| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 7
Расселение и поток генов
Популяция может приобрести новый аллель либо в результате мутации, возникшей у одного из её членов, либо в результате иммиграции носителя этого аллеля из другой популяции. Последний процесс создаёт поток генов.
Носитель нового аллеля, очевидно, приобрел его в результате мутации, произошедшей когда-то в прошлом, и, таким образом, поток генов можно рассматривать как запаздывающий эффект мутационного процесса. Тем не менее прямым и непосредственным источником некоторых изменений в популяции в каждый данный момент может быть иммиграция носителей различных аллелей из других популяций. Некоторые мутантные аллели, никогда не возникая в данной популяции, могут поступать в нее из соседних популяций. Таким образом, в конечном итоге источником изменчивости в популяции-реципиенте служат генные мутации, возникшие в какой-то момент в прошлом в каком-то другом месте; однако действенной причиной изменчивости в данное время и в данном месте является поток генов.
Поток генов зависит в свою очередь ещё от одного процесса — расселения, составляющего его физическую основу.
Какова эффективность потока генов в пространстве и времени? Сколь далеко может распространиться определённый ген в популяционной системе за некий разумный период, скажем за 1000 лет. При данном потенциале расселения и обычной интенсивности миграции с помощью или без помощи отбора? Этот основной вопрос будет обсуждён в данной главе, но окончательного ответа на него дано не будет.
Определения
Соотношение между названными выше процессами разъясняют следующие определения.
Расселение — это перемещение в пространстве особей, их спор, гамет или особых органов расселения. Оно может происходить как по территории, занимаемой данным видом, так и по территории, им не занимаемой.
Иммиграция — это особый случай расселения, при котором происходит перемещение из одной части популяционной системы в другую её часть. Короче говоря, иммиграция — это расселение в пределах территории, занимаемой данным видом.
Поток генов — это их перемещение внутри популяции или между популяциями. Этот процесс включает иммиграцию и скрещивание между иммигрантами и членами местной популяции, в результате которых в генофонд-реципиент вводятся инородные гены.
В одних случаях поток генов может привести к изменениям частот генов в популяции-реципиенте, тогда как в других случаях таких изменений не происходит. Желательно обозначить эти два случая разными терминами (Grant, 1980*), так как отсутствие для них специальных терминов привело к некоторой путанице в литературе. Грант (Grant, 1980*) предлагает называть поток генов между популяциями с одинаковыми частотами генов изогенным потоком генов, а поток генов между популяциями, различающимися по частоте генов, — генетически эффективным потоком генов.
Расселение
Единицами расселения у подвижных высших животных являются отдельные организмы, у сидячих морских животных — свободноплавающие личинки, у высших растений — пыльца и семена, а у низших растений и грибов— споры. Расселение представляет собой одну из нормальных стадий жизненного цикла. У всех организмов, в том числе и у сидячих, в жизненном цикле имеется, по-видимому, свободноживущая подвижная стадия, при помощи которой происходит расселение. Но при этом огромное большинство расселяющихся единиц, в том числе и отдельные особи подвижных животных, обычно остаются поблизости от места своего происхождения или от родной территории. Для них характерна также некоторая степень оседлости. Совместное действие этих двух факторов — расселения и оседлости — определяет пространственное распределение особей и их генов из поколения в поколение.
В разных случаях для измерения расселения используют разные единицы. Наиболее полезными представляются данные о распределении частот расстояний расселения (т. е. расстояний, на которые удалились взрослые половозрелые особи от места своего рождения, вылупления или местонахождения семени) для адекватной выборки особей. Такие данные были получены для техасской заборной игуаны (Sceloporus olivaceus); на молодых особей наносили метку, а затем вторично отлавливали их во взрослом состоянии (Blair, 1960*). Выпуская на волю помеченных генетическими маркерами особей Drosophila pseudoobscura и вылавливая их потомков спустя много месяцев, удалось установить расстояние, на которое расселяются мухи за несколько поколений (Dobzhansky, Wright, 1947*). Во многих исследованиях по практическим соображениям приходится вылавливать помеченных особей спустя короткое время после того, как они были выпущены, с тем чтобы определить расстояния, на которые они расселяются за какой-то период активного времени жизни. Так, Поуэлл и др. (Powell et al., 1967*), изучая группу Drosophila pseudoobscura, вылавливали меченых мух через 1 — 2 дня после того, как они были выпущены.
Яблоков и его коллеги предлагают три практические единицы измерения расселения, которые они использовали в своих исследованиях на ящерицах и других животных. Первая из них — так называемый радиус индивидуальной активности (Тимофеев-Ресовский и др., 1977*); это расстояние, покрываемое одной особью за день или за сезон. Вторая единица — средний радиус индивидуальной активности (Яблоков и др., 1980*) — среднеарифметическое расстояние между местом, где было выпущено животное, и местом, где оно было вновь поймано по прошествии определённого периода времени. Третья единица — это радиус репродуктивной активности, т. е. расстояние между местом рождения и местам размножения для 95% особей в любом данном поколении (Яблоков, 1986*).
У семенных растений существует две стадии расселения — расселение пыльцы и расселение семян, причём эти две стадии независимы одна от другой. Поэтому общее расстояние, на которое происходит расселение, в расчете на поколение определяется либо как разность между расстояниями, достигнутыми на этих двух стадиях, либо как их сумма.
Типы расселения
Данные о расстояниях, на которые расселяются взрослые особи Drosophila pseudoobscura и близкие к ним виды, приведены в табл. 7.1 (Powell et al., 1976*). Мух метили флуоресцентным красителем, затем выпускали и спустя 1 — 2 дня вылавливали по трём трансектам через определённые расстояния. В табл. 7.1 объединены данные по трём трансектам. Высокая частота расселения на короткие расстояния очевидна; довольно много также случаев расселения на средние расстояния, но всего несколько случаев расселения на большие расстояния.
Сравнимые данные по прыткой ящерице (Lacerta agilis) в горах Алтая приведены в табл. 7.2 (Яблоков и др., 1980*); это данные о расселении за один год. Большая часть популяции оставалась вблизи места, где животные были выпущены, но небольшая её доля расселилась довольно далеко во все стороны. Средний радиус индивидуальной активности для данной популяции составляет 30.6 м на одну особь за один год.
выловленных вторично по трём трансектам на разных расстояниях от места, где они были выпущены (по данным табл. 3 в работе Powell et al., 1976*) | |||
ста выпуска, м | Число меченных мух | ста выпуска, м | Число меченных мух |
| В месте выпуска | 123 | 440 | 19 |
| 40 | 115 | 480 | 11 |
| 80 | 116 | 520 | 9 |
| 120 | 91 | 560 | 2 |
| 160 | 67 | 600 | 4 |
| 200 | 65 | 640 | 3 |
| 240 | 67 | 680 | 11 |
| 280 | 55 | 720 | 6 |
| 320 | 45 | 760 | 6 |
| 360 | 29 | 800 | 2 |
| 400 | 35 | ||
В другой работе по расселению ящерицы, проводившейся в Техасе на протяжении 6 лет (Blair, 1960*), велись систематические наблюдения за техасской заборной игуаной (Sceloporus olivaceus). В этом случае определяли расстояние, на которое животные расселялись за всю свою жизнь — от вылупления до достижения половой зрелости. Характер расселения представлен в виде диаграммы на рис. 7.1. Как можно убедиться, большинство особей остается в радиусе 60 — 90 м от того места, где они вылупились, но некоторые отходят далеко, вплоть до 500 м от места вылупления.
Яблоков и др. (1980)* приводят следующие данные о среднем радиусе индивидуальной активности для особей разного пола и возраста в алтайской популяции прыткой ящерицы:
| Молодые особи | 40.6 м |
| Половозрелые самцы | 34.8 м |
| Половозрелые самки | 25.0 м |
Эти результаты типичны. Более высокая расселяемость самцов по сравнению с самками обнаружена также у техасской заборной игуаны (рис. 7.1), в группе Drosophila pseudoobscura (Powell et al., 1976*) и у различных млекопитающих. У мух и у млекопитающих наблюдалась также более высокая способность к расселению молодых особей по сравнению с половозрелыми.
на которые расселялась прыткая ящерица (Lacerta agilis) (Яблоков и др., 1980*) | ||
| Расстояние, м | Частота, % | |
| все ящерицы | самцы | |
| 0—5 | 60.1 | 61.2 |
| 6—10 | 25.9 | 19.7 |
| 11—30 | 10.0 | 13.7 |
| 31—60 | 2.5 | 3.3 |
| 61—100 | 1.0 | 1.1 |
| 101—200 | 0.4 | 0.7 |
| 201—400 | 0.07 | 0.2 |
| 401—700 | 0.03 | 0.1 |
Судя по некоторым данным, у полевок (Microtus) различия в поведении между особями одного пола и возраста, имеющими большой радиус активности и ведущими оседлый образ жизни, обусловлены генетически (Krebs et al., 1973*).
Колуэлл (Colwell, 1951*) провел экспериментальное исследование особенностей распространения пыльцы у сосны. Пыльцу сосны Культера (Pinus coulteri) метили радиоактивным фосфором и после её рассеяния ветром выявляли при помощи счетчика Гейгера — Мюллера и методом радиоавтографии. Партии пыльцы объемом 1 л помещали в сосуды, которые подвешивали кверху дном на высоте 3.5 м. В ясный день при слабом ветре пыльца медленно высыпалась из сосуда. По радиусам, отходящим в разных направлениях от точки распространения пыльцы, ставились с равномерными интервалами ловушки для пыльцы. Количество пыльцы, попавшей в разные ловушки, определяли по уровню радиоактивности.
Основная масса пыльцы распространялась в направлении ветра на расстояния от 3 до 9 м. За пределами этой зоны максимальной концентрации наблюдалось резкое снижение количества рассеянной пыльцы. На расстоянии 45 м и более от сосуда можно было обнаружить лишь небольшие количества пыльцы. Эти результаты означают, что в сосновом лесу женская шишка будет буквально засыпана пыльцой с соседних деревьев и получит некоторое, но не очень большое количество пыльцы от деревьев, находящихся на расстоянии нескольких десятков метров (Colwell, 1951*).
Как оказалось, расселение описанного выше типа широко распространено в природе. Мы привели характерные примеры; аналогичные примеры можно встретить у грызунов, птиц, медоносных пчел, бабочек, энтомофильных и орнитофильных растений и у семян, распространяемых ветром. В общем кривая частотного распределения расстояний, на которые происходит расселение, имеет максимум вблизи места появления организмов на свет и снижается по мере удаления от этого места. Большая часть особей остается вблизи своих «родных» мест, а некоторые расселяются на умеренно далекие расстояния.
Ожидаемый результат расселения такого типа состоит в том, что у свободно скрещивающихся организмов интербридинг происходит главным образом между ближайшими соседями. В ряду поколений это приведет к значительному количеству близкородственных скрещиваний несмотря на систему свободного скрещивания. Тесный инбридинг дополняется, однако, некоторым количестном отдаленных неродственных скрещиваний с иммигрантами — отдельными особями или гаметами. Эти скрещивания обеспечивают небольшой, но, вероятно, значимый обмен генами между особями данной популяции, находящимися на больших расстояниях друг от друга.
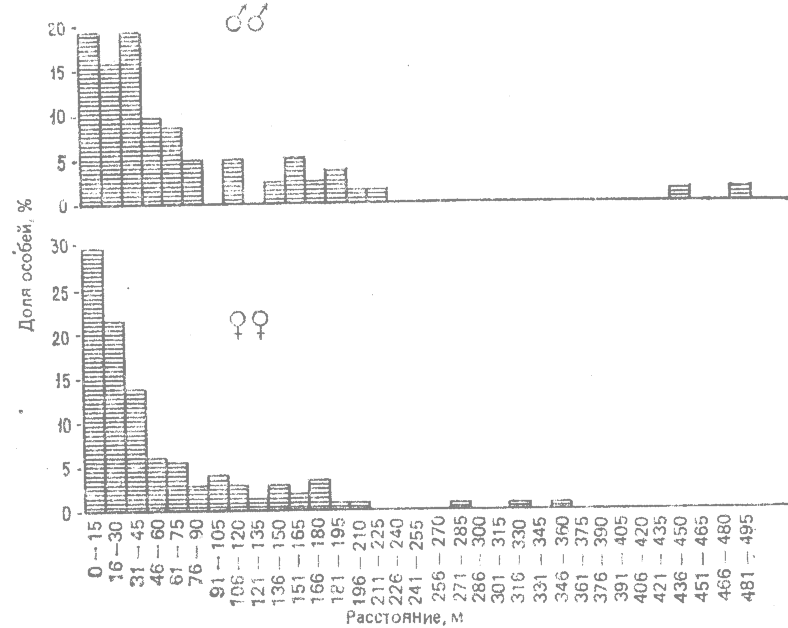 |
Рис. 7.1. Относительное количество самцоц и самок игуаны Sceloporus olivaceus, расселяющихся на разные расстояния от места вылулления до местонахождения половозрелых животных (Blair, 1960.*) |
Области расселения
Хотя частотное распределение расстояний, на которые происходит расселение, примерно одинаково для разных групп организмов, средние расстояния различны. Различия между главны ми группами по типичной области расселения очень велики. Эти различия представлены в табл. 7.3. Приведенные в ней средние радиусы расселения для большинства организмов относятся к одному поколению, а для дрозофилы — к некоторой части одного поколения. Эти данные соответствуют ожидаемым. Две противоположные крайние точки занимают высокоподвижные плодовые мухи и сидячие растения, различия между которыми достигает двух порядков величин. Перемещающиеся по земле ящерицы значительно менее непоседливы, чем крылатые мухи. Что касается улиток, то они расселяются ненамного шире, чем растения.
у разных организмов. | ||
| Организм | Среднее расстояние, м | Источник данных* |
| Двукрылые | ||
pseudoobscura (Калифорния) | 219, 631 в разных популяциях | Dobzhansky, Powell (1974); Powell et al., 1976 |
scura (Колорадо) | 176, 202 в разных популяциях | Crumpacer, Williams (1973) |
| Ящерицы | ||
| 49, 71 у самцов и самок | Blair (1960) | |
| 31 (средний радиус активности) | Яблоков и др. (1980) | |
| Улитка | ||
| Cepaea nemoralis | 8.1 | Lamotte (1951) |
| Травянистые растения | ||
| 6.3 (семянки) | Poole, Cairns (1949) | |
| 2.5 (семена) | Levin, Kerster (1969) | |
| 1.1 (семена) | Levin, Kerster (1968) | |
Интенсивность иммиграции
Интенсивность иммиграции (m) варьирует количественно от О до 1, где m=0 означает полное отсутствие иммигрирующих аллелей, а m=1 означает «затопление» популяции иммигрантами за время жизни одного поколения. В общем виде m равно отношению числа иммигрантов за одно поколение к числу исконных членов популяции плюс число иммигрантов, прибывших за данное поколение; или m равно отношению числа гамет, иммигрирующих за одно поколение, к общему числу гамет.
nigrospiracuta (по данным табл. 3 в работе Johnston, Heed, 1976*, и по личному сообщению Johnston) | |
ходит расселение, м | Интенсивность иммиграции, m |
| Место выпуска | 0.517 |
| 19—30 | 0.168 |
| 50—60 | 0.084 |
| 90—100 | 0.039 |
| 128—158 | 0.041 |
| 234—274 | 0.028 |
| 373—408 | 0.029 |
| 443—512 | 0.036 |
| 638—768 | 0.052 |
| 871—932 | 0.075 |
Одно из лучших количественных исследований интенсивности иммиграции провели Джонстон и Хид (Johnston, Heed, 1976*) на Microtus в Аризоне, Они представили данные о проценте меченых мух в выборках, взятых на различных рас стояниях от места выпуска. В табл. 7.4 приведены значения m, вычисленные по этим данным. Эти значения представляют собой взвешенные средние для выборок, взятых на каждом расстоянии в разные дни; вычисления проводил Джонстон (личное сообщение).
Общая средняя интенсивность иммиграции у этих мух m= 0.037.
Значение m, однако, не постоянно, а уменьшается с расстоянием примерно так же, как расстояния, на которые происходит расселение, в других использованных нами работах. Небольшое преобладание мух-мигрантов в отдаленных точках объясняется неравномерным распределением их природных источников пищи (Johnston, Heed, 1976*),
Поток генов
Поток генов составляет генетическую часть миграции. Миграция — это фактор, воздействующий на изменчивость популяции в том и только в том случае, если иммигранты генетически отличны от членов нативной популяции. Потока генов следует ожидать в случае миграции между популяциями или субпопуляциями, различающимися по частоте аллелей. Иммиграция в таком случае приведет к тому, что мы назвали генетически эффективным потоком генов (в отличие от изогенного потока генов).
Между интенсивностью миграции и изменением частоты аллелей существует определённая зависимость (Falconer, 1960; 1981*). Обозначим частоту данного аллеля среди исконных членов популяции через q0, среди иммигрантов — через qm, а в получающейся в результате смешанной популяции — через q. В та ком случае изменение частоты этого аллеля за одно поколение (Δq), обусловленное потоком генов, составит
Δq = m(qm—q0).
Новая частота аллеля в нативной популяции после иммиграции в одном поколении будет равна
q = m(qm—q0)+q0.
Например, если частота данного аллеля среди иммигрантов qm=0.5, среди исконных членов популяции q0=0.3, а скорость иммиграции m=0.001, то изменение частоты аллеля после одного поколения иммиграции Δq=0.002, а новая частота аллеля q=0.302.
В приведенной выше формуле принимается во внимание только m, т.е. доля популяции, мигрирующая в каждом поколении, а численность популяции N не учитывается. В своих работах по моделированию потока генов Слэткин (Slatkin, 1980; 1981a*, личное сообщение) обнаружил, что эффективное количество потока генов лучше измерять с учетом Nm, чем только m.
Вообще скорость изменения частоты гена в популяции, обусловленная потоком генов, зависит от двух факторов: 1) от интенсивности иммиграции, в каких бы величинах её не выражали, и 2) от различий в частоте данного аллеля между популяцией-донором и популяцией (или субпопуляцией) — реципиентом (Falconer, 1960; 1981*).
Расселение во времени
Исследования расселения и потока генов, как эмпирические, так и теоретические, касаются в большинстве случаев изменений, происходящих на протяжении жизни одного поколения. Если, однако, мы ставим себе целью понять динамику природных популяций, то совершенно ясно, что необходимо рассматривать эти процессы в длинном ряду поколений. В этом и следующем разделах сделана попытка ввести в обсуждение фактор времени (см. также Grant, 1980; 1985*).
У организма, который расселяется во всех направлениях, во 2-м поколении некоторые особи будут мигрировать дальше в том же направлении, что и часть особей в 1-м поколении. В последующих поколениях также некоторая доля миграций будет расширять ареал рассматриваемого организма в том же самом направлении. Умножив среднее расстояние расселения за одно поколение на число поколений, можно получить разумную численную оценку общего линейного расстояния, которое может покрыть данный организм путем постепенной миграции в биологическом времени. Такие оценки, полученные для различных орга низмов, можно привести к одной общей сравнительной шкале хронологического времени, вводя время генерации для каждого организма.
в результате постепенных миграций с течением времени (объяснения см. в тексте) | |||
| Вид | стояние, покры- ваемое за одно поколение | Число поко- лений в год | селения за 1000 лет (экс- траполяция) |
| Drosophila pseudoobscura и родственные формы | 176; 361 | 7 | 1234; 2527 |
| Scelopopus olivaceus | 49; 71 | 1 | 49; 71 |
| Cepaea nemoralis | 8.1 | 1 | 8.1 |
| Senecio jacobaea | 6.3 | 0.5 | 3.2 |
| Liatris aspera | 2.5; 3.5 | 0.2 | 0.5; 0.7 |
| Phlox pilosa | 1.1; 2.4 | 0.3 | 0.3; 0.7 |
Виды, фигурировавшие в табл. 7.3, вновь представлены в табл. 7.5 с указанием времени генерации. Для четырёх видов использованы средние расстояния расселения. Для Drosophila и Sceloporus они взяты непосредственно из табл. 7.3; для Liatris и Phlox меньшие значения относятся к расселению семян, а большие соответствуют сумме расстояний, на которые расселяются семена и пыльца. Последний столбец содержит оценки кумулятивного расстояния, на которое каждый вид мог бы расселиться или мигрировать за 1000 лет в результате ряда последовательных средних расселений в одном и том же направлении.
Мы видим, что высокоподвижная Drosophila pseudoobscura за 1000 лет может расселиться или мигрировать на 1200 — 2500км. Большее из этих расстояний почти равно длине или ширине её видового ареала в западной части Северной Америки. Растения с сидячим образом жизни, напротив, за 1000—10000 лет расселяются или мигрируют всего на несколько километров. Это расстояние составляет лишь незначительную долю их видового ареала. Ящерица и лягушка по способности к расселению занимают промежуточное положение.
Поток генов во времени
Нас здесь интересует генетически эффективный поток генов, происходящий на протяжении ряда поколений, в который вовлечены три или большее число популяций или субпопуляций. Мы будем применять формулу, приведенную ранее для простого случая, к несколько более сложному случаю с участием трёх поколений и четырёх популяций.
Допустим, что существуют четыре полуизолированные популяции (А, В, С и D), распространенные по трансекте, тянущейся с востока на запад. Популяция А содержит новый аллель G2, частота которого в поколении 0 равна 1.0; остальные популяции содержат прежний аллель (G1) с исходной частотой 100%. Аллель G2 в селективном отношении не лучше, но и не хуже аллеля G1. Между соседними популяциями имеет место миграция в обоих направлениях со скоростью m=0.1. Спустя три поколения частота нового аллеля G2 в четырёх популяциях составит:
популяция A q = 0.755
популяция В q = 0.219
популяция С q = 0.025
популяция D q = 0.001
Совершенно очевидно, что частота нового аллеля на каждом этапе его миграционного пути резко уменьшается. И это несмотря на то, что в нашем примере исходное различие по частотам аллеля между популяцией А и другими популяциями очень велико, в сущности максимально, и скорость миграции относительно высока. В некотором ограниченном ряду генераций (большем, чем в этом примере) на одном из этапов миграции новый мигрирующий аллель G2 окажется таким редким, что его шансы попасть в следующую выборку эмигрантов будут очень незначительны. Процесс генетически эффективного потока генов временно прекратится.
На протяжении длинного ряда поколений при продолжающемся потоке генов частоты аллелей во всех четырёх популяциях будут приближаться к равновесным, но на это потребуется много времени.
В предыдущем разделе мы пришли к выводу, что расстояния, на которые происходит расселение, с увеличением числа поколений приобретают существенную дополнительную компоненту. По расстояниям, на которые расселяется популяция за одно поколение, можно путем экстраполяции оценить расстояние, на которое она сможет расселиться с течением времени (табл. 7.5). Теперь же мы видим, что миграция нового аллеля в пространстве и во времени происходит по-разному. В каждом поколении новым аллелем обладает обычно лишь некоторая доля эмигрантов, причём величина этой доли в каждом последующем поколении уменьшается. Генетически эффективный поток генов в той мере, в какой он определяется одной лишь скоростью миграции, ограничен в пространстве гораздо сильнее, чем поэтапная миграция. Генетически эффективный поток генов довольно значительно буксует по сравнению с процессом расселения (см, также Grant, 1980*).
Попытаемся рассмотреть эти выводы применительно к проблеме миграции генов в обширной популяционной системе. Допустим, что эта система протянулась на 1000 км и что отрезок времени составляет 1000 лет. Может ли отдельный ген, не имеющий селективного преимущества, распространиться в этой системе за указанный срок?
Если этот ген принадлежит растению, т. е. сидячему организму, или малоподвижному животному, например улитке, то ответ, очевидно, должен быть отрицательным; скорость их расселения слишком мала, как это видно из табл. 7.5. Высокоподвижное быстро размножающееся животное, подобное дрозофиле, может легко расселиться за предоставленное время на 1000-километровое расстояние путем поэтапной миграции (табл. 7.5). Её способность к расселению вполне соответствует поставленной задаче. Однако мы не вправе допустить, что генетически эффективный поток генов, составляющий всего лишь часть потенциала расселения, соответствует этой же задаче у того же самого организма.
До сих пор мы считали мигрирующий аллель G2 нейтральным в отношении отбора. Изменим это допущение и придадим ему селективное преимущество перед обычным(и) и широко распространенным(и) в популяционной системе аллелем(ями). Это создаёт комбинацию сил — поток генов и отбор, которые способствуют распространению аллелей. Однако миграция аллеля G2 всё ещё будет протекать медленно, так как отбор требует времени. Поскольку аллель G2 при проникновении в новую популяцию обладает низкой начальной частотой, понадобится отбор на протяжении многих поколений, для того чтобы его частота повысилась до уровня, обеспечивающего его передачу следующей популяции, и, однажды возникнув, этот процесс должен повторяться вновь и вновь. В случае ступенчатого потока генов под контролем объединенных сил миграции и отбора нам следует принимать во внимание отбор, происходящий на каждой ступени миграции в течение многих поколений.
Здесь эта проблема рассмотрена в качественном аспекте на предварительном уровне. Рассмотрение её в количественном аспекте очень желательно, однако сопряжено с трудностями, так как ступенчатый поток генов представляет собой стохастический процесс или же, что ещё больше осложняет дело, — ряд стохастических процессов, каждый из которых может привести к весьма разнообразным результатам (Slatkin, личное сообщение).
Быть может, мы занижаем оценки расселения?
Количественные данные о расселении, примеры которых были представлены выше, создают основу для наших экстраполяции относительно потока генов в пространстве и во времени. Нельзя исключать возможности того, что в имеющихся данных о расселении существует неопределённая тенденция к преобладанию коротких и средних расстояний.
Количественные исследования расселения вынужденно проводятся на ограниченных опытных площадках. Особи, споры или другие единицы расселения, находящиеся за пределами изучаемой территории, не отлавливаются и не заносятся в таблицы. Кроме того, количественное изучение расселения проводится при «нормальной» погоде; а между тем многие организмы могут расселяться на большие расстояния в периоды сильных ветров или наводнений.
Представление о возможности расселения на далекие расстояния возникло на основании колонизации океанических островов и существования разорванных ареалов некоторых типов. Оно подтверждается данными о переносе пыльцы и насекомых через океаны с помощью ветра и обнаружением отдельных представителей некоторых видов птиц за тысячи километров от их обычных ареалов. Такое расселение можно наблюдать в некоторых местах во время сильных ветров. По данным Савила (Savile, 1972*, личное сообщение), один шторм в Арктике может унести семена, плоды и некоторые вегетативные диаспоры за сотни и даже за 2000 километров.
Койн и др. (Coyne et al., 1982*) изучали расселение Drosophila pseudoobscura на обширной территории в гористой местности в пустыне, выпуская и вновь отлавливая мух; оказалось, что некоторые особи проделывают путь до 15 км.
Таким образом, мы вправе считать, что имеющиеся количественные оценки расселения слишком занижены. Кроме того, данные другого рода, а именно — существование в некоторых группах отрицательной корреляции между подвижностью и географическим расообразованием, указывают на то, что поток генов действительно эффективен на достаточно обширных территориях. Соответствующие данные представлены в гл. 23.
Заключение
Играет ли поток генов существенную роль в природных популяциях в более широком масштабе, чем локальный? Может ли генетически эффективный поток генов распространить вариации по обширной популяционной системе? В настоящее время по этому вопросу существуют две противоположные точки зрения.
Maйр (Mayr, 1954; 1963; 1970*) — главный сторонник представления о том, что масштабы и скорость потока генов достаточны для того, чтобы гомогенизировать обширные популяциониые системы. Противоположная точка зрения, которой придерживаются Эрлих и Равен (Ehrlich, Raven, 1969*) и другие авторы (Levin, 1979; 1981; Ehrlich, White, 1980*), состоит в том, что поток генов слишком ограничен, чтобы оказывать сколько-нибудь существенное воздействие за пределами локальной популяции.
Первый взгляд основан на морфологическом единообразии больших популяционных систем широко распространенных видов; второй — на количественных исследованиях расселения. Ни тот, ни другой не подкреплены убедительными данными. Морфологическое единообразие может быть обусловлено происхождением от общего предка, так же как и потоком генов; что же касается количественных исследований расселения, то они в большинстве случаев ограничиваются, изучением изменений, происходящих в одном поколении.
Анализ, проведенный в этой главе и в других работах (Grant, 1980; 1985; Slatkin, 1981b; 1985*), позволяет, казалось бы, считать, что такие высокоподвижные животные, как дрозофила, соответствуют точке зрения Майра; сидячие растения — точке зрения Эрлиха и Равена; а организмы других типов занимают промежуточное положение. Однако ввиду неясности количественного аспекта имеющихся данных этот анализ неубедителен. Если генетически эффективный поток генов во времени, сопровождаемый или не сопровождаемый отбором, составляет слишком малую долю расселения, популяции подвижных животных будут соответствовать модели Эрлиха и Равена. Если же расстояния, на которые происходит расселение, существенно больше принятых в настоящее время оценок, то для популяций сидячих растений скорее подходила бы модель Майра.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |