| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 9
Соседства
Один из факторов, влияющих на структуру популяций, — это миграция. Структуру популяций следует рассматривать в свете особенностей расселения и иммиграции. В зависимости от характера этих процессов фактически скрещивающиеся группы в локальной популяции перекрёстно-оплодотворяющихся организмов бывают меньше, и часто гораздо меньше, чем сама локальная популяция.
Различают две субпопуляционные единицы: панмиктическую единицу (Wright, 1943*) и соседство (Wright, 1946*).
Панмиктическая единица
Панмиктическая единица — это группа, в пределах которой происходит случайное спаривание (Wright, 1943; 1946*). У организмов с облитатным свободным скрещиванием и отсутствием предпочтений при спаривании это вполне реальная единица. Если предпочтительное скрещивание у животных является обычным, то панмиктическая единица, как несвойственная им, будет полезна главным образом в качестве некой идеальной абстракции (Яблоков, 1986*). Число особей в панмиктической единице обычно обозначают через Ne. Важный фактор, определяющий величину Ne, — это скорость миграции (m). Другим фактором, влияющим на величину Ne, считают плотность популяции; эта зависимость требует дальнейшего изучения.
Следует отметить, что в панмиктичеcкой единице имеет место известный инбридинг, доля которого равна 1/Ne (Wright, 1943*).
Концепция соседства
Панмиктическая единица существует лишь при том условии, что случайное спаривание носит локальный характер. Но во многих реальных ситуациях система скрещивания вызывает сильные отклонения от локальной панмиксии. Поэтому была предложена (Wright, 1946*) более общая субпопуляционная единица — соседство, пригодная для более широкого круга систем скрещивания.
Большинство последующих авторов не делали различия между соседствами и панмиктическими единицами. Для практических целей это обычно допустимо. В обоих случаях величина субпопуляции выражается одним и тем же символом Ne. Однако между соседством и панмиктической единицей существует тонкое различие, зависящее от того, носит ли случайное спаривание локальный характер или нет. При локальном случайном спаривании соседство соответствует панмиктической единице во всех отношениях. В противном же случае соседство соответствует производной чисто теоретической панмиктической единицы. В этом случае теоретическая панмиктическая единица используется для того, чтобы установить некий стандарт, из которого можно вывести величину соседства.
Концепция соседства исходит из двух допущений. Во-первых, вероятность соединения гамет в данной популяции уменьшается с увеличением расстояния между родительскими особями; это можно выразить как распределение вероятности спаривания. Во-вторых, доля инбридинга в панмиктической единице величиной Ne составляет 1/Ne. Таким образом, в обширной непрерывной популяции в каждом поколении происходит некоторый инбридинг.
Имея дело с популяцией, в которой не происходит случайного скрещивания, можно определить действительную долю инбридинга, а затем определить величину панмиктической единицы с такой же степенью инбридинга по формуле l/Ne и вычислить значение Ne, т. е. величину соседства. Фактор l/Ne можно использовать для выведения величины Ne в тех популяциях, где не имеет места локальная панмиксия.
Область, занимаемая соседством, и его величина
Область, занимаемая соседством в непрерывной популяции, соответствует величине пространства, занимаемого в среднем Ne особями. Важный фактор, оказывающий влияние на эту величину, — среднее квадратичное отклонение расстояний, на которые происходит расселение в популяции (σ). Источником данных для этой статистики служат частотные распределения расстояний, на которые происходит расселение, обсуждавшиеся в гл. 7.
В популяции, характеризующейся двумерным распределением в пространстве, область, занимаемая соседством, ограничена окружностью радиусом 2σ. Внутри этой окружности находится 86.5% родителей тех особей, которые локализованы в центре области, занимаемой соседством (Wright, 1946*). Иными словами, если мы возьмем особь, находящуюся в центре круга с таким радиусом, что родители этой особи с вероятностью 86.5% происходят из этого же круга, то соответствующая окружность будет границей области, занимаемой соседством.
Если популяция имеет линейную конфигурацию, то соседство представляет собой полосу длиной 3.5σ. Полоса такого размера вмещает 92.4% родителей особей, находящихся в центре этой полосы.
Величины 2σ; 3.5σ; 86.5% и 92.4% связаны с характеристиками нормального частотного распределения. Статистики для линейного соседства рассчитывают из нормальной кривой, в которой 92.4% значений попадает в интервал, заключенный между значениями — 1.75σ и + 1.75σ ( = 3.5σ). Соседству в форме круга соответствует двумерное распределение, где в окружности радиусом 2σ содержится 86.5% выборки (N. Fowler, личное сообщение).
Величина соседства — это число скрещивающихся особей в области, занимаемой этим соседством. Поэтому практический подход к оценке величины соседства состоит в том, чтобы получить среднее квадратичное отклонение расстояний, на которые происходит расселение (σ) из данных по расселению. Затем для двумерной популяции можно очертить в реальной популяции окружность с радиусом 2σ и подсчитать число находящихся внутри нее особей.
Эволюционные свойства больших популяций с соседствами различной величины
Большую непрерывную популяцию можно рассматривать как ряд перекрывающихся соседств. Соседства могут быть большими или маленькими по сравнению с величиной всей популяции.
Скорость миграции — фактор, влияющий на величину соседств. С увеличением m возрастает и Ne. При высоких значениях m большая непрерывная популяция приближается к широкой панмиксии, хотя не обязательно достигает её. При низких же значениях m большая непрерывная популяция имеет тенденцию распадаться на инбредные группы, т. е. на мелкие соседства.
Исходя из теоретических соображений, можно считать, что если величина соседства невелика, то в большой непрерывной популяции возможна значительная локальная дифференциация. Представим себе непрерывную свободно скрещивающуюся полиморфную популяцию, в которой отбор отсутствует. Если Ne = 1000 или более, то следует ожидать незначительной региональной дифференциации на расы (не считая эффектов отбора). Но если Ne=100, то возможно образование региональных рас; если же Ne=10, то могут образоваться локальные расы без участия отбора (Wright, 1943*).
| Организм | Ne | Источник данных* |
| Плодовые мухи | ||
| 4922; 6436 | ||
| D. pseudoobscura Колорадо | 3239; 6479 | » » |
| D. nigraspiracula | 21 256 55 483 145 780 | Johnston, Heed, 1973 |
| Птицы | ||
| Parus major | 770; 1806 | Barrowclough, 1980 |
| Melospira Melodia | 892 | » » |
| Larus argentatus | 1 453 | » » |
| Puffinus puffinus | 52 157 | » » |
| Грызуны | ||
| Peromyscus polionotus | 240–360 | Schields, 1982 |
| Mus musculus | 12 | » » |
| Ящерица и лягушка | ||
| Sceloporus olivaceus | 125–270 | |
| Rana pipiens | 112–446 | Schields, 1982 |
| Улитка | ||
| Cepaea Nemoralis | иногда до 12 000 | Greenwood, 1974; 1976 |
| Травянистые растения | ||
| Phlox pilosa | 75–282 | Levin, Kerster, 1968, 1974 |
| Liatris aspera | 30–191 | » » |
| Lithospermum caroliniense | ~4 | Kerster, Levin, 1968 |
| Lupinus texensis | 42–94 | Schaal, 1980 |
В случае небольших соседств в большой непрерывной популяции со свободным скрещиванием предсказанный результат был подтвержден на компьютерной модели (Turner et ai,, 1982*). Представим себе гипотетическую популяцию из 10 000 однолетних растений, полиморфную по двум аллелям одного гена при отсутствии селективных различий между трёмя генотипами. В исходном поколении все три генотипа распределены случайным образом в большой непрерывной популяции. Растения свободно скрещиваются, но опыляются ближайшими соседями, так что величина соседства мала (Ne = 4.4—5.2).
Если соседство сохранит такую величину и структуру размножения в течение 100 или нескольких сотен поколений, то первоначально гомогенная популяция разделится на пятна сходных генотипов или локальных рас, как это показано на рис. 9.1 (Turner et al, 1982*).
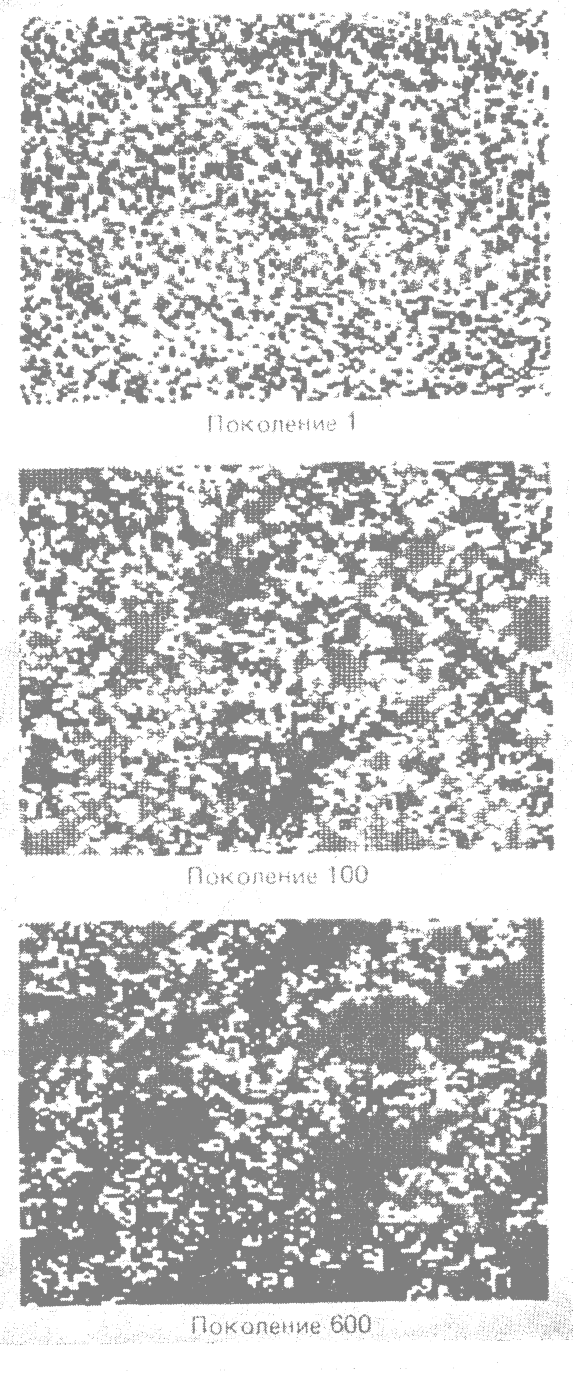 |
Рис. 9.1. Пространственное распределение трёх генотипов в разные периоды времени в модельной популяции растений. Чёрный цвет — генотип AA; серый — генотип aa; белый — генотип Aa. Объяснения смотри в тексте. (Turner, Stephens, Anderson, 1982*) |
Оценка величины соседств
Неоднократно предпринимались попытки измерить эволюционно значимую величину соседства в реальных популяциях различных организмов. Некоторые характерные оценки приведены в табл. 9.1. Как можно видеть, в популяциях некоторых видов дрозофил, птиц и улиток соседства имеют достаточно большую величину (Ne>1000).
У разных других видов соседства относительно невелики и соответственно их популяции довольно далеки от широкой панмиксии. Так, у техасской заборной игуаны (Sceloporus olivaceus) Ne=225—270, но, вероятно, ближе к 225, а область, занимаемая соседством, представляет собой круг, радиусом 213м (Kerster, 1964*). У флокса (Plox pilosa) Ne = 75—282, а область, занимаемая соседством, имеет 4.4—5.2 м в диаметре (Levin, Kerster, 1968; 1974*).
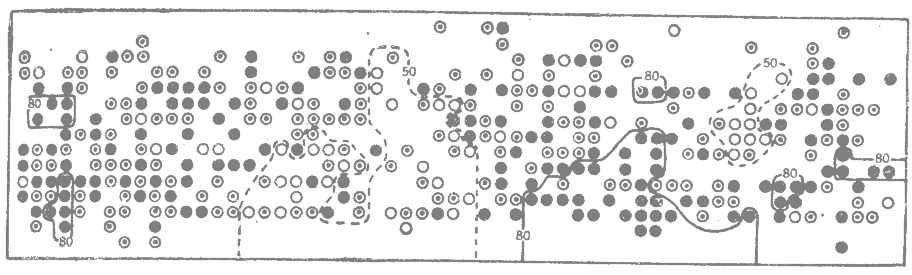 |
Рис. 9.2. Пространственное распределение трёх генотипов по одному ферментному локусу (эстераза-3) у домовой мыши (Mus musculus) в одной половине амбара. Кружками обозначено местонахождение отдельных мышей в момент поимки (черные кружки — гомозиготы -SS; белые кружки — гомозиготы ММ; кружки с точкой—гетерозиготы MS). (Selander, 1970.*) |
В некоторых диких популяциях домовой мыши (Mus musculus) воспроизводящиеся группы представляют собой очень маленькие полуизолированные субпопуляции, состоящие из нескольких особей (Selander, 1970; DeFries, McClearn, 1972; Shields, 1982*). Популяция диких мышей, обитавшая в одном амбаре в Техасе, была полиморфна по гену фермента эстеразы-3. По одной стороне этого амбара были расставлены мышеловки, расположенные в виде решетки, и пойманных в них мышей обследовали на наличие этого генетического маркера. Локализация в момент поимки особей, различающихся по генотипу в отношении эстеразы-3, показана на рис. 9.2. Особи, имеющие один и тот же генотип, часто образуют скопления на небольших участках (Selander, 1970*).
Однако у домовых мышей не всегда наблюдается такая картина: Бейкер (Baker, 1981*), работавший с другим генетическим маркеров, обнаружил между популяциями мышей на фермах в Нью Йорке значительную миграцию и обмен генами.
Очень сильное отклонение от широкой панмиксии выявлено у Lithospermum caroliniense (Boraginaceae). У этого растения соседства занимают области диаметром 4 м при эффективных величинах примерно 4 индивидуума (Kerster, Levin, 1968*).
Если воспроизводящиеся группы очень невелики, подобно описанным выше для некоторых природных популяций, то создаются условия, благоприятные для дифференциации на локальные расы без всякого участия отбора.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |