| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 8
Рекомбинация
Мутационный процесс и поток генов могут создать в популяции изменчивость по единичным генам. Если в результате таких первичных процессов возникает аллельная изменчивость по двум или большему числу генов, то создаётся почва для действия вторичного процесса — рекомбинации, В результате рекомбинации новые аллели, носителями которых первоначально, вероятно, были разные особи, могут сочетаться в одном генотипе. За счет рекомбинации число различающихся генотипов в популяции может увеличиться; этот процесс превращает небольшой первоначальный запас изменчивости по множественным генам в гораздо более значительное количество генотипической изменчивости.
Процесс рекомбинации
Допустим, что в популяции диплоидных организмов, размножающихся половым путем, в двух независимо распределяющихся генах А и В возникли новые мутации. Допустим далее, что носителями мутантных аллелей (а и b) первоначально были разные особи с генотипами АаВВ и ААВb соответственно. Теперь может начаться процесс рекомбинации, слагающийся из следующих этапов: 1) скрещивание между носителями различных мутантных аллелей: АаВВ×ААВb; 2) появление в F1 гетерозигот по двум генам АаВb (помимо других типов); 3) независимое распределение гамет с образованием четырёх классов гамет — АВ, Аb, аВ и ab; 4) образование в F2 девяти различных генотипов — ААВВ, …, aabb.
Большую часть этих девяти генотипов составляют новые генотипы. В начале процесса в популяции было три генотипа (ААВВ, АаВВ и ААВb); спустя два поколения она содержала девять генотипов, в том числе такие новые рекомбинантные типы, как ааВb и aabb,
Для того чтобы произошла рекомбинация, гены А и В необязательно должны быть независимы. Гены A и B могут рекомбинироваться, находясь в разных хромосомах или же в разных локусах одной хромосомы. Сцепление, если только оно не слишком тесное, снижает частоту рекомбинаций, но не предотвращает их образования.
Следует сказать несколько слов о терминологии. В рекомбинации генов участвуют два процесса: независимое распределение негомологичных хромосом и кроссинговер между негомологичными хромосомами. Молекулярные биологи и микробиологи, используя термин рекомбинация, имеют в виду исключительно второй процесс. Биологи, имеющие дело с организмами и популяциями, используют этот термин в его классическом смысле, т. е. имея в виду как сцепленные, так и несцепленные гены; именно в этом смысле мы пользуемся им в этой книге.
Количество генотипической изменчивости
Допустим, что число отдельных генов, присутствующих в двух аллельных формах, возрастает в арифметической прогрессии (2, 3, …, n). Число диплоидных генотипов возрастает при этом по экспоненте (З2, З3, …, Зn). В общем число возможных диплоидных генотипов (g) равно 3n.
Как мы видели выше, два отдельных гена (A и B), каждый из которых представлен двумя аллельными формами, могут образовать 9 генотипов, т. е. g=З2. Согласно менделевской генетике, тригибридное скрещивание с участием трёх генов (A, B и С) даёт 27 генотипов (g = 33).
Сцепление нарушает частоты рекомбинантных типов, но не изменяет общего числа возможных типов. Если отдельные гены не сцеплены между собой, то гетерозиготы по двум или по многим генам дают различные рекомбинации с определёнными частотами. Если гены сцеплены, но могут быть разделены в результате кроссинговера, то рекомбинанты всё же образуются, но с более низкими частотами, пропорциональными силе сцепления.
Полиморфные гены обычно представлены в природных популяциях множественными аллелями. В таких случаях для определения числа возможных генотипов следует возвести в n-ю степень не 3, а какое-то большее число. Общая формула для возможного числа диплоидных генотипов (g), выражаемая через n (число отдельных генов) и r (число аллелей каждого гена), имеет вид
| g == | [ | r(r+1) 2 | ] | n |
Рассмотрим применение этой формулы для случая всего двух отдельных генов с различным числом аллелей. Результаты представлены графически на рис. 8.1. Индивидуальная изменчивость, обусловленная рекомбинацией, быстро возрастает с увеличением в арифметической прогрессии числа аллелей в этих двух локусах.
Рассмотрим далее генотипическую изменчивость, возможную при наличии множественных аллелей более чем в двух локусах. Некоторые примеры приведены в табл. 8.1. Как показывает таблица, в результате рекомбинации между пятью генами, каждый из которых имеет по 10 аллелей, может возникнуть полмиллиона генотипов. Если выйти за пределы этой таблицы и рассмотреть случай шести несцепленных локусов с 10 или более аллелями в каждом, то окажется, что число диплоидных рекомбинантов исчисляется в миллиардах.
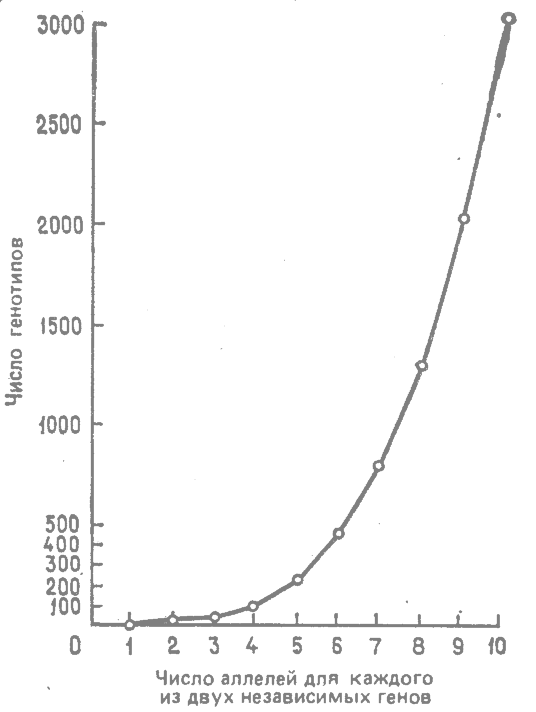 |
Рис. 8.1. Возрастание рекомбинационной генотипической изменчивости с увеличением числа аллелей в каждом из двух отдельных локусов. (Grant, 1963.*) |
При генетическом изучении природных популяций высших животных и растений в большинстве случаев обнаруживается, что они полиморфны по разным генам. Допущения, лежащие в основе приведенных выше численных примеров, нельзя считать нереалистичными; скорее они слишком занижены.
Очевидно, что рекомбинация представляет собой механизм, создающий огромные количества индивидуальной генотипической изменчивости. При наличии умеренного полиморфизма всего по нескольким несцепленным генам из этой генной изменчивости путем рекомбинации может возникнуть астрономическое число генотипов. При умеренной генной изменчивости число рекомбинантов вполне может оказаться выше общего числа особей данного вида. Именно по причине рекомбинации у организмов с половым размножением две особи, развивающиеся из разных зигот, никогда не бывают совершенно одинаковыми в генотипическом отношении.
в результате рекомбинаций между различным числом отдельных генов, каждый из которых имеет разное число аллелей (Grant, 1963*) | |||||||||||||||||||||||||
лей каждого гена | Число генов | ||||||||||||||||||||||||
| 2 | 3 | 4 | 5 | n | |||||||||||||||||||||
| 2 | 3n | ||||||||||||||||||||||||
| 3 | |||||||||||||||||||||||||
| 4 | |||||||||||||||||||||||||
| 5 | |||||||||||||||||||||||||
| 6 | |||||||||||||||||||||||||
| 7 | |||||||||||||||||||||||||
| 8 | |||||||||||||||||||||||||
| 9 | |||||||||||||||||||||||||
| 10 | |||||||||||||||||||||||||
| r |
|
|
|
|
| ||||||||||||||||||||
Рекомбинация и мутагенез
Рекомбинация сцепленных генов происходит в результате кроссинговера. Если сцепленные гены занимают локусы, находящиеся очень близко друг от друга, то кроссинговер случается редко. Появление рекомбинантов среди потомков в этом случае также наблюдается редко. Рекомбинанты ведут себя так же, как и мутанты.
Сходство между редкими рекомбинантами и мутантами иллюстрирует следующая модель. Два тесно сцепленных гена, А и В, регулируют сходные процессы и могут замещать друг друга.
Иными словами, нормальный фенотип создаётся либо двумя доминантными аллелями Ab/aB , либо одним из этих доминантных аллелей (А или В). Гетерозиготы по двум генам Ab/aB имеют нормальный фенотип и обычно размножаются в чистоте. Изредка между локусами А и В происходит кроссинговер, в результате чего возникает некоторое число гамет ab. Эти гаметы дают зиготы aabb, имеющие «мутантный» фенотип и размножающиеся в чистоте по отклоняющемуся признаку. Если кроссинговер между А и В происходит с частотой, сравнимой с частотой мутаций, то отличить такой редкий рекомбинант от мутанта обычными методами невозможно.
При помощи тонкого генетического анализа у Drosophila melanogaster, Zea mays и некоторых грибов было выявлено несколько случаев, соответствующих описанной выше общей модели. У дрозофилы кроссоверы встречались с частотой 0.00026 и, возможно, даже ещё реже. У кукурузы (Zea mays) имеется сложный ген А, определяющий окраску зерен и других частей растения; этот ген состоит из двух лежащих рядом субгенов, которые изредка рекомбинируют, давая формы, внешне сходные с мутантами (см. обзор Grant, 1975, гл. 4).
Из этих данных следует, что в любой группе непроанализированных мутантных форм, по всей вероятности, содержится, помимо настоящих генных мутантов, некоторое число редких рекомбинантов.
Адаптивная ценность
Среди большого числа возникающих рекомбинаций некоторые могут превосходить по адаптивной ценности исходный тип. Это объясняется тем, что на адаптивную ценность каждого аллеля могут оказывать влияние другие гены данного комплемента. Взаимодействие между генами — один из факторов, определяющих функцию каждого отдельного гена.
Тимофеев-Ресовский (1934a,b)* измерял жизнеспособность различных мутантов Drosophila pseudoobscura при 25°С и выражал её в процентах от жизнеспособности особей дикого типа. У всех форм, мутантных по одному гену, жизнеспособность была понижена. Некоторые рекомбинанты, содержащие по два мутантных аллеля, обладали в одних случаях более низкой, а в других случаях более высокой жизнеспособностью, чем любая из форм, мутантных по одному аллелю. Формы, возникшие путем рекомбинации некоторых мутантных типов, имели более высокую адаптивную ценность по сравнению с мутантами по одному аллелю и, как видно из следующих данных, почти соответствовали дикому типу.
| Miniature | — | жизнеспособность | 69% |
| Bobbed | — | » | 85% |
| Miniature-bobbed | — | » | 97% |
Рекомбинация и сложность
Сложные фенотипические признаки определяются не единичными генами, а их сочетаниями. Подобные сочетания генов состоят из аллелей, которые действуют совместно и согласованно. Рекомбинация — это тот механизм, который объединяет генные сочетания.
Рекомбинация играет важную роль почти у всех организмов. Во всех царствах живого мира существуют те или иные способы обмена генетическим материалом. У эукариот главным способом, обеспечивающим рекомбинацию, служит половое размножение, а у прокариот тот же результат достигается за счет парасексуальных механизмов.
Хотя рекомбинация обнаружена во всех основных группах, её относительное значение у разных организмов весьма различно. Роль рекомбинации наиболее велика у высших животных, у которых она обеспечивается облигатным половым размножением, большим числом хромосом и другими особенностями генетической системы. Противоположная ситуация имеет место у бактерий и вирусов, которые благополучно существуют при минимальной степени рекомбинации. Промежуточное положение по обычному уровню комбинативиой изменчивости занимают высшие растения.
Эти широкие различия коррелируют со сложностью организмов. Бактерии — наиболее простые организмы с простым генотипом. Важные жизненные функции, как, например, способность синтезировать некоторые необходимые метаболиты, могут определяться у них единичными генами; новые простые функции такого рода нередко, по-видимому, возникают в результате генной мутации и естественного отбора.
В невероятно сложном генотипе того или иного высшего животного роль каждого единичного гена относительно меньше. Здесь фенотипические признаки, обладающие адаптивной ценностью, в большинстве случаев определяются сочетаниями генов, причём обычно очень сложными сочетаниями. Поэтому механизмы возникновения рекомбинации генов имеют у них существенное значение.
Роль рекомбинации в эволюции
Эволюционные изменения признаков, детерминируемых одним геном, могут возникнуть в результате сочетания мутационного процесса и отбора. Это сочетание играет наибольшую роль в эволюции бактерий. Оно влияет также на простые признаки многоклеточных организмов, выступая здесь, однако, лишь на вторых ролях в общем процессе изменения.
Эволюция новых сложных признаков у многоклеточных организмов начинается с возникновения изменчивости по множественным генам и завершается закреплением в популяции новой адаптивной комбинации генов. Рекомбинация — важный промежуточный этап в этом процессе.
Процесс начинается с мутаций в двух или нескольких генах. Мутантные аллели, в случае если они рецессивны, могут оставаться в течение многих поколений в диплоидном состоянии, не экспрессируясь. Диплоидное состояние представляет собой хранилище для мутационной и полигенной изменчивости, а ключом к нему служит половое размножение; оно создаёт всевозможных рекомбинантов из имеющегося в генофонде «сырья»*.
Биологическая функция пола состоит в производстве множества рекомбинантных типов. Вероятность того, что какая-либо определённая комбинация генов может быть собрана в одном ряду поколений за счет одного лишь мутационного процесса и без полового размножения, практически равна нулю. Рассмотрим возникновение производного генотипа abc от предкового генотипа ABC у гаплоидного организма. Если бы это превращение зависело от ряда мутаций в бесполых линиях, то оно протекало бы крайне медленно. Однако в популяции организма с половым размножением новый генотип abc может образоваться всего за два поколения в результате скрещивания между трёмя линиями, каждая из которых несёт по одному аллелю — a, b или с (Wright, 1931; Miller, 1932*).
Процесс рекомбинации слеп в отношении адаптивной ценности образующихся рекомбинантов. Он механически создаёт как негодные, так и полезные в адаптивном смысле типы рекомбинантов. Совершенно очевидно, что желательно свести долю первых до минимума. В любом сложном организме чужие гены из отдаленно родственной популяции вряд ли могут гармонично сочетаться с коадаптированными генами нативной популяции. Один из путей снижения доли плохо адаптированных рекомбинантов состоит поэтому в возведении преград, препятствующих широкой гибридизации.
Организация популяций в нескрещивающиеся между собой биологические виды, каждый из которых поддерживает свой особый коадаптированный генофонд, представляет собой, таким образом, естественное следствие полового размножения. Биологические виды — практическое следствие полового размножения. Для рекомбинации необходим половой механизм, а пол в свою очередь требует видовой организации (Dobzhansky, 1937b*). Вопрос этот рассмотрен также в работе Grant, 1981a*.
Процесс рекомбинации, действуя в границах вида, даёт широкое разнообразие рекомбинантов. Некоторые из них могут оказаться более совершенными по степени адаптации. Проблема перемещается теперь с создания многочисленных рекомбинантных типов на сохранение некоторых лучших из них. Половой механизм, создающий в одном поколении ценное сочетание генов, в следующем поколении неумолимо вновь разъединит их. Теперь решающий момент заключается в том, чтобы закрепить новые, более совершенные рекомбинанты.
Отбор мог бы в принципе постепенно заменить предковое сочетание генов каким-либо новым их сочетанием. Но отбор — это слишком неэффективный и медленный способ закрепления новой генной комбинации в обширной свободно скрещивающейся популяции.
Более благоприятные условия для закрепления новой адаптивной генной комбинации создаёт инбридинг, сопровождающийся отбором среди продуктов последнего (Grant, 1963; Shields, 1982**).
Инбридинг может вызываться различными причинами. Малые размеры популяции вынуждают свободно скрещивающийся организм к инбридингу. Локализованные типы расселения в обширной популяции также способствуют инбридингу (Bateman, 1950; Shields, 1982*). Система скрещивания, благоприятствующая родственным скрещиваниям или самооплодотворению, приведет к инбридингу независимо от величины популяции. Два важных особых случая закрепления генных комбинаций с помощью инбридинга и отбора обсуждаются в дальнейшем при рассмотрении дрейфа генов (гл. 16) и квантового видообразования (гл. 24 и 25).
Формирование и закрепление рекомбинантных типов требует различных, и в сущности несовместимых, условий: свободного скрещивания в одних случаях и инбридинга — в других. Эта несовместимость может быть преодолена чередованием циклов широкого свободного скрещивания и инбридинга. Так, популяция, которая обычно бывает многочисленной, может проходить через «узкое горлышко» низкой численности. В преимущественно самооплодотворяющейся группе растений или животных могут время от времени наступать периоды свободного скрещивания.
Инбридинг представляет собой половое размножение в ограниченной форме. Организмы могут сделать ещё один шаг в сторону его ограничения, совершенно отказавшись от полового размножения на протяжении короткого или длинного ряда последовательных поколений. Чередование одного полового поколения с рядом бесполых поколений — благоприятное компромиссное решение, делающее возможным как образование новых комбинаций генов, так и их последующую репликацию; об этом писали Райт (Wright, 1931*), а позднее и другие авторы. Жизненные циклы, в которых существует равновесие между половыми и бесполыми поколениями, фактически более или менее обычны во всех царствах эукариот*.
В итоге мы располагаем одним набором эволюционных сил — сочетанием мутационного процесса и отбора, позволяющим дать адекватное объяснение первоначальным стадиям эволюции и изменениям простых признаков у высших организмов, Исследователи молекулярной эволюции иногда склонны подчеркивать всеобщее значение системы «мутационный процесс — отбор» (например, Beadle, 1963; Jukes, 1966*). Они пытаются также объяснять в этих терминах органическую эволюцию вообще (Beadle, 1963*).
Однако эволюцию многоклеточных организмов нельзя удовлетворительно объяснить с помощью системы «мутационный процесс—отбор». Сложность структуры и функций таких организмов требует столь же сложных генных комбинаций. Это требование выдвигает рекомбинацию "на первый план. Затем развивается ряд генетических систем, способствующих рекомбинации и регулирующих её: пол, вид, инбридинг и вторичная асексуальность.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |