| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 37
Биологические аспекты
Система классификации приматов
Отряд приматов отличается своим разнообразием, а главное — диапазоном этого разнообразия. Расстояние, разделяющее такие крайние типы, как лемур и человекообразная обезьяна или человек, огромно. Разнообразие в других крупных отрядах млекопитающих, как, например, копытные, в общем не выходит за пределы одной грады, но у приматов оно соответствует ряду град.
О степени разнообразия в отряде приматов ясно свидетельствует табл. 37.1. Отряд этот делится на 4 подотряда, 12 семейств, 47 родов и 177 видов. Типы животных, приведённые в табл. 37.1, отражают диапазон разнообразия, в котором можно выделить четыре грады: полуобезьяны (лемуры, лори, долгопяты), низшие обезьяны, человекообразные обезьяны, человек.
Приматы, занимающие низший конец этого диапазона град, приближаются к несколько более примитивному отряду насекомоядных. Промежуточными формами между двумя отрядами служат тупайи, которых иногда относят к приматам. Приматы, возможно, произошли от насекомоядных. Остатки насекомоядных обнаруживаются в палеонтологической летописи вплоть до мелового периода, а остатки приматов — вплоть до палеоцена.
Ископаемые остатки свидетельствуют о том, что первыми приматами были полуобезьяны, что они были широко распространены и что в палеоцене и эоцене у них происходила адаптивная радиация. В эоцене и олигоцене некоторые боковые линии вымерли, а другие сохранились до настоящего времени. Выжившие главные ветви соответствуют различным подотрядам и надсемействам, перечисленным в табл. 37.1 (Simpson, 1967*).
Гоминоиды
Hominoidea — главная ветвь приматов, обособившаяся в олигоцене. Среди её вымерших ископаемых представителей можно назвать олигоценовых Propliopithecus и Aegyptopithecus, миоценовых Proconsul, Ramapithecus, Sivapithecus и Dryopithecus и плиоценового Gigantopithecus (Pilbeam, 1985; Campbell, 1985, Fleagle et al, 1986*).
Таблица 37.1. Система классификации современных приматов (по данным Walker, 1975; Colbert, 1980; Eisenberg, 1981*) | ||
| Крупные группы | Роды и виды | Географическое распространение |
Подотряд Lemuroidae | ||
Сем. Lemuridae Лемуры | 5 родов (Lemur и др), 16 видов | Мадагаскар |
Сем. Indridae Индри | 3 рода (Indri и др), 4 вида | Мадагаскар |
Сем. Daubentoniidae Айе-Айе | 1 род (Daubentonia), 1 вид | Мадагаскар |
Сем. Lorisidae Лори | 4 рода (Loris и др), 5 видов | Тропическая Африка и острова Юго-Восточной Азии |
Сем. Galagidae | 2 рода (Galago и др), 6 видов | Тропическая Африка |
Подотряд Tarsioidea | ||
Сем. Tarsiidae Долгопяты | 1 род (Tarsius), 3 вида | Острова Юго-Восточной Азии |
Подотряд Platyrrhini | ||
Сем. Sebidae Обезьяны Нового Света | 10 родов (Cebus, Alouatatta и др), ~30 видов | Тропические области Америки |
Сем. Callitrichidae Игрунки | 5 родов (Callithrix и др) ~35 видов | Тропические области Южной Америки до Панамы |
Подотряд Catarrhini | ||
Надсемейство Cercopithecoidea | ||
Сем. Cercopithecoidae Мартышки, павианы | 10 родов (Macaca, Papio и др), 66 видов | Африка, острова Юго-Восточной Азии и Япония |
Надсемейство Hominoidea | ||
Сем. Hominoidae Гиббон, Сиаманг | 2 рода (Hylobates, Symphalangus), ~7 видов | Юго-Восточная Азия и острова этого региона |
Сем. Pongidae Человекообразные обезьяны | 3 рода (Pongo, Pan, Gorilla), 3 (или 4) вида | Сумматра и Калимантан (Pongo); тропическая Африка (Pan и Gorilla) |
Сем. Gominidae Человек | 1 род, 1 вид | Весь земной шар |
Современных представителей гоминоидов делят на три группы: гиббоны, человекообразные обезьяны и человек. Гиббонов теперь выделили в самостоятельное семейство — Hilobatidae. В систематике человекообразных обезьян и человека в настоящее время существуют два альтернативных направления. Традиционная система относит человека и человекообразных обезьян к двум разным семействам — Hominidae и Pongidae. Близкое родство между человеком и африканскими человекообразными обезьянами, обнаруженное на молекулярном уровне (см. гл. 32), привело к объединению этих родов в одно подсемейство (Hominidae) расширенного семейства Hominidae (Miyamoto et al., 1988*). Такой подход оправдан молекулярными данными, но его сторонники пренебрегают морфологическими, экологическими и поведенческими признаками. Я считаю, что для таксономических целей различия между человеком и человекообразными обезьянами в морфологии, экологии и поведении важнее их сходства по некоторым отдельным макромолекулам. Поэтому в данной книге сохранена традиционная система (табл. 37.1).
| Род и вид | Эпоха | Возраст (приблизительный) млн. лет до н.э. |
Australopithecus | От плиоцена до плейстоцена | |
A. afarensis | 3.75—2.5 | |
A. africanus | 3.0 —2.5 | |
A. boisei | 2.5 —1.2 | |
A. robustus | 2.1 —1.5 | |
Homo | От плейстоцена до современной эпохи | |
H. habilis | 2.1 —1.5 | |
H. erectus | 1.5 —0.3 | |
H. sapiens | 0.3 —современная эпоха | |
От человекообразных обезьян, по крайней мере в начале плиоцена, а возможно, и в позднем миоцене, отделилась антропоидная линия, состоящая из двух родов — Australopithecus и Homo (Wolpoff, 1980; Pilbeam, 1984; Campbell, 1985*). Возраст этих двух родов и относящихся к ним видов указан в табл. 37.2. Эти данные непрерывно пополняются по мере обнаружения новых ископаемых остатков и применения новых методов датировки.
Плио-плейстоценовые Australopithecinae известны главным образом из Южной, и Восточной Африки. По строению черепа, лица и объему головного мозга они были сходны с человекообразными обезьянами (рис. 37.1), но по признакам зубов, вертикальному положению тела и передвижению на двух ногах близки к человеку. Различают 4 вида: изящные Australopithecus afarensis и A. africanus и массивные A. boisei и A. robustus; два последних представляют собой, возможно, географические расы одного вида. A. afarensis — более старая предковая форма; A. africanus появился позднее. Эти два более крупных вида, по-видимому, дивергировали от ветви двух изящных видов. Все Australopithecinae вымерли до конца плейстоцена.
Род Homo относится к другой линии, представители которой обладают большим головным мозгом. Различают три вида: Н. habilis, H. erectus и Н. sapiens (рис. 37.1). Все три жили в Африке в плейстоцене. H. habilis — древнейший представитель рода Homo и является переходной формой между Australopithecus и Homo. H. habilis, H. erectus и H. sapiens, очевидно, образуют ряд последовательных видов, переходивших один в другой с течением времени. Принимаемые в настоящее время оценки их возраста приведены в табл. 37.2.
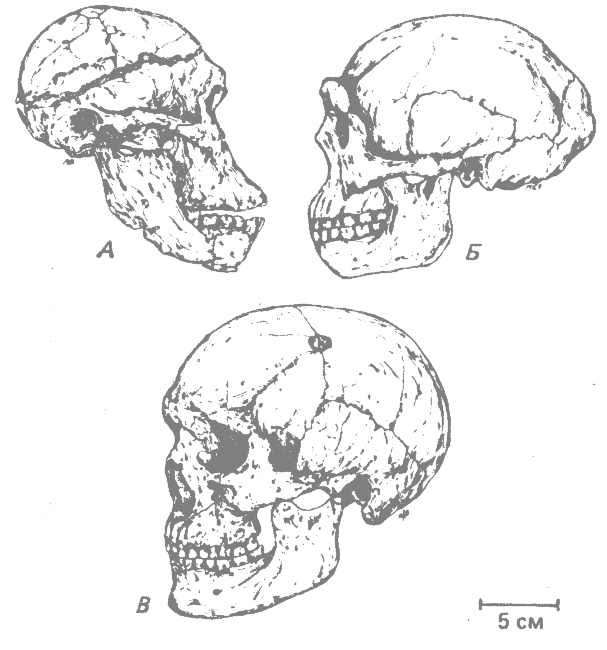 |
Рис. 37.1. Черепа ископаемых антропоидов. Все рисунки выполнены в одинаковом масштабе. A. Australopithecus africanus, нижний плейстоцен, Южная Африка. Б. Homo erectus, средний плейстоцен, Пекин. В. Homo sapiens, верхний плейстоцен, Пекин. (Из Brace, Nelson, Korn, 1971.*) |
Homo перекрывался во времени с Australopithecus, H. habilis сосуществовал с A. boisei и A. robustus (Klein, 1977*). Однако на каждом отдельном временном уровне всегда существовал лишь один вид Homo. Несмотря на то что род Homo близок к Australopithecus и что Н. habilis, по всей вероятности, образует связующее звено между ними, вопрос о точном строении филогенетического древа антропоидов вызывает большие разногласия и породил множество самых разных гипотез.
Наследие древесного образа жизни
Приматы обитают преимущественно в областях с теплым климатом (см. табл. 37.1); они живут главным образом на деревьях, а такие наземные формы, как павианы и человек, составляют исключение; приматы в основном всеядные. Древесный образ жизни привел к развитию ряда приспособлений, которые впоследствии использовал человек.
Млекопитающие, живущие на деревьях, должны уметь точно оценивать расстояния, фокусировать взгляд на определённых объектах, удерживаться на ветках и схватывать предметы, годные в пищу. В соответствии с этими требованиями у приматов имеются такие приспособительные особенности, как хорошо развитое зрение, хватательные передние конечности с противопоставляющимся большим пальцем и хвост, служащий балансиром.
Острота зрения приматов обеспечивается сочетанием ряда признаков. Глаза хорошо развиты. Они направлены вперед, что создаёт возможность одновременной фокусировки обоих глаз на одном и том же объекте и оценки расстояния по параллаксу. Наличие желтого пятна на сетчатке улучшает фокусировку. Наконец, приматы обладают цветовым зрением, которого лишены многие группы млекопитающих и которое, вероятно, способствует улучшению объемного зрения.
Зрение служит в свою очередь одним из существенных компонентов высшей нервной деятельности приматов, которая также представляет собой сложную черту. Важное значение для высшей нервной деятельности имеет прежде всего головной мозг, хорошо развитый у всех высших приматов. Однако то, что этот мозг может узнать о внешнем мире, в значительной степени зависит от сочетания глаз и передних конечностей приматов. Другие органы чувств (в особенности слух) играют дополнительную роль.
Высшая нервная деятельность приматов основана главным образом на использовании глаз, способных к одновременной фокусировке, и хватательных или осязательных кистей. У других животных эта деятельность имеет иную основу. У собак, например, она зависит прежде всего от сочетания способных к фокусировке глаз, обоняния и очень чувствительных ушей; у пчел —от сочетания сложных глаз, цветового зрения и гибких антенн, воспринимающих запахи.
Сочетание глаз — рука было и остается той основой, которая давала возможность приматам, предшествовавшим антропоидам, концентрировать внимание на отдельных объектах. У антропоидов это же самое сочетание легло в основу изготовления орудий и их использования, экспериментирования и решения задач.
Наследие наземного образа жизни
Человекообразные обезьяны обитают в лесах и ведут, во всяком случае частично, древесный образ жизни. Гиббоны проводят большую часть своей жизни на деревьях и передвигаются при помощи брахиации и прыжков. Шимпанзе и гориллы — полуназемные животные и передвигаются по земле, опираясь на подошвы ступней и тыльную сторону пальцев передних конечностей.
В отличие от этого Australopithecus и Homo были и остаются обитателями светлых лесов, саванн и равнин. Способом передвижения у них служит ходьба или бег на двух ногах. Хождение на двух ногах и её следствие — вертикальное положение тела — наиболее яркие отличительные признаки антропоидов. Это разделение между гоминидами и понгидами прослеживается менее чётко в случае Australopithecus afarensis и Homo habilis, которые, хотя и ходили на двух ногах, но, вероятно, вели в значительной степени древесный образ жизни (Susman, Stern, 1982; Lewin, 1983*).
В середине и в конце третичного периода протяженность лесных местообитаний постепенно сокращалась, уступая место степям. В результате изменения климата и растительности возникали новые местообитания. Заселение нового равнинного местообитания, возможно, определило дивергенцию примитивных антропоидов от их предков — человекообразных обезьян, обитавших в лесах. Передвижение на двух ногах и вертикальное положение тела были частью того сочетания адаптивных признаков, которое сделало антропоидов приспособленными к жизни в саваннах и на равнинах. Большая часть данных о дальнейшей эволюции антропоидов в плейстоцене, которыми мы располагаем, указывает на всё большее выпрямление тела при откинутой назад шее и голове и на сопровождавшее его улучшение походки.
Передвижение на двух ногах имело важное следствие: оно освободило руки, которые могли теперь заняться изготовлением и использованием орудий и оружия. Самые первые каменные орудия найдены в плиоцене, а несколько более сложные — в раннем плейстоцене (Pilbeam, 1984; Susman, 1988*). Умение обращаться с предметами — важная составная часть высшей нервной деятельности древесных приматов, как это уже отмечалось. А теперь с переходом от древесного образа жизни к наземному и с освобождением рук от участия в передвижении манипуляция предметами могла развиваться в полной мере. Первым результатом этого было развитие простейших ремесел.
(Campbell, 1985*) | ||
| Вид | Средняя ёмкость, см³ | Диапазон, см³ |
Pan troglodytes | 383 | 282—500 |
Australopithecus afarensis | 418 | 385—450 |
A. africanus | 452 | 428—500 |
Homo habilis | 638 | 509—752 |
H. erectus | 930 | 727—1225 |
H. sapiens | 1350 | ~1000—2000 |
Развитие высшей нервной деятельности у приматов имеет большую адаптивную ценность, и ему, несомненно, благоприятствует естественный отбор. У приматов, не относящихся к антропоидам, выявляется эволюционное направление в сторону развития высшей нервной деятельности, достигающей очень высокого уровня у шимпанзе и орангутана и ещё более высокого — у антропоидов.
Среди антропоидов, если судить по объему черепа, Homo erectus обладал более развитым интеллектом, чем австралопитек (см. рис. 37.1). Диапазоны колебаний емкости черепной коробки у этих двух форм приведены в табл. 37.3.
Весьма вероятно, что более высокое развитие интеллекта и более совершенные орудия и оружие у Homo erectus или у его предков дали ему преимущество перед австралопитеками в прямой и косвенной конкуренции между этими двумя группами в плейстоцене. Во всяком случае, австралопитеки вымерли, а H. erectus выжил и позднее эволюционировал в H. sapiens.
Расовая изменчивость
Для современной исторической эпохи характерно распространение Homo sapiens по всему Земному шару и его дифференцировка на ряд географических и бесчисленное множество локальных рас. Гарн (Garn, 1961*) различает следующие географические расы человека:
Азиатско-монголоидная
Американские индейцы
Индийская (Индия, Пакистан и т. д.)
Европеоидная белая
Африканская негроидная
Австралийская негроидная
Меланезийская
Микронезийская
Полинезийская
Недавно Кавалли-Сфорца и др. (Cavalli-Sforza et al., 1988*) на основании совокупности данных по генетическому полиморфизму, лингвистических и археологических данных установили порядок ветвления в филогении человека; составленный ими перечень рас несколько отличается от приведенного выше. I. Дивергенция на африканскую негроидную и другую расу-1. II. Дивергенция другой расы-1 на северо-евразийскую и другую расу-2. III. Дивергенция северо-евразийской расы на кавказскую и другую расу-3. IV. Дивергенция другой расы-3 на северо-восточную, азиатскую и арктическую американскую. V. Тем временем другая раса-2 дивергировала на юго-восточную азиатскую и расы островов Тихого океана, с одной стороны, и новогвинейскую и аборигенную австралийскую — с другой.
Это типичные географические расы, подобные расам у других видов млекопитающих. Некоторые расовые признаки размеров тела и его пропорций, пигментации и т. п. коррелируют с климатом (см. Coon, 1955; Garn, 1961*). Хорошо известно, например, что расовая изменчивость цвета кожи коррелирует с количеством солнечного света в данной области.
Цвет кожи у человека регулирует синтез витамина D. Определенное количество этого витамина необходимо для нормального роста; недостаток его вызывает рахит, а избыток — обызвествление костей. Витамин D синтезируется в коже, и его синтез регулируется солнечным светом: чем больше солнца, тем больше витамина D.
Темнокожие расы живут в тех областях, где много солнца. Темные пигменты, содержащиеся в коже, плохо пропускают солнечные лучи и предотвращают синтез избыточных количеств витамина D. Напротив, белая раса, изначально обитавшая в северных частях Европы, не могла бы синтезировать достаточное количество витамина, не имея светлой кожи, которая пропускает максимум солнечных лучей. Но в жарких странах белая кожа поглощает слишком много солнечного света, и поэтому представители белой расы, оказавшись в таком климате, покрываются загаром, что представляет собой фенотипическую модификацию, препятствующую избыточному поглощению света. Что касается представителей черных рас, то, если бы им пришлось жить на севере Европы в естественных условиях, они, вероятно, погибли бы от рахита. Таким образом, различия в цвете кожи представляют собой адаптивный признак, зависящий от интенсивности солнечного излучения в данной местности (Loomis, 1967*).
Подобные факты свидетельствуют о роли естественного отбора в развитии по крайней мере некоторых расовых признаков Homo sapiens. Локальная расовая изменчивость иногда носит случайный характер, позволяя предполагать, что в её возникновении участвуют также дрейф генов или отбор и дрейф (см. гл. 16). В других случаях характер расовой изменчивости у человека указывает на регулирующее влияние миграции и потока генов (см. гл. 20).
Заключение
Некоторые стороны эволюции антропоидов, обсуждавшиеся в этой главе, представляют собой продолжение эволюции приматов, не относящихся к антропоидам. Так, мы обнаружили эволюционные направления, приведшие к использованию руки, развитию высшей нервной деятельности, изменению положения тела т. д. Мы видим указания на квантовый сдвиг к новому способу передвижения в местообитании нового типа, на межвидовой отбор между отдельными линиями антропоидов; образование географических рас у выжившего и широко распространившегося вида Homo sapiens.
Все эти аспекты эволюции человека представляют собой часть более обширной картины эволюции органического мира. Их можно объяснить как результат действия эволюционных факторов и сил, описанных в более ранних главах этой книги.
В связи с этим следует указать, что за 10 000 лет человек может пройти через 600 поколений. За этот недолгий период отбор или отбор в сочетании с дрейфом генов могли вызвать существенные эволюционные изменения. А за более длительные периоды, на протяжении которых, согласно ископаемым данным, протекала эволюция антропоидов, могли, конечно, произойти гораздо более глубокие эволюционные изменения.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |