самых ранних и самых древних) первично имеют пять пальцев. Иное число пальцев в разных группах четвероногих связано с утратой того или иного их числа. В эмбрионах птиц все еще можно отыскать след их происхождения от древних пятипалых предков. В течение эмбрионального развития у птиц закладываются пять пальцев, но два в итоге рассасываются. Три остающихся пальца редуцированы, присутствует лишь несколько видоизмененных фаланг, служащих для закрепления маховых перьев. Число фаланг у птиц 1-2-1 в противоположность 2-3-4 у археоптерикса и дейнонихозавров.
Кроме того, у птиц кости пястья и передний ряд костей запястья срастаются в единую пястнозапястную кость. Если вам не нравится такое название, вы можете называть ее пряжкой. Это тоже будет правильно. У дейнонихозавров все кости запястья и пястья свободны, но об археоптериксе такого не скажешь. У него кости пясти не срастаются, но сильно сдавлены, таким образом, пряжка еще не сформирована, но начало процессу положено.
Полулунная кость запястья, которая есть у археоптерикса - это, скорее, птичий признак, но птицы разделяют эту особенность не только с археоптериксом, но и с дейнонихозаврами, у которых она является самостоятельным образованием. У птиц полулунная кость входит в состав пряжки. Она позволяет сворачивать кисть так, что та оказывается почти параллельна предплечью.
|
| Сравнение строения кисти. E динозавр Орнитолестес, F археоптерикс, G птица мелового периода Синорнис, H Гоацин, I курица |
Подобно своим динозавровым родственникам археоптерикс не имел надкаракоидного сухожилия. Эта важная адаптация позволяет птицам поднять крыло выше спины, тем самым птица может сделать мощный взмах, необходимый для машущего полета. Не ясно, был ли археоптерикс способен к машущему полету. С одной стороны, отсутствие надкоракоидного сухожилия показывает, что поднять крыло выше спины он не мог, а с другой – какая-то примитивная форма машущего полета, не свойственная современным птицам, у него все же могла быть.
Лопатка археоптерикса, как и у дейнонихозавров, в отдаленном конце имеет расширение. У птиц такого расширения нет. Суставная ямка лопатки у археоптерикса занимает промежуточное боковое положение. У птиц она смещена в спинном направлении. А связанный с лопаткой коракоид по образцу птиц имеет выступающий отросток. У дейнонихозавров этой особенности нет. В соответствующем положении на коракоиде у них присутствует бугристость, которая рассматривается палеонтологами как предшественник выступающего отростка. Сам корокоид археоптерикса устроен по образцу дейнонихозавров, а не птиц. Он плоский и имеет грубо квадратную форму. У птиц коракоид вытянут и принимает форму распорки.
Птицы также отличаются и от археоптерикса, и от дейнонихозавров наличием киля – выроста грудины, служащего для прикрепления летательной мускулатуры.
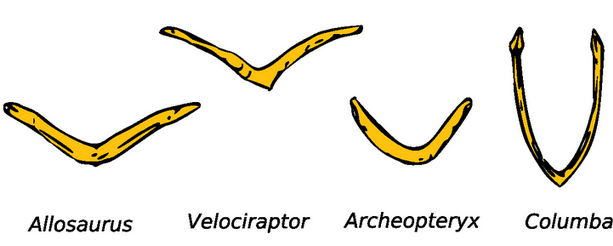
Ключицы, сросшиеся в вилочку – еще один признак, который роднит птиц, археоптерикса и дейнонихозавров. А ребра разделяют группы. Брюшные ребра есть у археоптерикса, его родственников и множества рептилий, у птиц их нет. Грудные ребра птиц скрепляются между собой выростами – крючковатыми отростками. Их нет у археоптерикса, хотя они есть у некоторых дейнонихозавров.
|
| Вилочка у звероногих динозавров, археоптерикса и голубя. Слева направо аллозавр, велоцираптор, археоптерикс, голубь. |
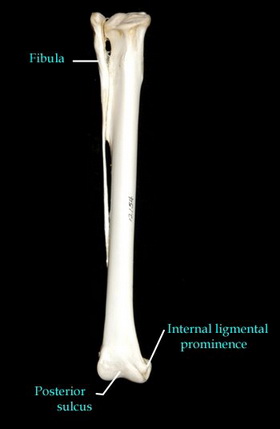
По строению нижних конечностей археоптерикс с его удлиненным вторым пальцем, украшенным хищным серповидным когтем, присоединяется к компании дейнонихозавров. В конце концов, именно из-за этого когтя они получили свое имя – "ужасный коготь". Археоптерикс также разделяет с дейнонихозаврами строение голени и стопы. Стопа и голень птиц перестроены. Прежде всего, у птиц редуцирована малая берцовая кость. Она уменьшилась до тонкой щепы и приросла к большой берцовой. У археоптерикса малая берцовая кость полностью развита. Ближе к стопе – у птиц пяточная и таранная кости срастаются друг с другом и с большой берцовой. Археоптерикс эту характеристику также не разделяет. Более того, таранная кость археоптерикса снабжена развитым восходящим отростком. Птицы эту характеристику утратили. Она обнаруживается в течении эмбрионального развития, но исчезает на более поздних стадиях. Здесь анатомия археоптерикса снова соответствует типу строения ближайших родственников, но не современных птиц. Птицы обзавелись цевкой – костью, образованной срастанием части костей плюсны и предплюсны. У археоптерикса цевки нет, но, как и в случае с пряжкой, соответствующие кости сдавлены. Таким образом, цевка находится в стадии формирования.
|
|
| Большая берцовая кость и малая берцовая кость лысого орла. Fibula - малая берцовая кость. http://www.royalbcmuseum.bc.ca/Natural_History/Bones/atlas/atlas.htm | Цевка беркута. http://www.royalbcmuseum.bc.ca/Natural_History/Bones/Species-Pages/GOEA.htm#Tarsus |
Оперение – то, что отличает птиц и археоптерикса от всех ныне живущих животных, но не от дейнонихозавров. Последние также обладали оперением. Причем некоторые имели асимметричные перья (асимметрия пера – необходимое условие для полета).
Как легко заметить, у археоптерикса нет почти ничего специфически птичьего. Его анатомия в гораздо большей степени рептилийная, чем птичья. В скелете археоптерикса сочетаются одновременно черты дейнонихозавров и птиц, поэтому он представляет собой мост между дейнонихозаврами и более поздними птицами, которые в свою очередь промежуточны по сравнению с современными птицами. Если бы мы сделали этот вывод 40 лет назад, могли бы претендовать на открытие. К сожалению, нас опередил палеонтолог Джон Остром. Он предпринял скрупулезный обзор анатомии археоптерикса и всех потенциально родственных групп животных. В итоге Остром заключил, что археоптерикс по существу – маленький оперенный динозавр.
Следующая диаграмма показывает числовое соотношение рептилийных и птичьих особенностей анатомии у ранних птиц и наиболее близких к ним динозавров. Список сравниваемых особенностей см. в приложении 3.
|
Динозавры: Гаплохейрус, Анхиорнис, Ксиотингия, Каудиптерикс, Микрораптор, Эпидендрозавр, Эпидексиптерикс. Базальные птицы: Археоптерикс, Джехолорнис, Эоконфуциусорнис, Сапеорнис, Конфуциусорнис, Янорнис |
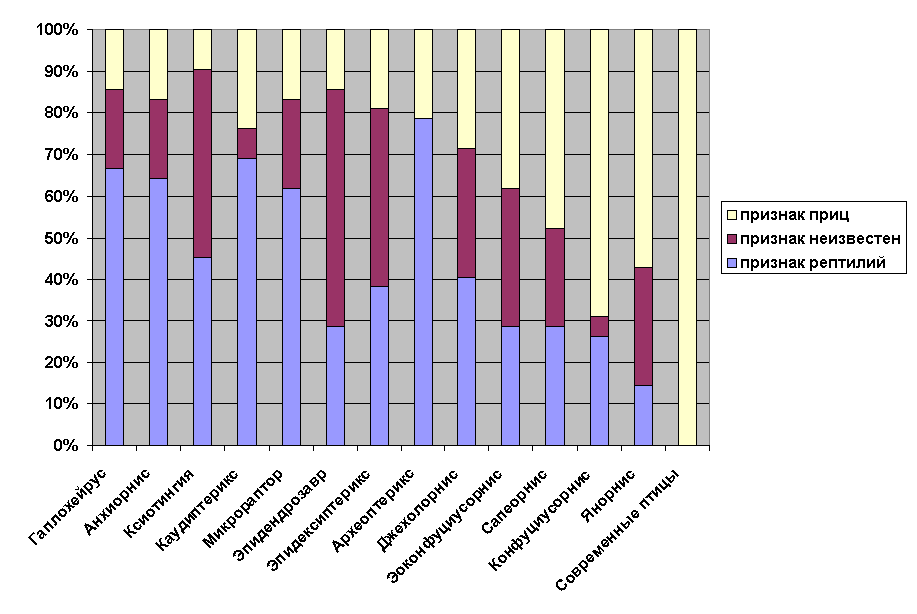
Я говорил, что представленное сравнение археоптерикса и его родственников является частичным и упрощенным. Для составления диаграммы я использовал 42 ключевых признака, наиболее простых и понятных. Палеонтологи обычно используют в своих исследованиях от 100 до 300 признаков. При этом они сравнивают значительно большее количество представителей и используют алгоритмы, основанные на теории вероятностей. Я надеюсь, что это даст некоторое представления об уровне упрощения, на котором нам приходится говорить об изучении переходных форм (см. также примечание a).
Как видно из диаграммы, археоптерикс действительно может быть описан как оперенный динозавр. Кроме того, он не одинок в своем роде. И если археоптерикс стоит ближе к динозаврам, чем к птицам, то другие формы, например Эоконфуциусорнис, находятся где-то на полпути.
Таким образом, единство в скелетной анатомии и существование ряда переходных форм указывают на происхождение птиц от одной из линий звероногих динозавров. Но кости не одиноки. Есть ряд других, несвязанных данных, подтверждающих этот вывод. Действительно, если птицы ведут свою родословную от общего предка с дейнонихозаврами, то мы должны обнаружить единство в аспектах биологии между птицами и близкими к ним звероногими. В то же время, биология птиц и звероногих должна контрастировать с биологией других групп животных. Все, что нам остается сделать, сравнить биологию птиц и звероногих в тех аспектах, которые нам известны.
Размер тела.
Способность к полету накладывает сильное ограничение на размер тела. Неслучайно птицы, утратившие способность к полету, могут приобретать очень большой размер тела (например, страусы и моа).
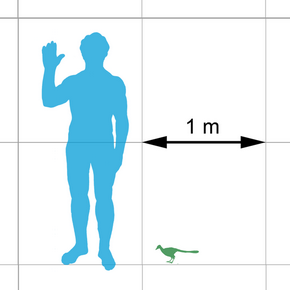
По контрасту, те дейнонихозавры, анатомия которых в прошлом служила эталоном для сравнения с птицами – дейноних и велоцираптор – крупные животные. Физически невозможно, чтобы животные такого размера могли освоить полет. Поэтому долгое время приходилось предполагать, что более близкие родственники птиц были существенно меньше в размере. Предположение было подтверждено с открытием микрораптора, который был описан в 2000 году как "самый маленький из известных динозавров". Девять лет спустя был описан анхиорнис – дейнонихозавр еще меньшего размера. Фактически, анхиорнис даже меньше микрораптора. Анализ скелетной анатомии показывает, что анхиорнис и микрораптор стоят ближе других к птицам. На сегодняшний день они остаются самыми маленькими и легкими из всех динозавров [39, 80, 98, 99].
 |
 |
| Археоптерикс | Анхиорнис |
 |
 |
| Микрораптор | Дейноних |
Мозг.
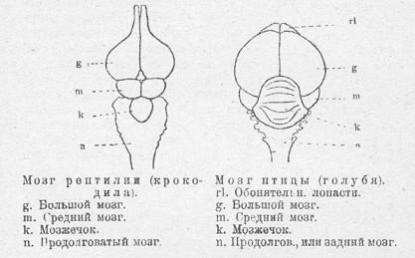
В данном случае обнаруживаются еще более детальные различия. Мозг современных птиц значительно отличается от мозга рептилий. Полушария головного мозга у птиц сильно увеличены. Также чрезвычайно увеличились мозжечок и зрительные доли. Что, конечно, связано со способностью к полету. По контрасту обонятельные луковицы сократились в размерах.
|
См. также сравнение строения мозга крокодила, археоптерикса, позднемеловой птицы и голубя.
|
Мозг не окаменевает. Означает ли это, что мы никогда не сможем ничего узнать о мозге вымерших животных? Не обязательно. Головной мозг окружен твердой мозговой оболочкой, которая отражает строение мозга. И если череп хорошо сохранен, есть возможность создать (физический или предпочтительно компьютерный) слепок мозга.
Мозг большинства звероногих динозавров, чьи черепа позволяют реконструкцию, в целом устроен по типичному для археозавров образцу. Но эта картина начинает изменяться, когда мы обращаемся к ближайшим родственникам птиц. В отличие от других звероногих, их мозг ближе к птичьему образцу: относительный размер больших полушарий, мозжечка и зрительных долей увеличивается, однако, не достигает характерных для птиц соотношений. Обонятельные луковицы, напротив, остаются развитыми или даже увеличиваются. Сокращение этих частей мозга произошло позднее. Действительно, обонятельные луковицы археоптерикса все еще соответствуют рептилийному, а не птичьему образцу. В целом, мозг археоптерикса характеризуется промежуточным состоянием между рептилийным и птичьим, его <мозжечок и большие полушария увеличены, но не столь существенно, как у современных птиц [1, 31, 43, 45, 60].
??
Размножение.
Птицы, как и большинство рептилий, размножаются, откладывая яйца. Форма яиц отличается в различных группах. Яйца крокодилов овальные. У черепах – овальные или шарообразные, у большинства динозавров – шарообразные. Яйца птиц асимметричны – один полюс яйца шире другого. Яйца звероногих, близкородственных птицам, также асимметричны.
Еще более важная характеристика яиц – микроструктура скорлупы. Микроструктура птичьих яиц существенно отличается от микроструктуры рептилий, включая крокодилов, завроподов и птицетазовых динозавров. Однако она практически идентична у птиц и близкородственных звероногих!
Черепахи, змеи, крокодилы кладут яйца однократно. То же справедливо в отношении динозавров. Мы знаем это, поскольку яйца в кладках динозавров лежат беспорядочно единой группой. По контрасту птицы откладывают не все яйца разом, а последовательно через промежутки времени, что связано с ограничением в размерах тела. Изучая сохранившиеся кладки близких родственников дейнонихозавров и птиц, – овирапторидов – палеонтологи обратили внимание, что в их гнездах яйца располагаются не в общей массе, а объединены в пары. Такое пространственное расположение интерпретируется как признак того, что самки овирапторидов откладывали по два яйца через некоторое время. Гипотеза подтверждается находкой овирапторида только с двумя яйцами в полости тела.
|
Кладка овираптора. Википедия. |
.jpg)
Название овирапторидов восходит к описанию в 1924 году динозавра, останки которого были обнаружены поверх гнезда. Палеонтологи заключили, что это был хищник, разорявший чужие гнезда. Таким образом, динозавр получил имя овираптор – крадущий яйца. Однако открытие новых экземпляров, заставило пересмотреть вывод. Были описаны экземпляры овирапторидов, сидевших в центре гнезда в типично птичьей позе и накрывавших яйца передними конечностями. Выяснилось также, что подобное поведение было характерно и для некоторых дейнонихозавров. Неизвестно, с чем именно связано такое поведение. Одно очевидное объяснение связывает присутствие родителя с охраной кладки, другое – с буквальным высиживанием яиц. Независимо от того, какое из этих предположений правильное, данные показывают, что близкие родственники птиц заботились о своих кладках так же, как и современные птицы. Возможно, даже согревали яйца теплом своего тела. Такое предположение не кажется невероятным, если вспомнить, что близкие родственники птиц (овирапториды здесь примыкают к дейнонихозаврам) имели оперение [5, 8, 35, 38, 42, 44, 103, 104].
Перья.
Поскольку в оперении археоптерикса присутствуют перья с типично птичьей структурой, было ясно, что археоптерикс мог унаследовать их от предков (см. приложение 1). Предположение подтвердилось только на рубеже XX и XXI веков, когда впервые были описаны овирапториды с сохраненными перьями. На сегодняшний день перья известны в трех наиболее близких к птицам семействах звероногих и не в каких других. Описаны звероногие с симметричными и асимметричными перьями Ннапомню, асимметрия – необходимое аэродиномическое условие для полета. В общих чертах оперение близких родственников птиц соответствует птичьему: пучок длинных перьев на кончике хвоста (рулевые перья у птиц), длинные и крупные перья на передних конечностях (маховые перья у птиц) и контурные перья тела [41, 56, 78, 81, 93, 94, 118].
Поведение.
В 2005 году палеонтологи представили миру экземпляр хорошо сохранившегося дейнонихозавра, у которого голова была повернута назад, а морда частично накрыта передней конечностью. Аналогичное положение тело можно наблюдать у современных птиц во время сна. Голова у птиц, как предплечья и голени у человека, именно та часть тела, которая обеспечивает наибольшую потерю тепла и соответственно охлаждение тела. Поэтому птицы типично во время сна накрывают голову крылом, что уменьшает теплопотерю. Предположительно, упомянутый динозавр спал в момент внезапной смерти [101].
|
|
Реконструкция положения тела Mei Long. Художник James Gurney. Использование только одной задней конечности для опоры во время сна характерно для современных птиц. Эта деталь реконструкции опирается на данные о связи птиц и дейнонихозавров и не может быть проверена на основе изучения известных окаменелостей. |
Спящий попугай. |


Перечисленные здесь особенности указывают на единство биологии одной ветви звероногих динозавров и птиц. Однако обратите внимание, что проведенное сопоставление не имеет отношения к переходности или непереходности археоптерикса и прочих форм, о которых шла речь выше. Переходный статус зависит от анатомии. Сравнение биологии птиц и родственных звероногих преследует другие цели. Оно демонстрирует:
Во-первых, интегральный характер изучения эволюции, предполагающий привлечение информации из разных областей биологии.
Во-вторых, как на основе выделения переходных форм можно выдвигать гипотезы, в поддержку которых постепенно накапливаются данные.
"Точные предки тираннозавров невыяснены". Палеонтолог Роберт Кэррол в 1989 году [128].
Чтобы искать предков какой-либо группы животных, сначала нужно установить диагностические черты, присущие только ее представителям. Тираннозавриды – типичные хищные звероногие динозавры. Это часто крупные животные, передвигавшиеся на двух задних конечностях. Их зубы крупные, зазубренные и загнутые назад. Эти зубы не оставляют сомнений на счет кулинарных пристрастий. Они имели большие головы и большие хвосты, служившие противовесом для головы и туловища. Но чем они отличались от других звероногих и своих ближайших родственников? Правильная классификация окаменелостей – сложная задача, и вы сейчас в этом убедитесь, поскольку нам придется углубиться в еще более сложные тонкости скелетной анатомии.
У тираннозаврид:
Носовые кости срастались друг с другом.
Теменные кости также срастались, образуя стреловидный гребень.
Зубы, связанные с межчелюстной костью обладают U-образной формой.
Сама межчелюстная кость имеет большое боковое отверстие.
Основная клиновидная кость имеет большую подмыщелковую полость.
Надугловая кость снабжена выраженным прямым гребнем.
Досочленовный отросток укорочен.
Скуловая кость частично пневматизирована и имеет окно.
Чешуйчатая кость в задней части имеет большое отверстие.
Отверстие в задней части нижней челюсти значительно уменьшено.
Верхняя затылочная кость не формирует большое затылочное отверстие.
Поперечные отростки шейных позвонков вытянуты. Они служат для прикрепления мощных мышц, удерживающих большую голову.
Передние конечности чрезвычайно маленькие относительно задних. Функция столь маленьких конечностей не выявлена до сих пор.
Конец лучевой кости сглажен.
Присутствуют только два пальца.
Второй палец передней конечности существенно больше.
Лобковая кость заканчивается очень большим расширением в форме ботинка (около 50% длинны всей кости).
Подвздошная кость примерно равна в длине бедру.
Третья кость в плюсне сжата второй и четвертой. Такая структура называется арктометатарсус. Арктометатарсус – адаптация к бегу.
|
|
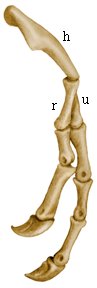
Передняя конечность тираннозавр. Обозначения. h – плечевая кость, r – лучевая кость, u – локтевая кость. |
Арктометатарзус у тираннозавра рекса. Википедия. |
.JPG)
Если вы обескуражены этим панегириком анатомии, утешайтесь тем, что это еще упрощенный список.
Теперь, когда мы определили отличительные особенности тираннозаврид, можно искать переходные формы к ним. Но самое интересное, что мы можем частично предсказать, на что будет похожа такая форма. Если мы знаем ближайших родственников тираннозавров, значит, сравнивая анатомию всех связанных групп, можно вывести черты, которые разделили их предковые формы. Если мы правильно определили родство, то переоценка уже описанных окаменелостей и/или обнаружение новых подтвердят гипотезу. В противном случае они опровергнут построенную схему эволюционных отношений. В сущности, именно так выдвигаются и проверяются эволюционные гипотезы.
Итак, что мы можем предположить с уверенностью о еще не найденных переходных формах? Здесь пригодятся пояснения. В интересующей нас группе звероногих динозавров выделяют две подгруппы – карнозавров и целурозавров. Ранее к карнозаврам относили всех звероногих большого размера, а к целурозаврам – среднего и маленького размеров. Но с течением времени выяснилось, что огромные тираннозавриды анатомически принадлежат в большей мере к целурозаврам, чем к карнозаврам. Пришлось гипотезу об отличиях в размерах тела отбросить. Позднее выяснилось, что представители нескольких семейств целурозавров независимо увеличивались в размерах тела. Таким образом, целурозавры изначально маленькие животные. Это приводит к неизбежному заключению, что предок гигантского тираннозавра должен иметь скромный размер.
Строение передних конечностей предковой формы также можно без труда предвидеть. Тираннозавриды в анатомии передних конечностей сильно отклонились от своих родственников. Комбинируя информацию из разных семейств целурозавров, приходится заключить, что переходная форма к типичным тираннозавридам должна быть трехпалой, кости передних конечностей должны быть длинными и хорошо развитыми. Мы можем даже указать число фаланг пальцев. Это то же число, что у археоптерикса, дейнонихозавров и большинства других целурозавров – 2-3-4.
Арктометатарсус – еще одна особенность, которую мы не ожидаем встретить у предка тираннозаврид. Арктометатарсус известен в разных группах, но он не является, насколько мы знаем, наследственной особенностью целурозавров, так что мы не ожидаем его увидеть у ранних тираннозаврид.
Можно попытаться спрогнозировать и некоторые другие особенности, но это потребует более сложного анализа. Остановимся на достигнутом.
Так известны ли нам предки для семейства динозавров, которое захватило внимание даже далеких от палеонтологии людей? Тираннозавр был впервые описан в 1905 году, а его самый старый и хорошо сохранившийся на сегодняшний день предок – 101 год спустя. Останки динозавра, получившего имя гуанлонг, были раскопаны в нулевых годах XXI века на западе пустыни Гоби. Возраст отложений, в которых были найдены кости, составляет 160 миллионов лет, поздний юрский период. Анализ скелета со всей очевидностью показывает, что гуанлонг занимает промежуточное положение между ранними целурозаврами и тираннозавридами мелового периода. Он также подтверждает нашу эволюционную гипотезу о том, какими были предки тираннозаврид [90].
Гуанлонг в сравнении с более поздними тираннозаврами – небольшое животное, длинна тела ок. 3 метров. Кости передних конечностей не редуцированы, так что его передние конечности хорошо развитые и длинные (ок. 60% длинны задних). Три развитых пальца присутствуют, число фаланг соответствует типичному для целурозавров – 2-3-4. Второй палец не увеличен относительно других. Интересно, присутствует полулунная кость запястья. Ни у одного другого тираннозаврида во взрослом состоянии полулунная кость не обнаруживается в связи с редукцией передних конечностей.
Есть несколько других особенностей, которые не встречаются у тираннозаврид. Например, нижнечелюстное отверстие гуанлонга было большим в размере, у него также присутствовало тазовое отверстие.
Чтобы не усложнять описание, я сведу все особенности в таблицу.
|
Признак |
Тираннозавриды |
Гуанлонг |
|
Сращение носовых костей |
+ |
+ |
|
Сращение теменных костей |
+ |
+ |
|
Зубы, связанные с межчелюстной костью U-образной формы |
+ |
+ |
|
Межчелюстная кость имеет большое боковое отверстие. |
+ |
+ |
|
Основная клиновидная кость имеет большую подмыщелковую полость. |
+ |
+ |
|
Надугловая кость снабжена выраженным прямым гребнем |
+ |
- |
|
Досочленовный отросток укорочен. |
+ |
+ |
|
Скуловая кость частично пневматизирована и имеет окно. |
+ |
+ |
|
Чешуйчатая кость в задней части имеет большое отверстие. |
+ |
- |
|
Отверстие в задней части нижней челюсти значительно уменьшено. |
+ |
- |
|
Верхняя затылочная кость не формирует большое затылочное отверстие. |
+ |
+ |
|
Поперечные отростки шейных позвонков вытянуты. |
+ |
+ |
|
Передние конечности чрезвычайно маленькие относительно задних. |
+ |
- |
|
Конец лучевой кости сглажен. |
+ |
+ |
|
Присутствуют только два пальца. |
+ |
- |
|
Второй палец передней конечности существенно больше. |
+ |
- |
|
Лобковая кость заканчивается очень большим расширением в форме ботинка (ок. 50% длинны всей кости). |
+ |
+ |
|
Нет тазовых отверстий |
+ |
- |
|
Подвздошная кость примерно равна в длине бедру. |
+ |
+ |
|
Арктометатарсус присутствует |
+ |
- |
Три особенности не попали в таблицу, так как их нельзя обозначить как "есть или нет". Они сами находятся в промежуточном состоянии. Это 1) Относительный размер межчелюстных зубов, 2) выступ на чешуйчатой и квадратно-скуловой костях, 3) положение подвздошных костей. У гуанлонга подвздошные кости сближаются относительно средней линии тела, но не входят в контакт, как у других тираннозаврид.
|
|
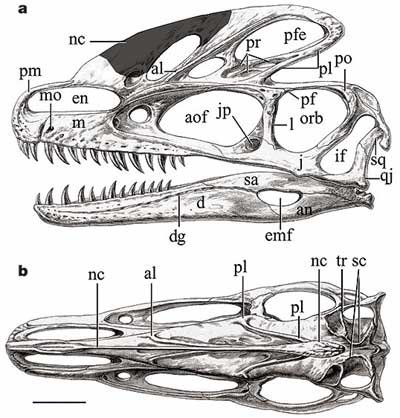
Череп гуанлонга. a. вид сбоку, b. вид сверху. Основные обозначения: an. Угловая кость, aof. Предглазничное отверстие, d. зубная кость, emf. внешнее нижнечелюстное отверстие, en. Внешняя ноздря, j. скуловая кость, jp. Отверстие в скуловой кости, l. слезная кость, m. Верхняя челюсть, nc. Носовой гребень, orb, Глазница, pm, Межчелюстная кость, qj. квадратно-скуловая кость, sa. надугловая кость, sc. Стреловидный гребень, sq. чешуйчатая кость. Черта масштаба 5 см. |
Череп тираннозавра. Показаны не все зубы. |

Гуанлонг не одинок в своем роде. Целурозавры подразделяются на несколько семейств, эволюционные отношения между которыми было очень трудно проследить из-за недостатка таких гуанлонгов. Но с течением времени искомые формы появились. Они подтвердили одни эволюционные гипотезы и опровергли другие. В 2009 китайские и американские палеонтологи подробно описали аналогичную гуанлонгу форму, на сей раз из семейства тех самых овирапторидов, охранявших свои гнезда. Динозавр получил имя инцизивозавр. Один абзац этого описания стоит процитировать полностью:
"Не считая известных пернатых динозавров, в джехольских отложениях были найдены бесчисленные переходные формы. Эти динозавры (например, лиаоцератопс, основные дейнонихозавры микрораптор и синовенатор, основной овирапторозавр каудиптерикс, переходной тираннозаврид дилонг, переходной теризинозаврид бэйпяозавр, основной орнитомимозавр шенцоузавр и ранний гадрозаврид Jinzhousaurus) обладают множеством особенностей, которые помещают их в основе признанных эволюционных групп, и поэтому они имеют важнейшее значение для разработки точных филогенетических деревьев. Сказанное также верно для инцизивозавра, который последовательно размещается в основе древа овирапторазавров". |
| Balanoff A. et al. "Cranial osteology of the theropod dinosaur Incisivosaurus gauthieri (Theropoda: Oviraptorosauria)" [3]. Внутренние ссылки опущены. |
Заинтересованным читателям я предлагаю самостоятельно изучить анатомию упомянутых динозавров, а также признаки их семейств, создать сравнительные таблицы и установить самим, являются ли перечисленные животные переходными формами или нет. Впрочем, эта работа уже проделана на профессиональном уровне. Палеонтологи, используя теорию вероятностей, вычислили эволюционные отношения между целурозаврами, определяя степень неслучайного совпадения анатомических черт.
|
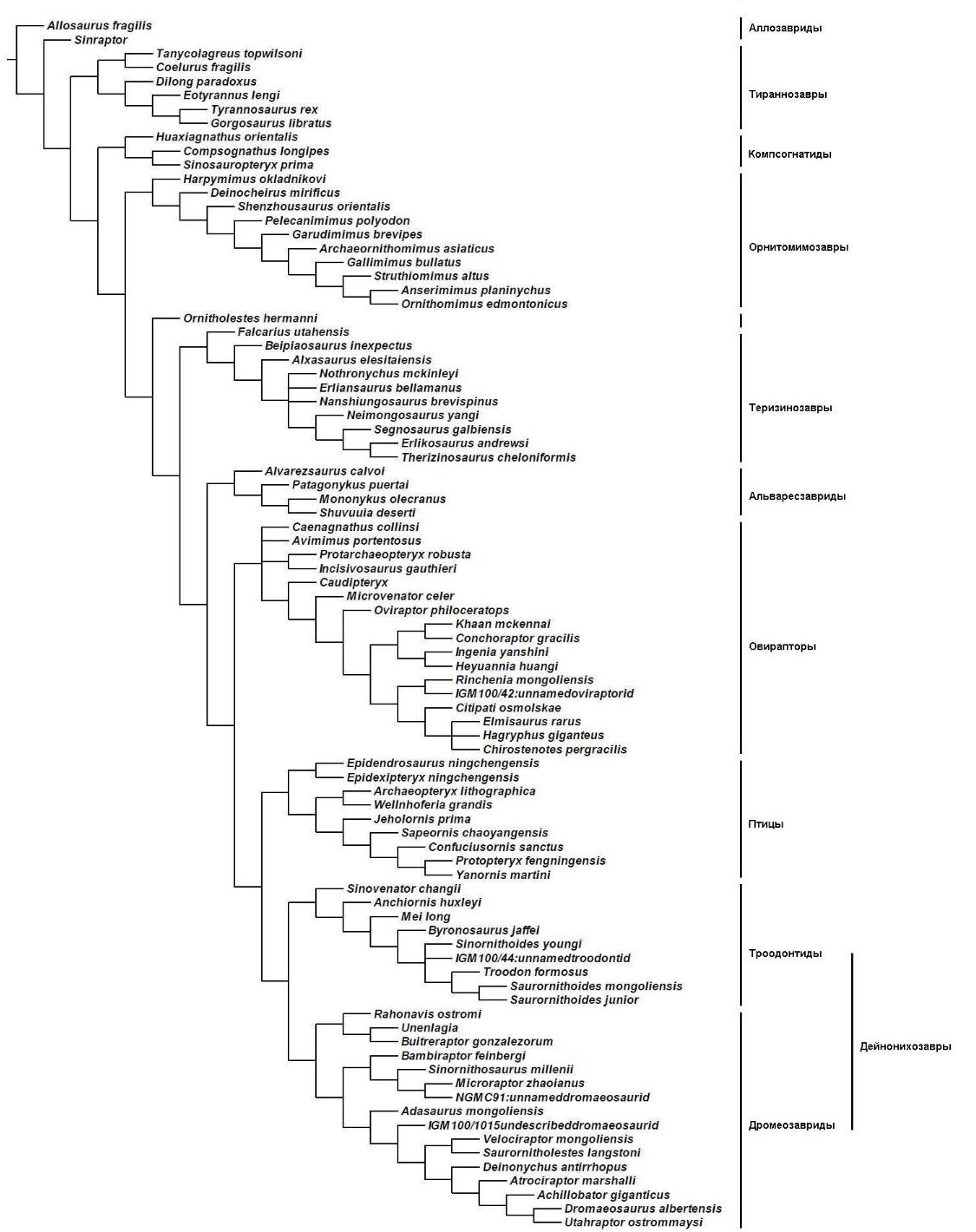
Схема (кладограмма) эволюционных отношений целурозавров. Из упоминавшихся выше форм присутствуют: микрораптор, синовенатор, археоптерикс, каудиптерикс, дилонг, бейпяозавр, инцизивозавр. |
Но, может быть, я просто пытаюсь выдать желаемое за действительное? Правда ли, что все переходные формы среди целурозавров (и вообще среди позвоночных животных) уже найдены, и палеонтологам осталось лишь вставлять новые роды в установленную схему? Нет. Хотя некоторые эволюционные отношения среди целурозавров, как думают, наконец-то надежно установлены, пробелы в нашем знании не исчезают все сразу. Связи между поздними (меловой период) целурозаврами стали более или менее ясными. Но ранняя эволюция целурозавров плохо изучена. Как сегодня считается (и это еще одна гипотеза), корни известных семейств следует искать в отложениях не позднее середины юрского периода. Примечательно, что до настоящего времени палеонтологам известно сравнительно немного хорошо сохранившихся динозавров этого временного интервала. Если где-то существует формация, хранящая изобилие экземпляров динозавров середины Юрского периода, то пока ее никто не обнаружил. Остается лишь надеяться на подобное открытие. Но палеонтология – "медленная" наука.
Также можно указать трудности другого рода. Из приведенной выше кладограммы следует, что семейство альваресзаврид ближе связано с семейством теризинозаврид. Альваресзавриды – очень специализированные динозавры, своей анатомией бросившие вызов фантазии палеонтологов. Кто может быть их родственником – палеонтологическая загадка, в сравнении с которой меркнут даже дзенские коаны. С кем только альваресзаврид не роднили. В них видели даже птиц, утративших способность к полету – мезозойских страусов. Пока что переходная форма от примитивных целурозавров к альваресзавридам не найдена. Когда она будет найдена (а никто из палеонтологов не сомневается в такой возможности), можно будет проверить, правильно ли палеонтологи оценили совпадения анатомических черт альваресзаврид и теризинозаврид, действительно ли они – двоюродные братья.
Более того, представленная выше схема эволюционных отношений также является гипотезой. Она неизбежно должна подвергаться проверке по мере описания новых окаменелостей и переоценки старых.
UPD: Искомый альваресзаврид был описан во время подготовки этой статьи. Я не стал переписывать первоначальный текст для иллюстрации того, как могут устаревать данные. Динозавр гаплочериус частично подтвердил связь альваресзаврид с теризинозавридами. Поскольку гаплочериус один из самых ранних целурозавров, я также добавил его в таблицу для сравнения с близкими родственниками птиц [14].
Часть 2. Креационисты vs. переходные формы.
Если существование переходных форм – самое лучшее свидетельство в пользу эволюционной теории, то это также самая большая проблема для креационистов. Десятилетия назад они решили, что если эволюция фактически имела место, значит, христианство полностью ложно. Если существуют переходные формы, то не существует Бога, который послал сына своего единородного на смерть, дабы ранами его мы исцелились. Это очень странное богословие. На самом деле, как существование бога может зависеть от строения передней конечности археоптерикса? И тем не менее, креационисты настаивают именно на таком, единственно верном, по их мнению, богословии. Креационизм был отвергнут множеством богословов и множеством христианских церквей, а также представителями других религий. Однако креационисты создали мощное социальное движение, которое способствовало распространению их богословия на глобальном уровне. Креационисты неколебимо убеждены в своей правоте. Они оправдывают это тем, что Бог через священные тексты открыл им истину, а Бог просто не может ошибаться. Отсюда с неизбежностью следует, что не могло быть никакой эволюции, а значит и переходных форм. Но как быть с примерами переходных форм, которые описаны палеонтологами?
Креационисты могут не соглашаться по богословским вопросам, они могут принадлежать к разным конфессиям и даже к разным религиям. Иногда одни креационисты отрицают некоторые аргументы против эволюции других креационистов. Однако все они едины в отрицании теории эволюции. По их словам, в пользу теории эволюции нет абсолютно никаких свидетельств. В том числе нет никаких переходных форм.
"В окаменелостях не обнаружено никаких переходных форм". |
Генри Моррис. "Сотворение и современный христианин" [131]. Курсив в оригинале. |
"...Но ... переходных звеньев в останках нет". |
Генри Моррис. "Библейские основы современной науки" [130]. |
"Никаких переходных форм от одной крупной группы живых организмов к другой вообще не было найдено". |
"Жизнь - как она возникла? Путем эволюции или путем сотворения?" [126] |
"...все без исключения классы, отряды и семейства появляются в отложениях внезапно, без каких-либо предшествующих промежуточных или переходных форм". |
Уильям Гладсхауэр. "Как возник наш мир" [124]. |
"Пропавшие звенья все еще остаются пропавшими". |
| Томас Хайнц. "Творение или эволюция" [139]. |
"...если эволюция действительно имела отношение к возникновению этих уникальных и хорошо приспособленных существ, то должно было быть множество очевидных переходных форм, но ни одно такое животное не найдено... Все то, что сохранилось в течение истории существования жизни, являет нам знаменательное отсутствие переходных форм, наличия которых требует теория эволюции. " |
| Дуэйн Гиш. "Сотворение мира, эволюция и исторические свидетельства." [123] |
"Кроме археоптерикса в летописи окаменелостей нет ничего, что хотя бы позволило предполагать существование убедительной связи между животными различных типов". |
Сильвия Бейкер. "Камень преткновения: Верна ли теория эволюции?" [121] |
"Полная неудача интенсивных поисков палеонтологов в течение более чем столетия показала, что промежуточные формы в природе никогда не существовали." |
| Сергей Вертьянов. "Общая биология" [122]. |
"Эволюционистам по-прежнему трудно найти объяснение тому, что все окаменелости неизменно разбиваются на основные группы, а огромные пробелы так и остаются незаполненными." |
| Ариэль Рос. "В начале..." [135] |
Научные труды полны предположений о возможном и, конечно же, несомненном, существовании в прошлом переходных форм. Часто эти предположения — не более чем следствие игры воображения в сочетании с несколькими неубедительными свидетельствами. |
Дж. Маклин и др. "Очевидность сотворения мира". [129] |
"Опираясь на неисчислимые миллионы исследованных ископаемых, ученые на сегодняшний день каталогизировали свыше 250 000 видов ископаемых животных, которые выставлены в музеях всего мира. И среди них невозможно встретить ни одного неоспоримого промежуточного звена." |
Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
"Если эволюция протекала в течение миллионов лет путем последовательности незначительных изменений, логично было бы предположить, что в природе должно быть больше переходных форм, чем статичных классов. Однако на сегодняшний день их не обнаружено вовсе! ... В окаменелостях не обнаружено ... ни одной переходной формы." |
Дэвид Роузвер. "Наука о сотворении мира, доказывающая правоту Библии" [125]. |
Теперь я опишу несколько способов, при помощи которых креационисты избавляются от переходных форм, и проанализирую их. Креационистские аргументы включают.
1. Одно из двух.
2. Мозаичные формы.
3. Подделки.
4. Временные парадоксы.
5. Трудности с эволюционными отношениями.
6. Где миллионы переходных форм?
7. Их нет: признают даже эволюционисты.
Первый способ состоит просто в отрицании переходной природы представленной формы. Креационисты определяют, согласуясь с необъясняемыми ими методами, к какой именно группе относится форма, а затем подчеркивают одни черты и полностью игнорируют другие. Так археоптерикса они считают птицей.
Археоптерикс был самой настоящей птицей, а никакой не переходной формой. |
Дэвид Роузвер. "Наука о сотворении мира, доказывающая правоту Библии" [125]. |
"...в ее лице [археоптерикс] мы имеем не промежуточный вид, а вполне нормальную птицу". |
Уильям Гладсхауэр. "Как возник наш мир" [124]. |
Если же говорить об археоптериксе - так называемой промежуточной форме... Это была не полуптица, это была настоящая птица. А переходные формы с недокрыльями и полуперьями так и не были найдены. |
| Дуэйн Гиш. "Сотворение мира, эволюция и исторические свидетельства." [123] |
"...крылья археоптерикса были полностью развитыми и функциональными". |
Пол Тейлор. "Сотворение. Иллюстрированная книга ответов" [138]. |
"Археоптерикс обладал практически всеми признаками птицы". |
| Сергей Вертьянов. "Общая биология" [122]. |
"Это существо имеет некоторые признаки рептилии, оставаясь впрочем совершенно нормальной птицей. О рептилии напоминают зубы по бокам клюва и хвост из нескольких позвонков... Но крылья и перья уже полностью птичьи... Нет оснований считать археоптерикса чем-то промежуточным". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
Соответственно креационисты перечисляют все птичьи особенности и пропускают все рептилийные особенности, как будто их и нет. Для этого им не приходится слишком стараться. Как уже говорилось, популярные описания археоптерикса очень скудны, и креационисты не стремятся выйти за эти описания. Зубы, когти и длинный хвост. Когти есть у некоторых современных птиц, – говорят креационисты – зубы были у некоторых вымерших, а хвост – это хвост.
Удивительна ирония ситуации. На заре становления современного креационного движения (примерно 60-ые годы XX века), когда креационисты решили считать археоптерикса птицей, эта точка зрения была более правдоподобной, чем сегодня. С течением времени список птичьих особенностей археоптерикса неуклонно сокращался. Происходило это по двум причинам. Во-первых, выяснялось, что некоторые птичьи особенности, вроде противопоставленного большого пальца задних конечностей, у археоптерикса диагностировались по ошибке. Во-вторых, птичьи особенности постепенно обнаруживались у ближайших родственников археоптерикса. На данный момент, у археоптерикса фактически нет ни одной сугубо птичьей особенности, которой не было бы у его родственников. И как мы видели выше, строение его скелета в гораздо большей степени соответствует рептилийному. Из составленной мной диаграмы следует Составленная мной диаграмма показывает, что к птичьим чертам у археоптерикса можно отнести около 20%, но даже эти черты археоптерикс разделяет с родственными динозаврами. Если бы креационисты говорили, что археоптерикс – рептилия и не имеет никакого отношения к птицам, это было бы гораздо правдоподобнее. Но их читателя не могут этого заметить. Все, что есть у них в голове – зубы, когти и длинный хвост.
"Поэтому многие ученые в пылу фантазии наделяют археоптерикса большим количеством признаков пресмыкающегося, чем он их имеет. Так называемые признаки пресмыкающихся у археоптерикса заключаются в наличии когтей на крыльях, зубов, позвонкового хвоста и маленькой грудины. Но это не признаки настоящей переходной формы, потому что они все еще встречаются и у птиц!". |
| Уильям Гладсхауэр. "Как возник наш мир" [124]. |
Здесь креационист Гладсхауэр допускает явную грубую ошибку. У современных птиц зубы не встречаются.
Признаки рептилии, которые находят у археоптерикса - это когтеподобные окончания крыльев, наличие зубов и позвонки, которые выдаются из хвоста. Считается, что он был посредственным летуном, так как у него был маленький киль на грудине... Современные птицы не имеют зубов, но у некоторых древних птиц, несомненных птиц, зубы были..." |
| Дуэйн Гиш. "Сотворение мира, эволюция и исторические свидетельства." [123] |
В процитированной статье Гиш сообщает ошибочные и вводящие в заблуждения сведения. Киля у археоптерикса не было вовсе. Оцените также расплывчатые формулировки, которые использует Гиш, чтобы приуменьшить значение других нетипичных для птиц характеристик - "когтеподобные окончания" и "позвонки, которые выдаются из хвоста". Гиш почему-то не в состоянии недвусмысленно признать, что археоптерикс обладал полноценными когтями. Креационисты (и Гиш не исключение) выдвигают на первый план сформированность перьев археоптерикса, но когда речь заходит о когтях, желание расписать что-то во всех подробностях улетучивается. Это тем более странно, потому что Гиш пишет о существовании когтей у современных птиц. Гиш не в состоянии признать, что все элементы хвоста археоптерикса в деталях строения имеют рептилийную анатомию. Более того, он хотя и упоминает хвост археоптерикса, но оказывается не в состоянии объяснить его строения, и просто обходит его молчанием! Однако самое важное, что Гиш полностью игнорирует все остальные многочисленные черты строения, свидетельствующие о переходном статусе археоптерикса.
"Еще один пример предполагаемого утраченного звена - окаменелые отпечатки животного, названного археоптериксом... Большинство эволюционистов утверждает, что это существо является связующим звеном между птицами и рептилиями. Окаменелости показывают, что археоптерикс имел перья, крылья и клюв, как у птицы. Однако у этого ископаемого были признаки, которые заставили палеонтологов предположить его родство с рептилиями - зубы на клюве и когти на крыльях". |
| Дж. Маклин и др. "Очевидность сотворения мира". [129] |
Archaeopteryx обладает некоторыми чертами рептилии, такими, как зубы, длинный хвост, когти на крыльях и костная структура, напоминающая в определенных аспектах рептильную. Есть у него и птичьи признаки, например, полностью развитые перья, вилочка (грудная кость) и цепкий большой палец. Часть основных рептильных характеристик Archaeopteryx присуща не одним только рептилиям. У целого ряда ископаемых птиц есть зубы, а когти на крыльях встречаются и у некоторых современных птиц. |
| Ариэль Рос. "В начале..." [135] Внутренние ссылки опущены. |
Однако креационисты не могут изменить аргументы и назвать археоптерикса рептилией. В этом случае им пришлось бы объяснять своим сторонникам, как птица стала рептилией, и почему они столько лет твердили о том, что у новоявленной рептилии "совершенное во всех отношениях оперение". Поэтому археоптерикс у креационистов остается птицей и, надо полагать, останется в дальнейшем.
Чтобы отклонить эволюционное родство птиц с дейнонихозаврами, креационисты очень часто цитируют палеоорнитолога Алана Федучию и заимствуют у него аргументы. Федучия – орнитолог, много работавший над эволюцией птиц. Он предлагает альтернативную гипотезу о родстве птиц, которая, правда, пока не получила подтверждений. И даже притом что Федучия критикуется подавляющим большинством своих коллег, креационисты широко цитируют его как эксперта и предпочитают его мнение мнению всех остальных. Когда Федучию спросили, что он думает об использовании его работ креационистами, тот ответил:
"Креационисты исказят любые предложенные им аргументы, и они поставили меня в один ряд с такими светилами, как Стивен Джей Гулд, поэтому меня это нисколько не беспокоит. археоптерикс - наполовину рептилия, наполовину птица, как не сними колоду, так что он - Розетский камень эволюции, независимо от того, связан ли он родством с динозаврами или нет. Эти креационисты путают мелкие детали эволюции с неоспоримым фактом эволюции. Животные и растения изменялись...". |
| Алан Федучия. Интервью [59]. |
Трудно подобрать более точное сравнение. Креационисты, рассматривая любую переходную форму, пытаются отнести ее к одной группе, подчеркнув соответствующие особенности и игнорируя другие. Что это если не попытка так снять колоду, чтобы переходной статус формы исчез?
Не все креационистские обзоры археоптерикса и эволюции птиц одинаковы. Наиболее подробные рассмотрения принадлежат американцу Дуэйну Гишу, который больше всех прочих креационистов занимался окаменелостями, пытаясь доказать отсутствие переходных форм, и немцам Юнкеру и Шереру. Гиш написал по этой теме две книги. В упоминаемом обзоре, Гиш настаивает на том, что археоптерикс все-таки птица. Он собирает все данные в поддержку этой идеи, которые только может найти. Примечательно, Гиш собирает множество информации, главным образом уклоняясь от рассмотрения анатомии. Он упоминает лишь несколько особенностей скелета археоптерикса, все из которых неоднозначны и долгое время были дискуссионными. Но в то же время, Гиш оставляет без внимания все бесспорные рептилийные особенности, будто их вовсе нет. Между тем, многие из них были известны 50 и 100 лет назад. Повторюсь, эта давняя работа Гиша – одна из лучших, изданных до настоящего времени креационистами. Любопытно, что это не единственный пример, когда Гиш выделял одни особенности и обходил молчанием другие. Более подробно см [27]. Аналогичную тактику используют Юнкер и Шерер. Обсуждая ту же тему, они сваливают в кучу массу несвязанных данных, которые просто не имеют отношения к сути вопроса. Но даже их, вероятно, лучшее рассмотрение археоптерикса остается очень поверхностным (я предлагаю читателю обратиться к тексту их книги и убедиться в этом самостоятельно).
Многие анатомические характеристики переходных форм настолько очевидно свидетельствуют об их промежуточном статусе, что лучшее, что могут сделать креационисты, - попросту игнорировать такие особенности. Прекрасный пример этой нехитрой аргументации можно найти у австралийского креациониста Джонатана Сарфати. В своей книге Сарфати перечисляет птичьи характеристики археоптерикса, в том числе ценой явных ошибок, и просто умалчивает обо всех рептилийных чертах. Вот что он пишет:
"Археоптерикс обладал полностью сформированными маховыми перьями (в том числе асимметричными бородками и особыми бороздками на вентральной стороне, усиливающими перо, как и у современных летающих птиц), классическими эллиптическими крыльями, как лесные птицы наших дней, и вилочкой, к которой прикреплялись мышцы, опускающие крыло. Его мозг был в точности таким же, как у летающих птиц, с развитым мозжечком и зрительной корой. Тот факт, что у археоптерикса были зубы, не имеет никакого отношения к его статусу "переходной формы" - у многих вымерших птиц были зубы, а у многих современных рептилий их нет. Кроме того, археоптериксу, как и другим птицам, была свойственна подвижность и верхней, и нижней челюсти. У большинства позвоночных, в том числе у рептилий подвижна только нижняя челюсть". |
| Джонатан Сарфати. "Несостоятельность теории эволюции" [136]. Курсив добавлен. |
Сведения Сарфати о мозге археоптерикса и подвижности верхней челюсти просто ошибочны. Лучшая реконструкция мозга археоптерикса, проведенная в 2004 году с помощью компьютерной томографии, показала, что хотя его мозг в основных чертах соответствовал птичьему образцу, все же в его строении прослеживается ряд рептилийных черт. Поэтому мозг археоптерикса имеет промежуточное строение. Об этом шла речь выше. Столь же неправильны сведения о подвижности верхней челюсти у археоптерикса. Строение верхних челюстей не позволили бы типичные для птиц движения. Те особенности костей черепа, которые присутствуют у археоптерикса и делают возможным подвижность верхней челюсти у птиц, присутствуют также у некоторых динозавров. По тем характеристикам, которые подразумевает Сарфати, археоптерикс не больше птица, чем динозавр [36]. То же верно в отношении упомянутой вилочки. Она хорошо известна у звероногих динозавров, да еще у тех, которые ближе всех стоят к птицам [54]. А крылья археоптерикса по строению весьма далеки от современных птиц. Не случайно Сарфати тут ссылается на их форму, а не на строение, как это он делает, например, в случае с перьями. Но самое важное, Сарфати ничего не может противопоставить множеству рептилийных черт археоптерикса и лучшее, что он может придумать, - сделать вид, будто их нет.
Археоптерикс – пример переходной формы №1. Год за годом, десятилетие за десятилетием креационисты говорят, что археоптерикс не может считаться переходной формой. Однако за несколько десятилетий они так и не преступили к работе, которую нужно было проделать давным-давно, и которую проделали ученые еще в 19 веке.
В 1912 году английский философ Бертран Рассел, говоря об изменении мировоззрения людей в XIX столетии, писал:
""Происхождение видов" Дарвина убедило мир, что видовые различия животных и растений не являются жесткими и неизменными... Было показано, что различие между человеком и низшими животными, которое человеческому разумению кажется неимоверно большим, есть достижение постепенное, предполагающее существование промежуточных существ, которых нельзя без колебаний ни включить в человеческую семью, ни исключить из нее". |
| Бертран Рассел. "Мистицизм и логика" [134]. |
На момент написания этих строк почти все открытия предков и родственников людей были еще впереди. В начале, в середине, в конце XX века, и в начале XXI креационисты отвергали и отвергают высказанную Расселом и другими эволюционистами точку зрения. По мнению креационистов, напротив, нет никаких промежуточных существ между человеком и другими животными. А все предполагаемые "переходные звенья" между человеком и другими приматами – либо обезьяны, либо люди.
"До настоящего времени не было ни одной палеонтологической находки, которая подтвердила бы правильность эволюционной теории происхождения человека. Окаменелые останки принадлежат либо явно человеку, либо обезьянам". |
Дж. Маклин и др. "Очевидность сотворения мира". [129] |
"Креационисты утверждают, что человек всегда был человеком, а обезьяны всегда были обезьянами. Нет никаких доказательств, что это неверно". |
Пол Тейлор. "Сотворение. Иллюстрированная книга ответов" [138]. |
"Есть множество окаменелостей настоящих обезьян и настоящих людей, но нет ничего промежуточного. Человекообезьяны или обезьяночеловека не существовало никогда". |
Генри Моррис. "Библейские основы современной науки" [130]. |
"Таким образом, налицо доказательства, что вера в "обезьянолюдей" необоснованна. Напротив, все отличительные признаки людей свидетельствуют о сотворении, происшедшем отдельно и обособленно от всех животных... Всякие обезьяноподобные существа, жившие в прошлом, были не кем иным, как человекообразными обезьянами... но не людьми. |
"Жизнь - как она возникла? Путем эволюции или путем сотворения?" [126] |
"...не существует ни одного достоверно зафиксированного случая находки предков человека". |
Кен Хэм и др. "Книга Ответов" [140]. |
"...различные находки представляют собой либо действительно представителей рода человеческого (например, неандертальцы или Homo erectus), либо существ, не относящихся к людям, - таких, как австралопитеки..." |
Джонатан Сарфати. "Несостоятельность теории эволюции" [136]. |
"Однако постепенно исследователи склоняются к мнению, что все находки ископаемых, имеющие непосредственное отношение к предполагаемой эволюции, могут быть поделены на две группы: Homo sapiens (человек разумный) и Australopithecus (южная обезьяна)." |
Уильям Гладсхауэр. "Как возник наш мир" [124]. |
Креационисты главным образом списывают друг у друга аргументы. Так появляется огромное количество книг, фильмов, лекций и пр. по существу одинакового содержания. Большинство креационистов не проводят самостоятельного анализа материала. Но человеческое происхождение – особая тема. Ею независимо занимались несколько авторитетных креационистов (археоптериксу и другим примерам переходных форм повезло меньше). Так каким образом следует разграничивать обезьян отдельно, людей отдельно? Вот результаты, к которым пришли креационисты.
| Экземпляр | Cuozzo (1998) |
Gish (1985) |
Mehlert (1996) |
Bowden (1981) Menton (1988) Taylor (1992) Gish (1979) |
Baker (1976) Taylor and Van Bebber (1995) |
Taylor (1996) Lubenow (1992) |
Line (2005) |
|
|---|---|---|---|---|---|---|---|---|
 |
ER 1813 (510 cc) |
Обезьяна | Обезьяна | Обезьяна | Обезьяна | Обезьяна | Обезьяна | Человек ??(2) |
| Java (940 cc) |
Обезьяна | Обезьяна | Человек | Обезьяна | Обезьяна | Человек | Человек | |
 |
Peking (915- 1225 cc) |
Обезьяна | Обезьяна | Человек | Обезьяна | Человек | Человек | Человек |
 |
ER 1470 (750 cc) |
Обезьяна | Обезьяна | Обезьяна | Человек | Человек | Человек(1) | Человек ??(3) |
 |
ER 3733 (850 cc) |
Обезьяна | Человек | Человек | Человек | Человек | Человек | Человек |
 |
WT 15000 (880 cc) |
Обезьяна | Человек | Человек | Человек | Человек | Человек | Человек |
Не смотря на все категорические заявления об однозначном разделении имеющихся окаменелостей на обезьян и людей, мы видим у креационистов те самые колебания, о которых писал Рассел на заре палеоантропологии. Иронично, креационисты сами подтверждают то, что пытаются опровергнуть. Причем в вопросе людей и обезьян они иногда меняют точку зрения на противоположную. Из таблицы видно, например, что Тейлор трижды изменял разделение на людей и обезьян, Гиш дважды. В результате вчерашние обезьяны у креационистов вдруг приобретали иную анатомию, превращаясь в людей. И только превращаться в переходные формы им не позволено.
Креационисты предлагают еще одно объяснение существованию известных переходных форм. Они утверждают, что бог по своему усмотрению сочетал в некоторых животных признаки разных групп. Таким образом, то, что ошибочно принимают за переходные формы, – ничто иное, как результат божьего промысла.
"Все формы, рассматриваемые как промежуточные звенья, проявили себя типичными мозаичными формами, комбинацией признаков из двух и более групп. Так, например, археоптерикс (Archaeopteryx) проявляет признаки рептилий и птиц." |
| Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
"Не логичнее ли предположить, что мозаичные формы являются совершенно особыми творениями, которые созданы Творцом по особому замыслу? Ведь Конструктору всего живого не требовалась наша таксономическая система, где птицы, звери и гады строго разделены между собой?" |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
Уместно рассмотреть подход креационистов к оценке окаменелостей. Ирония ситуации состоит в том, что среди креационистов нет ни одного палеонтолога. Те креационисты, которые больше других писали об окаменелостях, профессионально не связаны с палеонтологией или даже биологией вообще. Дуэйн Гиш – биохимик, Карл Виланд – врач, Джонатан Сарфати – химик, Кен Хэм специализировался в прикладных науках и т. д. Причем все они оставили научную карьеру и формально или фактически являются проповедниками. Большинство же креационистов не занимаются даже первичным изучением научной литературы. Вместо этого они без малейшей проверки переписывают свои аргументы у тех же Гиша или Хэма. Однако есть более компетентные креационисты, заслужившие похвалу даже от критиков. Примечательно, это те самые авторы, которые предложили идею мозаичных форм.
Так немецкие креационисты Юнкер и Шерер, обсуждая переходные формы, отмечают, что в их строении действительно сочетаются признаки изолированных групп. Но Юнкер и Шерер настаивают, что эти животные не переходные формы, а причудливые комбинации различных черт, объяснимые в модели создания. Они также утверждают, что мозаичные формы широко встречаются в природе сегодня.
Аналогичные аргументы предлагает американский креационист Курт Вайс. В работе, посвященной переходным формам, Вайс пишет, что "креационная палеонтология" находится в зачатке. Он убеждает креационистов не отрицать существование переходных форм, так как это может сделать их аргументы уязвимыми для эволюционистов. Вместо этого Вайс предлагает, чтобы креационисты подыскали им собственное объяснение и собственное название. Он также формулирует 4 прогноза эволюционной теории о том, какие окаменелости должны быть найдены, и отмечает, что частично эти прогнозы соответствуют палеонтологическим находкам. Однако в заключении Вайс выражает надежду, что креационисты смогут дать этим примерам лучшее объяснение. Относительно примеров переходных окаменелостей он пишет:
"Из четырех дарвинистских ожиданий промежуточные стратоморфические звенья оказались наиболее разочаровывающими для классических дарвинистов. Существующий на сегодняшний день недостаток каких-либо стратоморфических промежуточных звеньев привел к разработке теории прерывистого равновесия и способствовал ее принятию. Свидетельство для второго ожидания Дарвина – видовых стратоморфических промежуточных звеньев – включает такие виды как барагванатия (между риниофитами и плауновидными), пикайя (между иглокожими и хордовыми), пургаториус (между тупайии и приматами) и проконсул (между негоминидными приматами и гоминидами). Третье ожидание Дарвина – стратаморфические промежуточные звенья более высоких таксонов – было подтверждено такими примерами как группа млекопитающеподобных рептилий между рептилиями и млекопитающими, и фенакодонтами между лошадьми и их предполагаемыми предками. Четвертое ожидание Дарвина – стратоморфические серии – было подтверждено такими примерами как серия ранних птиц, серия китов, различные серии каинозойских млекопитающих (например, серии лошадей, верблюдов, слонов, свиней, титанозавров и др.), цепочки приматов кантиус и плезиадапус и цепочка гоминидов. Свидетельство для существования стратоморфических промежуточных звеньев не только для одного уровня (уровня вида и выше уровня вида), а для трех типов ожидается в соответствии с макроэволюционной теорией. Соответственно, креационисты должны принять данный факт. Конечно, нельзя говорить, что традиционная креационная теория предсказала любую из упомянутых находок". |
| Kurt Wise. "Towards a Creationist Understanding of Transitional Forms" [87]. Внутренние ссылки опущены |
Следует еще раз подчеркнуть, что Вайс не принимает переходного статуса перечисляемых им форм. Он надеется обосновать их существование в результате божественного сотворения и последующего изменения в рамках изначально созданных видов. Вайс известен высказыванием: "Если все свидетельство во вселенной обратится против креационизма, я первым признаю это, но я все равно останусь креационистом, так как, судя по всему, Слово Бога подразумевает креационизм". В этих словах Вайс явно демонстрирует догматическую природу креационизма. Креационисты не в состоянии изменить свои представления о мире, даже если все свидетельство во вселенной будет говорить в пользу необходимости такого изменения. По контрасту наука, под которую маскируется креационизм, ценна именно тем, что стремится описать мир таким, каков он есть, независимо от каких-либо ожиданий, предпочтений или верований. Сама попытка замаскировать религиозный догматизм под нечто научное, которая зовется креационизмом, порождена тем, что знания о мире, полученные наукой, противоречат верованиям, основанным на буквальном прочтении священных текстов, и приверженцы таковых верований стремятся что-то противопоставить научным выводам.
Очевидно, Вайс пошел дальше, чем кто-либо другой из креационистов, в признании, что есть окаменелости, которые можно справедливо расценить как подтверждение теории эволюции. Это не случайно. Вайс не палеонтолог, он – геолог, выпускник Гарварда, ученик знаменитого палеонтолога Стивена Джей Гулда. Выше я отметил, что без длительного курса палеонтологии и анатомии примеры переходных форм будут для человека пустым звуком. Теперь мы сталкиваемся с подтверждением этих слов. Вайс, Юнкер и Шерер явно не видят возможности отрицать очевидное – сочетание признаков отдаленных групп животных в окаменелостях, поэтому им ничего не остается, как признать данный факт. И они пытаются найти другое объяснение.
Однако концепция мозаичных форм противоречит отрицанию комбинации признаков, с которым мы сталкивались выше. Очевидно, кто-то из креационистов неправ. Археоптерикс, например, не может быть одновременно и птицей, и мозаичной формой. Если у него нет комбинации признаков, значит, Вайс, Юнкер и Шерер и др. неправы. Но если такая комбинация присутствует, значит, неправы Гиш, Сарфати и другие креационисты.
Удивительно, некоторые креационисты используют оба аргумента одновременно. Те же Юнкер и Шерер называют произвольно одних вымерших животных, например, рыбами, а других мозаичными формами рыб и амфибий. Причем они не объясняют, каким образом приходят к своим выводам, какие методы используют. Читателю предлагается просто поверить на слово. Другие креационисты отрицают, что у изученных форм прослеживаются комбинации особенностей, характерных для изолированных групп, но оставляют идею мозаичных форм про запас.
И все же неспособность креационистов согласится друг с другом даже более внушительна. Выше мы видели, как Курт Вайс убеждал креационистов не отрицать существование переходных форм, использовать для них другое название и говорил, что найденные окаменелости соответствуют теории эволюции. В конце статьи Вайс благодарит нескольких креационистов, за ценные комментарии и замечания к его рукописи. Среди упомянутых креационистов Карл Виланд. Однако несколько лет спустя Карл Виланд вместе с тремя другими авторами издает "Книгу ответов", в которой читаем:
"Дарвин предполагал, что в окаменелостях будут найдены миллионы переходных форм, однако вплоть до нынешнего дня обнаружена лишь горстка более чем сомнительных форм, в "переходности" которых никто не уверен... систематическое отсутствие предполагаемых эволюционных "звеньев" в глобальном масштабе – весомое свидетельство в поддержку библейской точки зрения на Сотворение мира". |
| Кен Хэм и др. "Книга Ответов" [140]. Курсив в оригинале. |
Как Виланд может делать подобные утверждения, если он помогал Вайсу улучшить упомянутую статью? Можно, конечно, предположить, что Виланд и его соавторы не согласны с Вайсом. Но почему они делают столь однозначные выводы, если знают, что даже среди креационистов не все согласились бы с ними.
Теперь рассмотрим саму концепцию мозаичных форм. Прежде всего, бросается в глаза, что это богословская концепция. Попытки найти укрытие в богословии вовсе не редкость в моделях креационистов. Когда креационисты не в состоянии объяснить нечто, они возлагают ответственность за его существование на бога. Так, к примеру, они не могли объяснить, почему мы можем видеть свет от космических объектов, удаленных от нас на десятки тысяч или миллионы световых лет, если вселенной, по их мнению, всего 10,000 лет. Некоторые креационисты утверждали, что бог преднамеренно создал свет, движущийся от объектов, которые еще не существовали. Когда генетики начали сравнивать длинные фрагменты ДНК разных видов, выяснилось, что геномы содержат большое количество атрофированных генов, и что эти атрофированные последовательности разделены между группами, которые креационисты считают созданными отдельно (человек и обезьяны, например). Тот же Карл Виланд предположил, что такие последовательности были специально созданы богом для испытания крепости веры креационистов. Правда, под влиянием критики он отказался от этого аргумента.
Поскольку мозаичные формы – это богословская, а не научная идея она должна оцениваться на богословских и умозрительных основаниях, а не научных. На самом деле, не можем же мы создать математический алгоритм, применить его к богословскому объяснению и дать заключение, что оно верно с вероятностью X, при P < Y.
Явный недостаток концепции мозаичных форм состоит в том, что для нее не существует никаких оснований, ни умозрительных, ни богословских. Более того, избавляясь от переходных форм ссылкой на прихоть бога, креационисты симулируют, что знают, как именно творил бог, что он намеривался сделать. Именно такую теологию высмеивал еще в XIX веке А. К. Толстой:
Отчего б не понемногу
Введены во бытие мы?
И не хочешь ли уж богу
Ты предписывать приемы?
Способ, как творил Создатель,
Что считал он боле кстати,
Знать не может председатель
Комитета о печати.
Ограничивать так смело
Всесторонность Божьей власти -
Ведь такое, Миша, дело
Пахнет ересью отчасти!
Ту же самую мысль, но высказанную на полном серьезе, мы находим у богословов в XX веке, на сей раз в официальном документе:
"Братство Епископов приходит к общему выводу: 67-ой Генеральный съезд подтверждает, что он верит в замечательную способность Бога творить любым способом, и в этом подтверждении отклоняет жесткий догматизм "Креационного" движения..." |
Итак, концепция мозаичных форм, как минимум, сомнительна с богословской точки зрения. Но если ее рассмотреть внимательно, становится ясно, что эта идея просто невероятна по трем причинам.
1. Идея мозаичных форм абсолютно бесплодна. Используя ее креационисты не в состоянии прогнозировать, какие мозаичные формы мы можем обнаружить, а какие нет. Бог по своему усмотрению может сочетать любые признаки в любых животных. То есть идея мозаичных форм предельно широка. Она может быть сформулирована кратко, как "возможно все". С другой стороны, эволюционисты прогнозируют, какие мозаичные формы должны быть обнаружены. Причем, по крайней мере, часть этих прогнозов была подтверждена. Складывается конфузная ситуация: креационисты, используя правильную теорию, не могут делать каких-либо прогнозов, в то время как эволюционисты, используя неправильную теорию, дают правильные прогнозы.
2. Если рассмотреть предполагаемые мозаичные формы внимательно, становится очевидным, что бог творил их не просто так, а по определенной схеме. Как если бы он желал подражать эволюции, симулировать ее, сверяясь с филогенетическими деревьями из работ палеонтологов. Бог сотворил мозаичные формы между рептилиями и птицами, в полном согласии с Генри Хаксли. Он почему-то не пожелал сотворить мозаичные формы между птеродактилями и птицами, или птицами и млекопитающими. Он сочетал в некоторых животных признаки рыб и амфибий, но не рыб и рептилий. Снова эволюционисты успешнее креационистов понимают и описывают предполагаемую работу бога.
3. Еще один вывод, который с неизбежностью следует из анализа мозаичных форм – бог комбинировал признаки только двух групп, не более. Насколько можно судить, в природе отсутствуют формы, сочетающие комплексы диагностических признаков, характеризующих сразу три изолированные группы. Мы можем ожидать такой результат от эволюции, так как потомковая группа имеет только одну предковую. Например, птицы имеют предками только рептилий, а, скажем, не рептилий и млекопитающих одновременно. Однако бог не мог иметь таких ограничений. Он в принципе мог бы объединять признаки любых групп, но почему-то этого не делал. Еще раз, такое впечатление, что он моделировал эволюцию.
И вот здесь креационисты сказали бы, что я ошибаюсь. На самом деле есть формы, у которых сочетаются признаки нескольких групп одновременно. Юнкер и Шерер приводят в качестве примера археоптерикса. По их словам, у археоптерикса есть признаки одновременно четырех групп. Действительно ли это так? Рассмотрим их признаки в таблице.
| Признак | Группа |
| Конические зубы | Крокодилы |
| Четырехлучевой тазовый пояс | Птицетазовые динозавры |
| Двояковогнутые позвонки | Ихтеозавры |
| Специализированные когти | Ящеротазовые динозавры |
Однако легко показать, что этот список порожден недостатком знаний.
Прежде всего, двояковогнутые позвонки присущи не только ихтеозаврам. Как мы видели, они также свойственны ближайшим родственникам археоптерикса.
Четырехлучевой таз – ошибка. Авторы используют весьма устаревшую реконструкцию археоптерикса (см. их рисунок 8.68).
Специализированные когти соответствуют эволюционным отношениям. Птицы связаны с ящеротазовыми динозаврами (конечно, не со всеми сразу), более точно с дейнонихозаврами.
Конические зубы – крепкий орешек. Действительно зубы археоптерикса и его ближайших родственников значительно отличаются. Долгое время приходилось предполагать, что были и такие дейнонихозавры, которые имели конические зубы крокодильего типа. Этот прогноз был подтвержден в 90-ых годах XX века.
Итак, археоптерикс сочетает в себе признаки только двух родственных по эволюционным представлениям групп.
Есть еще кандидаты? Креационисты думают, что есть. Это причудливый утконос. Кен Хэм, объясняет своим слушателям:
"Дело в том, что у утконоса клюв как у утки, а хвост как у бобра. Шерсть у него как у медведя, перепончатые лапы как у выдры, когти как у рептилии, он кладет яйца как черепаха, у него шпоры как у петуха, и он ядовит как змея. ... Для эволюционистов утконос – настоящая проблема. Они верят, что за миллионы лет одни животные эволюционировали в других, но вопрос вот в чем: от какого животного произошёл утконос? Такое впечатление, что он мог произойти от кого угодно! Мне кажется, всякий раз, когда эволюционист смотрит на утконоса, Бог улыбается. Может быть, Он для этого его и создал". |
Представленный здесь перечень основан на внешних и общих подобиях. Простой экскурс в зоологию расставляет все на свои места. Примечательно, что в сотрудничестве с другими креационистами Хэм приводит значительно укороченный список:
"У утконоса утиный клюв и перепончатые лапы, но никому не приходит в голову утверждать, будто утконос произошел от утки". |
| Кен Хэм и др. "Книга Ответов" |
Разумеется, перепончатые лапы – вездесущая особенность в животном мире. Разумеется, у утконоса нет какого-то особенного утиного клюва, отличающегося уникальным строением. Креационисты обобщают здесь признаки по внешней форме, и не в состоянии привести примера структурного сходства. Если же мы будем принимать в расчет внешние признаки, можно будет сказать, что человек – мозаичная форма между птицами и млекопитающими. Как птица он ходит на двух задних ногах, а как млекопитающее имеет три слуховые косточки. Чтобы утверждать совпадение черт в разных формах, необходимо доказать гомологию. А этого креационисты сделать не в состоянии. Более того, роговые покровы челюстей у птиц и утконоса структурно отличны. Они образованы различными типами белков. Клювы птиц составлены из β-кератина, в то время как "клюв" утконоса образован α-кератином. Таким образом, эти образования, не смотря на кажущееся подобие, негомологичны.
В целом, предлагаемые креационистами мозаичные формы, как минимум крайне сомнительны. Юнкер и Шерер утверждают, что мозаичные формы широко распространены в природе. Часть их примеров связана с утратой признака, но утрата не может быть свидетельством мозаичности.
Попугаи и туканы не имеют сросшихся ключиц, у паламодей нет крючковатых отростков ребер, у совиных попугаев нет киля или он сильно уменьшен. Однако если группа теряет признак, она не становится мозаичной. Ведь объяснение о мозаичных формах состоит в том, что бог сочетает признаки разных групп, а не отнимает их. Мы знаем, что безногие амфибии, змеи и двуходки потеряли конечности. Но креационисты не называют их мозаичными. И действительно, кем они стали – мозаичной формой между рептилиями и червями? Точно также некоторые амфибии утратили легкие. В этом случае мне даже сложно предположить, в чем их мозаичность.
Другие примеры не лучше.
Крокодил имеет четырехкамерное сердце, как птица. Сердце крокодила действительно разделено на четыре камеры, но не как у птиц. В отличие от сердца птиц сердце крокодила специализировано, что свидетельствует о параллельном происхождении деления на четыре камеры. Характерная особенность кровеносной системы крокодилов – способность переходить от разделения венозной и артериальной крови к ее смешению. В основании сосудов, отходящих от сердца крокодила, имеется так называемое паницциево отверстие, через которое артериальная кровь смешивается с венозной и попадает в правую дугу аорты. Если паницциево отверстие закрыто, то по левой дуге аорты идет венозная кровь, несущая к желудку не кислород, а углекислоту. У птиц нет ни левой дуги аорты, ни паницциева отверстия.
"Высиживающий потомство из яиц питон вырабатывает собственное тепло". Авторы явно не замечают, что это не анатомическая особенность вообще.
Итак, легко заметить, что креационистам удается обнаружить мозаичные формы в современном мире дорогой ценой. Им приходится выделять организмы по единичным признакам (иногда по отсутствию признаков) или по поверхностным сходствам, что приводит к абсурду. Конечно, все эти примеры несопоставимы с тем, чем являются переходные формы, обнаруживаемые в окаменелостях.
Креационисты возражают: в окаменелостях мы можем встретить комбинацию признаков, но никто пока не нашел частично развитого признака, признака находящегося на промежуточной стадии развития. И это подтверждение мозаичности, а вовсе не переходности известных примеров.
"От истинно переходной формы следует ожидать, что она продемонстрирует медленное изменение отдельных признаков". |
| Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
"Ни один из палеонтологов не нашел еще полу-чешуйки - полу-пера и полу-лапы – полукрыла. ... Главная особенность мозаичных форм - подчеркнем еще раз – это совершенно законченные в своем формировании структуры – перья, шерсть, чешуя и т. п., хотя в одном организме сочетаются структуры, свойственные разным классам животных". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
"Описанные животные удивительно сочетают в себе части тел pазных видов, как своеобразные фрагменты мозаики, но такие фрагменты (перья археоптерикса, утиный нос и шерстный покров утконоса, зубы и челюстные суставы зверозубого ящера) имеют у них вполне законченную фоpму, а отнюдь не промежуточную." |
| Сергей Вертьянов. "Общая биология" [122]. |
"Однако в обоих случаях мы имеем дело не с переходными, а с мозаичными формами. У них нет никаких переходных структур, их перья полностью развиты и функционируют, нет ни одного лишнего или рудиментарного органа... У археоптерикса были зубы и челюсти рептилии, а перья и крылья - птицы, но все органы были полностью сформированы и служили нуждам организма". |
| Генри Моррис. "Библейские основы современной науки" [130]. |
И снова креационисты за десятилетия не потрудились проделать необходимую работу. Промежуточные органы и структуры есть и в большом количестве. Передняя конечность археоптерикса с развитыми пальцами, полулунной костью, начавшимся срастанием пястных костей, снабженная ассиметричными летательными перьями – есть ничто иное как "полукрыло". Еще один пример – частично сформированная цевка археоптерикса. Суставная ямка лопатки у археоптерикса имеет промежуточное положение. Промежуточное строение имеет и коракоид археоптерикса. Как было сказано выше, он плоский, грубо квадратной формы, что резко контрастирует с характерным для птиц строением. Однако в полном соответствии с птичьей анатомией он снабжен выступающим вперед отростком. У другой древней птицы - конфуциусорниса - шейные позвонки имеют промежуточное строение. Только одна суставная поверхность позвонка имеет отличительно птичью форму, другая остается ближе к рептилийному состоянию. Теперь по ранним птицам и их ближайшим родственникам можно проследить формирование типичной для птиц кисти с сокращением числа фаланг и преобразованием пальцев. Также можно проследить развитие крестцового отдела позвоночника, число позвонков в котором увеличивалось вплоть до современного количества. Уже упомянутые шейные позвонки дают и другую интересную информацию. У птиц они имеют характерные пустоты, связанные с дыханием. Окаменелости позволяют установить, как возникали эти особенности из небольших впадин на телах позвонков с последующим расширением пустот. Еще один такой пример - положение подвздошных костей гуанлонга. Для большего количества примеров см. [133].
|
| Модификация грудины и формирование киля у мезозойских птиц, вид снизу. a. джехолорнис, две пластины грудины не срастаются. b. Эоконфуциусорнис, части грудины срастаются между собой, но киль отсутствует. c-d. два разных конфуциусорниса, начальные стадии формирования киля. У некоторых современных птиц киль утрачен. В классе птиц выделяют надотряд бескилевых птиц, почти все из которых утратили способность к полету. Поэтому они не нуждаются в особом креплении для летательной мускулатуры. Из бескилевых летать могут только тинаму, однако полет дается им с трудом и летают они редко. |
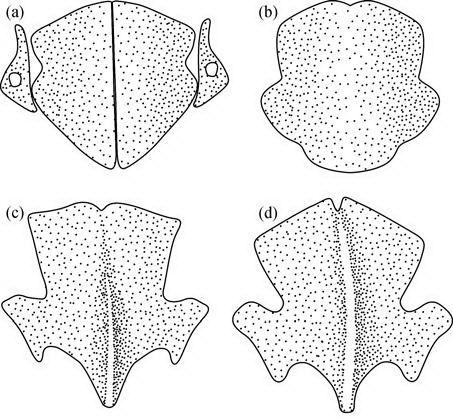
Любопытно, несуществующие промежуточные состояния органов упоминают даже креационисты, которые отрицают их существование. Юнкер и Шерер пишут:
"Следующим... "примитивным признаком" археоптерикса является отсутствие киля на грудине. Но он отсутствует почти полностью и у новозеландских совиных попугаев, а у большинства буревестников киль развит очень мало". |
| Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
"Развит очень мало" - что это, если не промежуточное состояние структуры? Любопытно, что мы можем обнаружить несостоятельность концепции "мозаичных форм", основываясь на работах самих креационистов. В этой связи важно обратить внимание, что креационисты готовы увидеть мозаичные формы в любых видах, кроме одного – человека. Когда речь заходит о промежуточных звеньях между древними гоминидами и человеком, креационисты не могут сказать, что бог по какой-то причине творил мозаичные формы между обезьянами и людьми. Такое предположение не согласуется с их верой в уникальность сотворения человека. Поэтому единственное, что им остается – пытаться провести черту "одно из двух" (см. выше).
Подводя итог обсуждению "мозаичных форм", можно сделать три вывода:
Во-первых, идея о существовании мозаичных форм по существу скрытое молчаливое признание переходных форм только под другим названием.
Во-вторых, если мозаичные формы были сотворены богом по своему усмотрению, то он творил так, как будто хотел симулировать эволюцию.
В-третьих, основываясь на креационизме, вы никогда не сможете прогнозировать, какие мозаичные формы могут быть обнаружены в будущем. Но вы можете успешно делать подобные прогнозы, используя эволюционные гипотезы.
Креационисты иногда утверждают, что некоторые окаменелости были преднамеренно подделаны эволюционистами для доказательства теории. Ожидаемый пример – все тот же многострадальный археоптерикс. Когда в 80-ых годах XX века астроном Фред Хойл с группой единомышленников заявил, что лондонский экземпляр археоптерикса – подделка, некоторые креационисты поспешили воспользоваться его аргументами. Но переисследования экземпляров не показало следов подделки, а обнаружение новых экземпляров уничтожило саму возможность. Поэтому креационисты перестали говорить о подделке археоптерикса. Хотя некоторые креационисты упоминают возможность подделки, не делая, впрочем, никаких выводов. Воистину юрская птица у креационистов успела побывать всем, чем угодно.
Другая подделка – химера птицы и динозавра, купленная на черном рынке в Китае. О ней сообщили некоторые СМИ до того, как исследование экземпляра было завершено. Научно-популярный журнал Национального географического общества США посвятил находке сенсационную статью. Но спустя некоторое время подделка была раскрыта. Креационисты намекают, что данный случай свидетельствует о нечестности эволюционистов, и что таких подделок может быть больше.
"События вокруг археораптора дают нам редкую возможность взглянуть на идеологический и журналистский подход, определяющий отношение общества к "недостающим звеньям"". |
| Стивен Остин, "Археораптор" [132]. |
В действительности окаменелость была склеена китайским фермером, без сомнения для того, чтобы побольше заработать на ее продаже. Кустарные раскопки в китайской провинции Ляонин стали весьма популярными среди сельских жителей после обнаружения богатых "кладбищ" окаменелостей. Искать здесь следы эволюционного заговора глупо. Вовсе не вера в эволюцию заставляет китайских фермеров искать ценные кости.
Любопытно, креационисты называют ляонинскую химеру латинским названием – археораптор, как будто ученые дали звучное имя подделке, сравнимое, например, с археоптериксом. В действительности это название никогда не существовало. Я не думаю, что креационисты говорят об археорапторе из-за нечестности. Скорее, я предполагаю, что они не знакомы с процессом принятия названий в науке. Чтобы появился новый род, он должен быть формально описан в научной литературе на основе экземпляра в соответствии с принципами "Международного кодекса зоологической номенклатуры". Археораптор никогда не был описан, никогда не был частью науки. Он был журналистской сенсацией. Но журналисты всегда ищут сенсации. Эволюционная сенсация здесь ничуть не хуже и не лучше креационной. Правда, Национальное географическое общество не публикует креационных сенсаций. Но учитывая историю с ляонинской химерой, креационисты должны быть этим, скорее, довольны.
Еще одна любимая креационистами подделка – пилтдаунский человек. На сей раз действительно часть науки. Его латинское имя действительно существовало, правда, креационисты, напротив, используют его не так уж часто. В отличие от ляонинской химеры пилтдаунский человек около 30 лет был в почете у ученых. Конечно, некоторые исследователи и тогда считали его подделкой, а в конце своей "карьеры" он оказался в основном забыт. И все же не следует приуменьшать его роль. Мейнстримная точка зрения была на стороне пилтдаунского человека. Тем не менее, пилтдаунский человек устарел. Пилтдаунский обман был разоблачен в 50-ых годах XX века. Креационное движение оформилось в следующем десятилетии. В те годы разоблачение пилтдаунского человека было событием. Но сегодня, 50 лет спустя пилтдаунский человек уже давно не часть палеантропологии. Он больше не актуален. Тем не менее, креационисты уделяют ему больше внимания, чем подлинным и важным окаменелостям, как будто еще вчера пилтдаунский человек владел умами.
Нежелание креационистов расставаться с пилтдаунским человеком вполне объяснимо. Это лучший пример явного обмана в истории изучения эволюции человека, который они могут найти. И он очень важен для их попыток нивелировать переходные окаменелости в человеческой родословной. Креационисты рассматривают ископаемых гоминидов неизменно однобоко. Сначала они рассматривают все находки, оказавшиеся ошибками, чтобы создать у читателей впечатление, что история палеонтропологии – это нечто вроде череды ошибок и обманов. Затем они описывают значимые находки, создавая впечатление, что последние столь же сомнительны, как и первые. Конечно, увлекаясь разоблачениями, креационисты избегают сравнения скелетных особенностей.
4. Трудности с эволюционными отношениями.
Когда креационисты имеют дело с несколькими примерами переходных форм, они часто указывают, что эволюционисты не могут выстроить их в четкий последовательный ряд от первого предка до последнего потомка. Так российский креационист Алфёров пишет об эволюции лошадей:
"...если лошадок расставить в воображаемой эволюционной последовательности, то окажется такая "эволюция" количества ребер: сначала 18, потом 15, потом 19, наконец снова 18. Подобные же вариации наблюдаются и в количестве позвонков". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
Чтобы разобраться с последовательными рядами окаменелостей, потребуется экскурс в историю биологии.
На заре эволюционной биологии большую популярность получила концепция направленной эволюции (ортогенез). Предполагалось, что в силу тех или иных причин формы жизни обнаруживают стремление к определенному состоянию, которое часто считалось "лучшим". Соответственно палеонтологи пытались воссоздать прямую последовательность форм от предка к потомку. Поначалу это не вызывало серьезных трудностей, поскольку небольшое количество описанных родов и видов проще разместить в последовательный ряд. И трудно сказать, это возможность легко составить ряд предок-потомок убеждала палеонтологов в направленности эволюции, или убежденность в прямолинейном ее характере побуждала их создавать такие реконструкции. Мы можем видеть, как эти представления нашли отражение в исследовании эволюции лошадей.
Начало исследований эволюции лошадей связано с именами двух знаменитых палеонтологов XIX века – Эдварда Копа и Чарльза Марша. Оба ученых питали друг к другу неприязнь и стремились опередить друг друга в количестве найденных и описанных окаменелостей, обнаруженных новых видов и т. д. Но примечательно, что независимо друг от друга и, не имея никакого желания соглашаться с мнением друг друга, они все же пришли к сходным взглядам на эволюцию лошадей. Оба заключили, что эволюция лошадей была направленной, последовательной и прогрессивной. Когда знаменитый биолог-эволюционист Томас Генри Хаксли посетил США, Чарльз Марш продемонстрировал ему собранные окаменелости и рассказал о своих взглядах на эволюцию лошадей. Хаксли, интересовавшийся этой темой, попросил Марша сделать рисунок, иллюстрирующий эволюцию лошадей, и Марш изобразил то, что легло в основу канонической схемы эволюции.
|
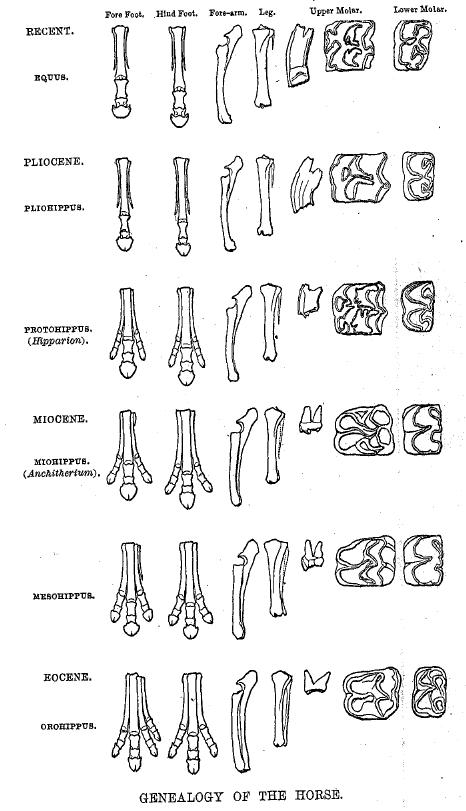
Нетрудно заметить, что рисунок четко отобразил идею Марша о направленной эволюции.
Другой влиятельный палеонтолог ученик Копа Генри Осборн в начале XX века также настаивал на направленном характере эволюции. Осборн утверждал, что линейная эволюция обнаруживается во всех группах млекопитающих, которые он исследовал. Хотя позднее Осборн отметил, что, возможно, более одной формы лошадей существовали в одно и то же время. Однако он не придал этому наблюдению значения.
|
Еще один знаменитый рисунок направленной эволюции. Впервые опубликован Осборном в 1917 году. Рисунок сопровождался следующим комментарием: "Стадии эволюции рога у титанотерий. Показывает, что это важное оружие возникает в результате эволюционной тенденции, то есть ортогенетически, а не в результате отбора случайностей или случайной генетической изменчивости". В современной классификации титанотерии называются бронтотериевыми.
Однако по мере обнаружения все большего числа окаменелостей, становилось все сложнее размещать их в последовательные ряды. Палеонтологи начали осознавать, что реальная эволюционная история не укладывается в линейные схемы от предка к потомку. Первым, кто предложил разветвленную модель эволюции лошадей, был сотрудник Осборна Уильям Гидли. Еще в 1907 он писал о том, что, возможно, существовало несколько ответвлений от эволюционной линии лошадиных, которые какое-то время эволюционировали изолированно друг от друга.
|
Обозначения снизу вверх: Эоцен – Олигоцен – Миоцен – Плиоцен – Плейстоцен – современность. |
Схема эволюции лошадиных по Гидли. Сопровождалась следующим описанием: "Вероятные отношения четырех подсемейств ... отображены ... в диаграмме".
В последующих десятилетиях идея линейной, направленной эволюции все больше подвергалась сомнению. В начале 50-ых другой выдающийся палеонтолог Джордж Гейлорд Симпсон издал популярную книгу об ископаемых лошадях, в которой представил уже широко признанное профессиональными палеонтологами разветвленное эволюционное древо этой группы и разъяснял, почему идея направленной эволюции неправдоподобна. Симпсон указал, что эволюция лошадей не следовала за какими-либо неизменными тенденциями. Он также отрицал существование главных и побочных ветвей. Действительно, на каких основаниях мы считаем одну ветвь главной, а вторую побочной? Допустим, мы считаем, что все вымершие ветви побочные. Но что если выживет две ветви или больше? Или в конечном счете вымирают все представители всех ветвей. Какая будет главной, а какая побочной?
|
Схема эволюционных отношений лошадей из книги Симпсона, 1951 год. Grazing horses – лошади, питавшиеся травой. Browsing horses – лошади, питавшиеся листвой. |
С этой информацией в памяти мы теперь можем оценить критику креационистов. Легко заметить, что в понимании эволюции лошадей и в понимании эволюции крупных групп вообще креационисты остались в 19 веке. Они продолжают нападать на схему направленной эволюции, несмотря на то что она была оставлена как минимум полвека тому назад. Поразительно и то, что креационисты ниспровергают теорию направленной эволюции и полагают при этом, что доказывают несостоятельность идей Дарвина и доминирующей в современной биологии теории, которые как раз рассматривают эволюцию как ненаправленный процесс. Я думаю, что это одно из наиболее ярких подтверждений их некомпетентности.
Однако легко понять, почему креационисты пребывают в подобных заблуждениях. Дело в том, что они знакомы с эволюционной биологией почти исключительно по популярным книжкам и учебникам. Такие источники неизбежно дают упрощенное описание и очень медленно изменяются. Даже неоправданно медленно. В частности до сих пор множество изданий воспроизводит эволюционный ряд лошадей Марша, сопровождаемый описанием предполагаемого направления эволюции. Эта невероятно устаревшая схема издается, даже не смотря на то, что сейчас известно, что не все включенные формы принадлежат одной эволюционной линии. Черпая информацию из таких источников и игнорируя науку, креационисты оказываются в плену искаженных представлений о предмете, который они отрицают.
Любопытно, те креационисты, которые все же сталкивались с современными данными, пытались утверждать, что наконец-то ученые сами отвергли схему эволюции лошадей.
"Сегодня многие ученые, в том числе и убежденные эволюционисты, не считают схему, изображающую эволюцию лошади, научно достоверной. Однако эта схема по-прежнему используется в научных трудах и учебниках как фактическое доказательство теории эволюции.". |
| Дж. Маклин и др. "Очевидность сотворения мира". [129] |
Тем не менее следует отметить, что теперь уже сами эволюционисты ставят под сомнение достоверность традиционной схемы развития лошади, разработанной О. Ч. Маршем. |
| Ариэль Рос. "В начале..." [135] Внутренние ссылки опущены. |
Не выходя за рамки популярных источников, они не осознают, что каноническая схема была пересмотрена около 100 лет назад, еще до их рождения. В процитированной выше книге Рос ссылается на нескольких палеонтологов, чтобы подтвердить неприемлемость схемы эволюции лошадей XIX века. Но он не единым словом не упоминает, что эта схема заменена современными реконструкциями. Причем знания об эволюции лошадей стали более детальными, а количество переходных форм в лошадиной родословной только увеличилось.
Знакомство с изданиями креационистов показывает, что они представляют себе эволюцию как монотонное направленное изменение от одного типа строения в направлении другого ("от рыб к амфибиям", "от рептилий к птицам"). Креационист Алферов следующим образом говорит об утконосе:
"Так от кого же и к кому является утконос переходной формой: от рептилий к зверям или от зверей к птицам? Вдобавок, нужно учесть, что он так и не "перешел" от кого-то к кому-то, а просто остался утконосом". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
Сходным образом креационисты ссылаются на латимерию, отмечая, что этот представитель кистеперых рыб не обнаруживает стремления к выходу на сушу. Очевидно, они думают, что в процессе эволюции организмы должны обнаруживать стремление к определенному варианту анатомии, физиологии и образа жизни.
Они также часто подчеркивают, что палеонтологи так и не смогли обнаружить четких цепочек переходных форм от первого предка до последнего потомка, подразумевая, что это говорит об отсутствии переходных форм. Но такие цепочки имеют смысл только, если эволюция – направленный процесс. Палеонтологи обнаружили, что это ожидание не работает. Они показали на множестве разных групп, что, как правило, в истории группы появляется множество разных эволюционных линий, каждая из которых развивается по-своему, без предопределенного направления. Соответственно, палеонтологи сосредоточились на изучении разделения эволюционных линий и прослеживании их истории. Выстраивание в ряд предков и потомков уступило место вычислению степени родства. Рассмотрите еще раз кладограмму целурозавров. Со временем целурозавры разделились на отдельные эволюционные ветви, которые также продолжали ветвиться. В Меловом периоде некоторые эволюционные ветви так изменились, что оказалось крайне затруднительным установить их родственные связи.
То же можно сказать о ранней эволюции птиц. Сегодня мы знаем, что первые птицы дали начало, по крайней мере, двум большим группам птиц – веерохвостым (эволюция этой группы продолжается до сих пор) и противоположным птицам, все из которых вымерли в Меловом периоде. Каждая из этих больших ветвей разделилась на множество больших и маленьких. В каждой из этих линий разные виды воспользовались своим вариантом адаптации. Мы знаем, что в разных линиях утрата одних признаков и приобретение других происходили независимо. Так, одни птицы утратили зубы, а другие их сохранили. Одни птицы адаптировались к водному образу жизни, другие – к древесному, третьи потеряли способность к полету и превратились в мезозойских страусов. Современные отряды птиц (например, водоплавающие) частично возникли в результате этой радиации.
Кистеперые рыбы "избрали" аналогичный путь. В то время как одна эволюционная линия породила различных обитателей мелких пресных водоемов, перешедших в последствии к полуводному образу жизни; другая линия воспользовалась во многом противоположной возможностью – адаптацией к глубоководному образу жизни в море. Креационисты всерьез полагают, что все представители группы должны обнаруживать стремление к некоему "более прогрессивному" состоянию. Вероятно, такая точка зрения все еще преподавалась в период зарождения современного креационизма, когда наличие своей собственной экологии у каждого вида не было оценено по достоинству. Но даже к тому времени биологи, включая Феодосия Добжанского, Эрнста Майра и уже упомянутого Джорджа Гейлорда Симпсона, показали, что эта концепция игнорирует реальность и не может объяснить накопленные данные.
Интересно также обратить внимание, что сам аргумент о недостатке четких эволюционных реконструкций является попыткой уклониться от рассмотрения сути вопроса. Креационисты настаивают на отсутствии переходных форм. Когда появляются соответствующие примеры, они ссылаются на отсутствие четкого ряда от предка к потомку, уклоняясь тем самым от своего первоначального аргумента. Является ли археоптерикс дедушкой конфуциусорниса или же его троюродным прадедушкой, к сути дела отношения не имеет. Они все равно промежуточны, как не крути.
Столь же бессмысленны ссылки креационистов на побочные, тупиковые ветви эволюции. Они подразумевают, что если животное принадлежит к побочной ветви, то его вид не может считаться переходным. Но если есть побочная ветвь, то есть и основная. Подобные аргументы по существу ведут к признанию эволюции, удивительно, что креационисты не в состоянии это заметить.
5. Временные парадоксы.
Есть способ избавиться от переходных форм без учета их анатомии. Креационисты говорят: если переходная форма (предковая) жила в одно и то же время с потомковой, то ее нельзя считать переходной. То же верно, если предполагаемые потомки предшествуют предполагаемому предку, то есть если потомков находят в более древних отложениях. Я соглашаюсь со второй частью аргумента, но не соглашаюсь с первой.
Если мы обнаруживаем, что предполагаемый предшественник жил в одно время со своими эволюционными родственниками по нисходящей линии, мы с полным основанием можем считать его переходной формой. Креационисты думают иначе. Я долго не мог понять, почему. Такое представление абсурдно в высшей степени. Оно фактически предполагает, что как только один вид дает начало другому, первый должен автоматически вымереть. Но потомки европейских колонистов заселили Северную и Южную Америку, Австралию, а европейцы все еще здравствуют. То же верно для многих видов животных и растений, которые колонисты и их потомки завезли с собой. Действительно, после случая видообразования родительский вид может вымереть по разным причинам, но его вымирание не обусловлено просто появлением дочернего вида. Еще неправдоподобнее мысль, что то же предположение об автоматическом вымирании прародителя применимо к родам или семействам. Но креационисты никогда не объясняли причин для обсуждаемого постулата. Они обычно ссылаются на него как на нечто само собой разумеющееся, очевидное, не требующее пояснений. Иногда даже в риторической форме.
"Наконец, самое важное, что рядом с археоптериксом в тех же слоях найдены и обычные птицы, поэтому он никак не может считаться их предком". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
"все [о лошадях] предполагаемые "промежуточные формы" найдены погребенными в одних и тех же геологических формациях, которые указывают, что они жили в одно время и не могли быть предками друг друга". |
Пол Тейлор. "Сотворение. Иллюстрированная книга ответов" [138]. |
"Согласно теории эволюции, животные, поднимаясь по эволюционной лестнице, становились все более способными к выживанию. Почему же тогда сохранились "низшие" семейства обезьян, но не остался ни один представитель предполагаемых промежуточных форм, казалось бы, дальше продвинувшихся в эволюции?" |
"Жизнь - как она возникла? Путем эволюции или путем сотворения?" [126] |
"Если человек произошел от обезьяны, то почему обезьяны все еще существуют?". |
Понять смысл подобных выводов можно, если вспомнить, что креационисты не придают значения видообразованию или вообще отрицают его возможность. Они представляют себе эволюцию, как постепенное накопление новых особенностей в рамках старого вида, а механизм появления новых черт, как внутривидовую гонку вооружений. По представлениям креационистов, вымирать должны не виды или роды, а представители того же самого вида. Разумеется, они не могут сосуществовать со своими более продвинутыми потомками.
Фактически креационисты используют плохо осмысленную комбинацию ламаркизма и дарвинизма, игнорируют исследования в эволюционной биологии после 30-ых годов XX века и даже не осознают этого. Однако не все. Ошибочность рассматриваемого аргумента понята некоторыми креационистами. Наибольшая креационная организация – "Ответы в Бытии" – разместила на своем сайте перечень "Аргументов, которые, как мы думаем, креационисты НЕ должны использовать".
Среди них и аргумент о невозможности сосуществования предка и потомка. Поэтому нет нужды тратить время на пояснение ошибочности этого аргумента.
Так как на счет временных парадоксов? Можно ли привести примеры переходов, в которых переходные формы предшествуют потомкам. Археоптерикс и Гуанлонг входят в их число. Здесь аргумент временных парадоксов попросту не работает. Есть примеры сосуществования переходных форм и потомковых. И есть такие, где потомковые формы опережают предложенные переходные формы во времени. Если даже применить жесткий отсев переходных форм по времени, их число сократится, но они не исчезнут. Правда, креационисты оспорили бы предшествующий статус археоптерикса. Они утверждают, что современные птицы и сосуществовали с археоптериксом, и предшествовали ему. Вот на чем основаны эти утверждения.
"Что касается хронологий, то они ошибочны. Настоящие птицы существовали по меньшей мере одновременно с археоптериксом, так что последний вряд ли мог быть их предком." |
| Генри Моррис. "Библейские основы современной науки" [130]. |
Информация о нахождении современных птиц и археоптерикса в тех же отложениях основана на работе американского палеонтолога Джеймса Дженсена, который описал в 1981 несколько изолированных костей как новый род и вид древних птиц. Однако в 1989 он переклассифицировал экземпляры как останки динозавров и птерозавров. Удивительно, но 20 лет спустя креационисты все еще используют устаревшую информацию. Кроме того, находки Дженсена все же моложе археоптерикса и обнаружены в разных отложениях. Все экземпляры археоптерикса найдены в Германии, а находки Дженсена сделаны в Северной Америке.
Более серьезное возражение связано с обнаружением протоависа, предшествующего археоптериксу на 75 миллионов лет. Как указано выше, подобное открытие лишило бы археоптерикса его переходного статуса.
"В 1984 году в Техасе были найдены ископаемые останки птиц. Их возраст по определению эволюционистов на "миллионы лет" превышает возраст, приписываемый археоптериксу. И эти птицы ничем не отличаются от современных".. |
| Цит по: Даниил Сысоев "Летопись начала" [137]. |
"Кости и дополнительные свидетельства о других птицах [274] (которые, по словам их первооткрывателей-эволюционистов, были, по существу, идентичны птицам, живущим в настоящее время) были найдены в формациях, которые эволюционисты определяют как такие же "старые" или "старше", чем археоптерикс" |
Пол Тейлор. "Сотворение. Иллюстрированная книга ответов" [138]. |
"Самой удивительной находкой была окаменевшая птица, обладающая всеми признаками современных птиц, но возраст которой оценивается на десятки миллионов лет больше археоптерикса. Так что, переходная форма возникла намного позже самих птиц?" |
Уильям Гладсхауэр. "Как возник наш мир" [124]. |
"Не так давно были обнаружены еще два "предка" птиц... Один был найден примерно на том же уровне в геологической таблице, что и Archaeopteryx, а другой - чуть ниже. По поводу этих ископаемых видов среди ученых идет оживленный спор" |
Ариэль Рос. "В начале..." [135] Внутренние ссылки опущены. |
"Археоптерикс был самой настоящей птицей, а никакой не переходной формой. Об этом говорит и тот факт, что окаменелость, названная Proavis (sic), очень похожая на современную птицу, была обнаружена в геологической колонне гораздо ниже, чем археоптерикс" |
Дэвид Роузвер. "Наука о сотворении мира, доказывающая правоту Библии" [125]. |
Экземпляры, связанные с протоависом были раскопаны американским палеонтологом Санкаром Четарджи и его студентами в 70-ых годах XX века. Анализируя материал, Четарджи пришел к выводу, что останки принадлежат примитивной веерохвостой птице, которую он назвал протоавис. Однако мало кто согласится с этой интерпретацией. Большинство палеонтологов отметили, что кости сохранились плохо, и отделены друг от друга. Когда ископаемые останки заключены в кусок породы (как это имеет место, например, с экземплярами археоптерикса), значительно проще установить принадлежность материала одному организму. Ученые указали, что некоторые скелетные особенности протоависа типичны не для птиц, а, скорее, для других групп, например, динозавров. Поэтому в большинстве палеонтологи пришли к выводу, что находки Чатерджи представляют собой химеры. Так палеонтологи называют соединения останков разных животных, в том числе из разных систематических групп. Однако Чатерджи не согласился, и все остались при своих точках зрения.
|
| Берлинский экземпляр археоптерикса. Поскольку останки заключены в пластине известняка, можно убедиться, что все кости принадлежат одному животному. Экземпляры протоависа представлены изолированными элементами. |
Вновь креационисты безоговорочно следуют за мнением ученого, которого критикует большинство его коллег. Категоричность креационистов не оправдана и потому, что основные сторонники птичьего статуса протоависа – Чатерджи и российский палеонтолог Евгений Курочкин – не соглашаются друг с другом. Чатерджи считает, что кости протоависа сохраняют бугорки для закрепления перьев, а их строение говорит о способности к полету. Курочкин отвергает эти выводы. По его мнению, протоавис не мог летать [127]. И наконец, Чатерджи и Курочкин соглашаются с гипотезой о происхождении птиц от звероногих динозавров, хотя они и предлагают свои варианты этой гипотезы.
Описание протоависа как современной птицы – преувеличенный слух, возникший, несомненно, в результате многократных заимствований информации одними креационистами у других. Птиц делят на три группы: ящерохвостые, противоположные и веерохвостые. Только представители последней группы выжили до наших дней. Чатерджи утверждает, что протоавис ближе всего к веерохвостым. Но из этого не следует, что всякая веерохвостая птица – современная. Фактически все ранние веерохвостые птицы отличны от современных, они не принадлежат ни к одному из отрядов современных птиц. Даже первые представители современных отрядов отличаются от сегодняшних птиц. Если протоавис был современной птицей, почему креационисты не сообщают к какому отряду и семейству он относится? Они никогда не будут в состоянии обеспечить такую информацию. Фрагментарность экземпляров протоависа просто не даст возможности сделать такую оценку.
Однако ирония ситуации для креационистов состоит в том, что и Чатерджи, и Курочкин считают протоависа не просто вымершей птицей, а одной из переходных форм. Действительно, оба ученых описали и птичьи, и рептилийные особенности протоависа. К примеру, шейный позвоночник протоависа больше соответствует строению цератозавров, а не птиц. В терминологии креационистов - это пример "полуразвитой" части тела, которой не может быть. Можно также упомянуть наличие у протоависа четырех пальцев в передней конечности. Одна эта характеристика исключает его из современных птиц, поскольку все птицы, включая археоптерикса и его ближайших родственников, трехпалы.
|
| Реконструкция скелета протоависа Санкара Четарджи. Недостающие элементы обозначены только контуром. Можно видеть четыре пальца передних конечностей, нередуцированный хвостовой отдел позвоночника и соединение шеи с черепом сзади. Эти особенности не характерны для современных птиц. |
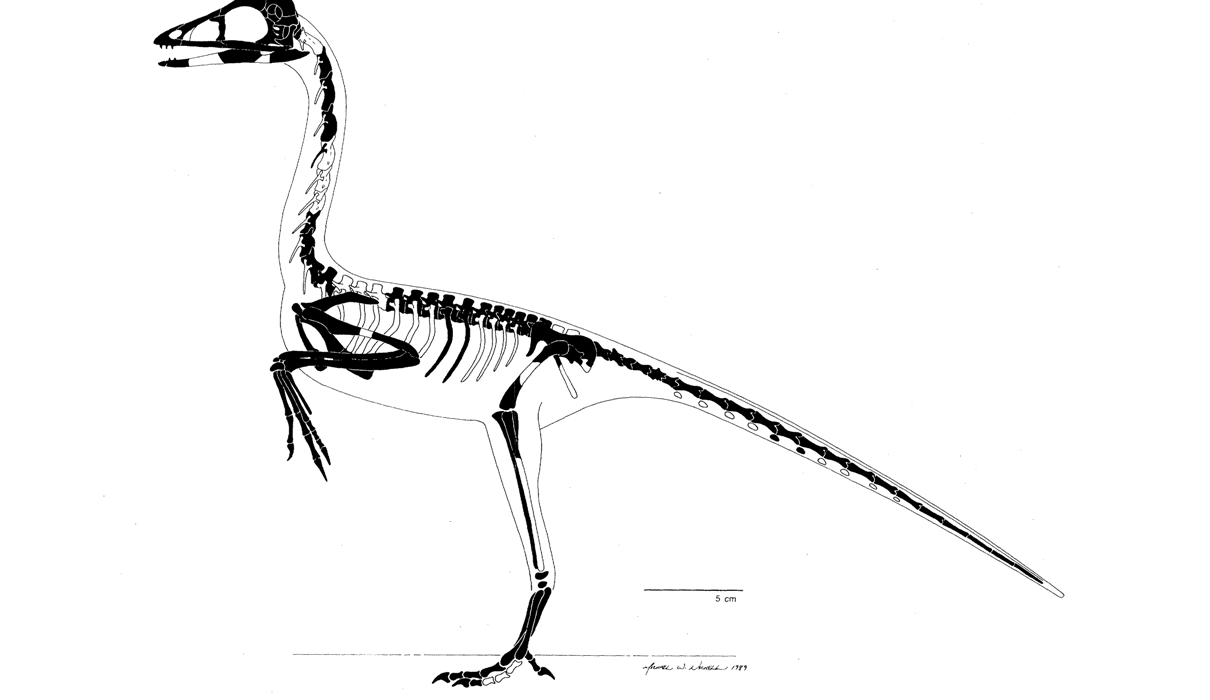
Итак, креационисты ничего не выигрывают от признания протоависа. Если они хотят игнорировать работу других палеонтологов, чтобы избавится от археоптерикса, то просто получают вместо одной переходной формы другую.
Поскольку проблематично определить, связан ли протоавис с пернатыми или нет, археоптерикс остается самой ранней птицей, и его переходной статус более чем очевиден. А предполагаемые временные парадоксы, которые могли бы лишить его этого статуса, отсутствуют.
6. Где миллионы переходных форм?
Креационисты часто подчеркивают, что, если эволюция верна, то должно быть очень много (возможно, миллионы переходных форм), хотя нет ни одной. Это настолько странный аргумент, что удивительно, почему он вообще существует. Сами креационисты почти всегда указывают, что палеонтологи описали по окаменелостям 250,000 видов. Казалось бы, креационисты должны осознавать непреодолимые математические препятствия для поиска миллионов в тысячах. Но, возможно, речь о том, что экземпляры переходных форм должны быть множественными. Это ожидание как раз соблюдено. К примеру, известно десять скелетных экземпляров археоптериксов и буквально сотни экземпляров конфуциусорнисов. Поэтому рассматриваемый аргумент не заслуживает дальнейшего внимания.
7. Их нет: признают даже эволюционисты.
Креационисты дают понять, что отсутствие переходных форм – простой, очевидный, неоспоримый факт. Чтобы рассеять сомнения в этом, они приглашают в качестве свидетелей тех, кто, казалось бы, в последнюю очередь признает такой факт – эволюционистов. Уж если эволюционисты согласятся с отсутствием переходных форм, то лучшего свидетельства и не нужно. В самом деле, креационисты дают понять, что палеонтологи нехотя признали неудачу с поиском переходных форм, и чтобы решить проблему, мучавшую еще Дарвина, стали придумывать теории, по которым переходы были, но окаменелостей от этих процессов остаться не могло. Так, по убеждению креационистов, появилась теория прерывистого равновесия. Креационисты пишут:
"Отсутствие ископаемых переходных форм вынудило эволюционистов предложить в конце 70-х годов новую модель эволюции, не требовавшую наличия промежуточных звеньев. Эта модель - теория прерывистого равновесия - утверждает, что эволюционные изменения происходили в геологическом смысле настолько быстро, что ископаемые свидетельства просто не успевали сохранятся". |
Кен Хэм и др. "Книга Ответов" [140]. |
"Сторонником этого взгляда выступает известный биолог Стивен Джей Гоулд. Идея заключается в том, что переход от одних отрядов и классов к другим происходит очень быстро, буквально в несколько поколений в одной обособившейся популяции предков. Такое быстрое изменение не оставляет следа в окаменелостях. Быстрое изменение сменяется длительным периодом накопления непроявляемых мутационных изменений, пока их количество не перейдет в качество и вновь в отдельной малой популяции, где наследственных изменений накопилось много, произойдет новый скачок эволюции". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
"Логика эволюционистов поразительна. Виды не подвергаются сильным изменениям на протяжении "сотен тысяч поколений" поэтому "эволюция проходит в основном быстро". Замечательный пример слепой веры в эволюцию. Так как нет никаких свидетельств ее существования ни в мире живых организмов, ни останках умерших, значит, она происходила чуть ли не мгновенно и в малых популяциях, "практически не оставляя следа в окаменелостях”". |
| Генри Моррис. "Библейские основы современной науки" [130]. |
"Так называемая теория "прерывистого равновесия", разработанная Элдреджем (Eldredge) и Гоулдом (Gould), гласит, что в малых изолированных сообществах происходили быстрые, скачкообразные перемены... Эти изменения протекали столь стремительно, что не оставили следа в летописи окаменелостей. Длительные периоды равновесия внезапно прерывались краткими, но глобальными переменами. Подобные гипотезы возможны потому, что геология не дает никакой информации о переходных формах". |
| Дэвид Роузвер. "Наука о сотворении мира, доказывающая правоту Библии" [125]. |
"В "новой" палеонтологии (основное понятие - "прерывистое равновесие" ... официально признано систематическое отсутствие промежуточных звеньев, и предпринимается попытка построить новую эволюционную модель, которая больше соответствовала бы палеонтологическим данным... Под давлением фактов палеонтология приходит в своем развитии к позиции, которая раньше подвергалась сильным нападкам со стороны теоретиков, сторонников эволюционного учения, к позиции, сформулированной в теории сотворения, сторонники которой всегда утверждали: "Никаких промежуточных звеньев не существует"". |
| Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
В действительности не могло быть ничего более далекого от правды. Давайте рассмотрим, что фактически говорили Гулд и Элдридж.
По их мнению, большинство ученых осознанно или даже бессознательно использовали неправдоподобную модель эволюции. Эта неправильная модель сводится к четырем постулатам.
1. Новые виды появляются путем преобразования представителей популяции старого вида. Мы уже сталкивались с этой идеей выше.
2. Появление вида происходит медленно и постепенно.
3. Появление вида происходит в пределах больших популяций.
4. Появление вида происходит в пределах всей территории, занимаемой популяцией.
Гулд и Элдридж утверждали, что сформулированная модель, с одной стороны, опровергается палеонтологической летописью, а с другой, не соответствует тому, что нам известно о видообразовании. Вместо описанной модели они предложили прерывистое равновесие, основанное на изучении популяций в современном мире.
Прежде всего, Гулд и Элдридж утверждали, что новые виды возникают не путем преобразования старых, а путем раскола или обособления. Дочерний вид занимает свою экологическую нишу и сосуществует с родительским. Как мы уже видели, эта идея не нова. Симпсон объяснял ее неспециалистам еще в 50-ых годах.
Далее, в модели Гулда и Элдриджа видообразование происходит в маленьких географически изолированных популяциях. Неудивительно, в 70-ых, когда Гулд и Элдридж создавали свою модель, этому механизму видообразования отдавалось предпочтение. Кроме того, адаптация происходит в течение видообразования. Если адаптация и география позволяют увеличение численности в популяции нового вида, то территория обитания вида быстро расширяется.
Учитывая сказанное, мы теперь можем вернуться к аргументам креационистов. Есть три ошибки в их понимании идеи прерывистого равновесия.
Во-первых, прерывистое равновесие не призвано заменить все прочие модели видообразования. Гулд и Элдридж подчеркивали, что другие механизмы также возможны.
Во-вторых, прерывистое равновесие основано не только и не столько на попытке отыскать объяснение недостатку переходных форм. В первую очередь модель прерывистого равновесия была призвана обосновать, почему не следует ожидать, что по окаменелостям можно проследить, как один вид постепенно перерождается в другой. Креационисты думают, что прерывистое равновесие не имеет никаких данных в свою поддержку, и было принято учеными исключительно потому, что позволяет объяснить отсутствие переходных форм. Это неверно. Гулд, Элдридж и другие ученые пытались найти такие исключительные условия, которые позволили бы по палеонтологическим данным изучить эволюцию видов в течение долгого времени и выявить, какая модель правильнее. Ответ мало похож на сенсацию: накопленные данные показывают, что в прошлом происходили и прерывистое равновесие, и градуализм.
Наконец, в-третьих, - и это самое главное – прерывистое равновесие объясняет видообразование, а не эволюцию крупных групп (родов, семейств, отрядов и пр.), как ошибочно думают креационисты. В модели прерывистого равновесия быстро происходит видообразование, а не происхождение новых типов строения. Гулд и Элдридж действительно говорили о недостатке переходных форм, но на уровне межвидовой эволюции и только. Гулд объясняет:
"Поскольку мы предложили прерывистое равновесие, чтобы объяснить тенденции [в палеонтологической летописи], меня приводит в ярость то, что креационисты снова и снова цитируют нас (не знаю, преднамеренно или по глупости) в качестве подтверждения отсутствия переходных форм в окаменелостях. Переходные формы, как правило, отсутствуют на уровне вида, но они в изобилии присутствуют между группами более высокого ранга... Продолжая искажение, некоторые креационисты приравняли теорию прерывистого равновесия с карикатурой взглядов выдающегося раннего генетика Ричарда Голдшмидта". |
| Stephen Jay Gould. "Evolution as Fact and Theory" [33]. |
Итак, попытка призвать эволюционистов в свидетели, чтобы подтвердить отсутствие переходных форм, основана на недоразумении. Но каков источник этого недоразумения? Изучая публикации креационистов, я пришел к выводу, что они почерпнули сведения о прерывистом равновесии из вторичных или третичных источников и были не в состоянии эти сведения понять. В этом можно убедиться, читая их издания. Так "отец современного креационизма" Генри Моррис, говоря о прерывистом равновесии, пишет:
"...многие палеонтологи новой школы взяли на вооружение марксистскую систему ... "прерывистого равновесия" ... Эта странная теория (называемая также теорией "обнадеживающего чудовища")..." |
| Генри Моррис. "Сотворение и современный христианин" [131]. |
Очевидно, он не понимает, что это две разные теории, не связанные друг с другом. Гипотеза "обнадеживающего монстра" была предложена Гольдшмидтом в 40-ых годах, а Гулд и Элдридж писали в 70-ых. Существенно, что Гольдшмит также говорил о видообразовании и не более, а вовсе не о том, что рептилия отложила бы яйцо, из которого вылупилась бы птица. Кроме того, к 70-ых годам гипотеза Голдшмита уже была отклонена.
Еще более интересное рассуждение мы находим у креациониста Роса. Он описывает прерывистое равновесие следующим образом:
"Они [эволюционисты] также заявляют, что изменения происходили в малых популяциях, в которых шансов на сохранение промежуточных форм в качестве окаменелостей должно быть меньше, то есть речь идет о модели прерывистого равновесия. Может быть, чем-то подобным и можно объяснить недостаток промежуточных звеньев между близкородственными видами, но в отношении гораздо более существенной проблемы — отсутствия таковых между основными группами организмов — данные предположения не имеют силы". |
| Ариэль Рос. "В начале..." [135] Внутренние ссылки опущены. |
Совершенно ясно, что Рос правильно догадался, в чем смысл модели, но явно не знает, что этот смысл присутствовал в ней изначально.
Иронично, ровно ту же ошибку делают более информированные креационисты Юнкер и Шерер:
"Может быть, новая теория сможет вполне достоверно ответить на вопрос, почему между отдельными видами и родами не имеется переходов... Однако эта теория не объясняет, как в царстве живых организмов смогли возникнуть большие основные типы (млекопитающие, птицы, четвероногие, а также отряды и семейства), так как подобные скачки развития в течение кратчайшего геологического периода ... даже и в малых популяциях представить невозможно". |
| Райнхарт Юнкер, Зигфрид Шерер. "История происхождения и развития жизни" [141]. |
Здесь мы снова сталкиваемся с ярким свидетельством некомпетентности креационистов.
|
Фрагмент титульного листа той самой работы, в которой Гулд и Элдридж предложили модель прерывистого равновесия. Красным выделено название рубрики, под которой опубликована статья – "видообразование". Любой, кто хотя бы взглянул на титульный лист, даже не читая текста, не совершил бы той ошибки, которую креационисты повторяли десятилетиями и продолжают повторять. |

Стоит вспомнить Курта Вайса. По контрасту с другими креационистами он упоминает прерывистое равновесие именно в связи с переходными формами на уровне вида и не более (см. раздел о мозаичных формах). Он же признает существование форм, которые справедливо можно расценить как переходные. Очевидно, учеба в Гарварде у одного из авторов теории прерывистого равновесия не прошла даром, Вайс в отличие от других креационистов знаком с предметом.
Любопытно, что, не осознавая своей некомпетентности, креационисты делают вывод, будто эволюционисты придумывают разные теории только для того, чтобы отклонить веру в бога. Генри Моррис так писал о модели прерывистого равновесия:
"Она полностью основана на отсутствии доказательств! Ее единственное достоинство состоит, видимо, в том, что она прекрасно соответствует марксистской философии многих ее ведущих сторонников, и что более важно, дает предлог, чтобы продолжать пренебрегать потрясающими свидетельствами креационистов, полученными ими на основании изучения окаменелостей". |
| Генри Моррис. "Сотворение и современный христианин" [131]. |
Очевидно, Моррис знаком с прерывистым равновесием из вторых, третьих рук. В противном случае он не пропустил бы десять страниц описания улиток и трилобитов, которое Гулд и Элдридж приводили в подтверждение своей модели. Разумеется, он также знал бы и о множестве других подтверждений, изданных палеонтологами позднее. Действительно, в одной из своих книг Моррис упоминает источник, из которого он почерпнул сведения о прерывистом равновесии. Это новостное сообщение о конференции палеонтологов [131].
Похожую конспирологию развивает российский креационист Алферов:
"Обратим внимание и на то, что для исключения Бога из картины мира люди готовы придумать самую фантастическую нелепость и невероятицу вроде этих "обнадеживающих уродов", которые позволили совершенно случайно создать все многообразие земной жизни. В такую чушь верить – это научно, а верить во Всемогущего и Всеведущего Творца – почему-то до сих пор считают недопустимым". |
| Алферов. "Православное мировоззрение и современное естествознание" [120]. |
Но если вдуматься, креационисты не осознают, что ситуация, в которой они оказались, еще плачевнее. Гулд и Элдридж говорят о недостатке переходных форм между видами. Это такие изменения, которые креационисты называют "микроэволюцией" и настоящей эволюцией не считают. Если бы подразумеваемые окаменелости существовали, креационисты никогда не согласились бы считать их свидетельством эволюции. С другой стороны, Гулд и Элдридж подтверждают существование тех переходных форм, которые, по словам креационистов, не существуют и не могут существовать.
В дополнение следует сказать несколько слов обо всей аргументации креационистов, касающейся переходных форм.
Прежде всего, обсуждая переходные формы, креационисты говорят о чем угодно, кроме того, что делает их переходными – их анатомии. Разделы, которые креационисты посвящают окаменелостям, часто объемны, но анатомические сравнения почти полностью отсутствуют. К примеру, книга "Жизнь, как она возникла" посвящает большой раздел (11 абзацев) вопросу невозможности эволюции птиц. Все же в этой связи археоптерикс кратко упомянут в нескольких предложениях в конце. В другом варианте креационисты сваливают в кучу массу несвязанных данных, цитат из устаревших источников, популярных изданий. Это дает возможность креационистам уклониться от упоминания особенностей строения вымерших организмов, которые они не могут объяснить в своей картине мира. Не будет преувеличением сказать, что креационисты бегут от анатомии как черт от ладана.
Сколько бы креационисты не говорили о переходных формах, они просто не описывают их. Фактически креационисты создали искусную форму риторики, которая позволяет много говорить об окаменелостях, на деле уклоняясь от рассмотрения их промежуточной природы. Прекрасный образец такой риторики можно найти в цитировавшейся выше статье креационного эксперта по окаменелостям Дуэйна Гиша. На примере его сочинения можно даже оценить эту риторику количественно! Так Гиш приводит 26 цитат из разных источников для подтверждения своего тезиса об отсутствии переходных форм и только 1 крайне скудное и ошибочное анатомическое сравнение при упоминании археоптерикса. Это сравнение было рассмотрено выше. Гиш избегает упоминания множества рептилийных характеристик археоптерикса, а также комплексной оценки его анатомии. При упоминании других форм он просто голословно утверждает, что они не имеют промежуточного характера без анализа анатомии. Итак, 26 цитат vs. 1 несостоятельное сравнение.
Иронично, некоторые креационисты введены в заблуждение своими собственными коллегами. Те из них, кто изучают эволюционную биологию по произведениям других креационистов, кажется, действительно убеждены, что краткие описания вымерших животных, прочитанные ими, – это все, что можно сказать о переходных формах. Они повторяют эти описания, будучи всерьез уверенными, что ученые обосновывают теорию эволюции на столь шатких и сомнительных данных. Следующим шагом, по всей видимости, становится убежденность в том, что ученые поступают так из злого умысла, из-за желания отрицать религию. Как выразился один российский креационист, говоря о переходных формах: "Это все сказки (sic) сочиненные для того, чтобы не слышать голоса Божия" [137].
Другая неизменная тенденция у креационистов – рассмотрение переходных форм изолированно. Тот же археоптерикс у них представляет эволюцию птиц один одинешенек, хотя цитируемые креационистами источники описывают и другие переходные формы. Насколько мне известно, ни один креационист не привел целиком тот упоминаемый Куртом Вайсом ряд переходных форм, связанный с эволюцией птиц, и не показал, на основе строения скелета, почему они не являются промежуточными формами, последовательно иллюстрирующими этапы эволюции класса птиц. Разумеется, результат такого сравнения испортил бы всю картину.
Отрицая существование переходных форм, креационисты подчеркивают существование четких промежутков между отдельными систематическими группами. Однако они никогда не определяют эти промежутки на основе анатомии, что весьма мудро с их стороны по двум причинам. Во-первых, в некоторых случаях такой промежуток невозможно найти. Попытка произвольно обозначить пробел привела бы к указанию на переходную форму, которая этот разрыв закрывает. Во-вторых, даже если разрыв существует, не исключено, что переходная форма, заполняющая его будет обнаружена в последствии. Подобная ситуация бросила бы тень на креационизм.
Впрочем, можно вспомнить одно исключение из этой тенденции. Марвин Лубенов - магистр богословия, считающийся у креационистов экспертом по разделению гоминидов на обезьян и людей, в своих изысканиях пришел к заключению, что нормальный человеческий объем мозга не ниже 700 кубических сантиметров. Заключение не лишено смысла, поскольку мозг современных горилл может иметь сопоставимый размер. И хотя граница здесь прочерчена по одному признаку, да и вряд ли Лубенов хотел обрисовать промежуток, который никогда не будет закрыт, все же перед нами фактически количественное разделение. Когда Лубенов изучал материал по человеческой эволюции, не существовало ни одного черепа гоминида, объем которого находился бы в диапазоне от 600 до 700 кубических сантиметров. Однако через 10 лет после публикации книги Лубенова, были описаны останки гоминидов из Грузии. Три черепа, найденные в одном местоположении и принадлежащие представителям одного вида, закрывают предполагаемый пробел. Черепа имеют объем 600, 650 и 780. Любопытно, креационисты главным образом обходят молчанием эти находки, а если и обсуждают, то уклончиво (включая Лубенова), не предлагая обоснований для своих выводов. Даже более примечательно, что Лубенов в основном тексте переиздания своей книги просто обошел молчанием упомянутые экземпляры [26, 28, 29].
|
Экземпляр D2282. Средний из трех черепов, найденных в Грузии. Оцененный объем мозга 650 кубических сантиметров. |
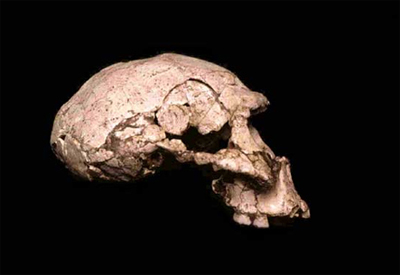
Еще один пример анатомического промежутка, которым можно воспользоваться, описан в книге "Жизнь, как она возникла..." Правда, авторы не намеревались описывать пробел, для которого никогда не будет переходных форм. Они пытаются продемонстрировать, что рептилий и птиц разделяет пропасть, эволюционный переход через которую просто немыслим. Так что же это за пропасть?
"Пресмыкающиеся – холоднокровные животные... высиживать своих птенцов приходится только птицам... чтобы птицы могли высиживать птенцов, эволюция должна была снабдить их новыми инстинктами, необходимыми для постройки гнезда, для насиживания яиц, для вскармливания птенцов... Перья – достояние только одних птиц... кости птиц тонкие и полые... благодаря ... воздушным мешкам, птицы поглощают из воздуха кислород гораздо эффективнее... острота зрения... у них лишь четыре пальца, а не пять, как у пресмыкающихся... пресмыкающиеся имеют трехкамерное сердце, а птицы – четырехкамерное... птиц от пресмыкающихся отличают клювы..." |
| "Жизнь - как она возникла? Путем эволюции или путем сотворения?" [126] |
Учитывая сказанное выше, легко заметить, что предполагаемая авторами пропасть просто не существует. Большую часть перечисленных особенностей птицы не "изобрели", а просто унаследовали от своих предков: перья, постройка гнезд, забота о потомстве, полые кости, четыре пальца задних конечностей, даже, возможно, насиживание яиц и воздушные мешки. Относительно холоднокровности авторы сообщают устаревшие сведения. В действительности динозавры обладали промежуточным метаболизмом. Метаболизм маленьких динозавров был ближе к птичьему, но по мере взросления становился более рептилийным. Форму сердца нельзя вывести из палеонтологических данных. Остроту зрения у вымерших рептилий точно установить невозможно, хотя ближайшие родственники птиц были хищниками, зрение для которых представляло большую важность, а их зрительные области мозга были увеличены. А вот клювов у самых ранних птиц не было. Любопытно, сразу же за упоминанием клювов, авторы обсуждают археоптерикса, не сообщая читателю, что у него-то как раз клюв отсутствовал. При этом авторы явно относят археоптерикса к птицам, что, казалось бы, не соответствует их собственным стандартам.
Креационисты весьма нетерпеливы, когда дело касается переходных форм. Я преднамеренно сосредоточил внимание на происхождении и эволюции птиц, поскольку свидетельства в пользу эволюции в данной области существовали давно, но в то же время исследователи столкнулись с существенными трудностями. Поэтому вопрос происхождения птиц позволяет демонстрировать, как работает наука, как выдвигаются, проверяются гипотезы, и как накапливаются свидетельства в их поддержку. Многие выдающиеся биологи, начиная с Томаса Хаксли, прогнозировали обнаружение переходных форм в ранней эволюции птиц. Однако многие из них ушли из жизни, прежде чем соответствующие открытия были сделаны. Самые поразительные экземпляры давно ожидаемых переходных форм были найдены только на рубеже XX и XXI веков в Китае. Эти окаменелости, которых, по словам креационистов, не должно быть, всегда были с нами. К счастью, они спокойно пережили бурный XX век. Их не было только лишь по той причине, что никто не проводил раскопок там, где они были захоронены. И хотя сегодня нет недостатка в "переходных" птицах, палеонтологи рассчитывают найти еще больше. Промежуток между археоптериксом и его ближайшими родственниками исчез совсем недавно. Но предшественники археоптерикса пока плохо известны. Палеонтологи очень хотели бы найти участок, столь же богатый окаменелостями рептилий и птиц, но средне- или позднеюрского возраста. Если он существует, пока никто не проводил там раскопок. И никак нельзя знать заранее, где он. Хотя есть гипотеза, что ранние этапы эволюции птиц были связаны с Азией. Будущее покажет, так это или нет.
Я не думаю, что креационисты смогут достичь большего успеха в вопросе переходных форм в будущем. У них были десятилетия на то, чтобы проделать необходимую работу. В сущности, они даже не попытались, если не считать ссылок на мозаичные формы. Комментарии креационистов о переходных формах сводятся к использованию устаревших источников; цитированию популярных книг и поиску цитат, которые они могут истолковать в свою пользу. В трудных случаях лучшее, что могут придумать креационисты - просто игнорировать неудобную информацию. Приводимые ими сведения показывают отсутствие знания анатомии и свидетельствуют, что креационисты не в состоянии отличить одну кость от другой.
Но следует вспомнить, что для креационистов отсутствие переходных форм – гарантия их религиозной веры. Некоторые креационисты вносят отсутствие переходных форм в список свидетельств в пользу существования Бога-Творца. Таким образом, находки палеонтологов могут представлять угрозу для всей их религии.
Самое удивительное в спорах о вымерших животных то, насколько они влиятельны после смерти. На основе их биологии некоторые люди на полном серьезе предлагают решать, какие ценности правильные, а какие нет.
И все же: можно ли продемонстрировать хоть один пример переходной формы? Ответ будет зависеть от того, знаете ли вы анатомию. В противном случае вам придется поверить на слово тому, кто вызывает у вас больше доверия.
Работая над темой, я пришел к убеждению, что популяризация переходных форм в противоположность своим целям создает у публики превратное впечатление об их характере, и поэтому должна быть изменена. Необходимо отказаться от представления переходных форм как наглядного свидетельства эволюции. Напротив, следует оговорить сложность работы с окаменелостями, необходимость специального образования и опыта. Конечно, популярная статья или книга не может отразить все анатомические сложности, но их существование важно подчеркнуть, объяснить, сколько информации осталось за скобками. Всегда можно указать, какие части тела (скелета) информативны для прослеживания родословной, сколько характеристик палеонтологи используют в исследованиях. Без такой коррекции популяризация изучения эволюции будет порождать искаженные представления.
Приложение 1. Эволюция перьев.
Традиционно биологи считали, что перья развились из чешуи рептилий. Для этой гипотезы есть несколько оснований. Во-первых, чешуи и перья – производные кожи. Во-вторых, и чешуи, и перья образованы белками одного класса – кератинами. Однако последнее обстоятельство не обязательно подразумевает происхождение одного из другого. Из кератинов также состоят волосы, но волосы не гомологичны ни перьям, ни чешуе. До настоящего времени палеонтология не обеспечила никаких подтверждений преобразований чешуи в перья. Креационисты постоянно указывали и указывают на это.
"Впечатляющим свидетельством происхождения птиц от рептилий (динозавров) были бы чешуйки, наполовину превратившиеся в перья". |
Кен Хэм и др. "Книга Ответов" [140]. |
"Еще никому не удалось обнаружить ископаемые промежуточные формы с полуразвитыми крыльями или перьями". |
Уильям Гладсхауэр. "Как возник наш мир" [124]. |
"...переходные формы с недокрыльями и полуперьями так и не были найдены." |
| Дуэйн Гиш. "Сотворение мира, эволюция и исторические свидетельства." [123] |
"Никаких животных с полуперьями-получешуёй не обнаружено". |
| Сергей Вертьянов. "Общая биология" [122]. |
"Если бы такой процесс протекал "по Дарвину", должно было остаться множество промежуточных форм с полу-крыльями и полу-перьями. В действительности никакой такой последовательности не найдено". |
Алферов. "Православное мировоззрение и современное естествознание" [120]. |
"...нет животных – ни живых, ни окаменелых – которые обладали бы переходными изменяющимися структурами (например, чешуя превращается в перья...). |
Генри Моррис. "Сотворение и современный христианин" [131]. |
"Здесь нет и речи о том, что это стадия на полпути между чешуей пресмыкающегося и птичьим пером, чего можно было бы ожидать у такого промежуточного существа. Нигде в летописи окаменелостей нет никаких признаков такой связи между чешуей и пером". |
Сильвия Бейкер. "Камень преткновения: Верна ли теория эволюции?" [121] |
Птичьи перья предположительно развились из чешуи какой-то древней рептилии. Всякий, кто изучал птичье перо под микроскопом, знает, что оно обладает сложной и узко специализированной структурой. Может быть, длительный процесс возникновения всех ее составляющих из чешуи ... оставил свои следы в горных породах? Пока ничего подобного не обнаружено. |
| Ариэль Рос. "В начале..." [135] |
Вероятное объяснение того, что мы не находим "полу-перьев" состоит в том, что шансов на их сохранение в окаменелостях практически нет. Следы перьев могут сохраниться только в уникальных условиях. Перья археоптерикса сохранились благодаря литографическому известняку, свойства которого приближаются к свойствам стекла. Поэтому литографический известняк весьма востребован. Без промышленного спроса на него мир, скорее всего, никогда бы и не слышал об археоптериксе. Но даже в таких, казалось бы, идеальных условиях перья сохраняются не всегда. Креационисты часто преувеличивают сохранность оперения археоптерикса. Часть перьев действительно сохранена превосходно, но не все. Некоторые экземпляры археоптерикса сохранились почти без перьев. И все же на сегодняшний день в распоряжении палеонтологов есть нечто большее, чем ссылки на бедность палеонтологической летописи. Потребовались годы, чтобы изучить и оценить важные окаменелости.
В начале 90-ых годов биологи Алан Браш и Ричард Прам использовали новые лабораторные методы, чтобы изучить биологию перьев и чешуй рептилий. В итоге они пришли к выводу, что предполагаемые сходства первых и вторых являются поверхностными. Это неизбежно вело к заключению, что перья не возникли путем модификации чешуй. Оба биолога независимо друг от друга предложили очень похожие модели эволюции перьев. В соответствии с этими моделями эволюция перьев началась с появления особого кожного образования – мешочка пера, из которого собственно и вырастают перья у современных птиц. А прото-перья представляли собой отростки в виде нитей [68, 69, 70]. В то же самое время палеонтологи описали нечто такое, что до того момента никому не удавалось обнаружить.
В 1998 палеонтологи описали звероногого динозавра со следами ранее не известного наружного покрова, образованного гибкими волокнами, а чуть позже представили миру двух звероногих с перьями современного типа строения. Поскольку наличие перьев у динозавров теперь было установлено, а звероногие с волокнистыми покровами принадлежали к той же эволюционной ветви, что и ранние птицы; авторы высказали осторожное предположение о том, что найденные волокна могли быть прото-перьями. Однако вскоре были высказаны сомнения. Скептики предположили, что пернатые динозавры на самом деле древние птицы, утратившие способность к полету, а покровные волокна в действительности – следы коллагена – белка соединительной ткани. Но новые и лучше сохраненные экземпляры сомнения не подтвердили.
На сегодняшний день сложилась следующая картина. В трех наиболее близких к птицам семействах целурозавров описаны представители с типично птичьими перьями. Описан даже неизвестный у современных птиц тип пера. В свою очередь волокнистые покровы обнаружены в четырех из пяти групп, связанных с птицами более дальним родством (см. кладограмму целурозавров). Таким образом, эволюционное древо целурозавров настоятельно указывает на возможность, что найденные покровные волокна могут быть предшественниками перьев. Учитывая эти данные, многие исследователи экспериментально определили их как прото-перья. Однако сделать более определенный вывод на основании эволюционных отношений нельзя. С помощью филогенетических деревьев можно прогнозировать неизвестные особенности биологии, но нельзя их диагностировать.
Поэтому, чтобы подтвердить происхождение перьев от нитевидных волокон, нужно обнаружить их структурное родство – гомологию. Сделать это очень проблематично по той простой причине, что покровные волокна целурозавров не обладают структурой. Действительно, какую структуру можно выявить у нити длинной 1-4 сантиметра? По мере описания новых экземпляров постепенно выяснили, что некоторые из этих волокон были полыми, что наводит на мысль о полом стержне пера. Выяснили также, что некоторые подобные волокна ветвились, что напоминает о бородках пера. Но все же самое сильное свидетельство родства перьев и покровных волокон состоит в том, что известны формы, у которых одновременно присутствовали оба типа покровных структур. Однако все это лишь косвенные свидетельства, которые не являются однозначными.
Самое важное свидетельство гомологии между перьями и покровными волокнами целурозавров появилось после работы аспиранта Джекоба Винзера, который вообще не планировал изучать динозавров. Винзеру первому пришла в голову идея поискать в экземплярах древних перьев микроскопические структуры, придающие им окраску. Эти структуры называются меланосомы. Название происходит от пигмента меланина, который меланосомы содержат. Меланосомы – клеточные органеллы, которые производятся специальными клетками и входят в состав пера во время его формирования. В результате перья приобретают окраску.
Винзер вместе с несколькими учеными проанализировали экземпляры древних перьев под электронным микроскопом и обнаружили меланосомы, что позволило им даже восстановить окраску перьев. Тот же подход был применен к изучению перьев птиц и покровных волокон звероногих, найденных в пределах одной формации. Окаменелости не подвели: перья птиц и динозавров, а также покровные волокна содержали два отличных типа меланосом, присутствующих в перьях современных птиц [106]. Вскоре меланосомы обнаружили также в перьях археоптерикса [7]. Таким образом, гипотеза о происхождении перьев от трубчатых покровных волокон получила лучшее на сегодняшний день подтверждение.
|
| Меланосомы в окаменевшем пере начала мелового периода. a. Один из исследованных образцов. Стрелка указывает точку, представленную ниже в увеличении. b. и c. Окаменевшее перо в сканирующем электронном микроскопе. Стрелки указывают меланосомы. |
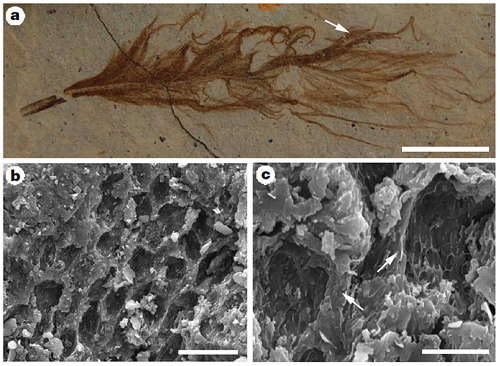
Так можно ли считать обнаружение прото-перьев доказанным? Все же излишняя категоричность ведет к догматизму. Правильнее было бы сказать, что накопленные на сегодняшний день данные говорят, что они обнаружены.
Приложение 2. Эволюция дыхательной системы.
Дыхательная система птиц значительно видоизменена. Легкие птиц прилегают к спинной стенке грудной клетки. Они строго ограничены в этом положении ребрами. При вдохе и выдохе легкие птиц почти не изменяются в объеме. Не смотря на это, они успешно функционируют, что становится возможным благодаря системе воздушных мешков – полых органов, расположенных между внутренними органами. Типично их девять: один межключичный, два шейных, два передних грудных, два задних грудных и два брюшных. В довершение всего на момент вдоха легкие заполнены воздухом. Работает такая система следующим образом. Движения грудины, ребер и плечевого пояса приводит к изменению давления, которое создает поток воздуха.
При вдохе воздух, содержащийся в легких, попадает в передние воздушные мешки, а воздух из внешней среды проходит в задние воздушные мешки. При выдохе насыщенный углекислым газом воздух из передних мешков высвобождается в окружающую среду, а воздух из задних мешков проходит в легкие и через передние воздушные мешки также попадает во внешнюю среду.
|
Преимущество описанной системы в том, что обогащенный кислородом воздух оказывается в легких и при вдохе, и при выдохе. В результате легкие птиц работают на 160% эффективнее, чем легкие млекопитающих. Если бы наши легкие вдруг стали столь же функциональными, как легкие птиц, мы стали бы получать кислорода в 2,5 раз больше. Правда, нам бы это совсем не понравилось.
Мы не можем ожидать, что легкие и воздушные мешки сохраняться в окаменелостях. Означает ли это, что мы никогда ничего не сможем узнать об эволюции дыхательной системы птиц? Не обязательно. Работа птичьей дыхательной системы в частности связано со скелетом, а значит, мы можем попытаться отыскать соответствующие признаки в окаменелостях. Воздушные мешки заходят в полости костей. Кроме того, функционирование птичьей дыхательной системы обеспечивается характерным строением грудных ребер и соответствующих им позвонков. Остается только описать эти признаки.
Шейные воздушные мешки заходят в шейные и частично в грудные позвонки.
Межключичный воздушный мешок связан с пустотами в грудине, плечевом поясе и локтевых костях.
Легкие также связаны с пустотами – в грудных позвонках и в смежных с ними ребрах.
Передние грудные мешки связаны с грудинными ребрами.
Задние грудные, крестцовые и хвостовые позвонки, а также бедро и элементы тазового пояса включают ответвления брюшных воздушных мешков.
К сожалению, диагностика большинства перечисленных черт требует хорошего сохранения костей. Тем не менее, многие из упомянутых особенностей на сегодняшний день описаны у звероногих динозавров, включая близких родственников птиц.
Данные о пустотах в костях свидетельствуют о присутствии у звероногих, по крайней мере, двух пар воздушных мешочков: затылочных и брюшных, а также о присутствии межключичного.
Строение практически не изменяющихся в объеме легких птиц и их физиология обеспечиваются анатомией грудной клетки, ребер и позвонков. Легкие прикреплены к позвоночной колонне, а в их спинную поверхность врезаются головки ребер, создавая углубления в легочной ткани. Грудные ребра птиц морфологически однородны по всему ряду. Они соединяются с позвонками двумя хорошо развитыми концами, отделенными друг от друга явной выемкой. Такое строение ребер не позволяет грудной клетке растягиваться в спинном и брюшном направлении. Боковые отростки позвонков соединяются с головками ребер в брюшном положении, в результате чего потолок грудной полости становится сильно бороздчатым, что также препятствует легким расширяться.
У звероногих динозавров грудные ребра имеют аналогичное строение. Они соединяются с позвонками двумя концами, которые разделены, но не столь значительно, как у птиц, а боковые отростки позвонков имеют сходное положение. Эти особенности строения звероногих свидетельствуют, что их легкие были в значительной мере подобны легким птиц, а именно строение и функционирование грудной клетки сильно ограничило бы изменение легких в объеме.
Учитывая все сказанное, можно в общих чертах описать дыхательную систему предковой группы птиц. Известные анатомические особенности показывают, что у звероногих динозавров дыхание осуществлялось за счет функционирования воздушных мешков, через которые воздух проходил в легкие, которые только в незначительной степени могли изменяться в объеме.
Хотя у звероногих присутствует ряд скелетных особенностей, которые являются типичными компонентами птичьей дыхательной системы, все же полный набор признаков не выявлен. Недавно появились данные о том, что строение задних конечностей звероногих не позволило бы брюшным мешкам функционировать по птичьему образцу. Поэтому дыхательная система звероногих может быть описана как промежуточная в анатомии и физиологии. Интересно, что функционирование брюшных воздушных мешков, вероятно, отличалось от того образца, который известен у современных птиц. Не следует забывать, что звероногие обладали также брюшными ребрами, которых у птиц нет. Есть некоторые свидетельства того, что брюшные ребра, возможно, были вовлечены в физиологию дыхания.
Недавно несколько биологов предложили модель четырех стадий эволюции дыхательной системы птиц. Более подробно см. [75].
Приложение 3. Сравнение анатомии дейнонихозавров, ранних и современных птиц.
Таблица анатомических характеристик дейнонихозавров и птиц, на основе которой построена диаграмма процентного соотношения птичьих и рептилийных особенностей у наиболее интересных переходных форм. Символ "?" обозначает, что сохранность окаменелостей не позволяет выявить анатомическую характеристику. Символом "*" отмечены случаи, когда особенность строения в палеонтологической литературе не описана, и не ясно, позволяют ли окаменелости ее диагностику. Промежуточные состояния анатомических особенностей обозначены с помощью символа "/". Таблица доступна на отдельной странице.
a. Я неоднократно подчеркиваю, что сообщаемые мной сведения о сравнении строения вымерших и современных животных упрощены. Все же следует разъяснить, что упрощения даже больше, чем оговорено в основном тексте. Во-первых, все черты строения рассматриваются как имеющие одинаковое значение. Во-вторых, я не выстраиваю анатомические черты в иерархическую схему. Вот в чем конкретно заключается каждое из этих упрощений.
Не все черты строения одинаково уместны для прослеживания эволюции. Поэтому необходимо отличать те, которые полезны для исследования эволюции, от тех, которые бесполезны. Прежде всего, следует выделить те черты, которые позволяют диагностировать принадлежность организма к своему виду, роду и т. д. Например, для археоптерикса одной из таких черт является редуцированная пятая кость плюсны. Такие особенности становятся полезными в прослеживании эволюционных отношений, если они наследуются потомковыми линиями. Другая группа черт - особенности, которые не наследуются от общего предка, а возникают независимо в разных группах вследствие параллельной (конвергентной) эволюции. Такие особенности часто усложняют прослеживание происхождения изучаемых групп, особенно в тех случаях, когда доступный для исследования материал фрагментарен. Так например, ранее по некоторым признакам птиц связывали с крокодилами и птицетазовыми динозаврами. Однако в дальнейшем было установлено, что уместные особенности независимо развились в этих группах. Третья группа особенностей - аннулирования, которые также могут затемнять эволюционную историю. Нередко разные группы теряют типичные и унаследованные от последнего общего предка черты. Так змеи, безногие ящерицы и безногие амфибии относятся к четвероногим животным, даже не смотря на то, что они утратили конечности (независимо, в результате конвергентной эволюции). Поэтому для анализа эволюционных отношений наибольший интерес представляют наиболее ранние формы, которые в наименьшей степени отклонились от предкового состояния и поэтому с большей вероятностью сохраняют наследственные черты. Наконец, выделяют такие черты, которые наследуются потомковой линией от предка. Именно они позволяют прослеживать эволюционные связи. У птиц к таковым относятся, например, перья.
Я говорю обо всех анатомических чертах как о "рептилийных" или "птичьих". Более точная картина возникает, если использовать иерархическую классификацию черт. Есть особенности, которые птицы разделяют со всеми архозаврами (предглазничное окно), с целурозаврами (полулунная кость запястья), с дейнонихозаврами (форма коракоида) и т. д. Такая иерархия дает возможность максимально точно проследить родственные связи.
b. В этой работе я стараюсь избегать специфически научной терминологии. Например, моя задача существенно упростилась, если бы я использовал такой полезный и удобный термин как "таксон". Отсутствие удобной терминологии делает текст несколько громоздким и невнятным. Однако такой подход позволяет обойтись без объемных разделов, служащих только введением в сложные научные вопросы. Включение таких разделов привело бы к значительной затрате места, времени и, вероятно, потере многих читателей.
c. Еще одна чрезвычайно интересная тема для популяризации знаний об эволюции заслуживает отдельного упоминания. В некоторых случаях качество ископаемого отчета позволяет проследить начальную эволюцию крупных групп позвоночных животных. Это дало возможность палеонтологам подтвердить, что на начальных этапах обособления эволюционной ветви ее родоначальники в строении стоят ближе к своим предкам и ближайшим родственникам, чем к более поздним потомкам. Например, в каменноугольном периоде возникли три большие ветви четвероногих животных: анапсиды (ныне вымершие), диапсиды (включают ящериц, змей, крокодилов, птиц, а также динозавров, плезиозавров, птерозавров и множество других вымерших) и синапсид (млекопитающие плюс их вымершие родственники и пеликозавры, также вымершие). Как можно легко понять, каждая ветвь характеризуется чрезвычайным разнообразием форм. Однако первые представители этих ветвей в анатомии весьма и весьма подобны друг другу. Многообразие форм (и прошлое, и наблюдаемое сегодня) возникает не сразу. То же верно, например, для отряда непарнокопытных млекопитающих. Современных непарнокопытных (лошади, носороги и тапиры) невозможно перепутать. Однако ранние представители соответствующих семейств отличаются друг от друга лишь несколькими мелкими особенностями скелета. В случае птиц ископаемый отчет не столь хорош, но картина во многом такая же. Ранние птицы настолько близки в анатомии к нескольким родственным семействам звероногих динозавров, что палеонтологам пришлось немало потрудиться, чтобы найти анатомические черты, разделяющие птиц и их ближайших родственников.
Такая информация позволяет показать эволюционные переходы на крупном масштабе и обеспечивает дополнительные свидетельства для теории единой общей родословной. К сожалению, мне не встречались научно-популярные обзоры по данной тематике. Традиционно переходы и переходные формы описываются на уровне упоминания отдельных родов. В своей статье я также следовал за этой традицией.
Промежуточность не только отдельных родов, но крупных групп также красноречиво свидетельствует против идей креационистов о характере ископаемого отчета. Креационисты настаивали и настаивают, что крупные группы появляются в летописи окаменелостей внезапно. Можно показать, что это ложно, но обсуждаемые данные порождают даже больше проблем для креационистов. Если в момент появления в отчете оказываются промежуточными сами группы, то вопрос внезапности теряет значение. Кроме того, важно отметить, что крупные группы появляются не в полном составе. В качестве примера можно рассмотреть уже упоминавшихся диапсид. Ящерицы, змеи, крокодилы, птицы, динозавры, плезиозавры, птерозавры и др. появляются намного позже ранних диапсид. Особо интересны змеи, которые возникли в меловом периоде. Таким образом, от ранних диапсид их отделяет несколько геологических периодов: пермский, триасовый и юрский. В свете таких данных предполагаемая внезапность немыслима.
Выражаю глубокую признательность Георгию Рюрикову (Институт проблем экологии и эволюции (ИПЭЭ РАН)) за рецензию первоначального варианта этой статьи, ценные замечания и предложения. Ответственность за любые возможные ошибки, неточности и огрехи ложится только на меня.
- Alonso P. et al. The avian nature of the brain and inner ear of Archaeopteryx. - Nature, 2004, v. 430, p. 666-669.
- Baier D. et al. A critical ligamentous mechanism in the evolution of avian flight. - Nature, 2007, v. 445, p. 307-310
- Balanoff A. et al. Cranial osteology of the theropod dinosaur Incisivosaurus gauthieri (Theropoda: Oviraptorosauria). - American Museum Novitates, 2009, n. 3651, 35pp.
- Bennett S. Ontogeny and Archaeopteryx. - Journal of Vertebrate Paleontology, 2008, v. 28, p. 535-542.
- Buffetaut E et al. Minute theropod eggs and embryo from the Lower Cretaceous of Thailand and the dinosaur-bird transition. - Naturwissenschaften, 2005, v. 92, p. 477-482.
- Butler R., Goswami A. Body size evolution in Mesozoic birds: little evidence for Cope's rule. - J Evol Biol, 2008, v. 21, p. 1673-1682
- Carney et al. New evidence on the colour and nature of the isolated Archaeopteryx feather. - Nature Communications, 2012, v. 3 doi: 10.1038/ncomms1642
- Chiappe L. Downsized Dinosaurs: The Evolutionary Transition to Modern Birds. - Evo Edu Outreach, 2009, v. 2 p. 248-256
- Chiappe L. et al. Anatomy and systematics of the Confuciusornithidae (Theropoda: Aves) from the Late Mesozoic of northeastern China. - Bulletin of the American Musuem of Natural History, 1999, v. 242, p. 1-88
- Chiappe L. The closest relatives of birds. - Ornitologia Neotropical, 2004, v. 15, p. 1-16
- Chiappe L., Dyke G. The early evolutionary history of birds. - Journal of the Paleontological Society of Korea, 2006, v. 22, p. 133-151.
- Chiappe L., Dyke G. The Mesozoic radiation of birds. - Annual Review of Ecology and Systematics, 2002, v. 33, p. 91-124.
- Chiappe L., Witmer, L. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. - University of California Press, Berkeley, California, 2002
- Choiniere J. et al. A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China. - Science, 2010, v. 327, p. 571-574
- Christiansen P., Bonde N. Axial and appendicular pneumaticity in Archaeopteryx. - Proceedings of the royal Society, Biological Sciences, 2000, v. 267, p. 2501-2505
- Christiansen P., Bonde N. Body plumage in Archaeopteryx: a review, and new evidence from the Berlin specimen. - Comptu Palevol, 2004, v. 3, p. 99-118
- Chuong C. et al. Adaptation to the sky: defining the feather with integument fossils from Mesozoic China and experimental evidence from molecular laboratories. - Special issue: Development and evolution of amniote integuments, guest editors, Chuong, C.-M., and Homberger, D. G., Journal of Experimental Zoology, Molecular and Developmental Evolution, 2003, v. 298B, p. 42-56
- Claessens L. Archosaurian respiration and the pelvic girdle aspiration breathing of crocodyliforms. - Proceedings of the Royal Society of London B: Biological Sciences, 2004,v. 271, p. 1461-1465
- Claessens L. Dinosaur gastralia; origin, morphology, and function. - Journal of Vertebrae Paleontology, 2004, v. 24, p. 89-106.
- Clark J. An Oviraptorid Skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, Preserved in an Avianlike Brooding Position Over an Oviraptorid Nest. - American Museum Novitates, 1999, n. 3265, p. 1-36.
- Codd J. et al. Avian-like breathing mechanics in maniraptoran dinosaurs. - Proceedings of the Royal Society of London, series B, 2008, v. 275, p. 157-161
- Dyke G., Kaiser G. (eds.). Living Dinosaurs: The Evolutionary History of Modern Birds. - John Wiley & Sons, London, 2011.
- Dyke G., Norell M. Caudipteryx as a non-avialan theropod rather than a flightless bird. - Acta Palaeontologica Polonica, 2005, v. 50, p. 101-16
- Eldredge N., Gould S. Punctuated equilibria: an alternative to phyletic gradualism. In Schopf T. (ed.) Models in Paleobiology. - Freeman, Cooper and Co., San Francisco., 1972, pp. 82-115.
- Elzanowski A. A new genus and species for the largest specimen of Archaeopteryx. - Acta Palaeontologica Polonica, 2001, v. 46, p. 519-532
- Foley J. Dmanisi and Answers in Genesis. http://pandasthumb.org/archives/2008/12/dmanisi-and-aig.html
- Foley J. Duane Gish quote about ER 1470. http://talkorigins.org/faqs/homs/cronin.html
- Foley J. Skull D2700. http://talkorigins.org/faqs/homs/d2700.html
- Foley J. The Dmanisi skulls and creationism. http://pandasthumb.org/archives/2005/05/the-dmanisi-sku.html
- Fountaine T. et al. The quality of the fossil record of Mesozoic birds. - Proc. R. Soc. B, 2005, v. 272, p. 289-294
- Franzosa J. Evolution of the Brain in Theropoda (Dinosauria). - Dissertation, 2004.
- Gish D. As a Transitional Form Archaeopteryx Won't Fly. - Acts & Facts, 1989, v. 18 (9). http://www.icr.org/article/321/
- Gould S. Evolution as Fact and Theory. in Hens Teeth and Horse's Toes: Further Reflections in Natural History. - New York: W. W. Norton & Co., 1983, p. 258-260.
- Greenwold M., Sawyer R. Linking the molecular evolution of avian beta (b) keratins to the evolution of feathers. - Journal of Experimental Zoology (Molecular and Developmental Evolution), 2011, v. 316, p. 609-616.
- Grellet-Tinner G. et al, Dinosaur eggs and nesting behaviors: a paleobiological investigation. - Palaeogeography, Palaeoclimatology, Palaeoecology, 2006, v. 232, p. 294-321.
- Hollida, C., Witmer L. Cranial Kinesis in Dinosaurs: Intracranial Joints, Protractor Muscles, and Their Significance for Cranial Evolution and Function in Diapsids. - Journal of Vertebrate Paleontology, 2008, v. 28, p. 1073-1088.
- Holtz T. A new phylogeny of the carnivorous dinosaurs. - Gaia, 1998. v. 15, p. 5-61
- Horner J. Dinosaur reproduction and parenting. -Annual Review of Earth and Planetary Sciences, 2000, v. 28, p. 19-45.
- Hu D. et al. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. - Nature, 2009, v. 461, p 640-643
- Hwang S. et al. New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from northeastern China. - American Museum Novitates, 2002, n. 3381, p. 1-44.
- Ji Q. et al. Two feathered dinosaurs from northeastern China. - Nature, 1998, v. 393, p. 753-761.
- Kavanau J. Roots of avian evolution: clues from relict reproductive behaviors. -Scientific Research and Essay, v. 2(8), p. 263-294.
- Kurochkin E. et al. A fossil brain from the Cretaceous of European Russia and avian sensory evolution. - Biology Letters, 2007, v. 3, p. 309-313
- Larson P. The theropod reproductive system: In: Aspects of Theropod Paleobiology, edited by Perez-Moreno, B. P., Holtz, T. Jr., Sanz, J. L., and Moratalla, J., Gaia, 2000, v. 15, p. 389-387.
- Larsson H. et al. Forebrain enlargement among nonavian theropod dinosaurs. - Journal of Vertebrate Paleontology, 2000, v. 20, p. 615-618.
- Lee M., Worthy T. Likelihood reinstates Archaeopteryx as a primitive bird. - Biology Letters, 2012, v. 23 p. 299-303.
- Li D. et al. Basal birds from China: a brief review. - Chinese Birds, 2010, v. 1, p. 83-96
- Li Q. et al. Reconstruction of Microraptor and the evolution of Iridescent Plumage. - Science, 2012, v. 335, p. 1215-1219.
- Martin L. et al. Confuciusornis sanctus compared to Archaeopteryx lithographica. - Naturwissenschaften, 1998, v. 85, p. 286-289
- Mayr G. et al. A well-preserved Archaeopteryx specimen with theropod features. - Science, 2005, v. 310, p. 1483-1486
- Mayr G. et al. The tenth skeletal specimen of Archaeopteryx. - Zoological Journal of the Linnean Society, 2007, v. 149, p. 97-116.
- Middleton K. The morphological basis of hallucal orientation in extant birds. - Journal of Morphology, 2001, v. 250, p. 51-60.
- Miller K. Taxonomy, transitional forms, and the fossil record. http://www.asa3.org/ASA/resources/Miller.html
- Nesbitt S. et al. The theropod furcula. - Journal of Morphology, 2009, v. 270, p. 856-879.
- Norell M. et al. A Velociraptor wishbone. - Nature, 1997, v. 389, p. 447
- Norell M., Xu X. Feathered Dinosaurs. - Annual Review of Earth and Planetary Sciences, 2005, v. 33, p. 277-299.
- O'Connor P., Claessens L. Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. - Nature, 2005, v. 436, p. 253-256
- O'Connor J. et al. A new species of Jeholornis with complete caudal integument. - Historical Biology, 2012, v. 24(1), p. 29-41
- Ornithologist and Evolutionary Biologist Alan Feduccia - Plucking Apart the Dino-Birds. - Discover magazine, 2003, V. 24, № 2. http://discovermagazine.com/2003/feb/breakdialogue
- Osmolska H. Evidence on relation of brain to endocranial cavity in oviraptorid dinosaurs. - Acta Palaeontologica Polonica, 2004, v. 49, p. 321-324.
- Ostrom J. Archaeopteryx and the origin of birds. - Biological Journal of the Linnean Society, 1976, v. 8, p. 91-182
- Padian K. et al. Dinosaurian growth rates and bird origins. - Nature, 2001, v. 412-408.
- Padian K., Chiappe L. The origin and early evolution of birds. - Biological Review, 1998, v. 73, p. 1-42.
- Prothero D. Bringing Fossils to Life: An Introduction to Paleobiology. (2nd ed.). - WCB/McGraw-Hill, New York: 2003.
- Prothero D. Evolution: What the Fossils Say and Why it Matters. - Columbia University Press, New York: 2007
- Prothero D., Schoch R. Horns, Tusks, and Flippers: The Evolution of Hoofed Mammals. - Johns Hopkins University Press, Baltimore: 2002.
- Provini P. et al. A new species of the basal bird Sapeornis from the Early Cretaceous of Liaoning, China. - Vertebrata PalAsiatica, 2009, v. 47, p. 194-207
- Prum R. S06-3 The theropod origin of feathers: congruence between development, paleontology, and phylogeny. - Acta Zoologica Sinica, 2006, v. 52, p. 129-130.
- Prum R., Brush A. The evolutionary origin and diversification of feathers. - The Quarterly Review of Biology, 2002, v. 77, p. 261-295.
- Prum R., Dyck J. A Hierarchical Model of Plumage: Morphology, Development, and Evolution. - Journal of Experimental Zoology (Molecular and developmental evolution), 2003, v. 298B, p. 73-90
- Quanguo L. et al. Plumage Color Patterns of an Extinct Dinosaur. - Science, 2010, v. 327, p. 1369-1372
- Sanz J. et al. A Nestling Bird from the Lower Cretaceous of Spain: Implications for Avian Skull and Neck Evolution. - Science, 1997, v. 276, p. 1543-1546
- Schachner E. et al. Evolution of the dinosauriform respiratory apparatus: new evidence from the postcranial axial skeleton. - The Anatomical Record, 2011, v. 294, p. 1532-1547.
- Schachner E. et al. Evolution of the Respiratory System in Nonavian Theropods: Evidence from Rib and Vertebral Morphology. - The anatomical record, 2009, v. 292, p. 1501-1513
- Sereno P. et al. Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina. - PLoS ONE, 2008, v. 3, p. e3303
- Sereno P. The Evolution of Dinosaurs, - Science, 1999, v. 284, p. 2137-2147.
- Sullivan C. et al. The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs. - Proceedings of the Royal Society, Series B., 2010, v. 277, p. 2027-2033
- Sumida S., Brochu C. Phylogenetic context for the origin of feathers: In: Evolutionary Origin of Feathers, organized by Maderson, P. F. A. and Homberger, D. A., American Zoologist, 2000, v. 40, p. 486-503.
- Switek B. The Branching Bush of Horse Evolution. http://laelaps.wordpress.com/2007/09/13/the-branching-bush-of-horse-evolution/
- Turner A. et al. A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight. - Science, 2007, v. 317, p. 1378-1381
- Turner A. Feather quill knobs in the Dinosaur Velociraptor. - Science, 2007, v. 317, p. 1721.
- Vinther J. et al. Structural coloration in a fossil feather. - Biology Letters, 2010, v. 6, 128-131
- Vinther J. et al. The colour of fossil feathers. - Biology Letters, 2008, v. 4, p. 522-525.
- Wedel M. Evidence for bird-like air sacs in saurischian dinosaurs. - Journal of Experimental Zoology, 2009, v. 311A p. 611-628.
- Wedel M. Postcranial pneumaticity in dinosaurs and the origin of the avian lung, Ph.D. dissertation, 2007, University of California, Berkeley, p. 112.
- Weishampel D., Dodson P, Osmolska H. (eds.) The dinosauria. University of California Press, Berkeley and Los Angeles, California, 2004.
- Wise K. Towards a Creationist Understanding of Transitional Forms. - Creation Ex Nihilo Technical Journal, 1995, v. 9, p. 216-222.
- Witmer L. New aspects of avian origins: roundtable report. In: Avian Paleontology at the Close of the 20th Century: Proceedings of the 4th International Meeting of the Society of Avian Paleontology and Evolution, Washington, D.C., 4-7 June 1996, editor Olson, S. L., associate editors, Wellnhofer P., Mourer-Chauvire C., Steadman D. W., and Martin L. D., Smithsonian Contributions to Paleobiology, 1999, n. 89, p. 327-334.
- Xu X. et al. A basal troodontid from the Early Cretaceous of China. - Nature, 2002, v. 415, p. 780-784
- Xu X. et al. A basal tyrannosauroid dinosaur from the Late Jurassic of China. - Nature, 2006, v. 439, p. 715-718
- Xu X. et al. A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. - Nature, 1999, v. 401, p. 262-266.
- Xu X. et al. A juvenile coelurosaurian theropod from China indicates arboreal habits, - Naturwissenschaften, 2002, v. 89, p. 394-398
- Xu X. et al. A new feather type in a nonavian theropod and the early evolution of feathers. - Proceedings of the National Academy of Science, 2009, v. 106, p. 832-834
- Xu X. et al. A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin. - Chinese Science Bulletin, 2008, v. 54, p. 430-435
- Xu X. et al. An Archaeopteryx-like theropod from China and the origin of Avialae. - Nature, 2011, v. 475, p. 465-470.
- Xu X. et al. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids: Nature, 2004, v. 431, p. 680-684
- Xu X. et al. Branched integumental structures in Sinornithosaurus and the origin of feathers. - Nature, 2001, v. 410, p. 200-204.
- Xu X. et al. Four-winged dinosaurs from China, - Nature, 2003, v. 421, p. 335-340.
- Xu X. et al. The smallest known non-avian theropod dinosaur. - Nature, 2000, v. 408, p. 705-708.
- Xu X. Feathered dinosaurs from China and the evolution of major avian characters. - Integrative Zoology, 2006, v. 1, p. 4-11.
- Xu X., Norell M. A new troodontid dinosaur from China with avian-like sleeping posture. - Nature, 2004, v. 431, p. 838-841
- Yu M. et al. The morphogenesis of feathers. - Nature, 2002, v. 420, p. 308-312.
- Zelenisky D. et al. Bird-like characteristics of troodontid theropod eggshell. - Cretaceous Research, 2002. v. 23, p. 297-305
- Zelenitskiy D. Reproductive traits of non-avian theropods. - Journal of the Paleontological Society of Korea, 2006, v. 22, p. 15-27.
- Zhang F. et al. A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers. - Nature, 2008, v. 455, p. 1105-1108
- Zhang F. et al. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. - Nature, 2010, v. 463, p. 1075-1078
- Zhang Z. et al. Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China. - Journal of Ornithology, 2009, v. 150, p. 783-790
- Zhang, F. et al. A primitive confuciusornithid bird from China and its implications of rearly avian flight. - Science in China, Series D, Earth Sciences, 2008, v. 51, p. 625-639.
- Zhou Z. et al. A new basal ornithurine bird (Jianchangornis microdonta gen. et sp. nov.), from the Lower Cretaceous of China. - Vertebrata PalAsiatica, 2009, v. 47, p. 399-310
- Zhou Z. et al. Gastroliths in Yanornis: an indication of the earliest radical diet-switching and gizzard plasticity in the lineage leading to living birds? Naturwissenschaften, - 2004, v. 91, p. 571-574.
- Zhou Z. The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence. - Naturwissenschaften, 2004, v. 91, p. 455-471
- Zhou Z., Li F. A new Lower Cretaceous bird from China and tooth reduction in early avian evolution. - Proceedings of the Royal Society, Series B, 2009, v. 277, p. 219-227
- Zhou Z., Zhang F. A long-tailed, seed-eating bird from the Early Cretaceous of China. - Nature, 2002, v. 418, p. 405-409
- Zhou Z., Zhang F. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. - Canadian Journal of Earth Sciences, 2003, v. 40, p. 731-747
- Zhou Z., Zhang F. Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. - Naturwissenschaften, 2003, v. 90, p. 220-225.
- Zhou Z., Zhang F. Mesozoic birds of China - A synoptic Review, - Vertebrata PalAsiatica, 2006, v. 44, p. 74-98.
- Zhou Z., Zhang F. Mesozoic birds of China: an introduction and review. - Acta Zoologica Sinica, 2004, v. 50, p. 913-920.
- Zhou Z., Zhang F. Origin and early evolution of feathers: evidence from the Early Cretaceous of China. - Acta Zoologica Sinica, 2006. v. 52, p. 125-128.
- Zhou Z., Zhang F. Two new ornithurine birds from the Early Cretaceous of western Liaoning, China. - Chinese Science Bulletin, 2001, v. 46, p. 1-7.
- Алферов. Православное мировоззрение и современное естествознание (Уроки креационной науки в старших классах средней школы). - М.: Паломник, 1998.
- Бейкер С. Камень преткновения: Верна ли теория эволюции? - М.: "Протестант", 1992. http://www.scienceandapologetics.com/pdf/sylvia_beker_-_kamen_pretknovenija_ili_prava_li_evolucija.pdf
- Вертьянов С. Ю. Общая биология: Учебник для 10-11 кл. общеобразовательных учреждений. - М.: Свято-Троицкая Сергиева Лавра, 2012 http://www.vertyanov.ru/books/vertyanov_uchebnik_po_biologii_10_11.pdf
- Гиш Д. Сотворение мира, эволюция и исторические свидетельства. - The American Biology Teacher, 1973(8?), p.132-140. http://www.evangelie.ru/content.php?r=364-%C4%F3%FD%E9%ED-%D2.-%C3%E8%F8.-%D1%CE%D2%C2%CE%D0%C5%CD%C8%C5-%CC%C8%D0%C0-%DD%C2%CE%CB%DE%D6%C8%DF-%C8-%C8%D1%D2%CE%D0%C8%D7%C5%D1%CA%C8%C5-%D1%C2%C8%C4%C5%D2%C5%CB%DC%D1%D2%C2%C0
- Гладсхауэр У. Как возник наш мир. - CLV, 2004. http://www.volodarmira.narod.ru/lib/00141.htm
- Роузвер Д. Наука о сотворении мира, доказывающая правоту Библии. - Симферополь: Крымское общество креационной науки, 1995. http://www.creationism.org/crimea/pdf/creatscience.pdf
- Жизнь - как она возникла? Путем эволюции или путем сотворения? - Watchtower bible and tract society of New York, 1992
- Курочкин Е. Н. Новые идеи о происхождении и ранней эволюции птиц - Достижения и проблемы орнитологии Северной Евразии на рубеже веков / Под ред. Курочкина Е. Н., Рахимова И. И. Казань: Изд - во МАГАРИФ, 2001. С. 68-96.
- Кэрролл Р. Палеонтология и эволюция позвоночных. - т. 2. - М., 1993.
- Маклин Дж., Окленд Р., Маклин Л. Очевидность сотворения мира. Происхождение планеты Земля. - Триада, 1991. http://jesuschrist2012.narod.ru/books/Conspicuity.htm
- Моррис Г. Библейские основы современной науки. http://www.librius.net/b/6210/read
- Моррис Г. Сотворение и современный христианин. - М.: "Протестант", 1993. http://www.volodarmira.narod.ru/lib/00206.htm
- Остин C. Археораптор. "Пернатый динозавр" Национального географического общества не умеет летать. - Institute for Creation Research, 2000, n. 321. http://anwarrior.narod.ru/creation/arheoraptor.html
- Пучков П.В., Дзеверин И. И. Геологическая летопись и радикальный "научный" креационизм. http://evolution.powernet.ru/polemics/cr31.html
- Рассел Б. Мистицизм и логика. - Почему я не христианин. - М.: Политиздат, 1987. http://philosophy.ru/library/russell/02/01.html
- Рос А. В начале... - Источник жизни, 2002. http://www.creationism.org/crimea/zip/ariel_ros-v_nachale.zip
- Сарфати Дж. Несостоятельность теории эволюции. - Симферополь: Христианский научно-апологетический центр, 2001. http://www.volodarmira.narod.ru/lib/16.html
- Сысоев Д. Летопись начала. Библейская история и современная наука. - Издательство Сретенского монастыря, 1999. http://shestodnev.narod.ru/archieve/dsysoev_letopis.pdf
- Тейлор П. Сотворение. Иллюстрированная книга ответов. - СПб.: Библия для всех, 1994. http://www.creationism.org/crimea/pdf/paul_teylor.pdf
- Томас Хайнц. Творение или эволюция: анализ теории эволюции в свете Священного Писания. http://www.volodarmira.narod.ru/lib/00143.htm
- Хэм К., Сарфати Дж., Виланд К. под ред. Бэттена Д. Книга Ответов: расширенная и обновленная. - Симферополь: Христианский научно-апологетический центр, 2000. http://volodarmira.narod.ru/lib/13.html
- Юнкер Р., Шерер З. и др. История происхождения и развития жизни. Основные положения и понятия для уроков биологии. - СПб.: КАЙРОС, 1997. http://www.creationism.org/crimea/tutorial/index.html
- Прам Р., Браш А. Динозавры или птицы: кто оперился первым? - В мире науки, 2003. №7 http://evolution.powernet.ru/library/feather_origin.html