| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 14
Типы индивидуального отбора
Естественный отбор на индивидуальном уровне принимает различные формы в зависимости от того, какие варианты элиминируются или сохраняются из поколения в поколение. Можно выделить четыре формы отбора: направленный, стабилизирующий, дизруптивный и уравновешивающий.
Направленный отбор
Этот тип отбора обсуждался в гл. 10 и 12. Направленный отбор вызывает прогрессивное, или направленное в одну сторону, изменение генетического состава популяции (рис. 14.1). Он происходит в тех случаях, когда популяция находится в процессе приспособления к новой среде или же когда происходит постепенное изменение среды, а вслед за ней и постепенное изменение популяции. Хорошим примером первой ситуации служит увеличение частоты меланистической формы Biston betularia и других бабочек при внезапном изменении среды, а именно — её загрязнении промышленными отходами (гл. 12). Вторую ситуацию, в которой возникает направленный отбор, иллюстрируют постепенные изменения, происходящие в среде, как, например, изменения многих климатических факторов.
Стабилизирующий отбор
Совершенно иной характер носит стабилизирующий отбор. Если популяция хорошо приспособлена к данной среде, которая остается стабильной, то главное действие отбора состоит в элиминации плохо приспособленных и крайних вариантов, возникающих в результате мутагенеза, потока генов, расщепления и рекомбинации. Таким образом, сохраняются несомненно приспособленные генотипы, тогда как плохо приспособленные формы «выпалываются». Это и есть стабилизирующий отбор.
Действие стабилизирующего отбора графически представлено на рис. 14.1. Предполагается, что генетическая изменчивость популяции имеет нормальное распределение, т. е. по каждому изменчивому и измеримому признаку основная масса особей группируется вблизи среднего значения этого признака для данной популяции, а крайние значения наблюдаются лишь у немногих особей. Под действием стабилизирующего отбора эти особи, находящиеся на концах кривой нормального распределения, элиминируются из поколения в поколение. Преимущественное размножение особей, у которых значения признаков близки к среднему для данной популяции, приводит к постоянству модальных значений во времени. В отличие от этого при направленном отборе происходит элиминация крайних генетических вариантов на одной стороне кривой, в результате чего значение средней в ряду последовательных поколений смещается (рис 14.1).
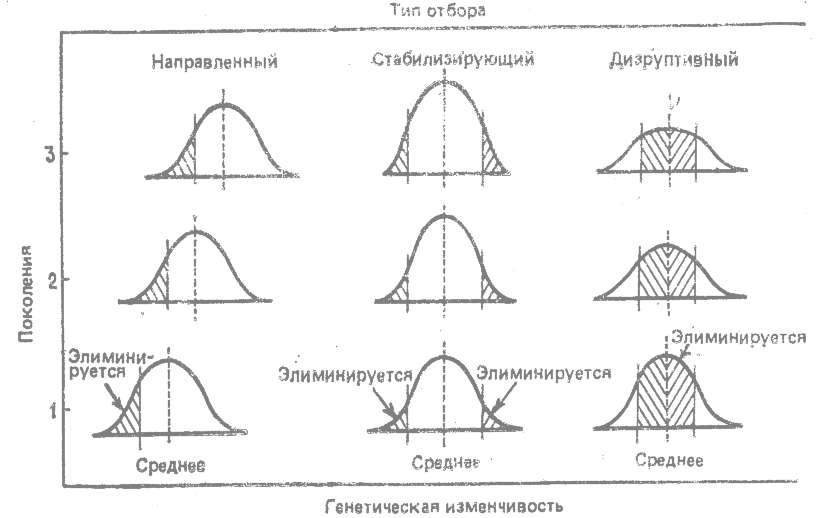 |
Рис. 14.1. Воздействие отбора трёх разных типов на генетическую изменчивость в популяции. |
Праут (Prout, 1962*) провел эксперимент по стабилизирующему отбору у Drosophila melanogaster. В качестве изменчивого признака он использовал время развития мух. В линии, подвергнутой стабилизирующему отбору, в каждом поколении отбирали особей, время развития которых приближалось к среднему значению. В результате такого стабилизирующего отбора в ряде поколений дисперсия времени развития в этой линии уменьшилась по сравнению с дисперсией в исходном поколении и в контрольных линиях, не подвергавшихся отбору.
Стабилизирующий отбор происходит везде и всюду, хотя заметить его трудно; это наиболее обычный тип отбора в природе. Мы уже приводили в качестве яркого примера направленного отбора березовую пяденицу (Biston beiularia), однако при этом нечего не было оказано о комплементарной роли стабилизирующего отбора в популяциях этой бабочки. До развития промышленности редкие меланистические мутанты элиминировались из популяции стабилизирующим отбором, который, вероятно, продолжался много веков. В измененной индустриальной среде, в которой обитают новые популяции, состоящие главным образом из меланистов, снова действует стабилизирующий отбор, устраняя появляющиеся время от времени светлые варианты.
Дизруптивный отбор
Дизруптивный отбор (известный также как разрушающий или разрывающий отбор) благоприятствует сохранению крайних типов и элиминации промежуточных. В результате он приводит к сохранению и усилению полиморфизма (рис. 14.1).
Рассмотрим вновь эксперимент Праута (Prout, 1962*) с отбором на время развития у Drosophila melanogaster. Одна линия дрозофилы была подвергнута дизруптивному отбору; для этого в каждом поколении скрещивали мух, вылуплявшихся раньше всех, с мухами, вылуплявшимися позже всех. Через ряд поколений это привело к увеличению дисперсии времени развития в сравнении с контрольными линиями, не подвергавшимися отбору.
Ряд экспериментов по дизруптивному отбору у Drosophila melanogaster провели Тудей и его сотрудники. В одном из таких экспериментов (Thoday, Boam, 1959*) производился отбор на число щетинок на теле*.
Отбор производился одновременно на большое число щетинок в группе «высоких» линий и на малое их число в группе «низких» линий. Этот дизруптивный отбор был направлен на то, чтобы разделить исходную популяцию на различные «высокие» и «низкие» субпопуляции. Но «высокие» и «низкие» линии постоянно скрещивались между собой, так что между ними всё время происходил обмен генами, действие которого противоположно действию дизруптивного отбора. Эксперимент продолжался таким образом до 36-го поколения.
В первых поколениях между «высокими» и «низкими» линиями не наблюдалось значительной дивергенции. Они начали дивергировать в 14-м поколении, а после 30-го поколения разошлись очень сильно. На рис. 14.2 представлены кривые изменения числа щетинок в одной «высокой» и одной «низкой» линиях, дивергировавших в результате отбора. Следует отметить, что «высокая» и «низкая» линии в этом эксперименте находятся в совершенно ином положении, нежели в описанном выше селекционном эксперименте с кукурузой (рис. 12.1); в эксперименте с кукурузой «высокая» и «низкая» линии были изолированы друг от друга, а в эксперименте с дрозофилой их скрещивали между собой. Как показал последний эксперимент, дизруптивный отбор может превалировать над потоком генов.
Эксперименты Стримса и Пиментела (Streams, Pimentel, 1961*) на Drosophila melanogaster интересны количественным подходом к взаимодействию между дизруптивным отбором и потоком генов. Эти авторы использовали четыре уровня потока генов — 6, 20, 50 и 0% (в контрольных линиях) и дизруптивный отбор двух интенсивностей — умеренный и сильный. Сильный дизруптивный отбор оказывал свое действие при 20%-ном потоке генов, но был неэффективен при 50%-ном потоке. Что касается умеренного дизруптивного отбора, то его действие не могло проявляться даже при слабом (6%-ном) потоке генов.
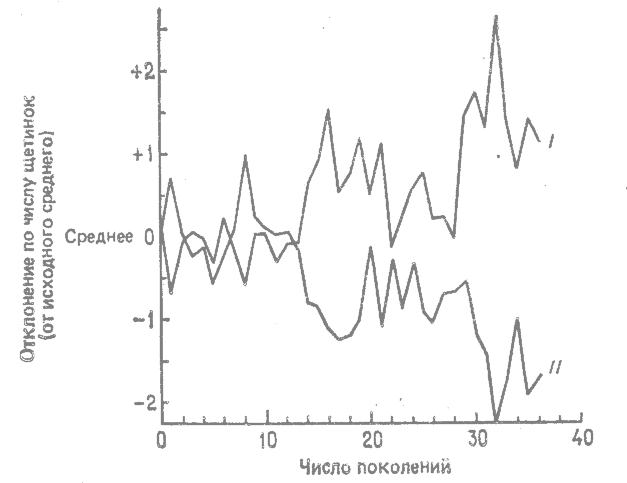 |
Рис. 14.2. Реакция на дизруптивный отбор по числу щетинок у Drosophila melanogaster в условиях максимального обмена генами. Объяснения см. в тексте. I — «высокая» линия; II — «низкая» линия. (Thoday, Boam, 1959.*) |
Дизруптивный отбор в природе
Эффективность дизруптивного отбора в экспериментальных условиях установлена достаточно хорошо. Теперь следует выяснить, какова его роль в природных популяциях. Здесь мы стоим на менее твердой почве и нам приходится говорить о некоторых вероятностях и возможностях, а не о бесспорных фактах.
Одна из возможных в природе ситуаций, в которой, вероятно, вступает в действие дизруптивный отбор, возникает в тех случаях, когда хорошо дифференцированные полиморфные типы обладают явным селективным преимуществом по сравнению со слабо дифференцированными полиморфными типами. Примером служит половой диморфизм: самки и самцы с хорошо дифференцированными вторичными половыми признаками спариваются и размножаются более успешно, чем различные промежуточные типы (интерсексы, гомосексуалы и т. д.).
Вторая возможная ситуация возникает в тех случаях, когда полиморфная популяция занимает неоднородное местообитание (Levene, 1953*). Считается, что в таком местообитании полиморфные типы приспосабливаются к различным нишам или субнишам и соответственно живут в основном в этих особых нишах. Так, морфа А может обладать более высокой приспособленностью в субнише А, но более низкой — в субнише В', тогда как морфа В процветает в субнише В', но не в А', Это предполагаемое соответствие между разными морфами полиморфной популяции и набором субниш в гетерогенном местообитании могло возникнуть в результате дизруптивного отбора.
Примером, который, по-видимому, подтверждает описанную выше модель, служит желтушка (Colias eurytheme). Самки этой североамериканской бабочки полиморфны по окраске крыльев: встречаются самки с оранжевыми и самки с белыми крыльями. Этот полиморфизм обусловлен единичным геном.
В середине лета в нескольких местностях в Калифорнии пики активности у белой и оранжевой форм наблюдаются в разное время дня: белая форма более активна рано утром и в позднее послеполуденное время, а оранжевая форма — в полдень; по-видимому, эти две морфы различаются по предпочитаемым температуре и влажности. Кроме того, наблюдается и сезонная изменчивость частоты двух цветных морф в калифорнийских популяциях: осенью, с понижением температуры, частота белой морфы возрастает (Hovanitz, 1953; Remington, 1954*). Как показывают физиологические исследования, пигментация крыльев играет важную роль в регуляции температуры у Colias eurytheme (Watt, 1969*). Таким образом, создаётся впечатление, что полиморфизм окраски у этого вида способствует расширению диапазона температур и продолжительности сезона, в пределах которых его популяции сохраняют активность.
Третья ситуация, в которой проявляется действие дизруптивного отбора в природе, наблюдается у растений при некоторых особых условиях. Представим себе популяцию растений, которая, будучи прикреплена к определённому месту, свободно скрещивается и занимает участок с двумя зонами, различающимися экологически, например, по типу почвы или по рельефу местности. В таких случаях у двух частей данной популяции растений нередко развиваются разные комплексы приспособительных при знаков. И такая дифференциация сохраняется, несмотря на скрещивание между ними. Почти несомненно, что главным регулирующим фактором в подобной ситуации служит дизруптивный отбор.
Высокогорный вид сосны Pinus albicaulis встречается в горах Сьерра-Невада в Калифорнии на границе древесной растительности и непосредственно над ней. Популяции, растущие на склонах гор, вплоть до границы леса, состоят из прямостоячих деревьев — обычной для этого вида формы роста. Выше границы древесной растительности этот вид представлен низкорослой стелющейся формой. Древовидная и стланиковая субпопуляции переходят одна в другую, и между ними происходит перекрёстное опыление при помощи ветра, о чем свидетельствует наличие особей промежуточного типа; однако в субальпийской и альпийской экологических зонах под влиянием дивергентных направлений отбора сохраняются соответственно различные формы роста. Аналогичные различия по обе стороны узкого пояса вдоль границы древесной растительности наблюдаются и у других высокогорных видов хвойных, а также у ив (Clausen, 1965*).
Другая группа примеров, иллюстрирующих действие такого же комплекса сил, относится к пастбищным злакам, произрастающим в районе свинцовых рудников в Уэльсе. В таких районах обычная для пастбищных лугов почва, не содержащая скольконибудь значительных количеств свинца, на отвалах резко переходит в почву с высоким содержанием свинца. Agrostis tenuis и другие виды злаков образуют в этом районе непрерывные популяции, область распространения которых захватывает районы с различными типами почвы. Генетическое единство таких популяций поддерживается благодаря перекрёстному опылению при помощи ветра. Тем не менее между субпопуляциями, растущими на пастбищных почвах и не переносящими свинец, и толерантными к свинцу субпопуляциями, растущими на отвалах, возникает слабая расовая дивергенция (Jain, Bradshaw, 1966; Antonovics, 1971*).
Уравновешивающий отбор
Гетерозиготы нередко превосходят соответствующие типы гомозигот по общей мощности или по тому или иному компоненту жизнеспособности, например по способности к конкуренции или по устойчивости к заболеваниям. Таким превосходством могут обладать генотипы, гетерозиготные по единичному гену или по целому блоку генов.
В тех случаях, когда гетерозигота Аа обладает селективным преимуществом по сравнению с одной или с обеими гомозиготами, отбор способствует сохранению в популяции обоих аллелей (А и а). В генофонде создаётся равновесная частота аллелей А и а, точный уровень которой определяется относительными селективными ценностями альтернативных типов. Однако, до тех пор пока частоты аллелей контролируются отбором, ни аллель A, ни аллель а не элиминируются. В популяции поддерживается состояние сбалансированного полиморфизма.
Отбор, благоприятствующий гетерозиготам, известен под названием уравновешивающего отбора (а также превосходства или преимущества гетерозигот).
При уравновешивающем отборе не происходит полного закрепления или элиминации альтернативных аллелей или блоков генов, как было описано выше; единицей, которой благоприятствует отбор, в данном случае оказывается гетерозиготная пара аллелей или сочетание генов. Это имеет известное отношение к вопросу об ожидаемых результатах направленного отбора в сбалансированной полиморфной системе. Полагают, что направленный отбор в сочетании с уравновешивающим приводит не к замене в генофонде одного гена или блока генов другим, а к ряду переходов от одной гетерозиготной комбинации к другой, например A1A2→A2A3→A3A4 (Lerner, 1954, р. 113—114*).
Сбалансированный полиморфизм, основанный на превосходстве гетерозигот, — явление, которое распространено довольно широко. Среди животных оно наблюдается, например, у прямокрылых, кур, мышей, человека и разных видов дрозофилы; среди растений оно обнаружено у кукурузы (Zea mays), ячменя (Hordeum sativum), Arabidopsis и Oenothera. Наиболее подробно превосходство гетерозигот изучено у Drosophila pseudoobscura, на примере которой оно будет описано в следующем разделе.
Превосходство гетерозигот у Drosophila pseudoobscura
Дикий североамериканский вид Drosophila pseudoobscura известен своей изменчивостью по хромосоме III. У этого вида (как отмечалось в гл. 4) описано 16 типов хромосомы III, отличающихся друг от друга по инверсиям. Все типы инверсий, выявляемые цитологически в хромосомах слюнных желез личинок, получили названия (Стандарт, Эрроухед, Чирикауа, Тимберлайн, Пайкс-Пик и т. д.) и буквенные обозначения (ST, AR, CH, TL, РР и т. д.). Большая часть популяций полиморфна по некоторым типам инверсий, и в них встречаются различные возможные гомозиготы и гетерозиготы.
Инвертированные участки хромосомы различаются как по содержащимся в них генам, так и по своим цитологическим признакам. Популяция, полиморфная по инверсиям ST, CH и AR, содержит не только хромосомы трёх разных типов, но и три раз ных набора генов в инвертированных участках; поэтому генотипы ST/CH, ST/AR и CH/AR гетерозиготны не только по инверсиям, но и по генам. Инверсии предотвращают эффективную рекомбинацию генов в гетерозиготах по инверсиям. Следовательно, полиморфизм по инверсиям у D. pseudoobscura одновременно представляет собой полиморфизм по блокам генов, которые остаются интактными из поколения в поколение.
и гомозигот по инверсиям в хромосоме III (данные Epling, Mitchel, Mattoni, 1953*) | ||
| Тип | Наблюдаемые частоты | Ожидаемые частоты |
| ST/CH | 0.232 | 0.168 |
| ST/AR | 0.232 | 0.168 |
| Все гетерозиготы | 0.830 | 0.724 |
| ST/ST | 0.134 | 0.184 |
| Все гомозиготы | 0.170 | 0.273 |
Добржанский и его школа изучали полиморфизм по инверсиям в природных популяциях D. pseudoobscura по всему ареалу этого вида. Наиболее обширные выборки в течение многих лет отбирали из некоторых популяций Сьерра-Невады и Сан-Джасинто (Калифорния). Кроме того, проводились многочисленные лабораторные эксперименты в популяционных ящиках с линиями мух, отловленных в тех же самых природных популяциях в Калифорнии.
В этих калифорнийских горных популяциях обычно встречаются инверсии ST, СН и AR; встречаются также, но с низкой частотой, инверсии TL, РР и SC. Нередко частота гетерозигот по инверсиям бывает выше той, которую следовало бы ожидать на основании равновесия Харди — Вайнбарга. Действительную частоту различных типов инверсий в гаметном фонде определяют с помощью соответствующих методов взятия выборок из популяции; на основании полученных данных по формуле Харди — Вайнберга вычисляют ожидаемую частоту гетерозиготных генотипов. Затем эту ожидаемую частоту сопоставляют с действительной частотой гетерозигот по инверсиям. При этом в определённое время года в природных популяциях постоянно обнаруживается статистически значимый избыток гетерозигот.
Так, в выборках, взятых в мае 1952 г. в горах Сан-Джасинто из одной популяции, полиморфной по пяти инверсиям, были обнаружены значительные различия между фактическими и ожидаемыми частотами гетерозигот и гомозигот по инверсиям (табл. 14.1). Данные табл. 14.1 показывают, что наблюдается устойчивый избыток гетерозигот и соответствующий недостаток гомозигот по сравнению с тем, чего следовало бы ожидать, исходя из формулы Харди — Вайнберга.
Наблюдавшиеся отклонения от равновесия Харди — Вайнберга можно объяснить на основании отбора в пользу гетерозигот. Один из способов проверки этой гипотезы состоит в том, чтобы сравнить частоты гетерозигот и гомозигот по инверсиям в выборке яиц, взятых из природной популяции, с их частотами в выборке взрослых мух. Оказалось, что соотношение гетерозиготных и гомозиготных генотипов в выборках яиц соответствует формуле Харди — Вайнберга. Недостаток гомозигот в выборках взрослых особей был, вероятно, обусловлен дифференциальной смертностью в период развития от яйца до взрослых стадий, действующей в пользу гетерозигот (Dobzhansky, Levene, 1948*).
Параллельные данные о превосходстве гетерозигот были получены для искусственных популяций, выращенных в популяционных ящиках. Основателями этих искусственных популяций служили линии мух, взятые из природных популяций Сан-Джасинто и несущие хромосомы ST, СН и AR. Популяционные ящики содержали в тепле и допускали возникновение в них перенаселенности. На стадии яйца соотношение гомозигот и гетерозигот по инверсиям соответствовало формуле Харди — Вайнберга. Однако на стадии взрослых особей в тех же самых искусственных популяциях наблюдался достоверный избыток гетерозигот по инверсиям (Dobzhansky, 1947a*).
Изучение развития и поведения мух показало, что более высокую приспособленность гетерозигот по инверсиям можно разложить на ряд различных компонентов: более высокая жизнеспособность на стадиях, предшествующих достижению половой зрелости; более высокая скорость развития; большая продолжительность жизни, плодовитость и быстрота нахождения брачного партнера (Moos, 1955; Dobzhansky, 1970, p. 137—138*).
Следует снова подчеркнуть, что превосходство гетерозигот проявляется только при определённых температуре, корме и плотности популяции; в отсутствие нужных условий, будь то природные или искусственные популяции, гетерозиготы по инверсиям теряют свое селективное преимущество перед гомозиготами.
Важно отметить, что природные популяции периодически действительно оказываются в таких условиях среды, которые приводят к проявлению превосходства гетерозигот. В этих популяциях, если не постоянно, то во всяком случае с перерывами, действует уравновешивающий отбор. Taкого прерывистого действия достаточно для постоянного сохранения сбалансированного полиморфизма. В прохладную погоду в начале лета (май и июнь) в природных популяциях возрастает частота хромосом AR и CH, а в жаркую летнюю погоду и осенью (с июля по сентябрь или октябрь) возрастает частота хромосом ST, однако эти изменения никогда не доходят до полного закрепления или элиминации (Dobzhansky, 1943; 1947b; 1948a*). Таким образом, уравновешивающий отбор способствует расширению сезонных границ популяции по сравнению с теми, какие, вероятно, могут быть у мономорфной популяции.
Факторы, благоприятствующие полиморфизму
Три типа отбора, обсуждавшиеся в этой и предшествующей главах, имеют непосредственное отношение к проблеме широкого полиморфизма, наблюдаемого в природных популяциях.
В пятидесятых годах была выдвинута гипотеза о том, что неоднородность среды должна способствовать полиморфизму: местообитание, в котором имеются многочисленные ниши и субниши, может предоставить условия, необходимые для существования, большему числу полиморфных типов, чем более однородная среда (da Cunha et al., 1950; da Cuhna, Dobzhansky, 1954; Dobzhansky, 1950b; Ludwig, 1950; Levene, 1953; обзоры см. Mayr, 1963; Hedrick et al., 1976; Lacy, 1982*). В пользу этой гипотезы свидетельствуют эмпирические данные по разным организмам, например по тропической Drosophila willistoni (da Cunha et al., 1950; 1959; da Cunha, Dobzhansky, 1954*), по питающейся грибами дрозофиле из восточной части США (Lacy, 1982*) и по средиземноморским моллюскам (Lavie, Nevo, 1981*).
Усиление полиморфизма в неоднородных средах сначала приписывали влиянию уравновешивающего отбора, и такое объяснение действительно в значительной мере справедливо, однако Левин (Levene, 1953*) показал, что в неоднородной среде полиморфизм может поддерживаться и за счет дизруптивного отбора.
Третий тип отбора, способствующий усилению полиморфизма, — это отбор, зависящий от частоты (Clarke, 1979*). Такой отбор, благоприятствующий редким вариантам., обычно связан с хищничеством и паразитизмом. Поскольку эти экологические взаимодействия часто встречаются в природе, было высказано мнение, что отбор, зависящий от частоты, служит, возможно, главным фактором полиморфной изменчивости в природных популяциях (Clarke, 1979*).
Возникновение в популяциях нейтральных в селективном отношении аллелей служит четвертым источником полиморфной изменчивости (Kimura, Ohta, 1971; Kimura, 1979*). Такие селективно нейтральные аллели несомненно существуют. Однако вопрос о том, сколь часто они встречаются в природных популяциях, в настоящее время вызывает значительные разногласия. Мы вернемся к этому в одной из последующих глав.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |