| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 18
Фенотипические модификации
Что такое приобретённый признак?
Теория наследования приобретённых признаков пользовалась всеобщим признанием на протяжении большей части истории биологии — начиная от древних греков и до XIX в. (Zirkle, 1946*). Её принимали и Ламарк, и Дарвин. Ошибочность этой доктрины продемонстрировал Вейсман в конце XIX в. и представители классической генетики — в начале XX в. Сторонники традиционной точки зрения, однако, не признали своего поражения. Эта концепция продолжала оказывать влияние на многие области науки до середины XX в. Дискуссии по этому вопросу, начавшись с появлением ключевых работ Вейсмана (Weismann, 1889—1892; 1892*), не прекращались до недавнего времени.
Одним из факторов, способствовавших продолжению дискуссий, была двойственность, присущая традиционной концепций приобретённых признаков. Некоторые приобретённые признаки действительно передаются по наследству (например, наследственные инфекционные заболевания), тогда как другие приобретённые признаки, называемые теперь фенотипическими модификациями, не наследуются.
Решение проблемы, предложенное Вейсманом (1889—1892; 1892*), было в то время большим шагом вперед. Он начал с того, что разделил организм на сому и зародышевую плазму, считая последнюю единственным носителем наследственности. Вейсман ввел категорию так называемых соматогенных вариаций. Это реакции соматических тканей или организма в целом на внешние воздействия; к числу таких реакций он относил различные повреждения и влияние упражнения или неупражнения. Вейсман считал, что коль скоро сома находится вне русла наследственности, то и соматогенные вариации не имеют отношения к наследственности. В подтверждение этого он приводил результаты наблюдений и экспериментов.
Часто указывают, что чёткое морфологическое разделение зародышевой плазмы и сомы далеко не универсально. Подобное разделение действительно имеет место в разных группах беспозвоночных животных, но отсутствует в других царствах (см. Buss, 1983*). Так, у растений из одной точки роста развиваются как вегетативные, так и репродуктивные органы. Подобные несообразности вызывали неудовлетворенность теорией зародышевой плазмы.
Вейсман (1892*) понимал это затруднение и пытался преодолеть его. Он указывал также, что представление о ненаследуемости соматогенных вариаций, по-видимому, справедливо применительно ко всем группам организмов, включая растения (Weismann, 1889—1892*). В этом он был не совсем прав.
В ранний период развития генетики вейсмановское разделение на зародышевую плазму и сому уступило место разделению на генотип и фенотип (Johannsen, 1911*). Первое разграничение — чисто морфологическое и не имеет всеобщего применения, а второе — генетическое и применимо ко всему живому. Разделение на генотип и фенотип позволяет прояснить проблему наследственности. Признаки, детерминируемые недавно приобретённым генетическим материалом соматических тканей, могут быть наследственными, если эти ткани затем участвуют в создании репродуктивных клеток, что вполне возможно у растений. Что же касается фенотипических признаков, то они никогда не наследуются, а поэтому фенотитипческие модификации также не передаются по наследству.
Басс (Buss, 1983*) недавно привлек внимание к одному предполагаемому изъяну в синтетической теории эволюции. Он рассуждает следующим образом. Синтетическая теория сосредоточена на генетических различиях между особями и пренебрегает генетической изменчивостью соматических тканей в пределах отдельных особей, полагая, что такие изменения никогда не наследуются. Это допущение в свою очередь основано на теории зародышевой плазмы Вейсмана, которая неприменима ко многим крупным группам. Синтетическая теория нуждается в изменениях, с тем чтобы в ней учитывалась возможность наследования некоторых типов соматической изменчивости (Buss, 1983*).
Ошибка Басса проистекает из принятой им предпосылки, что синтетическая теория опирается на вейсмановские представления о наследственности, тогда как она основывается на разделении генотипа и фенотипа.
Итак, следует различать приобретённые признаки двух типов: 1) фенотипические признаки, определяемые вновь приобретённым генетическим материалом, и 2) признаки, возникающие в результате фенотипических реакций на внешние факторы, т. е., иными словами, фенотипические модификации. Первые могут передаваться по наследству, а вторые не наследуются. Эти два типа приобретённых признаков играют совершенно разные роли в эволюции.
Мы рассмотрим их по отдельности. Фенотипические модификации обсуждаются в этой главе, а эффекты приобретённого генетического материала — в гл. 19.
Фенотипическая пластичность
Генотип — это совокупность всех генов данного организма, а фенотип — совокупность его признаков и свойств. От генотипа к фенотипу ведут длительные и сложные процессы действия генов и развития. Эти процессы протекают в определённой среде и находятся под её влиянием. Фенотипическое проявление признака представляет собой, таким образом, результат действия двух наборов факторов: генотипических детерминант и условий среды, в которой происходит развитие.
Любой генотип способен дать начало определённому спектру фенотипов, каждый из которых реализуется в определённых условиях среды. В этом смысле генотип можно представить себе как «норму реакции» (Johannsen, 1911*). Иными словами, действие генотипа нельзя считать ни жестко предопределённым, ни неограниченным: генотип может детерминировать ряд фенотипических признаков в пределах, установленных самим генотипом (Johannsen, 1911*).
Диапазон фенотипической изменчивости длины метелки и сроков цветения в различных средах был измерен в разных «семьях» однолетнего растения — костра мягкого (Bromus mollis) (Jain, 1978*). Между «семьями» были обнаружены большие различия по степени фенотипической пластичности. Некоторые генотипы порождают крайне разнообразные фенотипы; другие генотипы дают фенотипы с более узким диапазоном разнообразия.
Широкий спектр фенотипических проявлений легко продемонстрировать у растений, способных к вегетативному размножению. Для этого достаточно взять одно растение, разделить его на части и размножать эти части при различных условиях среды в фитотроне или в вегетационном домике.
Хорошим примером служит экспериментальное изучение реакций на внешние условия у сложноцветных группы Achillea millefolium (Clausen, Keck, Hiesey, 1948*), многолетних травянистых растений, которые можно размножать вегетативным путем. В Пасадене (шт. Калифорния) части одного и того же растения выращивали в разных камерах фитотрона, а также в открытом грунте зимой. Регулируемые среды различались по продолжительности светового периода, дневным и ночным температурам.
В типичном эксперименте шесть частей одного растения Achillea borealis из Сьюарда (Аляска) выращивали в течение 3.5 мес в пяти регулируемых средах в фитотроне и в шестой среде — открытом грунте. Различные ростовые реакции, наблюдавшиеся к концу экспериментального периода, изображены на рис. 18.1. Генотип этого растения обеспечивал хороший рост в одних условиях (длинные теплые дни) и слабый рост в других (короткие дни). Другие генотипы этого же вида реагировали на эти условия по-иному (Clausen, Keck, Hiesey, 1948*).
Приспособительные свойства фенотипических модификаций
Фенотипические реакции на нормальные изменения среды обычно носят приспособительный характер. Это можно видеть на примере листьев растений, находящихся на свету и в тени. У листьев, находящихся в тени («теневые листья»), площадь поверхности больше, что увеличивает их фотосинтетическую способность и компенсирует более низкую освещённость, тогда как у листьев, находящихся на солнце («солнечные листья»), площадь поверхности меньше, что понижает транспирацию и потери воды. Один и тот же генотип способен обеспечить развитие листьев этих двух типов в условиях сильной и слабой освещённости. Генетически детерминированная способность растения к таким различным фенотипческим реакциям даёт ему возможность приспосабливаться к варьирующим условиям освещёния.
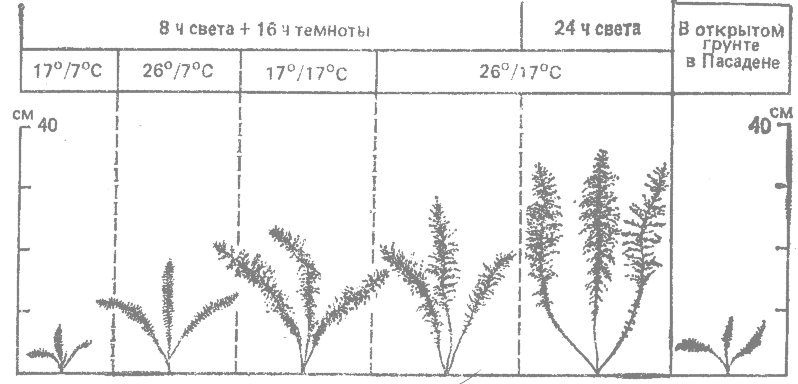 |
Рис. 18.1. Ростовые реакции отдельных частей одного растения Achillea borealis из Сьюарда (Аляска) в шести разных условиях. Первое число — дневная температура, а второе — ночная. Продолжительность периода роста составляла 3.5 мес. (Clausen, Keck, Hisey, 1948.*) |
Диапазон фенотипческой модифицируемости в разных крупных группах высших организмов весьма различен. Высшие растения, и в частности травянистые растения, характеризуются большой фенотипической пластичностью. Противоположную крайность представляют насекомые, отличающиеся крайне низкой фенотипической гибкостью.
Эти различия в пластичности непосредственно связаны с различиями в типах развития этих двух групп. Растительный организм развивается из точек роста, которые подвержены сильному воздействию факторов среды, преобладающих в период формирования его новых частей, так что «солнечные листья» появляются на ярком свету, а «теневые листья» — при слабой освещённости и т. д. В отличие от этого организм взрослого насекомого развивается внутри твердого наружного скелета, который формируется на одной из предшествующих стадий развития. Главные внешние свойства организма насекомого закладываются задолго до того, как они приобретают функциональное значение, и успевают принять окончательную форму к тому времени, когда они могут быть использованы.
Помимо этих различий в типах развития между растениями и насекомыми существуют более глубокие различия в стратегиях приспособления особей к колеблющимся условиям среды. Насекомые подвижны, а растения прикреплены к определённому месту. Взрослые насекомые в известных пределах могут приспосабливаться к переменным факторам среды, переходя в более теплое место в холодную погоду или в более влажное — при засухе, т. е. выбирая себе наиболее подходящее местообитание. Растение же, удерживаемое на одном месте корнями, не имеет выбора; для него главным средством индивидуального приспособления к изменениям среды служит способность к фенотвпическим модификациям.
Как типы фенотипических модификаций, так и диапазон фенотипической изменчивости зависят от потребностей организма. Поэтому весьма вероятно, что генотипы подвергались отбору на способность к проявлению приспособительных фенотипических реакций при изменении условий среды.
Роль фенотипических модификаций в эволюции
Фенотипические модификации охраняют отдельный организм от стрессовых воздействий среды и в той мере, в какой это удается, создают буфер, противостоящий отбору, определяемому средой. Теоретически популяция, состоящая из генотипов, способных обеспечить полный спектр приспособительных фенотипических реакций, вовсе не должна реагировать на давление естественного отбора. Практически фенотипическая пластичность сдерживает селективное воздействие среды.
Благодаря фенотипической пластичности популяция может выжить в среде, изменившейся неблагоприятным образом. Задержка элиминирующего действия отбора даёт популяции больше шансов на приобретение новых генетических вариаций в результате мутагенеза, потока генов и рекомбинаций, с помощью которых она может создать генотип, соответствующий новой неблагоприятной среде. Эту косвенную связь между фенотипическими модификациями и развитием новых адаптивных признаков называют эффектом Болдуина (Simpson, 1953b*).
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |