ГЛАВА 15. МОЛЕКУЛЯРНЫЙ КИСЛОРОД КАК ФАКТОР ЭВОЛЮЦИИ
Общепринято представление о том, что молекулярный кислород атмосферы имеет биогенное происхождение, и его появление непосредственно связано с формированием нового типа фотосинтеза, при котором в качестве донора электронов используется вода. В условиях первобытной Земли до возникновения выделяющих кислород фотосинтезирующих эубактерии единственным источником свободного кислорода была реакция фотолиза паров воды в атмосфере, происходящая под действием коротковолнового ультрафиолета. Однако количество "фотолитического" кислорода было ничтожным. Образующийся кислород использовался для окисления газов первобытной атмосферы и восстановленных минералов, входящих в состав земной коры.
Из всех организмов, осуществляющих фотосинтез с выделением O2, наиболее примитивно организованными являются фотосинтезирующие эубактерии (цианобактерий, прохлорофиты), и мы вправе предполагать, что появление молекулярного кислорода связано с этими организмами или с какими-то их весьма близкими предками.
До возникновения фотосинтезирующих эукариот, и в первую очередь высших растений, содержание свободного кислорода в атмосфере Земли было незначительным по сравнению с его содержанием в современной земной атмосфере. Однако, по проведенным подсчетам, для переключения организма с брожения на дыхание достаточна концентрация кислорода 0,2%, т. е. 0,01 его уровня в современной атмосфере. Появление и накопление O2 в земной атмосфере было событием, значение которого для последующей эволюции жизни на Земле трудно переоценить. Прежде всего это означало существенную перестройку всего, что сформировалось на Земле в "докислородную" эпоху, и в первую очередь касалось живых организмов.
Образование O2 в возрастающих количествах сделало возможным протекание окислительных реакций в широких масштабах. Изменился характер атмосферы: из восстановительной она стала окислительной. Последнее повлекло за собой существенные изменения в отношении донор-акцепторной проблемы. Если в условиях бескислородной атмосферы доминирующим было решение проблемы акцептора электронов, то в условиях кислородной атмосферы основной становится проблема донора электронов, поскольку с появлением O2 в атмосфере Земли образовался источник превосходного акцептора электронов.
ВЗАИМОДЕЙСТВИЕ ПРОКАРИОТ С МОЛЕКУЛЯРНЫМ КИСЛОРОДОМ
Первоначально молекулярный кислород появился внутри клетки, и это сразу же создало проблему взаимодействия клетки с O2. Очевидно, что у первых фотосинтезирующих организмов, продуцировавших молекулярный кислород, не было ферментных систем не только для выгодного использования этого акцептора, но и для его нейтрализации в клетке. Не было их также и у других существовавших анаэробных форм жизни. Поэтому можно предполагать, что первый тип взаимодействия с O2 базировался на резко отрицательном отношении к нему клетки. Пример этого — многочисленные данные по высокой токсичности молекулярного кислорода для современных облигатно анаэробных организмов62.
62 В этой связи интересны данные о том, что в период, предшествовавший появлению больших количеств свободного кислорода в атмосфере, прокариотное сообщество было разнообразнее, чем в последующее время. Разнообразие прокариотного сообщества значительно уменьшилось 1,5 млрд лет назад (см. рис. 52).
По мере накопления O2 становится постоянным компонентом внешней среды, и только локально могут быть созданы такие условия, где он отсутствует или содержится в следовых количествах. Это обусловило два возможных варианта последующего взаимодействия прокариот с молекулярным кислородом. Одни из существовавших анаэробных форм "ушли" в места обитания, где O2 практически отсутствует, и тем самым сохранили "облик бескислородной эпохи". Другие были вынуждены пойти по пути приспособления к "кислородным" условиям. Это означает, что они формировали новые метаболические реакции, служащие в первую очередь для нейтрализации отрицательного действия молекулярного кислорода.
Итак, следующий шаг на пути взаимодействия прокариот с кислородом — возможность существовать в присутствии O2, нейтрализуя его отрицательное действие. Определенное представление о формировавшихся системах защиты от молекулярного кислорода у прокариот можно получить, изучая представителей этой группы, располагающихся на разных ступенях эволюционной лестницы.
Токсические эффекты молекулярного кислорода и его производных
Как фактор внешней среды O2 воздействует на современные прокариотные организмы двояко: с одной стороны, он может быть абсолютно необходимым, с другой — с молекулярным кислородом и его производными связаны токсические эффекты для клеток.
Молекулярный кислород. Существует ряд гипотез, объясняющих чувствительность прокариот к O2. Согласно одной из них молекулярный кислород сам является токсическим соединением, агрессивное действие которого связано со способностью окислять клеточные метаболиты, необходимые для функционирования в восстановленном состоянии. Токсический эффект O2 зависит от условий, при которых происходит взаимодействие с ним организмов: концентрации растворенного O2, длительности экспозиции, состава окружающей среды.
Токсичность молекулярного кислорода, например, может быть следствием активного акцептирования им электронов с растворимых переносчиков, функционирующих в процессах брожения, что будет приводить к истощению внутриклеточного пула восстановленных доноров электронов, необходимых для биосинтезов. Действительно, было обнаружено, что активность растворимых флавопротеинов, способных функционировать как НАД(Ф)-H2-оксидазы, повышалась в 5 — 6 раз при выращивании Clostridium acetobutylicum в аэробных условиях сравнительно с анаэробными. Сдвиг под влиянием O2 электронных переносчиков в сторону их преимущественного нахождения в окисленном состоянии приводил к подавлению роста и изменению выхода продуктов брожения: прекращению синтеза масляной кислоты и накоплению более окисленного продукта — уксусной кислоты.
Наконец, для проявления токсического эффекта O2 вполне достаточно окисления им какого-либо одного ключевого метаболита или фермента, приводящего к их инактивации. Известны три ферментные системы прокариот, особо чувствительные к молекулярному кислороду: нитрогеназа, гидрогеназа и рибулозодифосфаткарбоксилаза.
Нитрогеназная система, катализирующая фиксацию молекулярного азота, как известно, состоит из двух металлопротеинов: белка, содержащего железо и молибден, и белка, в состав которого входит только железо. Каждый белок необходим для проявления каталитической активности. Молекулярный кислород оказывает повреждающее действие на оба белка нитрогеназы, но более чувствителен к O2 Fe-белок.
Чувствительность белков нитрогеназы к O2 определяется прежде всего чувствительностью их металлоцентров, которые участвуют как в связывании субстрата, так и в переносе электронов. Поскольку при этом может происходить и ступенчатое восстановление O2 по одноэлектронному механизму, в качестве продуктов такого восстановления возникают супероксидные ионы, перекись водорода и синглетный кислород, вносящие свой вклад в окислительное повреждение нитрогеназы.
Нитрогеназные белки являются не единственным компонентом азотфиксирующей системы, чувствительным к O2. Ферредоксины и флаводоксины, донирующие электроны на нитрогеназу, могут автоокисляться и подвергаться необратимым окислительным повреждениям.
Гидрогеназы многих прокариот также обнаруживают высокую чувствительность к молекулярному кислороду, которая in vitro в большой мере зависит от метода выделения и степени очистки. Как правило, более устойчивы к O2 неочищенные ферментные препараты. По сравнению с мембрансвязанным ферментом устойчивость к O2 гидрогеназы, отделенной от мембраны, обычно ниже. Фермент, полученный из клеток анаэробов, более чувствителен к O2, чем выделенный из клеток аэробных прокариот.
Каталитическая активность рибулозодифосфаткарбоксилазы, фермента, катализирующего фиксацию CO2 у подавляющего большинства автотрофных прокариот, зависит ст парциальных давлений СO2 и O2. При высокой концентрации O2 и низкой — CO2 преобладает оксигеназная реакция:

Молекулы O2 и СO2 конкурируют за каталитический центр фермента. И хотя сам фермент не обнаруживает повышенной чувствительности к молекулярному кислороду и, следовательно, не повреждается при его высокой концентрации, увеличение O2 в среде приводит к изменению функционирования рибулозодифосфаткарбоксилазы. Протекание ферментативной реакции по оксигеназному пути приводит к истощению клеточного пула молекул рибулозодифосфата и, как следствие этого, понижению активности восстановительного пентозофосфатного цикла в клетке.
Помимо существования в основной форме в биологических реакциях и под действием различных физико-химических факторов возникают продукты неполного восстановления O2, более реакционно способные и обладающие высокой токсичностью для клетки. Как известно, для полного восстановления молекулярного кислорода, приводящего к образованию молекулы воды, требуются 4 электрона:
O2 + 4H+ + 4e– ® 2H2O
У большинства прокариот имеются ферменты, катализирующие реакции одновременного переноса 4 электронов на O2, при которых не обнаружено каких-либо промежуточных продуктов восстановления O2. Это цитохромоксидазы и некоторые медьсодержащие ферменты. Возможно, что в этих реакциях и возникают короткоживущие продукты неполного восстановления O2, но они остаются связанными с ферментами, не выходят в цитоплазму и практически не наносят вреда клетке.
Супероксидный анион. Если восстановление молекулярного кислорода происходит ступенчато, то при переносе 1 электрона на O2 образуется надпероксидный (супероксидный) анион:
O2 + e– ® O2–.
Последний содержит неспаренный электрон, поэтому является отрицательно заряженным радикалом (анион-радикалом). Он может протонироваться с образованием нейтрального гидропероксидного радикала:
O2–. + H+ ® HO2.
В последнее время признание получила точка зрения, согласно которой основную опасность для организмов представляют продукты, образующиеся при одноэлектронном восстановлении молекулы O2, одним из которых является супероксидный анион.
Можно назвать много биохимических реакций, приводящих к его возникновению. Супероксидные анионы генерируются при взаимодействии с молекулами O2 различных компонентов (восстановленные флавины, хиноны, тиолы, FeS-белки), а также в реакциях, катализируемых рядом флавопротеиновых ферментов. Наконец, в процессе фотосинтеза имеет место поток электронов. Большинство реакций фотосинтеза — это реакции одноэлектронного переноса. Поэтому в системе часто возникают супероксидные анионы. Помимо реакций биологической природы O2–. могут возникать вне клетки в водных растворах при воздействии на них ультразвуком, в результате фотохимических, химических и электрохимических процессов.
Опасность любых реакционно активных соединений в значительной степени зависит от их стабильности. В этом плане ионы O2–. весьма опасны, так как время их "жизни" в водной среде продолжительнее, чем у остальных O2-производных радикалов. Поэтому экзогенно возникшие O2–. могут проникать в клетку и (наряду с эндогенными) участвовать в реакциях, приводящих к различным повреждениям: перекисном окислении ненасыщенных жирных кислот, окислении SH-групп белков, повреждении ДНК и др. Токсичность супероксидных анионов может увеличиваться за счет вторичных реакций, ведущих к образованию гидроксидных радикалов (OH.) и синглетного кислорода (*O2).
Многие прокариоты, относящиеся к разным физиологическим группам, в том числе и строго анаэробные виды, имеют специфическую защиту в виде фермента супероксиддисмутазы, осуществляющего перехват ионов O2–. и катализирующего их дисмутацию. Образующиеся супероксидные анионы дисмутируют в реакции, протекающей спонтанно (3) или катализируемой супероксиддисмутазой (4):
O2–. + O2–. + 2H+ ® H2O2 + *O2; (3)
O2–. + O2–. + 2H+ ® H2O2 + O2. (4)
Различия между обеими реакциями в их скоростях (скорость ферментативной реакции приблизительно на четыре порядка выше, чем спонтанной), а также в том, что при спонтанной реакции дисмутации одним из первоначально возникающих продуктов является синглетный кислород, в то время как при ферментативной реакции образующийся кислород находится в основном триплетном состоянии.
Гидроксидный радикал. Супероксиданион может взаимодействовать с H2O2 с образованием гидроксидного радикала (OH.), превосходящего O2–. по окислительной активности и токсичности:
O2–. + H2O2 + H+ ® O2 + H2O + OH.. (5)
Источником возникновения ОН. могут служить реакции одноэлектронного окисления перекиси водорода, катализируемые железосодержащими соединениями, всегда имеющимися в клетках:
H2O2 + Fe2+ ® Fe3+ + OH– + OH.. (6)
Помимо указанных выше реакций гидроксидные радикалы образуются также при радиолизе воды и в низких концентрациях обычно присутствуют в водных растворах. OH. из всех известных окислителей является самым сильным, вызывающим радиационные повреждения многих типов биополимеров.
Перекись водорода. Перенос 2 электронов на O2 приводит к образованию перекисного аниона (7) или перекиси водорода (8):
O2 + 2e– ® O22–; (7)
O2 + 2H+ + 2e– ® H2O2. (8)
Катализировать перенос 2 электронов на O2 могут содержащиеся в клетках прокариот оксидазы флавиновой природы и некоторые цитохромы. Источником H2O2 могут быть реакции автоокисления некоторых негемовых FeS-белков, а также описанные выше реакции дисмутации супероксидных радикалов (реакции 3 и 4). Перекись водорода образуется у всех аэробов и факультативных анаэробов, растущих в аэробных условиях, так что ее возникновение в клетках прокариот — естественный процесс.
Перекись водорода — наиболее стабильный из промежуточных продуктов восстановления O2, но и наименее реакционноспособный. У большинства аэробных прокариот H2O2 быстро разлагается с помощью гемсодержащих ферментов каталазы и пероксидазы. В отсутствие их H2O2 может накапливаться в летальных для организма концентрациях.
H2O2 вызывает окисление SH-групп в белках, перекисное окисление ненасыщенных жирных кислот. Однако эти реакции протекают с измеримыми скоростями, если концентрация H2O2 в клетке будет на четыре порядка выше той, которая обычно достигается in vivo. Поэтому не исключено, что перекись водорода опасна не из-за прямого взаимодействия с компонентами клетки, а потому, что, реагируя с O2–. (реакция 5) или ионами Fe2+ (реакция 6), может приводить к образованию гидроксидного радикала.
В 20-х гг. большой популярностью пользовалась теория, объясняющая токсичность O2 накоплением в клетке перекиси водорода. Однако позднее были обнаружены более токсичные для клетки формы O2 среди первичных и вторичных продуктов его одноэлектронного восстановления (O2, OH., *O2).
Синглетный кислород. В норме O2 находится в стабильном состоянии, называемом триплетным и характеризующемся наименьшим уровнем молекулярной энергии. В определенных условиях молекула O2 переходит в одно из двух возбужденных синглетных состояний (*O2), различающихся степенью энергизованности и длительностью "жизни". У большинства живых клеток в темноте основным источником синглетного кислорода служит спонтанная дисмутация супероксидных анионов (см. реакцию 3). Синглетный кислород может возникать также при взаимодействии двух радикалов:
O2–. + OH. ® OH– + *O2. (9)
Вероятно, любая биологическая система, в которой образуется O2, может быть активным источником синглетного кислорода. Однако последний возникает и в темновых ферментативных реакциях в отсутствие O2 .
Давно было известно, что на свету токсичность молекулярного кислорода для живых организмов повышается. Этому способствуют находящиеся в клетке вещества, поглощающие видимый свет, — фотосенсибилизаторы 63 Многие природные пигменты могут быть фотосенсибилизаторами. В клетках фотосинтезирующих организмов активными фотосенсибилизаторами являются хлорофиллы и фикобилипротеины. Окисление биологически важных молекул под влиянием видимого света в присутствии молекулярного кислорода и фотосенсибилизатора получило название фотодинамического эффекта.
63 Фотосенсибилизаторы — молекулы, способные поглощать свет и индуцировать химические реакции, которые в их отсутствие не происходят. Способность поглощать свет обусловлена наличием в молекулах хромофорных группировок, содержащих обычно циклические ядра. Известно более 400 веществ, обладающих свойствами фотосенсибилизаторов. Среди природных веществ фотосенсибилизаторами являются хлорофиллы, фикобилины, порфирины и промежуточные продукты их синтеза, ряд антибиотиков, хинин, рибофлавин и др. Некоторые фотосенсибилизаторы действуют только в присутствии O2, вызывая фотодинамический эффект.
Поглощение видимого света приводит к переходу молекулы фотосенсибилизатора в возбужденное синглетное состояние (*Д):
Д+hn ® *Д
Молекулы, перешедшие в синглетное состояние, могут возвращаться в основное (Д) или переходить в долгоживущее триплетное состояние (*Д), в котором они фотодинамически активны. Установлено несколько механизмов, с помощью которых возбужденная молекула (*Д) может вызывать окисление молекулы субстрата. Один из них связан с образованием синглетного кислорода. Молекула фотосенсибилизатора в триплетном состоянии реагирует с O2 и переводит его в возбужденное синглетное состояние:
TД + O2 ® Д + *O2.
Синглетный кислород окисляет молекулу субстрата (В):
B +*O2 ® ВO2.
Фотодинамический эффект обнаружен у всех живых организмов. У прокариот в результате фотодинамического действия индуцируются повреждения многих типов: утрата способности формировать колонии, повреждение ДНК, белков, клеточной мембраны. Причина повреждений — фотоокисление некоторых аминокислот (метионина, гистидина, триптофана и др.), нуклеозидов, липидов, полисахаридов и других клеточных компонентов.
Клетки содержат вещества, выполняющие функцию тушения синглетного кислорода и понижающие возможность структурных и иных повреждений, вызываемых им. Одним из "тушителей" синглетного кислорода служат каротиноиды, защищающие фотосинтезирующие организмы от летальных эффектов, фотосенсибилизируемых хлорофиллом. Перехватчиками *O2 являются также различные биологически активные соединения: липиды, аминокислоты, нуклеотиды, токоферолы и др.
Озон и атомарный кислород. Продуктами молекулярного кислорода являются также атомарный кислород (O) и озон (O3). Известно, что молекулярный кислород сильно поглощает свет в дальней УФ-области (160–240 нм). Поглощенный фотон вызывает диссоциацию молекулы кислорода на два атома:
O2 +hn ® 2O.
Затем спонтанно протекает реакция, приводящая к образованию молекулы озона:
O2 + O ® O3
Озон может возникать из молекулярного кислорода в воздухе при сильных электрических разрядах, а также при электролизе воды и в некоторых реакциях, где он сопровождает образование O2. В некоторых реакциях окисления с помощью озона образуется синглетный кислород. Как окислители озон и атомарный кислород сильнее O2. Озон может реагировать практически со всеми типами соединений с образованием радикалов.
Защитные механизмы клетки
Для нейтрализации токсических форм O2 существующие прокариоты выработали различные защитные механизмы, которые могут быть разделены на несколько типов. В основе систем защиты первого типа лежит активность специальных ферментов, для которых разложение токсических форм O2 является основной и в ряде случаев единственной функцией. В системах защиты второго типа для разрушения токсических форм O2 используются определенные клеточные метаболиты. Как правило, в этом случае участие в защите клетки от токсических эффектов производных O2 является не единственной функцией этих метаболитов. Наконец, к защитным механизмам особого типа может быть отнесен ряд приспособлений, выработанных прокариотами на разных уровнях: популяционном, физиологическом, структурном. Более вероятно, что они были созданы для других целей, но оказались полезными и для детоксикации O2.
Ферментные системы защиты. Передовой линией защиты от токсического действия производных O2 являются ферменты: супероксиддисмутаза, захватывающая молекулы О2 (реакция 4), каталаза и пероксидаза, улавливающие H2O2:
Они сводят до минимума концентрацию в клетке O2–. и H2O2 и не дают им возможности взаимодействовать с образованием ОН. (реакция 5).
Супероксиддисмутаза обнаружена у хемотрофных прокариот, использующих O2 (облигатно и факультативно аэробных форм), а также у изученных представителей из групп фотосинтезирующих прокариот. Среди анаэробов фермент найден у подавляющего большинства аэротолерантных форм. Исключение составляют некоторые молочнокислые бактерии, однако в клетках большинства из них содержатся высокие концентрации (до 30 мМ) ионов двухвалентного марганца. Оказалось, что Мп2+, для которого показана способность окисляться под действием O2, в таких концентрациях способен так же эффективно убирать образующиеся супероксидные ионы, как это делает супероксиддисмутаза, содержание которой в клетке обычно поддерживается на микромолярном уровне. Таким образом, у этих молочнокислых бактерий функцию нейтрализации O2 выполняют ионы Mn2+.
В клетках некоторых видов молочнокислых бактерий не найдено ни супероксиддисмутазы, ни высоких концентраций Mn2+. Эти виды характеризуются очень высокой чувствительностью к O2.
Среди облигатных анаэробов супероксиддисмутаза обнаружена у многих представителей рода Clostridium. Изучение их устойчивости к O2 обнаруживает четкую связь с содержанием в клетках этого фермента. Виды, имеющие супероксиддисмутазу, характеризуются умеренной или даже высокой устойчивостью к O2 по сравнению с видами, у которых этот фермент отсутствует. Супероксиддисмутаза найдена у разных видов строго анаэробных бактерий. Число организмов с не выявленной до сих пор супероксиддисмутазой очень мало.
Обнаружение супероксиддисмутазы у строгих анаэробов (гораздо более распространенное, чем предполагали раньше) ставит вопрос о ее физиологической роли у этих организмов. Способность последних расти только в бескислородной среде делает неясным функции фермента в данных условиях. Возможно, что только при попадании строгого анаэроба в неблагоприятные для него аэробные условия синтез фермента индуцируется молекулярным кислородом, что и обеспечивает организму защиту от O2 в этих условиях.
Супероксиддисмутаза — фермент, содержащий в активном центре в качестве простетической группы ионы металла. У прокариот — это атомы марганца и/или железа. Большинство изученных супероксиддисмутаз построено из двух идентичных субъединиц, каждая из которых содержит по одному атому металла. Fe- и Mn-ферменты сходны по аминокислотной последовательности. Попытки выявить связь между физиологическими и иными особенностями организмов и металлоформой содержащегося в них фермента не привели к определенному заключению. И та и другая формы супероксиддисмутазы обнаружены у представителей грамположительных и грамотрицательных прокариот, среди фото- и хемотрофов, облигатных анаэробов, аэробов и факультативно анаэробных форм. Более того, обе металлоформы супероксиддисмутазы могут присутствовать у одного организма и даже входить в состав молекулы одного фермента. Для некоторых видов показано, что синтез того или иного типа фермента зависит от наличия ионов металла в среде культивирования.
Супероксиддисмутаза изученных хемотрофных прокариот — не связанный с мембранами фермент, локализованный в цитоплазме. У Е. coli, в клетках которой обнаружены Fe-, Mn- и Fe/Mn-формы фермента, Fe-супероксиддисмутаза локализована в периплазматическом пространстве, а Mn-содержащий фермент — в цитоплазме. В связи с этим высказывается предположение, что металлоформы фермента играют разную роль в защите клетки от O2–.: Fe-содержащий фермент защищает клетку от экзогенных супероксидных анионов, а Mn-содержащий — от эндогенных.
Особо остро стоит проблема защиты от молекулярного кислорода и его производных в клетках цианобактерий. Вероятно, именно они впервые в наибольшей степени ощутили последствия токсических эффектов кислорода. Супероксиддисмутаза найдена у всех цианобактерий. В клетках Anacystis nidulans (Synechococcus) Fe-супероксиддисмутаза, составляющая до 90% от общего количества фермента, локализована в цитозоле клетки, а Mn-содержащая форма — в тилакоидах. Функция последней формы фермента сводится, вероятно, к перехвату ионов O2–., возникающих в процессе фотосинтетического электронного транспорта.
Каталаза и пероксидаза. Перекись водорода разрушается двумя классами родственных ферментов, катализирующих ее двухэлектронное восстановление до H2O и использующих в качестве донора электронов H2O2 в случае каталазы (реакция 10) или различные органические соединения в случае пероксидазы (реакция 11).
Каталазная и пероксидазная активности обнаружены у всех облигатно и факультативно аэробных прокариот. Среди облигатных анаэробов эти ферменты распространены значительно в меньшей степени, чем супероксиддисмутаза. Обнаружены многие строгие и аэротолерантные анаэробы, содержащие супероксиддисмутазу, но не содержащие каталазы. К их числу можно отнести и те молочнокислые бактерии, у которых дисмутация образующихся ионов O2–. обеспечивается Mn2+, находящимся в клетках в высоких концентрациях.
Отсутствие каталазы у молочнокислых бактерий связано с тем, что они не могут синтезировать гем — простетическую группу фермента, но способны к синтезу апофермента. При добавлении гемовых групп извне молочнокислые бактерии образуют гемсодержащую каталазу. У ряда молочнокислых бактерий обнаружена каталаза, не содержащая гемовой группы, названная поэтому псевдокаталазой. Выделенный фермент состоит из шести идентичных полипептидных цепей, соединенных между собой нековалентными силами. Каждая субъединица содержит 1 атом марганца.
Перекись водорода, возникающая в результате взаимодействия клеток с O2, устраняется и неферментативными путями. Известно, что ионы Fe2+ в водном растворе ускоряют восстановление H2O2 до H2O. В клетке всегда содержится некоторое количество ионов железа. Разрушение H2O2 может происходить и за счет выделяющихся в культуральную среду восстановленных веществ.
Для анаэробных прокариот, способных переносить контакт с O2 и его производными в относительно небольших масштабах, необходимо присутствие в клетках супероксиддисмутазы, "убирающей" O2–.. Наличие каталазы при этом не обязательно, поскольку возникающая в реакции дисмутации и других реакциях перекись водорода разлагается спонтанно или с участием неферментативных катализаторов, и организмы в целом справляются с ней в этих условиях. Таким образом, при осуществлении энергетического метаболизма анаэробного типа для устранения токсических эффектов O2. достаточно одной ферментной преграды в виде супероксиддисмутазы.
Резкое возрастание масштабов взаимодействия прокариот с O2 при функционировании метаболизма аэробного типа делает неэффективными неферментативные пути устранения H2O2. Для разложения перекиси водорода, образующейся в больших количествах, необходимы специальные ферменты, повышающие скорость разложения H2O2 на несколько порядков. Это обеспечивается каталазой и пероксидазой. Таким образом, в условиях активного взаимодействия клеток с O2, делающего возможным аэробную жизнь, система ферментной защиты от его токсических эффектов сформирована с участием супероксиддисмутазы, каталазы и пероксидазы в качестве необходимых компонентов (рис. 87).
Механизмы защиты с помощью клеточных метаболитов. Защита против одного из самых токсичных производных O2 — синглетного кислорода — осуществляется с помощью разных биологически важных молекул. Все виды тушения *O2 можно разделить на физические и химические. Физическим называют тушение, которое не приводит к разрушению тушителя:
*O2 + A ® O2 + A
Химическое тушение приводит к окислению тушителя:
*O2 + A ® O2 + Aок
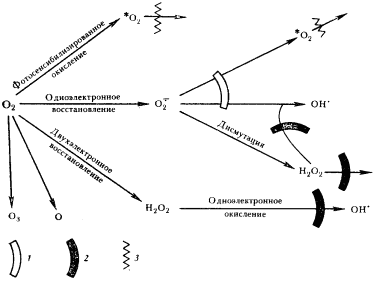
|
| Рис. 87. Системы клеточной защиты от токсических производных молекулярного кислорода: 1 — супероксиддисмутаза, ионы Mn2+; 2 — каталаза, пероксидаза; 3 — тушение каротиноидами |
Преимущественно по химическому механизму тушение *O2 осуществляется насыщенными жирными кислотами, липидами, аминокислотами, нуклеотидами и другими соединениями. Механизмы химического тушения разнообразны, но в большинстве случаев начальной стадией является образование лабильной циклической перекиси с последующим ее разложением, которое приводит к возникновению свободных радикалов. Химическое тушение *O2 может приводить в клетке к существенным деструктивным последствиям. К тушению в основном по физическому механизму способны молекулы разных химических соединений. Наиболее эффективны в этом отношении каротиноиды, широко распространенные в мире прокариот. Они обнаружены в клетках многих аэробных хемотрофов, являются обязательным компонентом пигментного аппарата всех фототрофов. В клетках фотосинтезирующих организмов, как отмечалось выше, активными фотосенсибилизаторами являются хлорофиллы. Однако возможность фотоокислительных эффектов в условиях функционирования фотосинтетического аппарата довольно низка, во-первых, из-за чрезвычайно короткого (10–11 с) времени пребывания хлорофилла в возбужденном состоянии и, во-вторых, из-за защиты клеток от фотоокисления каротиноидами.
Впервые роль каротиноидов в предотвращении летального эффекта, вызываемого фотоокислением, была показана при изучении бескаротиноидного мутанта пурпурной бактерии Rhodopseudomonas sphaeroides. Исходная культура хорошо росла фототрофно в анаэробных условиях, но могла также расти на свету, и в темноте в аэробных условиях. Полученный из нее мутант, лишенный каротиноидов, обладал низкой скоростью роста на свету в анаэробных условиях и в темноте в аэробных условиях, но быстро погибал при перенесении на свет + воздух.
Фотоокислительные повреждения могут развиваться и у нефотосинтезирующих прокариот, так как в их клетках также имеются окрашенные молекулы, поглощающие видимый свет, которые могут функционировать как фотосенсибилизаторы. Действие каротиноидов не ограничивается только их участием в защите от фотодинамического эффекта. Они гасят синглетное состояние кислорода независимо от того, в каких реакциях он возникает: на свету или в темноте.
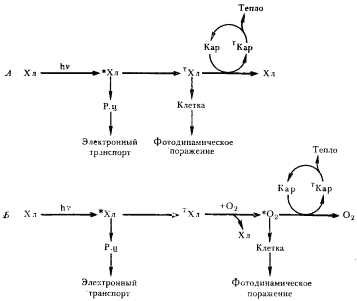
|
| Рис. 88. Механизмы защитного действия каротиноидов. А — тушение каротиноидами триплетного состояния хлорофилла. Б — тушение каротиноидами синглетного кислорода: *Хл — молекула хлорофилла в возбужденном синглетном состоянии; TХл — молекула хлорофилла в возбужденном триплетном состоянии; TKap — молекула каротиноида в возбужденном триплетном состоянии; Р. ц. — реакционный центр |
Механизм защитного действия каротиноидов у фотосинтезирующих организмов заключается в следующем (рис. 88). Молекула хлорофилла, поглотившая свет, быстро (10–12 с) переносит энергию синглетного возбужденного состояния в реакционный центр. Из 104 поглощенных квантов света приблизительно 4 приводят к переходу молекулы хлорофилла в возбужденное триплетное состояние. Возникает возможность фотодинамического поражения. Каротиноиды могут участвовать в трех защитных реакциях: 1) непосредственно тушить триплетное состояние хлорофилла, переводя его в основное состояние (рис. 88, А); возникающая при этом триплетная молекула каротиноида отдает избыточную энергию в виде тепла и возвращается в основное состояние; 2) триплетный хлорофилл не гасится каротиноидами; происходит его взаимодействие с O2, переводящее последний в возбужденное синглетное состояние; синглетный кислород гасится каротиноидами (рис. 88, Б); 3) синглетный кислород, не подвергшийся гашению каротиноидами по физическому механизму, может взаимодействовать с ними в химической реакции, приводящей к окислению каротиноидов. Участие каротиноидов в любой из трех описанных выше реакций будет снижать уровень образования в клетке *O2.
Приспособления прокариот, помогающие им в защите от токсических эффектов молекулярного кислорода. В клетках тех облигатно анаэробных клостридиев, у которых не обнаружено ни супероксиддисмутазы, ни каталазы, доступным средством нейтрализации O2 служит вытеснение его из среды культивирования с помощью активно выделяющихся газообразных продуктов (CO2 и H2), сопровождающих брожение, а также поглощение клеточной суспензией кислорода из среды, приводящее к гибели части клеток, но дающее возможность оставшимся размножаться в условиях пониженного содержания O2.
Молочнокислыми бактериями в направлении защиты от молекулярного кислорода сделан определенный шаг вперед. Эти бактерии — единственная группа прокариот, не имеющих гемсодержащей каталазы, способных расти в присутствии воздуха. Поиски механизмов нейтрализации O2 и его производных привели к обнаружению у них помимо супероксиддисмутазы и высокой внутриклеточной концентрации ионов Mn2+, осуществляющих разложение O2–., псевдокаталазы, а также каталазо- и пероксидазоподобной активности. У отдельных представителей молочнокислых бактерий просматривается более четко выраженная степень приспособленности к O2, приводящая к попыткам определенного его полезного использования. Для некоторых молочнокислых бактерий рода Lactobacillus показано ускорение гликолитического разложения глюкозы в аэробных условиях. Это связано с тем, что в аэробных условиях водород с НАД-H2 может прямо передаваться на O2, освобождая часть пировиноградной кислоты от ее акцепторной функции, как это происходит при обычном молочнокислом брожении. Освобожденная от этой "обязанности" пировиноградная кислота может теперь окисляться до ацетил-КоА, последующее метаболизирование которого до ацетата приводит к синтезу молекулы АТФ. Как можно видеть, участие кислорода в этом процессе прямо не связано с получением клеткой энергии (при передаче водорода с НАД-H2 на O2 энергия в форме АТФ не образуется), т. е. вся энергия получается за счет субстратного фосфорилирования, но O2, беря на себя акцепторную функцию, освобождает часть пирувата, которая может использоваться клеткой по энергетическому пути, что в конечном итоге приводит к повышению энергетического выхода брожения. Таким образом, прямое окисление части восстановленных переносчиков электронов в процессе брожения может иметь не только отрицательные, но и положительные последствия.
Как отмечалось выше, очень чувствителен к O2 процесс азотфиксации. Несмотря на это, способность фиксировать N2 широко распространена среди прокариот, различающихся отношением к молекулярному кислороду; она присуща хемотрофам и фототрофам, в том числе цианобактериям, осуществляющим кислородный фотосинтез. Фиксировать N2 могут свободноживущие формы и прокариоты, находящиеся в симбиозе с эукариотными организмами.
Изучение средств защиты этого процесса у прокариот показало, что в большинстве случаев она далека от 100%-ной эффективности. Среди аэробных азотфиксаторов можно выделить лишь немногие организмы, способные расти в среде с N2 в условиях равновесия с воздухом. Большинство может расти и фиксировать N2 только в условиях пониженной концентрации молекулярного кислорода, т. е. в микроаэробных условиях. Защита нитрогеназы в клетках факультативных анаэробов еще менее эффективна: они могут осуществлять активно фиксацию азота только в анаэробных условиях.
К числу аэробных азотфиксаторов относятся представители рода Azotobacter, у которых обнаружены различные защитные приспособления. Одно из них связано с резким увеличением дыхательной активности клеток, осуществляющих азотфиксацию в аэробных условиях. Дыхание в значительной мере служит в этом случае для "связывания" внутриклеточного O2. При этом обнаружены существенные перестройки как в клеточном строении азотобактера, выражающиеся в интенсивном развитии системы внутрицитоплазматических мембран, так и в организации самой дыхательной цепи, локализованной в этих мембранах. Дыхательная цепь Azotobacter vinelandii достаточно сложная, имеет разветвления на путях переноса электронов на уровне цитохрома b. Транспорт электронов, сопряженный с фосфорилированием, происходит по пути:
цитохромы b ® c4 ® c5 ® a1.
При осуществлении "дыхательной" защиты возрастает активность электронного транспорта по ветви: цитохромы b ® d, не связанной с запасанием энергии. Это приводит к тому, что несмотря на возрастание общей активности дыхания сопряжение электронного транспорта с запасанием энергии снижается. Таким образом, происходит "сжигание" части углеродных субстратов, которые используются для восстановления O2, без запасания при этом клеткой энергии.
В дополнение к вынужденному "принесению в жертву" части источников углерода высокие концентрации O2 вызывают в клетке обратимые изменения структуры нитрогеназы, делающие чувствительные к молекулярному кислороду участки менее доступными для него. Высказываются разные предположения относительно того, как осуществляется "конформационная" защита. Возможно, при этом происходит изменение взаимного расположения двух нитрогеназных белков. Не исключено участие в защите такого типа клеточной мембраны. Определенная стабилизация нитрогеназы в условиях высокой концентрации O2 происходит при добавлении к ферментному комплексу двухвалентных катионов. Наконец, обнаружены специальные защитные белки, образующие комплексы с нитрогеназными белками и приводящие к повышению их стабильности в присутствии O2. Никаких других функций, кроме защитной, у этих белков пока не найдено.
Большинство азотфиксирующих прокариот способны фиксировать молекулярный азот в микроаэробных условиях. К числу защитных приспособлений у них относятся: образование слизи, препятствующей диффузии в клетку O2 и тем самым создающей вокруг нее микроаэробную зону; формирование клеточных скоплений, затрудняющих доступ O2 к клеткам, расположенным внутри скопления, которым, таким образом, создаются более благоприятные условия для азотфиксации; существование азотфиксирующих видов в ассоциации с неазотфиксирующими аэробными гетеротрофами, защищающими нитрогеназу азотфиксаторов от доступа O2.
Специфические приспособления для защиты нитрогеназы от высоких концентраций O2 выработаны симбиотическими азотфиксаторами — клубеньковыми бактериями. Уже сами клубеньки, места активного размножения бактерий и фиксации ими N2, следует рассматривать как структуру, одним из назначений которой является ограничение доступа внутрь молекулярного кислорода. Эту же функцию выполняет содержащийся в клубеньках леггемоглобин (белок, аналогичный гемоглобину), способный активно связывать O2 и контролировать его поступление в бактероиды. В любом случае при осуществлении метаболизма аэробного типа дыхание также будет препятствовать накоплению в клетке молекулярного кислорода.
Наиболее остро стоит проблема защиты процесса азотфиксации от O2 в группе цианобактерий. У всех цианобактерий нитрогеназа чувствительна к O2, имеющему внеклеточное и внутриклеточное происхождение. В соответствии с этим у них можно выделить приспособления, направленные на защиту от экзогенного кислорода, и те, которые предназначены для нейтрализации O2. образующегося внутри клетки в процессе фотосинтеза.
Если все азотфиксирующие цианобактерий рассматривать под углом зрения степени защиты процесса азотфиксации от O2, то их можно разделить на две группы. К первой группе относятся цианобактерий, у которых защита азотфиксации от O2 наименее эффективна, поэтому вегетативные клетки могут фиксировать N2 только в анаэробных или микроаэробных условиях. Вторую группу составляют цианобактерий, у которых для осуществления азотфиксации в аэробных условиях сформированы специализированные клетки — гетероцисты.
У безгетероцистных цианобактерий защита нитрогеназы вегетативных клеток от O2, в первую очередь эндогенного, осуществляется с помощью разделения во времени процессов фотосинтеза и азотфиксации, непрерывного синтеза нитрогеназы, высокой активности супероксиддисмутазы в сочетании с каталазной и пероксидазной активностями. В центре филаментов некоторых безгетероцистных форм часто выделяются слабопигментированные вегетативные клетки, у которых предположительно подавлена способность к фотосинтетической фиксации CO2 и тем самым созданы более благоприятные условия для азотфиксации. (Это не гетероцисты, но, вероятно, именно из них впоследствии развились гетероцисты как центры азотфиксации в аэробных условиях). Средством защиты от экзогенного O2 служит синтез большого количества слизи, часто окружающей клетки азотфиксирующих цианобактерий. Существование в виде колониальных форм также может обеспечивать создание анаэробных условий для клеток, располагающихся в центральной части колонии.
Наиболее совершенна защита от эндогенного и экзогенного молекулярного кислорода в гетероцистах. Гетероцисты не способны к фотосинтетическому выделению O2. А высокие активности окислительного пентозофосфатного пути, поставляющего электроны в дыхательную цепь, где они акцептируются O2, повышенные уровни супероксиддисмутазы сравнительно с вегетативными клетками, образование гетероцистами молекулярного водорода, толстая многослойная оболочка, выполняющая функцию газового барьера, — все это надежно защищает азотфиксирующую систему в гетероцистах от инактивации молекулярным кислородом.
Таким образом, можно только предполагать, что механизмы нейтрализации молекулярного кислорода на различных этапах эволюции взаимодействия с ним клеток были неодинаковы. H2 каком-то этапе возникли ферментные реакции, катализирующие включение O2 в метаболизм прокариот.
МОЛЕКУЛЯРНЫЙ КИСЛОРОД В МЕТАБОЛИЗМЕ ПРОКАРИОТ
Тот факт, что все существующие на Земле прокариоты, даже строгие анаэробы, в присутствии O2 его поглощают, указывает на осуществление ими каких-то реакций взаимодействия с молекулярным кислородом. По отношению к O2 все прокариоты могут быть разделены на несколько физиологических групп (см. рис. 34). Такое подразделение говорит о необходимости или вреде молекулярного кислорода, но не раскрывает механизмов взаимодействия с ним клетки. Действительно, сейчас мы знаем, что O2 может быть необходим клетке для получения энергии или же для осуществления всего одной реакции, не имеющей энергетического значения.
| Неферментативное окисление | Ферментативное поглощение O2 (дыхание) | ||
| свободное окисление | окисление, сопряженное с запасанием энергии | ||
| нефосфорилируюшее | фосфорилирующее | ||
Рис. 89. Пути использования порции молекулярного кислорода, поглощенного клеткой. Объяснение см. в тексте (по Скулачеву, 1969).
H2 основании изучения энергетических процессов, происходящих в митохондриях животных клеток, В. П. Скулачев предложил следующую классификацию реакций взаимодействия клетки с молекулярным кислородом (рис. 89). Порцию поглощенного клеткой O2 можно разделить на две неравные части. Основная масса кислорода потребляется клеткой с участием клеточных ферментных систем. Поглощение клеткой какой-то части O2 не связано с ее ферментными системами. Иллюстрацией последнего служит хорошо известный факт активного поглощения кислорода суспензией убитых прогреванием клеток. В этом случае поглощение кислорода — чисто химический процесс, связанный с окислением определенных химических веществ клетки, например SH-групп клеточных белков. Нельзя исключить возможность протекания процессов аналогичной природы и в суспензии живых клеток. В свою очередь ферментативное поглощение молекулярного кислорода — дыхание64 — подразделяется на окисление, сопряженное с запасанием энергии, и свободное окисление, т. е. не связанное с запасанием энергии для клетки. Окислительные ферментативные реакции с участием O2, относимые к категории свободного окисления, — это реакции, в результате которых энергия выделяется в виде тепла65. К этой категории процессов относятся реакции, катализируемые моно- и диоксигеназами, в которых имеет место прямое включение кислорода в молекулу окисляемого вещества, а также реакции, катализируемые некоторыми оксидазами.
64 Термин "дыхание" впервые был введен для обозначения определенного процесса, связанного с жизнедеятельностью высших организмов (растений и животных). Два основных признака характеризовали этот процесс: газообмен с внешней средой с непременным участием O2; необходимость для жизнедеятельности организма. Принципиальное сходство процесса дыхания на клеточном уровне у всех высших организмов делало употребление этого термина удобным, а обозначаемое им понятие достаточно четким. Сложности возникли при применении термина "дыхание" для обозначения функционально аналогичных процессов у прокариот в силу их необычайного разнообразия. В нашем понимании термин "дыхание" распространяется на все процессы ферментативного поглощения клеткой молекулярного кислорода.
65 Реакции свободного окисления имеют важное значение в осуществлении терморегуляции у животных при охлаждении организма.
Ферментативное поглощение O2, сопряженное с запасанием энергии, подразделяется на процессы, не связанные с фосфорилированием, и процессы, сопровождающиеся фосфорилированием. В первом случае окисление, сопряженное с запасанием энергии, не связано с трансформированием свободной энергии в форму макроэргических фосфатных связей. Известно, что в клетке существуют две универсальные формы энергии: химическая и электрохимическая (DmH+). Один из путей получения энергии в форме трансмембранного электрохимического градиента Н+ связан с переносом электронов на O2. Энергия в этой форме может использоваться клеткой для совершения разного вида работы (см. рис. 27). Химическая энергия заключена в основном в соединениях, содержащих макроэргические фосфатные связи, и в первую, очередь в молекулах АТФ. Но на промежуточных этапах катаболических процессов, связанных в конечном итоге с поглощением O2, образуются метаболиты, содержащие богатые энергией связи, например тиоэфирные (C~S — КоА). Эти соединения могут непосредственно обеспечивать энергией некоторые биосинтетические процессы.
Наконец, при фосфорилирующем окислении энергия, высвобождаемая при электронном транспорте на молекулярный кислород и возникающая первоначально в форме DmH+, с помощью протонной АТФ-синтазы трансформируется в химическую форму в молекулах АТФ. В отличие от высших организмов, где достигнута высокая степень сопряжения между переносом электронов и фосфорилированием, т. е. этот путь предстает уже в сложившемся виде, у современных прокариот мы обнаруживаем различные пути переноса электронов и разные степени сопряжения электронного транспорта с фосфорилированием. Все перечисленные типы окислительных процессов с участием O2, протекающие в высокоорганизованной клетке, обнаруживаются и у прокариот.
В основу классификации, предложенной В. П. Скулачевым, положено рассмотрение всех реакций взаимодействия клетки с молекулярным кислородом под углом зрения их "энергетической значимости". По химическим механизмам, лежащим в основе этих реакций, все они могут быть разделены на 2 типа. К первому типу относятся реакции, катализируемые кислородными трансферазами, или диоксигеназами, в которых имеет место прямое присоединение молекулы кислорода к молекуле метаболита:
A + O2 ® AO2.
Одна молекула субстрата может акцептировать оба атома молекулы кислорода, как это имеет место в приведенной выше реакции. Акцепторами O2 могут быть молекулы двух разных субстратов:
X + Y + O2 ® XO + YO.
Все подобные реакции представляют собой свободное окисление и не связаны с получением клеткой энергии.
В реакциях второго типа электроны идут к кислороду, выполняющему функцию конечного акцептора. В этом случае 1, 2 или 4 электрона в зависимости от природы переносчика акцептируются молекулой кислорода, что приводит в конечном итоге к ее неполному (O2, H2O2) или полному (H2O) восстановлению. Реакции данного типа катализируются ферментами, называемыми оксидазами, и могут представлять собой свободное окисление и окисление, сопряженное с запасанием энергии. К реакциям свободного окисления относятся реакции, катализируемые растворимыми оксидазами, локализованными в цитозоле клетки. Помимо них у прокариот описан ряд связанных с мембранами оксидаз цитохромной и нецитохромной природы, перенос электронов с которых на O2 также не сопряжен с запасанием энергии.
Промежуточными по химическому механизму реакциями между приведенными выше являются реакции, в которых судьба каждого из двух атомов в молекуле кислорода различна:
A + O2 + ДH2 ® AO + H2O + Д.
В этом случае 1 атом поглощенной молекулы кислорода используется для окисления вещества путем прямого присоединения к нему. а другой восстанавливается до H2O в присутствии подходящего донора электронов. Обе реакции катализируются одним ферментом, принадлежащим к группе монооксигеназ, или оксигеназ (оксидаз) со смешанными функциями. Монооксигеназы в клетке многочисленны и разнообразны. Они катализируют реакции свободного окисления. Участие в процессах, сопряженных с запасанием клеткой энергии, маловероятно.
Таким образом, оксигеназы — это ферменты, катализирующие активирование O2 и последующее включение 1 или 2 его атомов в молекулы различных субстратов. Если субстратом (акцептором O2) служит водород, фермент называют оксидазой. В этом смысле оксидазы можно рассматривать как специализированный класс оксигеназ.
Оксигеназы играют важную роль в процессах биосинтеза, деградации и трансформации клеточных метаболитов: ароматических аминокислот, липидов, сахаров, порфиринов, витаминов. Субстратами, на которые воздействуют оксигеназы, часто служат сильно восстановленные не растворимые в воде соединения; их окисление приводит к тому, что продукты реакции становятся более растворимыми в воде и, следовательно, биологически активными, что важно для их последующего метаболизирования. У строго анаэробных прокариот кислород, включаемый в молекулу субстрата, происходит не из O2, а из других соединений, например воды.
Следовательно, всю совокупность взаимодействия молекулярного кислорода с клеткой, с точки зрения лежащих в основе этого химических механизмов, можно свести к участию O2 в двух типах реакций, в первом из которых он выступает в качестве конечного акцептора электронов, а во втором происходит его прямое внедрение в молекулу вещества. Только первый тип реакций с участием молекулярного кислорода может стать источником энергии для клетки. Поэтому для нас важно проанализировать эволюцию взаимодействия клетки с O2 по пути формирования ею систем, включающих использование молекулярного кислорода в качестве конечного акцептора электронов.
ФОРМИРОВАНИЕ "ОКСИДАЗНОГО МЕХАНИЗМА" ВЗАИМОДЕЙСТВИЯ С МОЛЕКУЛЯРНЫМ КИСЛОРОДОМ, СОПРЯЖЕННОГО С ЗАПАСАНИЕМ ЭНЕРГИИ
С появлением в атмосфере O2 возникла возможность переноса на него электронов. Чтобы этот перенос мог быть связан с получением энергии, необходимо было сформировать электронтранспортные цепи с определенным образом ориентированными в мембране переносчиками, обеспечивающими на отдельных этапах перемещение протонов через мембрану, а электронов — на O2, и ферментный комплекс, преобразующий возникающую при электронном транспорте электрохимическую энергию в химическую, запасаемую в молекулах АТФ.
Со сформированными электронтранспортными цепями, локализованными в мембране, содержащими все типы переносчиков и имеющими прямое отношение к получению клеткой энергии, мы уже встречаемся у рассмотренных в главах 13 и 14 анаэробных эубактерий с наиболее просто организованной энергетикой хемотрофного (брожение) и фототрофного (бескислородный фотосинтез) типа: некоторых пропионовокислых бактерий, всех фотосинтезирующих пурпурных и зеленых бактерий. В клеточных мембранах этих организмов локализованы и функционируют сопряженные с электронным транспортом АТФ-синтазы.
П. Митчелл высказал предположение, что система переноса электронов и протонов и переносящая протоны АТФаза возникли независимо друг от друга и, вероятно, неодновременно как разные способы генерации DmH+, необходимого для обеспечения энергией процесса избирательного транспорта питательных веществ в клетку. Последующая "встреча" обеих систем, в клетке положила начало сопряжению процессов транспорта электронов и фосфорилирования в результате обращения работы АТФазы. Это сделало возможным запасание свободной энергии окисления в молекулах АТФ. Близкий состав и аналогичная структура энергопреобразующих мембран, большое сходство механизмов сопряжения у разных групп прокариот и эукариот указывают на то, что возникшая на раннем этапе эволюции система сопряжения электронного транспорта и фосфорилирования была использована всеми организмами без принципиальных изменений.
О происхождении обратимой протонной АТФазы
Наиболее древнее происхождение имеет, вероятно, протонная АТФаза. Она обнаружена в клетках всех организмов, в том числе и у первичных анаэробов-бродильщиков, синтезирующих АТФ в реакциях субстратного фосфорилирования. Гипотетические первичные клетки получали всю энергию за счет субстратного фосфорилирования и имели слаборазвитые биосинтетические способности. Поступление необходимых органических соединений из внешней среды и выделение конечных продуктов брожения происходило по механизму пассивного унипорта (см. рис. 26). Первичные клетки, вероятно, не имели клеточной стенки, а были отграничены от окружающей срезы только элементарной мембраной. Очевидно, что активные транспортные процессы, обеспечивающие избирательный перенос веществ против их концентрационных градиентов, были необходимы на очень ранних этапах клеточной эволюции.
Для выполнения этой задачи в клетках и была сформирована локализованная в ЦПМ АТФ-зависимая протонная помпа. Энергия гидролиза АТФ, осуществляемого АТФазой, использовалась для выталкивания протонов из клетки во внешнюю среду. Гидролиз одной молекулы АТФ приводит к переносу 2 протонов и созданию таким путем трансмембранного электрохимического протонного градиента. Экспериментально это было показано для молочнокислых бактерий и клостридиев, у которых нет дыхания, но в ЦПМ локализованы АТФазы, расщепляющие молекулы АТФ, образующиеся при брожении.
Таким образом, использование АТФ для создания DmH+ на мембране — эволюционно очень древний механизм прокариотной клетки. Позднее возник механизм синтеза АТФ за счет DmH+. Для этого надо было изменить направление работы протонного АТФазного комплекса.
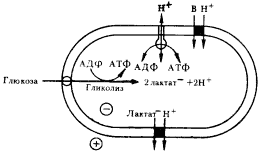
|
| Рис. 90. Схема энергетических и транспортных процессов у молочнокислых бактерий. Темный кружок — переносчик; В — молекула растворенного вещества; глюкоза поступает в клетку с помощью фосфотрансферазной системы. Остальные объяснения см. в тексте |
Обратимо функционирующие протонные АТФазы мы находим у первичных анаэробов, получающих энергию в процессе брожения. Обнаружено, что выделение во внешнюю среду молочной и уксусной кислот молочнокислыми бактериями и клостридиями приводит к созданию на ЦПМ протонного градиента. У стрептококков, осуществляющих гомоферментативное молочнокислое брожение, молочная кислота накапливается в клетке в виде аниона, для которого ЦПМ практически непроницаема. Выход лактата из клетки происходит в процессе Электронейтрального симпорта с протонами (рис. 90). Аналогичную картину наблюдают у Clostridium pasteurianum, у которого накапливающиеся в клетке ионы ацетата проходят через ЦПМ в недиссоциированной форме. Поскольку внутри клетки концентрации молочной и уксусной кислот в начале брожения всегда выше, чем во внешней среде, выход их осуществляется с помощью соответствующих переносчиков по концентрационному градиенту, т. е. в процессе облегченной диффузии, не требующей энергетических затрат. Транспорт H+ в симпорте с лактатом или ацетатом приводит к генерированию на ЦПМDmH+. При накоплении во внешней среде кислот их концентрационный градиент постепенно падает, в результате чего способность образовывать протонный градиент, связанный с выделением кислот из клетки, уменьшается. При высоких концентрациях молочной и уксусной кислот в среде образование DmH+ на мембране зависит только от гидролиза АТФ.
Электрохимическая энергия протонного градиента, возникающая при выделении из клетки кислот в процессе брожения, может использоваться для транспорта в нее растворимых веществ, а также для синтеза АТФ, который осуществляется при функционировании протонной АТФазы в обратном направлении, т. е. в АТФ-синтазной реакции. Выход энергии за счет выделения из клетки продуктов брожения может быть довольно значительным. При гомоферментативном молочнокислом брожении, по проведенным подсчетам, он может достигать 30% от общего количества энергии, вырабатываемой клеткой. Таким образом, у некоторых эубактерий, получающих энергию в процессе брожения, АТФ может синтезироваться в реакциях субстратного фосфорилирования и дополнительно за счет использования DmH+, образующегося при выходе конечных продуктов брожения в симпорте с протонами. Следовательно, эубактерий с облигатно бродильным типом энергетики уже имеют протонные АТФазы, функционирующие в направлении гидролиза и синтеза АТФ, т. е. катализирующие обратимое взаимопревращение двух видов метаболической энергии:
АТФ « DmH+
Наконец, у некоторых первично анаэробных эубактерий-бродильщиков обнаружена АТФ-синтазная активность, сопряженная с короткими фрагментами переноса электронов с помощью связанных с мембраной переносчиков (см. ниже).
Растворимые системы переноса электронов на O2 у первичных анаэробов
Как известно, перенос электронов лежит в основе всех окислительно-восстановительных процессов. В разных видах брожений, рассмотренных в гл. 13, перенос электронов (водорода) от одних органических молекул к другим обычно осуществляют растворимые НАД-зависимые дегидрогеназы:
где дг — соответствующие дегидрогеназы, содержащие НАД в качестве кофермента; А — молекула органического вещества, служащая акцептором электронов. Молекулы НАД-H2 используются в конструктивном метаболизме, обеспечивая восстановитель для биосинтетических процессов, а также в системе энергетического метаболизма, участвуя в решении "акцепторной проблемы". Электронный перенос в этом случае не приводит к получению клеткой энергии, она вырабатывается только в реакциях субстратного фосфорилирования.
У некоторых эубактерий описан прямой перенос электронов с растворимых НАД-зависимых ферментов на O2, приводящий к его восстановлению:
НАД-H2 + O2 ® НАД+ + H2O2
Окисление НАД-зависимых дегидрогеназ осуществляется также через посредство флавопротеинов, катализирующих перенос 1, 2 или 4 электронов на O2, что приводит к образованию супероксидного аниона, перекиси водорода или воды соответственно. O2 и H2O2 далее могут разлагаться ферментами, разобранными выше в этой главе.
У аэротолерантных анаэробов, таких как молочнокислые бактерии и некоторые клостридии, флавопротеины выполняют роль основного связующего звена между субстратом и молекулярным кислородом. Подобные системы могут быть полезными, например, для создания анаэробных условий в результате поглощения O2 из среды, но не имеют отношения к получению клеткой энергии. Восстановление O2, при котором в роли оксидаз, т. е. ферментов, непосредственно осуществляющих перенос электронов на молекулярный кислород, выступают флавопротеины, получило название "флавинового дыхания". В основном при флавиновом дыхании осуществляется двухэлектронный перенос на O2. Так, у молочнокислых бактерий рода Streptococcus 90% поглощенного O2 восстанавливается до H2O2.
Наконец, у некоторых эубактерий обнаружены оксидазы флавопротеиновой природы, катализирующие прямое окисление субстратов, например пировиноградной и молочной кислот, молекулярным кислородом:
Общая черта перечисленных выше путей электронного транспорта с участием одного-двух посредников на O2 — протекание реакций в цитозоле клетки, т. е. вне связи с клеточными мембранами, и отсутствие при этом запасания клеткой полезной энергии.
Принципиально важным шагом на пути создания электрон-транспортных систем, приводящим к получению клеткой энергии, явилось встраивание электронных переносчиков в мембрану.
Формирование связанных с мембраной путей переноса электронов в анаэробных условиях
В клетках первичных анаэробов обнаружены короткие пути переноса электронов, осуществляемого с помощью мембран-связанных переносчиков. В некоторых случаях такой перенос сопровождается перемещением протонов через мембрану и приводит к образованию DmH+ и синтезу АТФ. Одним из наиболее изученных путей такого типа является фумаратредуктазная система, приводящая к восстановлению фумарата до сукцината.
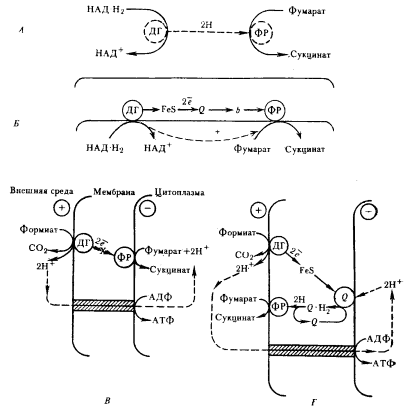
|
| Рис. 91. Функционирование фумаратредуктазной системы в растворимом (А) и связанном с мембраной (Б, В, Г) состоянии. Пунктиром обведены растворимые ферменты и изображены пути переноса H и H+ в цитозоле клетки: ДГ — дегидрогеназа; ФР — фумаратредуктаза; Q — переносчик хиноновой природы: b — цитохром; FeS — железосеросодержащий белок; X — электронный переносчик неизвестной природы |
Восстановление фумарата до сукцината может быть использовано для анаболических целей (необходимость сукцината для синтеза тетрапирролов) или же в катаболических процессах. В последнем случае все компоненты реакции могут быть растворимыми, и тогда процесс служит только для акцептирования электронов (рис. 91, 4), или же находиться в связанном с мембраной состоянии (рис. 91, Б–Г). По имеющимся данным, это не всегда приводит к синтезу АТФ. Образование протонного градиента на мембране при переносе электронов на фумарат зависит от состава и расположения электронных переносчиков.
Донорами электронов для восстановления фумарата могут служить НАД-H2, лактат, формиат или молекулярный водород, от которых электроны с помощью субстратспецифических дегидрогеназ переносятся на связанные с мембраной переносчики (рис. 91, Б). Среди переносчиков идентифицированы FeS-белки, менахинон и цитохромы типа b, однако перенос такого типа не связан с получением клеткой энергии.
Для образования протонного градиента в некоторых случаях достаточно, чтобы донор электронов и их конечный акцептор были расположены на разных сторонах мембраны. Поступление электронов на переносчик, локализованный на внешней стороне мембраны, приводит к выделению протонов в среду, а восстановление фумарата на другой стороне мембраны сопровождается их поглощением из цитоплазмы, при этом переноса протонов через мембрану не происходит. Разрядка образующегося протонного градиента с помощью АТФ-синтазы будет приводить к синтезу АТФ (рис. 91, Б). Если донор и акцептор электронов локализованы на одной стороне мембраны, тогда создание протонного градиента обеспечивается сочетанием переноса электронов и водорода по цепи, содержащей несколько переносчиков (рис. 91, Г). Перенос водорода через мембрану осуществляется с помощью хинонов. Дальнейшее усовершенствование электронного транспорта связано с включением в мембрану цитохромов.
Возможность синтеза АТФ при переносе электронов от НАД-H2, формиата, лактата, H2 на фумарат подтверждается соответствующими значениями окислительно-восстановительных потенциалов доноров и конечного акцептора электронов (см. табл. 11).
Функционирующее в системе клеточного катаболизма восстановление фумарата до сукцината обнаружено у ряда эубактерий, получающих энергию в процессе брожения. Одним из этапов на пути образования пропионовой кислоты при пропионовокислом брожении является восстановление фумарата до сукцината, катализируемое фумаратредуктазой (см. рис. 54). Фумаратредуктаза найдена также у некоторых клостридиев и молочнокислых стрептококков.
Хорошо известен связанный с мембраной фермент сукцинатдегидрогеназа катализирующий в ЦТК окисление сукцината до фумарата. Водород, акцептируемый в этой реакции флавинадениндинуклеотидом (ФАД), непосредственно поступает в дыхательную цепь (см. рис. 92). Поскольку фумаратредуктаза и сукцинатдегидрогеназа катализируют одну и ту же реакцию, но в разных направлениях, первоначально считали, что это один фермент. Сейчас показано, что реакции осуществляются разными ферментными белками. Информация о них содержится в разных генах. Синтез сукцинатдегидрогеназы индуцируется в аэробных, а фумаратредуктазы — в анаэробных условиях.
Ацетогенные клостридии оказались способными синтезировать ацетат из CO2 и H2:
2CO2 + 4H2 ® CH3-COOH + 2H2O.
Они могут расти хемолитоавтотрофно на среде, содержащей H2 в качестве единственного источника энергии. Следовательно, у этих организмов восстановление CO2 до ацетата, сопряженное в анаэробных условиях с окислением H2, должно быть связано с получением полезной энергии. Обнаружено, что в переносе электронов от H2 на CO2, ведущем к синтезу ацетата, участвуют флаводоксин, менахиноны и цитохромы типа b, т. е. переносчики того же типа, что и при функционировании фумарат-редуктазной системы.
Таким образом, у ряда первичных анаэробов, получающих энергию в процессах брожения, сформировались короткие, связанные с мембраной электронтранспортные цепи, функционирование которых ведет к образованию протонного градиента, используемого для синтеза АТФ. Из-за отсутствия подходящего конечного акцептора электронов в анаэробных условиях выход энергии в такого типа процессах низкий. Однако принципиальные основы для создания энергетики нового типа сформированы.
Для перехода к использованию энергии света необходимо было создание фоторецепторных молекул и "подключение" части из них к имеющимся электронтранспортным цепям. Такие фоторецепторы — Mg-порфирины — были сформированы. Фотосинтез начался, видимо, с создания системы фотоиндуцированного циклического электронного транспорта и служил сначала в качестве источника энергии, дополнительного к основному, которым являлись процессы брожения. Восстановитель первичные фотосинтезирующие организмы могли получать теми же путями, что и бродильщики, или же тратя для этого часть синтезированного АТФ в процессе обратного переноса электронов. Позднее для этой цели была сформирована способность прямого фотовосстановления НАД+, приведшая к созданию светозависимого нециклического электронного транспорта. Дальнейшее усовершенствование фотосинтетического аппарата привело к использованию воды в качестве донора электронов, побочным продуктом чего явилось образование молекулярного кислорода.
В результате фотосинтетического выделения O2 появилось химическое соединение, служащее активным окислителем. В ответ на появление O2 большинством прокариот были выработаны различные механизмы защиты. У некоторых линий приспособление к O2 на этом закончилось, в результате возникли анаэробные формы с разной степенью аэротолерантности.
Следующий важный шаг в формировании механизма использования молекулярного кислорода в качестве конечного акцептора электронов — использование этого процесса, таящего в себе большие энергетические возможности, для получения клеткой энергии. Действительно, количество энергии, освобождающейся при переносе пары электронов, зависит как от природы донора, так и от природы акцептора электронов. Hапример, окислительно-восстановительный потенциал НАД-H2 равен – 320 мВ, а молекулярного кислорода — +810 мВ. Для образования 1 молекулы АТФ необходим перенос пары электронов по электрохимическому градиенту, соответствующему разнице потенциалов приблизительно 200 мВ.
Для использования O2 в качестве конечного акцептора электронов в процессах, связанных с получением метаболической энергии представлялось наименее сложным превратить фотосинтетический электронный транспорт в дыхательный. С этой целью надо было добавить дегидрогеназы на низкопотенциальный конец цепи и цитохромоксидазы — на другой, взаимодействующий непосредственно с O2. Все необходимые типы переносчиков и обратимые протонные АТФазы уже были к этому времени сформированы.
Основная задача сводилась к созданию ферментной системы для четырехэлектронного восстановления O2 (цитохромоксидазы), при котором не освобождалось бы его токсических промежуточных продуктов.
Фосфорилирование, сопряженное с переносом электронов от субстратов в темновых окислительных реакциях, получило название окислительного фосфорилирования. Развитие механизма окислительного фосфорилирования позволило добиться наиболее полного извлечения свободной энергии из окисляемых субстратов.
Таким образом, появление молекулярного кислорода положило начало эволюции новых типов жизни в мире прокариот, в основе которых лежит получение энергии за счет процессов окислительного фосфорилирования.
М.В.Гусев, Л.А.Минеева © 1992-2001
Кафедра клеточной физиологии и иммунологии биологического факультета МГУ им. М.В.Ломоносова © 2000-2001
Страница обновлена 09.09.01
Оформление: А.В.Киташов