| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 23
Образование рас и постепенное видообразование
Эволюционная дивергенция
Дивергенция между близкими эволюционными линиями, если она протекает постепенно и продолжается долго, проходит с течением времени через ряд стадий, Общая предковая популяция даёт начало двум или более локальным расам, географическим расам, полувидам, биологическим видам и группам видов, последовательно сменяющим друг друга (рис, 23.1). Дивергенция может достигать самых высоких уровней — от рода до класса и типа.
Популяционные единицы и таксономические, категории представляют собой отдельные стадии процесса эволюционной дивергенции. Высшие таксономические категории представляют собой продукты макроэволюции. Нас здесь интересует процесс дивергенции на низших ступенях среднего уровня, т. е. на уровне рас и видов. Дивергенция на этих уровнях относится к проблемам видообразования.
Дивергенция на уровне рас и видов проявляется в трёх главных направлениях. Переход от низших к высшим уровням на ветвящемся филогенетическом древе сопровождается: 1) возрастанием дифференциации генотипа; 2) возрастанием морфологической, физиологической и поведенческой дифференциации; 3) более сильной изоляцией (рис. 23.1).
Усиление генетической дифференциаций с возрастанием уровня дивергенции иллюстрируется на примере группы Drosophila willistoni (Ayala et al., 1975*). Большое число популяций, относящихся к 14 таксонам, исследовали по 36 ферментным локусам. Сходства или различия между популяциями выражали с помощью показателя генетического сходства (I). Полученные данные сгруппировали в зависимости от уровня дивергенции и определили средний показатель сходства (I) для каждого уровня. Полученные результаты представлены в табл. 23.1; генетическое сходство уменьшается на трёх из четырёх ступенчатых повышений уровня дивергенции.
Следует иметь в виду, что в различных группах организмов три перечисленных выше набора признаков могут эволюционировать с разными скоростями. В некоторых группах относительно небольшая генотипическая дифференциация выливается в резко выраженную фенотипическую дифференциацию, тогда как в других группах наблюдается широкая генотипическая, но незначительная фенотипическая дифференциация. Как показало рассмотрение большой выборки групп растений и животных, репродуктивная изоляция лишь очень слабо коррелирует со степенью фенотипической дифференциации.
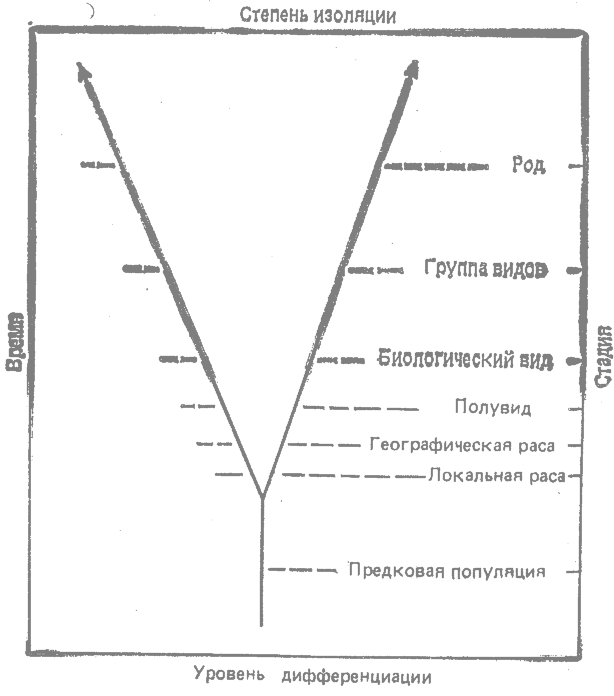 |
Рис. 23.1. Стадии дивергенции. |
Различия наблюдаются также в последовательности стадий. Некоторые из средних стадий видообразования в ряде случаев выпадают. Возможны также реверсии процесса дивергенции, кие отклоняющиеся способы видообразования будут рассмотрены позднее.
Факторы, участвующие в образовании рас
Образование рас, будь то локальные или географические, непрерывные или разобщённые расы, происходит под действием тех же эволюционных сил, которые обусловливают микроэволюционные изменения внутри популяций. Как уже говорилось, к основным процессам, создающим изменчивость, относятся мута генез, поток генов и рекомбинация, а к процессам, сортирующим возникшие изменения, — отбор и дрейф генов. Образование рас происходит в тех случаях, когда эти процессы протекают в разных направлениях (т. е. создают различные генофонды) в географически различных областях ареала данного вида.
между популяциями, расами и видами в группе Drosophila willistoni (Ayala et al., 1975*) | |
| Уровень дивергенции | I |
| Локальные популяции | 0.970 |
| Подвиды | 0.795 |
| Полувиды | 0.798 |
| Виды-двойники | 0.563 |
| Недвойниковые виды | 0.352 |
Роль отбора в образовании рас
Естественный отбор, несомненно, служит главной действующей силой в формировании рас. О роли отбора в образовании рас свидетельствуют две группы явлений. Во-первых, мы постоянно убеждаемся в том, что расовые признаки носят приспособительный характер по отношению к той среде, в которой обитает данная раса. Во-вторых, у разных видов, обитающих в средах со сходными диапазонами условий, нередко можно наблюдать параллельную расовую изменчивость.
Одно из обобщений систематики животных, известное под названием правила Бергмана, касается видов теплокровных позвоночных, которые встречаются как в теплых, так и в холодных областях. Правило Бергмана гласит, что такие виды обычно представлены в областях с холодным климатом крупными расами, а в областях с теплым климатом — мелкими расами. Это правило подтвердилось в отношении примерно 75—90% видов птиц и 60—80% видов млекопитающих в различных фаунах. Тем самым его можно признать справедливым обобщением, из которого имеются некоторые исключения (см. Mayr, 1963*).
Адаптивное значение различий в размерах тела, о которых говорится в правиле Бергмана, зависит от термодинамических свойств нагретого тела. В холодном климате большое нагретое тело обладает преимуществом, поскольку оно сохраняет тепло вследствие низкого отношения площади поверхности к объему. В умеренно теплом климате это преимущество исчезает. А в жарком климате небольшие размеры тела могут создавать определённое преимущество для регуляции температуры, так как высокое отношение площади поверхности к объему способствует рассеянию тепла.
Если бы подобная расовая изменчивость размеров тела была обнаружена лишь в нескольких отдельных случаях, то основания для утверждения о том, что эта изменчивость адаптивна, а следовательно, контролируется отбором, были бы весьма шаткими. Однако, поскольку подобная тенденция в изменении размеров тела наблюдается у многих различных видов в разных фаунах и служит основанием для создания правила Бергмана, представление о контролирующей роли отбора вполне оправдано.
Другое связанное с этим экогеографическое правило, правило Аллена, касается длины выступающих частей тела — ушей, носа, клюва, хвоста — у видов теплокровных животных, обитающих в различных климатических зонах. Согласно правилу Аллена, у рас данного вида птиц или млекопитающих, обитающих в холодных областях, выступающие части тела короче, чем у рас того же вида из теплых областей.
Эта общая тенденция, наблюдаемая в расовой изменчивости, подобно тенденции в изменчивости размеров тела, связана с проблемами терморегуляции у теплокровных животных. Сильно выступающие части тела рассеивают тепло в окружающую среду. Поэтому теплокровному животному, обитающему в холодном климате, невыгодно, чтобы эти части были длинными; в теплом же климате, напротив, сильно выступающие части тела дают определённую выгоду.
В различных группах животных наблюдаются многие другие упорядоченные элементы расовой изменчивости, имеющие, возможно, адаптивное значение. Более подробно от этом см. Rensch, 1960a, b; Mayr, 1963.*
Параллельная расовая изменчивость, коррелирующая с особенностями среды, обнаружена также у многих видов растений, занимающих обширные ареалы. Когда речь идет о растениях, то говорят обычно не об экогеографических правилах, а об экотипах. Виды растений, распространенные вдоль береговой линии, могут быть представлены стелющимися экотипами на песчаных дюнах и кустистыми экотипами на cкалах. Виды деревянистых растений на высоких горах могут быть представлены древовидными экотипами в субальпийской зоне и кустарниковыми стланиковыми экотипами в альпийской зоне (см. обсуждение вертикальных рас в гл. 20). Такие экотипы, несомненно, возникают в результате средового отбора. Имеется ряд обзоров, посвященных превосходным экспериментальным исследованиям экотипов у растений (см. Turesson, 1922; 1925; Clausen et al., 1940; 1948*).
Роль дрейфа генов
Случайная расовая изменчивость наблюдается у некоторых видов с колониальной популяционной системой. «Случайность» означает отсутствие явной корреляции между расовыми различиями и различиями в среде по трансектам, проведенным через область, занимаемую данным видом. Иллюстрацией служит новогвинейская галатея (Tanysiptera galatea) на территория Новой Гвинеи (Mayr, 1942*).
Новая Гвинея — очень большой остров (около 2500 км в длину и от 160 до 800 км в ширину). На нем можно встретить самые разнообразные типы климата и растительности — от вечнозеленого дождевого леса на одном конце до обнажающегося в сухое время года муссонного леса на другом конце. Новая Гвинея окружена многочисленными мелкими и средними островами, по климату и растительности сходными с соседними частями главного острова.
Главный остров, т. е. собственно Новую Гвинею, населяют три расы Tanysiptera galatea, а именно Т. g. galatea, T. g. meyeri и Т. g. minor. Эти расы распространены с перерывами от одного конца острова до другого. Они очень сходны. Пять других рас встречаются на некоторых из небольших соседних островов. Эти островные расы хорошо отличаются друг от друга и от рас, обитающих на соседних с ними частях главного острова, от которых они, возможно, произошли. Близкородственный вид Т. hydrocharts встречается на двух небольших островах, а также в одной части главного острова (рис. 23.2).
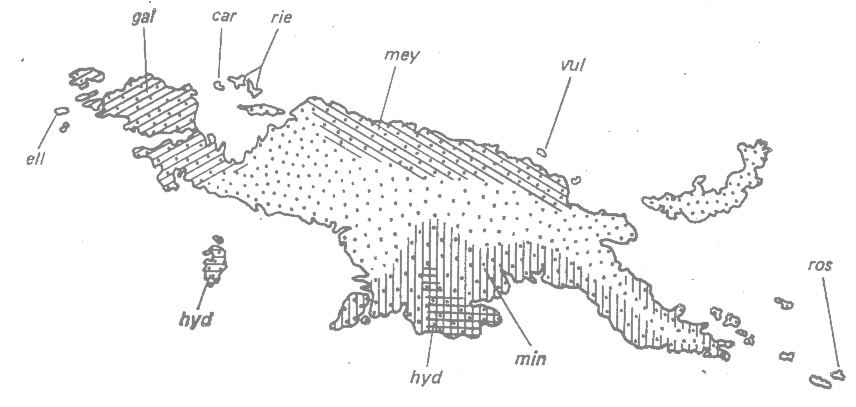 |
Рис. 23.2. Распределение рас галатеи (группа Tanysiptera galatea) на территории Новой Гвинеи. Главный остров: gal — Т. g. galatea; mey — T. g. meyeri; min — Т. g. minor; мелкие острова: ell—Т. g. ellioti; car —Т. g. carolinae; rie — T. g. riedelii; vul — T. g. vulcani; ros — T. g. rosseliana; обе области: hyd — T. hydrocharis, (Mayr, 1942.*) |
Майр (Mayr, 1942; 1954*) объясняет подобную расовую изменчивость следующим образом. Классическая расовая изменчивость на собственно Новой Гвинее — продукт отбора в большой популяционной системе с широким скрещиванием. Каждая из островных популяций была основана небольшим числом особей, мигрировавших с главного острова. Дрейф (или эффект основателя, по терминологии Майра) происходил в течение первых поколений колонистов, а в дальнейшем эти эффекты поддерживались благодаря пространственной изоляции от большой популяции главного острова. Все эти факторы привели к заметной дивергенции рас, населяющих малые острова, от соседней и, возможно, предковой популяции главного острова. Популяция, населяющая один из малых островов, дивергировала так сильно, что достигла видового ранга — Т. hydrocharis.
Нет необходимости допускать, что отбор не участвует в образовании рас в группе Т. galatea. Как указывалось в гл. 16, дрейф и отбор обычно действуют в природных популяциях совместно. Биологически реальным различием, касающимся факторов, ответственных за образование рас, следует считать различие между отбором в больших популяциях, с одной стороны, и взаимодействием отбор — дрейф генов — с другой.
Известно много других случаев расовой изменчивости, соответствующих тому, что наблюдается в группе Tanysiptera. Пример с группой Nigella aruensis (Ranunculaceae) на островах Киклады в Эгейском море и в прилежащих материковых областях описан выше (см. рис. 20.5).
Роль интрогрессии
Интрогрессия представляет собой особый способ естественной гибридизации, известный главным образом у растений. Интрогрессия заключается в образовании естественных гибридов F1 и их возвратном скрещивании с одним или обоими родительскими видами или полувидамя. Нередко возвратное скрещивание происходит многократно в ряде последовательных поколений. В результате возникает поток некоторых генов от вида или полувида, служащего донором, в популяционную систему, служащую реципиентом. Интрогрессия — это распространение потока генов и рекомбинации до видового уровня.
В тех случаях, когда два вида или полувида, А и В, маргинально-симпатричны и между ними происходит интрогрессия, в зоне перекрывания возникают расы, конвергентные по различным морфологическим признакам и экологическим предпочтениям. В области, занятой видом В, вид А в результате интрогрессии даёт начало расе, которая приближается к виду В по ряду признаков, и наоборот. Примеры этого выявлены у Iris, Trades cantia, Helianthus, Gilia и во многих других группах растений (см. обзор Heiser, 1973b; Grant, 1981*).
В данном случае, как и в случае дрейфа, не следует предполагать отсутствие отбора. Интрогрессия порождает изменения, а отбор сортирует их. Интрогрессивные расы — продукт совместного действия интрогрессии и отбора.
Роль потока генов
Следует ожидать, что экстенсивное расселение и поток генов должны задерживать образование рас. Ограниченное количество данных, которым мы располагаем, соответствует этому ожиданию.
Некоторые мигрирующие чешуекрылые занимают обширные ареалы при очень слабом развитии у них географических рас. У бабочки Vanessa cardui, отличающейся высокой подвижностью и распространенной почти по всему земному шару, на большей части её ареала образования географических рас не происходит. В Австралии и в Новой Зеландии эта бабочка представлена одной географической расой (Williams, 1958*). Линейчатый бражник (Hyles lineata), также интенсивно мигрирующий и широко распространенный вид, встречающийся по всей Северной и Южной Америке, за исключением крайнего севера, тоже представлен лишь одним подвидом (H. l. lineata) на большей части этого пространства. Еще по одному подвиду обитает в Уругвае и на Галапагосских островах (Schreiber, 1978*). Среди Lepidoptera можно найти и много других аналогичных примеров.
Противоположное положение вещей наблюдается у некоторых немигрирующих чешуекрылых, для которых характерен более «типичный» уровень образования рас. У девяти изученных в Скандинавии немигрирующих видов Lepidoptera была обнаружена хорошо выраженная географическая изменчивость в отличие от трёх мигрирующих видов из той же области, у которых географическая изменчивость выражена слабо или отсутствует вовсе (Williams, 1958*).
Параллельная ситуация наблюдается у гавайских цветочниц (Drepanididae). Принадлежащие к этой группе хорошие летуны — Himatione sanguinea, Vestiaria coccinea и Psittirostra psittacea — нe дифференцируются на подвиды на разных островах, если не считать один подвид H. sanguinea на острове Лайсан. В отличие от этого слабые летуны, такие, как Loxops virens и L. maculata, нередко бывают представлены на каждом острове или на группе небольших островов различными подвидами (Baldwin, 1953*).
Среди 23 североамериканских видов колибри (Trochilidae) есть как кочующие, так и оседлые формы. На основании данных о распространении и таксономии этой группы, собранных Джонсгардом (Johnsgard, 1983*), составлена табл. 23.2, выявляющая определённые корреляции: кочевой образ жизни коррелирует с монотипической структурой вида, а оседлый — с дифференциацией на географические подвиды. Два преимущественно кочующих вида (Setasphorus sasin и S. platycercus) имеют оседлые расы, которые выделяют в качестве подвидов.
ким образованием рас у североамериканских видов колибри (по данным Johnsgard, 1983*) | |||
| Географическая изменчивость | Образ жизни | ||
| сильно кочевой | кочевой, но с оседлыми расами | оседлый или главным образом оседлый | |
| Монотипические | 7 видов | — | 5 видов |
| 2–4 подвида | — | 2 вида | 7 видов |
Зарождающаяся репродуктивная изоляция
Репродуктивная изоляция и изоляция, обусловленная средой, могут создаваться и действительно возникают как побочный результат эволюционной дивергенции; об этом свидетельствуют следующие экспериментальные данные. Длительный эксперимент на Drosophila melanogaster был начат с одной популяции. Две совершенно идентичные популяции, полученные от этой исходной популяции, содержали в отдельных популяционных ящиках при различных температуре и влажности на протяжении 6 лет. Между этими двумя популяциями возникла сильная репродуктивная изоляция, обусловленная развивающимися между ними этологическими различиями и гибридной стерильностью. Этологическая изоляция детерминируется генами, распределенными по всем хромосомам, а гибридная стерильность имеет цитоплазматическую основу (Kilias, Alahiotis, 1982*).
Между разными географическими расами одного и того же вида нередко существует частичная репродуктивная изоляция. Расы, обитающие в разных географических областях, скорее всего имеют разные сезоны цветения или размножения. Столь же вероятно, что они имеют разные экологические потребности. Между географическими расами могут также существовать частичные внутренние преграды. Искусственные межрасовые гибриды F1 у травянистых цветковых растений обычно бывают полустерильны, а у их потомков F2 обычно проявляются некоторые признаки разрушения гибридов. Между отдельными парами географических рас можно наблюдать, во всяком случае в неполной или зарождающейся форме, все известные механизмы репродуктивной изоляции, описанные в гл. 21.
Рассмотрим случай, когда две географические расы какого-либо вида растений обитают в разных областях, в одной из которых климат прохладный и влажный, а в другой — теплый и сухой. Эти две расы будут подвергаться отбору на приспособленность к разным климатическим, а возможно, и разным эдафическим факторам; это первая стадия экологической изоляции. У таких рас цветение, по всей вероятности, происходит в разное время года; в результате возникает частичная сезонная изоляция. В прохладных влажных и теплых сухих районах, по-видимому, обитают разные группы пчел, а по мере того как местные расы растений приспосабливаются к обычно посещающим и опыляющим их видам пчел, в строении их цветков возникают изменения. Это приводит к зарождению механической и этологической изоляции, а может быть, и обеих сразу. Некоторые изменения в строении цветка, например изменение длины столбика, оказывают влияние на легкость скрещивания и могут привести к возникновению преграды несовместимости. Наконец, многочисленные генные различия, накапливаемые двумя такими расами за время их дифференциации, вполне могут привести к образованию у их гибридов каких-либо дисгармонирующих продуктов рекомбинации. Так возникает частичная нежизнеспособность или полустерильность гибридов, а также разрушение гибридов.
Таким образом, дивергенция на расовом уровне создаёт основу для возможного будущего видообразования.
Географическая теория видообразования
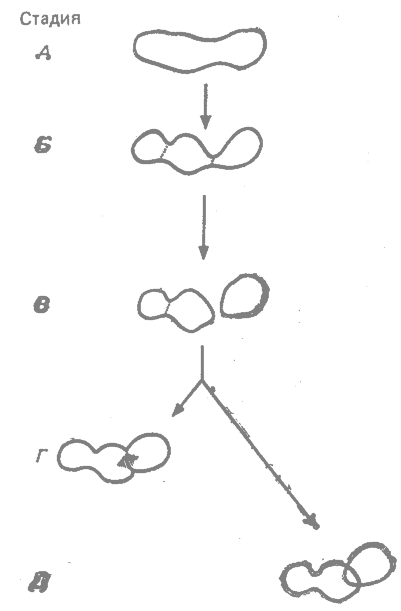 |
Рис. 23.3. Стадии географического видообразования. Объяснения см. в тексте. (Mayr, 1942, с изменениями.*) |
Согласно географической теории видообразования, созданной рядом авторов (К. Jordan, D. S. Jordan, Rensch, Mayr и др.) в первой половине XX в., биологические виды обычно возникают из географических рас (см. Mayr, 1942*). Пространственная изоляция — нормальная прелюдия к развитию репродуктивной изоляции.
Последовательность стадий при видообразовании по этой теории представлена на рис. 23.3. Широко распространенный однородный вид (рис. 23.3, А) дифференцируется на ряд непрерывных географических рас (Б), которые затем распадаются, превращаясь в разобщённые расы (В). Между разобщёнными расами в качестве побочного продукта дивергенции развивается репродуктивная изоляция. Затем в результате миграции контакт между этими расами восстанавливается. Если такой контакт наступает слишком рано, то он приводит к скрещиванию между расами и к вторичной интерградации (Г). Но если контакт происходит позднее, после того как репродуктивная изоляция достигла известного критического порога, дивергентные популяции могут обитать на одной и той же территории, не скрещиваясь между собой, и достигают таким образом, стадии молодых биологических видов (Д).
Один обычный способ видообразования, известный как географическое видообразование, протекает в этой последовательности — от предковой популяции через географические расы и аллопатрические полувиды до биологических видов. Все постулированные стадии действительно обнаружены в различных группах видов как у животных, так и у растений.
Ряд примеров, относящихся к одной естественной группе — видам гилии (Polemoniaсеае),— представлен на рис. 23.4. Все это однолетние диплоидные, преимущественно самоопыляющиеся растения пустынных и гористых районов американского юго-запада. В этой группе наблюдается следующая последовательность стадий (Grant, Grant, 1956; 1960*): 1) вид, состоящий из непрерывных географических рас (рис. 23.4, А); 2) разобщённые и различающиеся географические расы (Б); 3) сингамеон, состоящий из трёх маргинально-симпатрических полувидов, время от времени образующих гибриды (В); 4) пара стерильных при скрещивании друг с другом аллопатрических видов (Г); 5) пара стерильных при скрещивании друг с другом симпатрических видов (Д).
Пограничные случаи между географическими расами и вполне сформировавшимися видами приобретают в связи с этим особенно большое значение. Такие пограничные случаи, или полувиды, довольно обычны, что хорошо известно каждому систематику животных или растений. По мнению Майра (Mayr, 1942*), 12.5% всех таксономических видов птиц Северной Америки занимают промежуточное положение между видами и полувидами, так что орнитологи относят их то к одной, то к другой из этих таксономических категорий. Примерами служат два вида американских дятлов — Colaptes caper collaris и золотистый дятел (Colaptes auratus), а также два вида американских древесных славок — Dendroica auduboni и D. coronata.
Сравнимые примеры имеются во многих других группах животных и растений. Так, например, следует ли считать благородного оленя (Cervus etaphus), живущего в Евразии, и вапити (С. canadensis), распространенного в Северной Америке и Восточной Азии, географическими расами одного вида или самостоятельными аллопатрическими видами? В качестве аналогичного примера из царства растений можно привести Веймутову сосну (Pinus strobus), растущую в восточной части Северной Америки, и горную Веймутову сосну (P. monticola), растущую в западной её части.
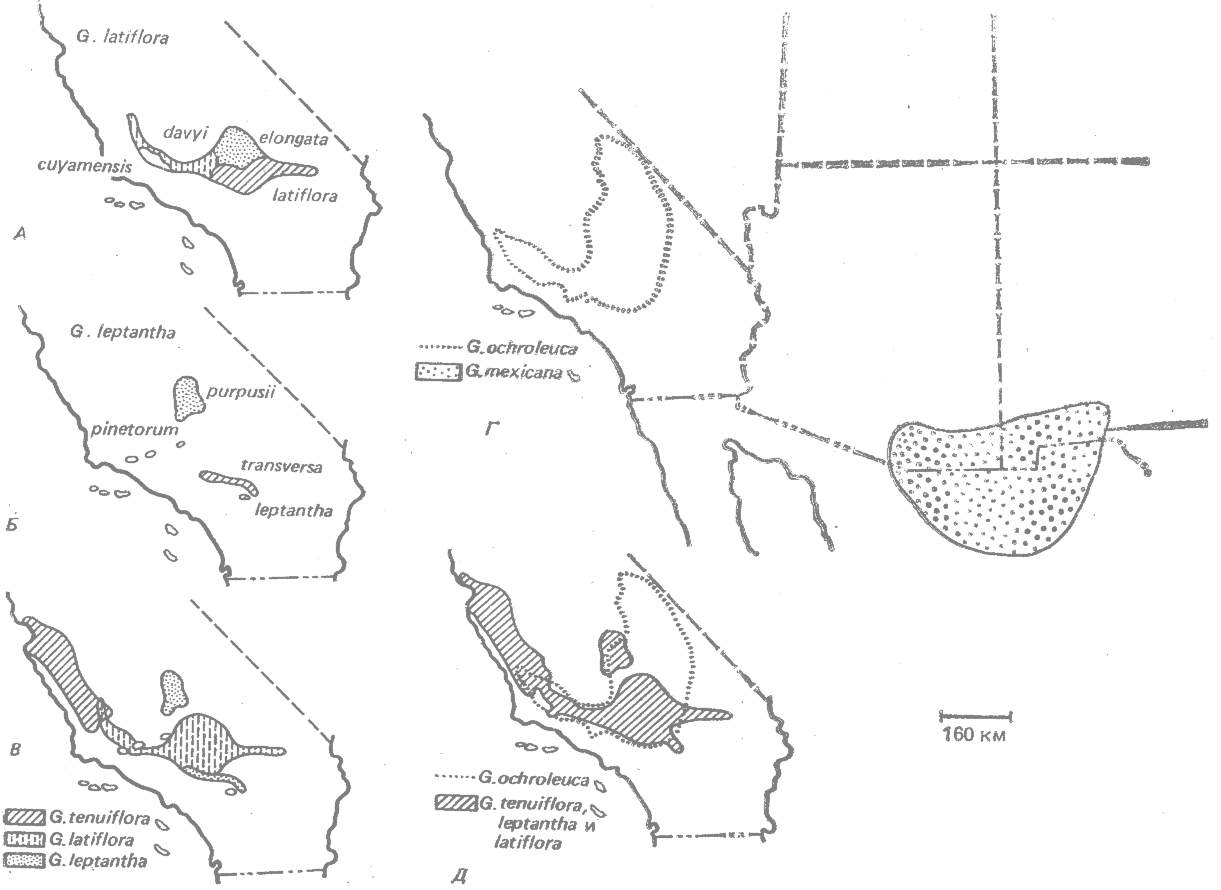 |
Рис. 23.4. Стадии географического видообразования у гилии (Polenomiaceae). Объяснения смотри в тексте. (По Grant, Grant, 1956; 1960*) |
Перекрывающиеся кольца рас
Особенно интересный тип пограничных случаев — цепочка интерградирующих рас А, В, С, D, E, образующих широкое кольцо с перекрыванием между формами А и Е. Расы А и Е представляют собой морфологически и экологически крайние типы в данной популяционной системе; они интерградируют в ряду А—В—С—D—Е. Если исходить только из этой особенности, то рассматриваемую популяционную систему можно считать обычным видом, состоящим из географических рас. Однако концевые и крайне различающиеся члены этого ряда, А и Е, сосуществуют симпатрически, а следовательно, отношения между ними соответствуют отношениям между симпатрическими видами.
Примером служит саламандра Ensatina eschscholtzii, обитающая на тихоокеанском склоне Северной Америки (Stebbins, 1949; 1957*). У этого вида наблюдается географическая изменчивость по типу окраски. У одной группы рас окраска красноватая, у другой — черная с желтым, а у третьей — черная с белым; при этом желтые или белые пятна могут быть мелкими или крупными (рис. 23.5).
Между различными расовыми типами окраски наблюдается полная интерградация (рис. 23.5 и 23.6). В Калифорнии эти интерградирующие расы образуют цепь, окружающую большое пространство: подвиды eschscholtzii (южная часть побережья Каланфорнии) — xanthoptica (центральная часть побережья) —platensis (Сьерра-Навада) — croceator—-klauberi (горы на юге Калифорнии) (рис. 23.6).
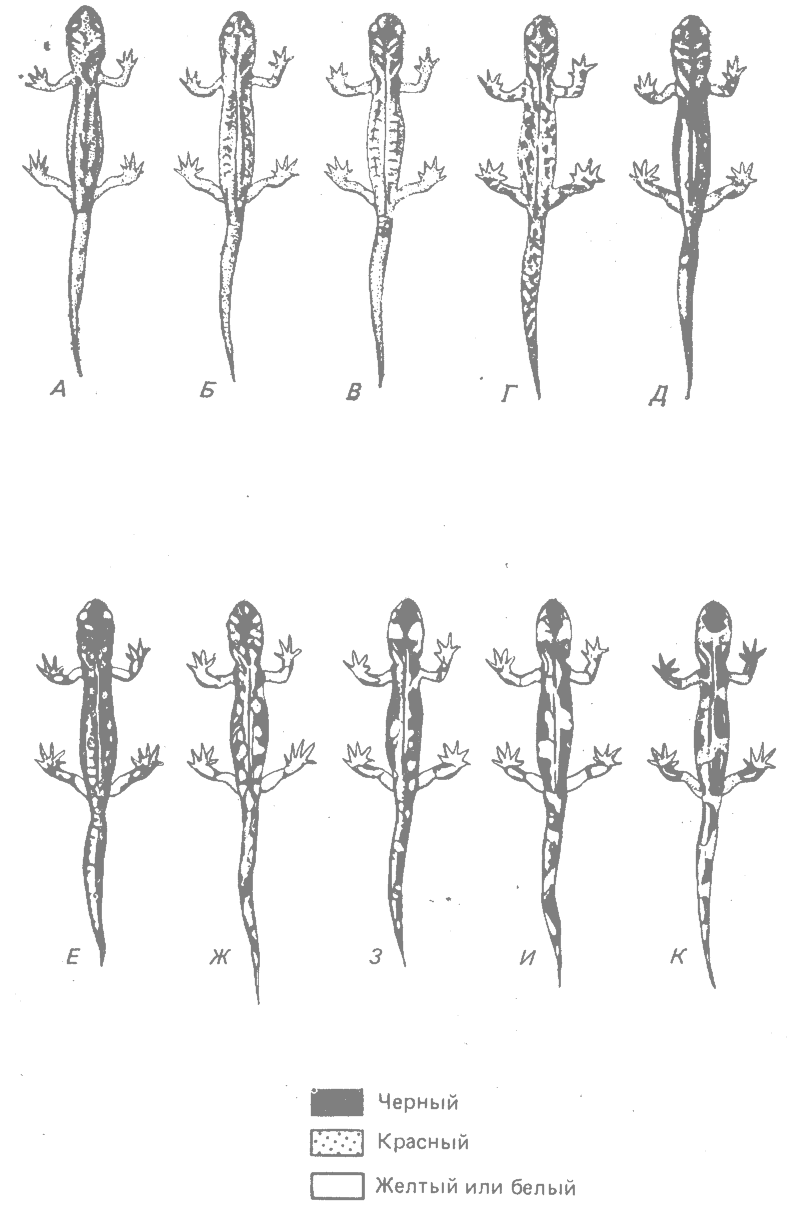 |
Рис. 23.5. Типы окраски тела у географических рас Ensatina eschscholtzii А, Е. е. eschscholtzii. Б. Е. е. xanihoptica. В. Е. е. oregonensis. Г. Е. е. picta. Д. Интерградация picta—platensis. Е. Е. е. platensis. Ж. Интерградация platensis—croceator 3. Е. е. croceator. И. Интерградация croceator—klauberi. К, Е. е. klauberi. (Из Stebbins, 1949.*) |
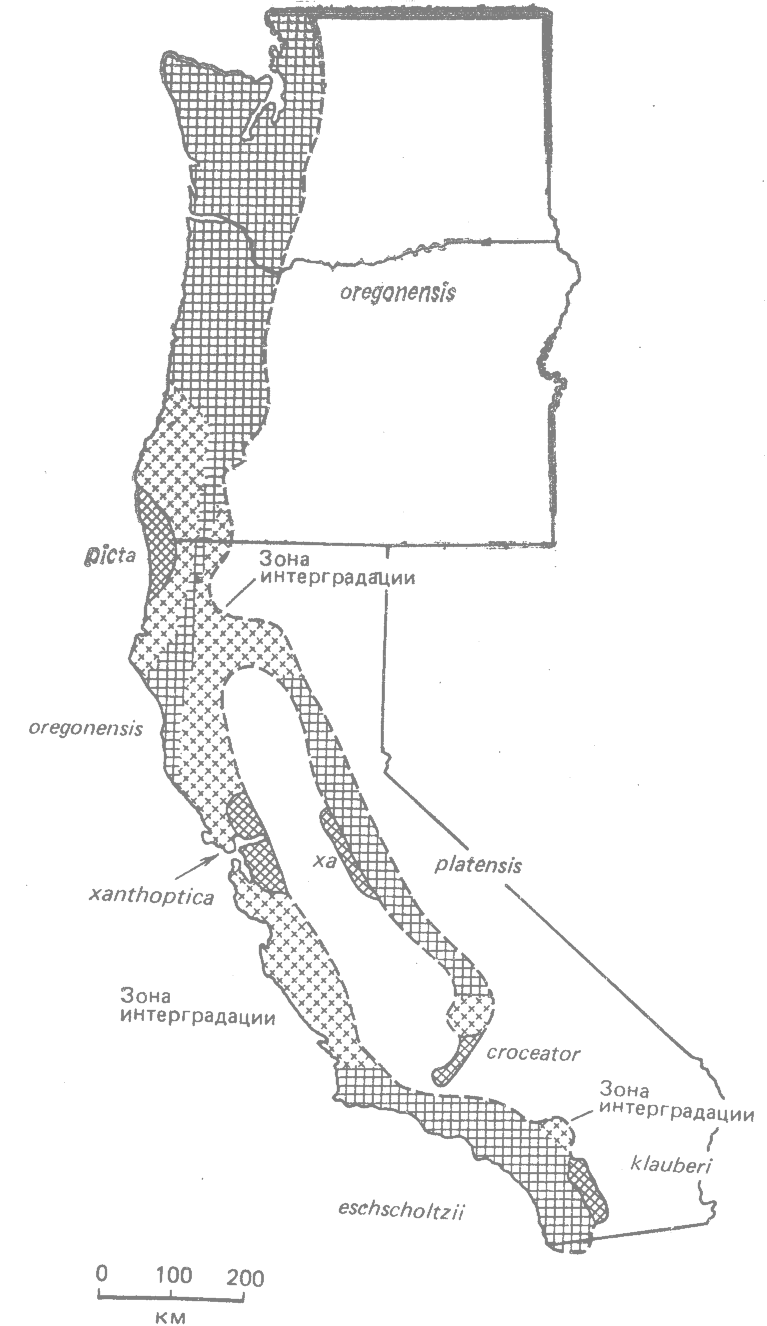 |
Рис. 23.6. Распространение географических рас Ensatina eschscholtzii в шт. Калифорния, Орегон и Вашингтон. Названия подвидов указаны на карте. Крестиками обозначены зоны интерградации между подвидами. (Stebbins, 1949; Wake, Yanev, 1986.*) |
Концевые звенья образуют краснотелая раса eschscholtzii и черная с белым klauberi или интерградирующие klauberi—croceator. Эти концевые расы перекрываются симпатрически в узкой зоне гор в южной Калифорнии (рис. 23.6) (Stebbins, 1949; 1957; Wake; Yanev, 1986*). В этой зоне контакта были обследованы четыре участка, где обитают оба подвида. В выборке из 86 особей, взятой в одном участке, не было ни одного гибрида, а в выборках из трёх других участков гибриды встречались с очень низкой частотой (Wake, Yanev, Brown, 1986).
У североамериканской пчелы Hoplitis producta (Megachilidae) наблюдается интерградация географических рас: gracilis— subgracilis—interior—producta (рис. 23.7). Кроме того, H. p. interior дала начало двум производным расам, ареал одной из которых (bernardina) перекрывается с ареалом gracilis (рис. 23.7) (Michener, 1947*).
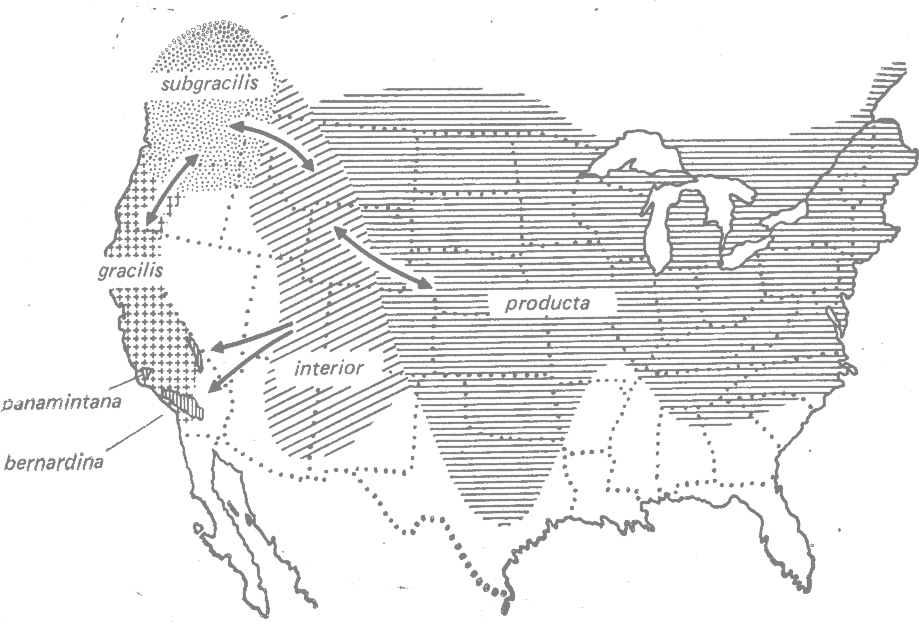 |
Рис. 23.7. Перекрывающееся кольцо рас у пчелы Hoplitis producta (Megachilidae). Интерградация между географическими расами показана двунаправленными стрелками. (Из Michener, 1947.*) |
Другие примеры кольцевого перекрывания описаны, в частности, у чаек (группа Larus argentatus), у оленьей мыши (Реromyscus maniculatus), у наземных улиток (группа Partula olympia) и у Drosophila paulistorum. Синицу (группа Parus major) в Евразии часто приводят в качестве примера перекрывающегося кольца рас; однако, поскольку в зонах перекрывания отмечена гибридизация, данный случай лучше отнести к категории сингамеонов (обзор см. Mayr, 1963*).
Реверсии дивергенции
Эволюционную дивергенцию на уровне рас и видов нельзя считать непоколебимым процессом. Дивергенция от географических рас до биологических видов происходит лишь до тех пор, пока продолжают действовать эволюционные силы, создающие эту дивергенцию. Процесс дивергенции может прекратиться на любой стадии. У некоторых видов географические расы возникли очень давно. Дивергенция может не только прекратиться, но и изменить свое направление на обратное на любой стадии, вплоть до возникновения полной стерильности между близкими видами. Естественная гибридизация, обычная у высших растений и хорошо известная во многих группах, представляет собой реверсию процесса дивергенции.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |