| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 36
Вымирание
Вымиранию подвержены группы разных размеров и разных рангов. Нам представляется полезным выделить пять модальных уровней вымирания: 1) вымирание вида на большей часта его ареала; 2) вымирание вида в целом; 3) вымирание филетических групп относительно низкого таксономического ранга, например родов или семейств; 4) вымирание групп высокого ранга, таких как отряды и классы: 5) массовое вымирание, охватывающее много разных групп в данную эпоху. В этой главе будут приведены примеры, относящиеся к разным уровням вымирания.
В общем, группы высокого таксономического ранга сохраняются в течение более длительного времени, чем филетические группы низких рантов, а последние в свою очередь существуют в среднем дольше, чем виды. Вымирание уготовано огромному большинству видов, но царства и типы, как правило, бессмертны.
Рассматривая вымирание надвидовых групп, удобно провести дальнейшее разделение. Некоторые такие группы вымирают, не оставляя филогенетических потомков, как, например, птерозавры. Другие группы вымирают, но дают начало потомкам, образующим отдельную группу; так, группа рептилий Therapsida вымерла в середине мезозоя, но дала начало млекопитающим.
Мы можем установить причину вымирания, если она представляет собой прямое или косвенное следствие деятельности человека в историческое время. Известно, например, что странствующий голубь, бескрылая гагара и многие другие виды птиц (см. Dorst, 1974*), а также бизон на большей части своего некогда обширного ареала (см. Hornday, 1889*) вымерли по вине человека.
В тех случаях, когда вымирание происходило в геологическом прошлом, причины его установить трудно. Неясны причины, как мы увидим, исчезновения в четвертичном периоде видов, особенно хорошо представленных в палеонтологической летописи, и неудивительно, что по мере продвижения вглубь геологического времени и перехода от видового уровня к крупным группам и массовым вымираниям, проблема всё усложняется*.
Некоторые плейстоценовые и постплейстоценовые копытные
События, приведшие к почти полному истреблению крупного травоядного животного новым хищником — белым человеком — иллюстрриуются хорошо документированной и печальной историей бизона (Bison bison), изложенной Хорндеем (Hornaday, 1889*).
Первоначальный ареал бизона простирался от Невады и Орегона до Джорджии и Пенсильвании и от северной части Мексики до северо-западных территорий Канады (рис. 36.1, А). По имеющимся оценкам в начале появления европейских переселенцев по этой огромной области бродило 50 млн. бизонов. Дикие бизоны существовали в восточной части США до 1760— 1800 гг., на Среднем Западе — до 1810—1825 гг., в Техасе — до 1837 г., а в Орегоне — до 1838 г.
Охота и разрушение местообитаний, производимые белым человеком, привели к постепенному исчезновению бизона из центральных и восточных частей США и из южных и крайне западных частей его первоначального ареала, а к 1870 г. этот ареал был сведен до небольшой области на Великих равнинах (рис. 36.1, Б). Примерно в это же время (1870 г.) человек перешел от сравнительно ограниченной охоты на бизонов к их систематическому истреблению. Спустя 10 лет, в 1889 г., ареал бизона состоял из одного большого и пяти маленьких разобщённых участков (рис. 36.1, В). К концу 1888 г. в этих шести местностях обитало примерно всего 830 диких бизонов (рис. 36.1, Г), а в 1889 г. их число сократилось до 541 особи (Walker, 1975*). Тогда специалисты по охране природы занялись спасением бизонов от полного вымирания, и в настоящее время в нескольких Национальных парках и в парках некоторых штатов существуют охраняемые стада этого вида животных.
Ареал Megaloceros giganteus — гигантского оленя, жившего в плейстоцене и в начале голоцена, — простирался от Ирландии через всю Европу до Сибири и Китая, а на юге — до Северной Африки. Он процветал в Ирландии в течение периода, непосредственно следовавшего за отступлением льдов в конце последнего оледенения, и вымер там примерно 11 000 лет назад. Возможно, что в некоторых частях материковой Евразии он дожил до начала голоцена (Gould, 1974*).
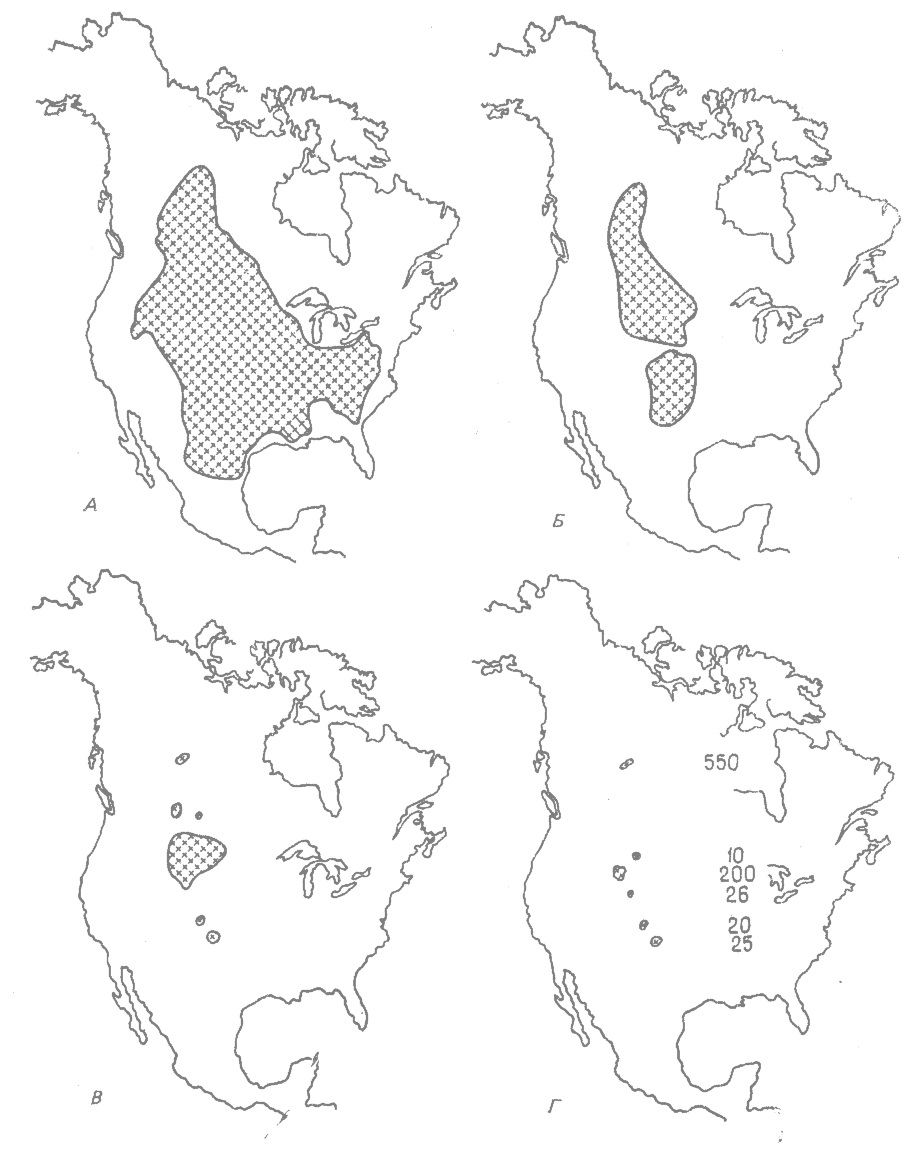 |
Рис. 36.1. Сокращение ареала бизона с начала XVIII до конца XIX в. А. Первоначальный ареал. Б. Ареал в 1870 г. В. Ареал в 1880 г. Г. Ареал и число особей к концу 1888 г. (Из Hornaday, 1889.*) |
Большая часть ископаемых остатков Megaloceros giganteus найдена в озерных отложениях и торфяных болотах Ирландии. Это был гигантский олень высотой около двух метров на уровне плеч. Самцы обладали огромными рогами весом до 25 кг и размахом примерно до 3 м. Это максимальные размеры; подробные результаты измерений приводит Гулд (Gould, 1974*). Ни у одного другого представителя семейства оленей не было таких крупных рогов.
Многие авторы отмечали, что большие и тяжелые рога Megaloceros giganteus, которые олень ежегодно сбрасывал и которые отрастали вновь, вероятно, создавали тяжелую физическую нагрузку и физиологически истощали животных. Среди выдвигавшихся ранее причин вымирания М. giganteus особенно большое внимание уделялось тем неудобствам, которые, возможно, терпели животные из-за таких больших рогов; эти рога просто считали неадаптивными. Вполне возможно, что рога были фактором, способствовавшим вымиранию животного, но, как указывал Симпсон (Simpson, 1949, 1967*), нет причин считать, что эти рога не имели адаптивного значения в тот период, когда они развились и когда вид процветал.
Симпсон (Simpson, 1949, 1967*) указывал далее, что величину рогов М. giganteus следует рассматривать не в качестве изолированного признака, а как результат соотношения скоростей роста разных частей тела в процессе филогенеза. Среди представителей семейства оленей филогенетическая скорость роста рогов значительно превышает скорость роста тела, так что самые крупные виды оленей должны иметь пропорционально самые большие рога. У М. giganteus с его большим телом рога были примерно такими, каких и следовало бы ожидать у очень крупного оленя.
Следовательно, возможно, что отбор благоприятствовал увеличению размеров тела М. giganteus, а большие рога развились в качестве побочного эффекта (Simpson, 1949, 1967*). Увеличение размеров тела — обычная тенденция в эволюции млекопитающих, дающая им ряд преимуществ (см. гл. 28). Возможно также, что половой отбор благоприятствовал увеличению рогов у М. giganteus, потому что это давало преимущество самцам при спаривании, а побочным эффектом были большие размеры тела (Gould, 1974*).
Причины вымирания М. giganteus, по крайней мере в Ирландии и, вероятно, в других местах, можно искать также и в других аспектах его жизни. Он, по-видимому, обитал на открытых поросших травой местах, перемежавшихся с участками леса. Такого рода растительность достигла расцвета в один из периодов потепления к концу последнего оледенения, а именно в это время и процветал М. giganteus. Несколько позднее в тот же период степи сменились лесами, и М. giganteus, который не был лесным оленем, вероятно, не смог выжить в густом лесу. Неблагоприятные для этого оленя изменения климата и растительности в период последнего оледенения, по-видимому, и были причиной его вымирания (Gould, 1973, 1974*).
Лошадь (Equus) вымерла в Северной Америке в начале голоцена, примерно 8000—10 000 лет назад. Как уже говорилось в гл. 28, эволюция Equus происходила главным образом в Северной Америке, откуда животное мигрировало на три других материка. В плейстоцене в Северной Америке существовали огромные табуны лошадей, принадлежавших, вероятно, к нескольким видам Equus. Затем род Equus полностью вымер в Северной Америке (Simpson, 1951* и личное сообщение).
Спустя много тысячелетий, в 1519 г., испанцы ввезли в Северную Америку европейскую домашнюю лошадь (Equus caballas). Интродукция оказалась очень успешной, и домашняя лошадь образовала дикие табуны на древней родине своих предков.
Причины вымирания Equus в Северной Америке неизвестны. И тайна эта усугубляется тем, что Equus удалось вновь успешно закрепиться в диком состоянии на этом материке спустя несколько тысяч лет после того, как она там вымерла.
Высказывались различные предположения о причинах этого вымирания, но все они малоубедительны. Не оказалось ли аборигенная североамериканская лошадь жертвой оледенения? Но ведь Equus пережила плейстоцен, а кроме того, она была распространена и за пределами границы льдов. Была ли лошадь истреблена первыми американскими индейцами, с которыми она жила в одно время? Такую возможность исключить нельзя. Но ведь индейцы охотились на бизона и тем не менее не истребили его. Возможно также, что лошадь погибла от какого-то эпидемического заболевания. Однако в палеонтологической летописи не сохранилось ничего, что свидетельствовало бы о такой эпидемии (Simpson, 1951*).
Те же самые события происходили параллельно в Южной Америке. Здесь род Equus также процветал в плейстоцене и вымер в начале голоцена; затем в историческую эпоху лошадь была интродуцирована из Европы, после чего она одичала. Причины её вымирания в Южной Америке также неизвестны (Simpson, 1951*).
Саблезубые кошки
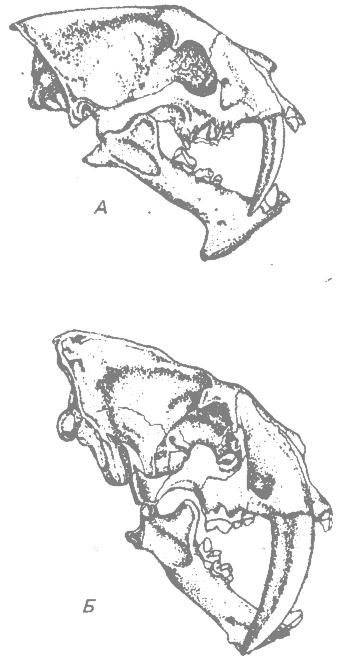
Саблезубые кошки (подсемейство Machairodontinae) — одна из двух главных ветвей сем. Felidae; другую ветвь образуют настоящие кошки (подсемейство Felinae). Саблезубые отличались от настоящих кошачьих крупными размерами верхних клыков и другими, связанными с этим, признаками челюстей и зубов (рис, 36.2).
Саблезубые кошки жили в Северной Америке и Евразии в течение почти всего третичного периода, начиная с олигоцена, а в плейстоцене проникли в Южную Америку. Увеличение размеров резцов и общих размеров тела происходило на протяжении всего третичного периода. Черепа Hoplophoneus — характерного олигоценового представителя группы в Северной Америке, — и Smilodon — более крупного плейстоценового представителя, жившего в Северной и Южной Америке, показаны на рис. 36.2, А и Б. В плейстоцене вся группа саблезубых кошек вымерла (Romer, 1966*).
 |
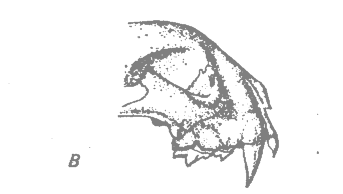 |
Рис. 36.2. Черепа двух саблезубых кошек (А и Б) и одной настоящей кошки (В). A. Hoplophoneus; олигоцен, длина черепа 15.9 см. Б. Smilodon; плейстоцен, длина черепа 30.5 см. В. Pseudaelurus; плиоцен, длина черепа 14.6 см.. Показаны клыки для сравнения. (Из Romer, 1966*, с изменениями.) |
В одно время было принято считать причиной вымирания саблезубых кошек большие размеры их клыков, которые, подобно рогам Megaloceros giganteus, рассматривали как неадаптивный признак; высказывалось даже мнение, что они мешали кусанию. На самом деле крупные клыки не мешали кусанию. Челюсти саблезубых кошек могли широко раскрываться и плотно сжиматься.
Возможно, что саблезубые кошки были специализированы к охоте на таких крупных, медленно передвигавшихся толстокожих растительноядных животных, как мастодонты и ленивцы. Их длинные клыки при разомкнутых челюстях могли служить эффективными органами для закалывания и разрывания добычи; в этом им помогали мощные шея и плечевой пояс.
В течение плейстоцена рады толстокожих растительноядных значительно поредели. Снижение численности, а затем и вымирание добычи этого типа сократило или закрыло адаптивную зону саблезубых кошек. Тем временем у представителей подсемейства Felinae в семействе кошачьих совершенствовался альтернативный способ охоты на животных, которые быстро передвигаются, — нападение из засады и преследование. Поскольку в плейстоцене адаптивная зона самих саблезубых кошек стала закрываться, а альтернативные адаптивные зоны были уже заняты, этим кошкам пришел конец. Они представляют собой яркий пример группы, которая была успешно специализирована на одной стадии эволюции, но на более поздней стадии, когда условия изменились, оказалась «чересчур специализированной» (Simpson, 1953*).
(Kurten, Andercon, 1980*) | ||
| Эпоха | Век | Датировка (приблизительная) |
| Ранчолабрейский | ||
| Ирвингтонский | ||
| Бланкский | ||
Массовое вымирание североамериканских млекопитающих в позднем кайнозое
Конец плиоцена и плейстоцен характеризуются тектонической активностью, ухудшением климата, изменениями в географическом распространении организмов и обновлением биоты. Среди млекопитающих которые были доминирующими наземными животными в течение всего третичного периода, происходило массовое вымирание. При этом фауна северных континентов пострадала сильнее, чем фауна тропических и южных областей суши. Здесь мы сосредоточим внимание на событиях в Северной Америке*. Хронология соответствующего периода времени на этом континенте представлена в табл. 36.1.
Известно, что в позднем кайнозое в Северной Америке вымерло 338 видов млекопитающих, представленных в палеонтологической летописи. Некоторые группы вымерли в Северной Америке, но сохранились на других континентах; например: лошади, верблюды, ламы, сайга, як, водосвинка и очковый медведь. Другие группы вымерли полностью; например: мегатерии, гигантский броненосец, глиптодонты, гигантский бобр, мастодонты, мамонты, шерстистый носорог и саблезубые кошки (Axelrod, 1967; Kurten, Anderson, 1980*).
в разное время в позднем кайнозое (Kurten, Anderson, 1980*) | |
| Век и его подразделения | Число видов |
| 12 | |
| 49 | |
| 27 | |
| 49 | |
| 1 | |
| 138 | |
| 39 | |
| 14 | |
| 33 | |
| 3 | |
| 89 | |
| 7 | |
| 21 | |
| 77 | |
| 3 | |
| 108 | |
| 3 | |
| 338 | |
| Виды, найденные в плейстоцено-голоценовых отложениях и живущие до сих пор. | 229 |
Вымирание затронуло 38 семейств, принадлежащих к 9 отрядам из 45 семейств и 12 отрядов, представленных тогда на континенте (Kurten, Anderson, 1980*).
Вымиранию млекопитающих в позднем плейстоцене уделялось много внимания как в научной, так и в популярной литературе. Необходимо представлять себе, что вымирание происходило на всем протяжении 3.5 млн. лет (Anderson, Kurten, 1980*). Распределение вымирания видов во времени показано в табл. 36.2.
Вымирание затронуло млекопитающих всех размеров. Интересно указать, что вымирание видов мелких млекопитающих достигло наивысшей скорости в бланкском веке, а вымирание крупных — в висконсинской фазе ранчолабрейского века (Kurten, Anderson, 1980*).
Учитывая большое разнообразие событий вымирания в отношении сроков, таксономического родства, общих размеров животных, жизненных циклов и экогеотрафичеоких областей, было бы, вероятно, бесперспективным искать какие-то единые особые причины (Kurten, Anderson, 1980*). Здесь необходимо рассмотреть ряд таких факторов, как изменения климата, изменения растительности, конкуренция, хищничество, заболевания и т. п. Эти факторы, вероятно, влияли на разные виды по-разному. Некоторые сочетания факторов могли оказывать на некоторые виды очень неблагоприятное или даже фатальное воздействие. Если и существовала некая единая причина, то это была, вероятно, не какая-то конкретная причина, а общее орографическое и климатическое изменение, происходившее в данный период, которое привело к сложным последствиям в различных экосистемах.
Любое существенное изменение климата или другое физическое изменение неизбежно изменяет равновесие между видами, благоприятствуя одним за счет других, и это приводит к замещению видов.
Ниже будут рассмотрены три гипотезы о причинах вымирания.
Согласно гипотезе перепромысла, которую выдвинул Мартин (Martin, 1973*), причиной вымирания крупных млекопитающих в позднем плейстоцене было их массовое уничтожение первобытным человеком. Когда примерно 11 500 лет назад первые индейцы, жившие за счет охоты, пришли в Северную Америку из Сибири, они нашли здесь обильные новые пищевые ресурсы — населявших этот континент в те времена крупных млекопитающих. Эти млекопитающие, среди которых были мастодонт, мамонт, лошадь, верблюд и бизон, прежде никогда не подвергались истреблению со стороны человека. Популяция человека быстро разрасталась и расселялась по Северной и Южной Америке благодаря обилию пищи, обеспечиваемому крупной дичью. В результате интенсивной охоты большая часть этих животных вымерла. Некоторые из них вымерли, по-видимому, примерно в то же время, когда, согласно предположениям, произошел взрыв численности первобытного человека, т. е. 10—11 тысяч лет назад (Martin, Wright, 1967; Martin, 1973*).
Первые индейцы, несомненно, оказали воздействие на местную фауну млекопитающих и вполне возможно, что они полностью истребили некоторые виды; однако сомнительно, чтобы при своем примитивном охотничьем снаряжении они могли вызвать те грандиозные события, которые постулирует гипотеза перепромысла. Указывают, что плейстоценовая фауна Австралии, которая содержала ряд крупных животных, таких, как настоящий красный кенгуру, сосуществовала с человеком не менее 7000 лет (Gillespie et al., 1978*). В Северной Америке скорость вымирания орнитофауны в позднем плейстоцене была близка к скорости вымирания млекопитающих (Grayson, 1977*).
Аксельрод (Axelrod, 1967*) выдвинул в качестве причины вымирания млекопитающих в плейстоцене новые климатические условия, в особенности холод. Для третичного периода был характерен в общем ровный климат, и млекопитающие были адаптированы к такому климату. Климат плейстоцена отличался резкими колебаниями с периодами жестоких холодов. При очень сильных холодах отдельные особи млекопитающих погибли либо просто от физиологического воздействия низких температур, либо от голода, и эта гибель отдельных животных могла в конечном счете привести к вымиранию вида в целом. Новый холодный климат оказывал, возможно, также неблагоприятные косвенные воздействия на многих млекопитающих, изменяя растительность в их местообитаниях.
С климатической гипотезой Аксельрода согласуется существование определённой зависимости между общими размерами животных и их вымиранием в плейстоцене. В эту эпоху среди мелких млекопитающих гибель была относительно невелика. Большинство мелких млекопитающих во время холодов могут укрыться в норах или гнездах. Кроме того, у мелких млекопитающих репродуктивные циклы обычно короткие и им легче адаптироваться к климату с резкими сезонными колебаниями. У многих мелких млекопитающих детеныши успевают достигнуть зрелости за короткое благоприятное время года (Axelrod, 1967*). Вымирание крупных млекопитающих, по большей части неспособных приспосабливаться таким образом к очень холодному времени года, достигло в позднем плейстоцене самого высокого уровня.
Ряду фактов климатическая гипотеза не может дать объяснения. Почему многие виды североамериканских млекопитающих вымерли только в позднем плейстоцене, а не в позднем плиоцене или раннем плейстоцене, когда, собственно, произошел переход к ледниковому климату? Климатическая гипотеза не позволяет объяснить более высокую степень вымирания мелких млекопитающих по сравнению с крупными в Бланкском веке. Другая проблема, не укладывающаяся в климатическую гипотезу, — вымирание многих крупных млекопитающих не только на северных континентах, но и в Африке, которая не испытала в полной мере воздействий ледникового периода (Simpson, личное сообщение).
Брайсон и др. (Bryson et al., 1970*) провели реконструкцию изменений климата и растительности на Великих равнинах в позднем плейстоцене. На протяжении большей части позднего плейстоцена ледяной щит ограждал Великие равнины от арктического воздуха. В этих условиях могли существовать высокие злаки и крупные травоядные. Но отступление ледяного щита в результате потепления в конце плейстоцена сняло эту преграду и открыло доступ арктическому воздуху к Великим равнинам. Холодные иссушающие ветры привели к смене высоких злаков низкими ксерофитами. Сочетание жестоких холодов и недостатка пищи могло бы объяснить исчезновение многих крупных травоядных на Великих равнинах к концу плейстоцена (Bryson et al., 1970*; М. Bolick, личное сообщение).
Динозавры
Динозавры, делящиеся на два отряда, Saurischia и Ornithischia, занимали доминирующее положение среди наземных животных на всём протяжении мезозоя. За эту эру возникло большое разнообразие форм, о чем свидетельствует разделение динозавров на 25 семейств и 218 родов (Colbert, 1961*). По своим размерам эти формы варьировали от мелких до таких гигантов, как бронтозавр и тираннозавр, а по типу питания — от травоядных до хищников. Динозавры были распространены по всему земному шару и доминировали на суше в течение более чем 100 млн. лет.
Затем в позднем мелу вся группа динозавров исчезла из палеонтологической летописи. Вымирание произошло за относительно короткий, по геологическим масштабам времени, период, но всё же исчезновение различных форм было разделено промежутками в сотни тысяч и миллионы лет (Colbert, 1961; Simpson, 1968; Simpson, Beck, 1965; Van Valen, Sloan, 1977; Sloan et al., 1986*). Возможно, что некоторые динозавры дожили до раннего палеоцена (Sloan et al., 1986*).
По вопросу о причинах вымирания динозавров существуют различные мнения. Согласно одной школе, этой причиной было изменение климата; её мы обсудим ниже. Другая школа выдвигает на первый план биотические факторы, такие, как истребление млекопитающими яиц динозавров, конкуренция со стороны млекопитающих, чрезмерное уничтожение растительности динозаврами и заболевания. Третья школа постулирует физическую катастрофу в масштабах всего земного шара; этот вопрос мы обсудим позднее.
Ван Вален и Слоан (Van Valen, Sloan, 1977*) изучали последовательную смену ископаемых организмов в Хелл-Крик-Филд (Монтана) в интервале от позднего мела до палеоцена. Это непрерывная последовательность наземных растений и животных, в которой нет разрывов. Более древняя меловая флора была субтропической; флора палеоцена соответствовала растительности умеренного пояса и содержала листопадные двудольные и папоротники. Такая растительность, характерная для умеренного пояса, пришла с более далекого севера, включилась в местную флору и вытеснила прежнюю субтропическую растительность. Этот сдвиг указывает на изменение климата от субтропического до умеренного с холодными зимами.
С субтропической флорой было связано сообщество наземных животных позднего мела, в которое входили многочисленные динозавры, другие рептилии и примитивные млекопитающие, в том числе сумчатые. Эту фауну в раннем палеоцене заменило другое сообщество животных, содержавшее примитивных плацентарных млекопитающих и некоторых рептилий, но не динозавров. Семь видов динозавров, принадлежавших к прежнему сообществу, сохранились в течение переходного периода, но постепенно встречались всё реже и реже и наконец исчезли. Исчезновение динозавров в Хелл-Криж-Филд происходило медленно и постепенно, на протяжении нескольких миллионов лет, но в последние 300 000 лет процесс этот ускорился (Van Valen, Sloan, 1977; Sloan et al., 1986*).
Изложенные здесь данные указывают на то, что главной причиной вымирания динозавров в Монтане было наступление нового климата с холодными зимами и связанная с этим растительность, характерная для умеренного пояса. Кроме того, возможно, что вновь появившиеся плацентарные млекопитающие оказались в условиях нового климата и растительности более конкурентоспособными, чем динозавры (Van Valen, Sloan et al., 1977; Sloan et al., 1986*).
Разные авторы подчёркивали роль климата, и в особенности смены ровного климата относительно неустойчивым, как главную причину вымирания динозавров в масштабе всего земного шара (Axelrod, Bailey, 1968; Spotilla et al., 1973; Benton, 1979; Hallam, 1987*). Ровный теплый климат преобладал на протяжении большей части мезозоя, и эволюция динозавров протекала в таких климатических условиях примерно 130 млн. лет. Во второй половине мелового периода произошло изменение в сторону ещё менее ровного континентального климата с ещё более резкими колебаниями температуры.
Динозавры были, вероятно, эктотермными (Benton, 1979*), а не гомойотермньми животными, как полагал Бэккер (Bakker, 1975*) без достаточных к тому оснований. В теплом климате эктотермность была бы успешной стратегией. Однако в условиях континентального климата с очень резкими различиями в крайних температурах, эктотермная рептилия должна была испытывать температурные стрессы (Spotilla et al., 1973; Benton, 1979*).
Некоторые группы рептилий мелового периода могли избегать воздействия крайне низких или крайне высоких температур, забираясь в норы или впадая в спячку (ящерицы, змеи), другие — погружаясь в воду (черепахи), а третьи — обитая в тропических или субтропических реках (крокодилы); таким образом они дожили до современной эпохи. Но наземные динозавры не могли прибегнуть к таким способам (Axelrod, Bailey, 1968*).
Массовое вымирание в позднем мелу
В позднем мелу произошло одно из крупнейших массовых вымираний всех времен. Век рептилий пришел к концу. Вымерли морские ихтиозавры и плезиозавры, летающие птерозавры и динозавры, жившие на суше.
Вдобавок к этому в морях вымерла процветавшая прежде группа моллюсков — аммонитов, а также многие фораминиферы.
Споры о причинах вымирания динозавров принимают ещё более широкие масштабы при обсуждении массовых вымираний, охвативших почти все группы животных в позднем мелу.
Астероидная теория
В настоящее время ведутся оживленные дискуссии вокруг астероидной гипотезы, которую выдвинули Альварес и др. (Alvarez, 1983; Alvarez et al., 1980, 1984*). Как свидетельствуют косвенные данные примерно 65 млн. лет назад на землю упал астероид диаметром 10 км. При его входе в верхние слои атмосферы образовалось облако пыли, сохранявшееся в течение 2 или 3 лет. В морских осадках на границе между мелом и третичным периодом образовался прослой, обогащенный иридием — металлом, относящимся к семейству платины, который редок в земной коре, но содержится в больших количествах в астероидах.
Предполагается далее, что столкновение с астероидом и его последствия были причиной массового вымирания в конце мелового периода. Это предположение основано на изменениях, обнаруженных в ископаемой биоте иридиевого прослоя. В данной вертикальной последовательности та или иная группа иногда исчезает на границе этого прослоя. Так обстоит дело с аммонитами и другими морскими беспозвоночными в Дании, фораминиферами в Италии и у берегов. Африки, радиоляриями в Карибском море и пыльцой покрытосеменных в Нью-Мексико (Alvarez, 1983; Alvarez et al., 1984; Hsu et al., 1982*).
Первый вопрос состоит в том, действительно ли имело место столкновение с астероидом. Среди геологов в настоящее время существуют большие разногласия по этому поводу. Некоторые геологи считают, что иридий не внеземного, а вулканического происхождения и что обогащенный им прослой формировался на протяжении длительного времени (Officer, Drake, 1983, 1985*). Другая проблема касается кристаллической структуры кварцевых зерен, вкрапленных в иридий. Бохор и др. (Bohor et al., 1984, 1987*) считают, что характер этой кристаллической структуры указывает на столкновение, а не на извержение вулкана, т. е. высказываются в пользу астероидной гипотезы, но некоторые другие исследователи не убеждены в этом (Hallam, 1984*). Отсутствие следов столкновения с таким крупным астероидом создаёт трудную проблему. Это, однако, можно объяснить, допустив, что астероид упал в океан. Но в таком случае так называемый «ударный» кварц с его особой кристаллической структурой оказывается слишком далеко от места падения, а это заставляет усомниться в том, что он возник в результате столкновения (Hallam, 1987*). Этот геологический вопрос до сих пор остается неразрешенным (см. Kerr, 1988*).
Следующий вопрос состоит в том, могло ли вообще столкновение с крупным астероидом вызвать массовое вымирание на всём земном шаре. Этот аспект проблемы биологи-эволюционисты в состоянии рассмотреть вполне компетентно.
Альварес (Alvarez, 1983; Alvarez et al., 1980*) предлагает следующее объяснение массового вымирания. Слой пыли, образовавшийся в результате столкновения с астероидов, преградил доступ солнечному свету и подавил фотосинтез как у наземных растений, так и у морских водорослей на год или на несколько лет. За этим последовала голодная смерть морских и наземных животных, сначала растительноядных, а затем хищников. В этой гипотезе есть несколько слабых мест.
1. Исчезновение какого-либо вида в одном или нескольких местонахождениях ископаемых не означает, что данный вид вымер. В других местах популяции этого вида могли выжить.
2. Остается неясным, мог ли постулируемый слой пыли подавить фотосинтез в такой степени, чтобы вызвать голод в широких масштабах. Облака пыли, возникавшие при очень сильных извержениях вулканов в исторические времена, ограничивали доступ солнечного света и вызывали понижение температуры в областях, удаленных от вулкана, но не приводили к полному прекращению фотосинтеза или вымиранию растительноядных видов в столь отдаленных областях (см. Stommel, Stommel, 1979; Stothers, 1984*).
3. Астероидная гипотеза не позволяет объяснить, почему некоторые группы вымерли, а другие выжили. В этом суть проблемы, как указывали также Симпсон (Simpson, 1968*), Майр (Mayr, 1982*) и Колберт (Colbert, личное сообщение).
4. Астероидная гипотеза требует допустить, что все вымирания произошли за короткое время, измеряемое годами или десятилетиями, непосредственно следовавшими за катастрофой. Одновременность вымираний не была доказана. Как свидетельствуют имеющиеся данные некоторые группы, в том числе динозавры и фораминиферы, исчезали постепенно на протяжении длительного геологического времени (Simpson, Beck, 1965; Van Valen, Sloan, 1977; Sloan et al., 1986; Hallam, 1987*).
Столкновение с крупным астероидом, если оно в самом деле произошло, несомненно должно было иметь гибельные биологические последствия. Можно предполагать, что оно вызвало локальное или региональное вымирание некоторого числа видов. Но я скептически отношусь к мысли, что оно могло быть главной причиной вымирания в масштабах всего земного шара.
Астероидная теория — позднейшая из ряда теорий, объясняющих массовое вымирание в меловом периоде теми или иными катастрофами. В выдвигавшихся ранее гипотезах фигурировали вспышки радиации от взорвавшейся новой звезды и летальные дозы космических лучей, достигших поверхности Земли при обращении её магнитного поля. Все эти теории выдвигались физиками. Они представляются упрощенческими и неадекватными попытками истолкования очень сложных биологических явлений.
Большинство эволюционистов-биологов предпочитают ту или иную разновидность климатической или комбинированной климатико-биологической теории массового вымирания в меловом периоде. Поздний мел был временем поднятия материков, горообразовательных процессов, вулканической активности и изменений климата. По меркам мелового (но не четвертичного) периода климат стал неровным. Изменение физической среды влечет за собой множественные биотические события. Изменяется конкурентное равновесие между видами, в результате чего одни виды получают преимущество, а другие оказываются в невыгодном положении, что ведет к замещению видов. Вымирание видов при этом должно быть избирательным, постепенным и растянутым во времени, что и наблюдается на самом деле.
Периоды массового вымирания
Давно установлено, что в геологической истории жизни были периоды массового вымирания, разделенные длительными периодами постепенного эволюционного изменения. Палеонтологическая летопись позволяет предполагать состояние эпизодической эволюции — нечто среднее между теорией катастроф Кювье и строгим униформизмом Лайеля: эпизодическое возникновение периодов массового вымирания, во время которых многие прежние группы гибнут, после чего происходит развитие и адаптивная радиация новых групп.
В самом деле, основой для проведения границ между геологическими эрами и периодами частично служат промежутки между этими эпизодическими массовыми вымираниями, в особенности вымираниями животных. Определяющие признаки эпизодической эволюции животных отражены в названиях эр (палеозойская и т. п.).
Существенная черта эпизодического вымирания — высокая скорость этого процесса, наблюдаемая в совершенно не связанных между собой группах примерно в одно и то же время. Так, у аммоноидов и у рептилий отмечено быстрое вымирание в поздней перми, позднем триасе и позднем мелу (Newell, 1967*).
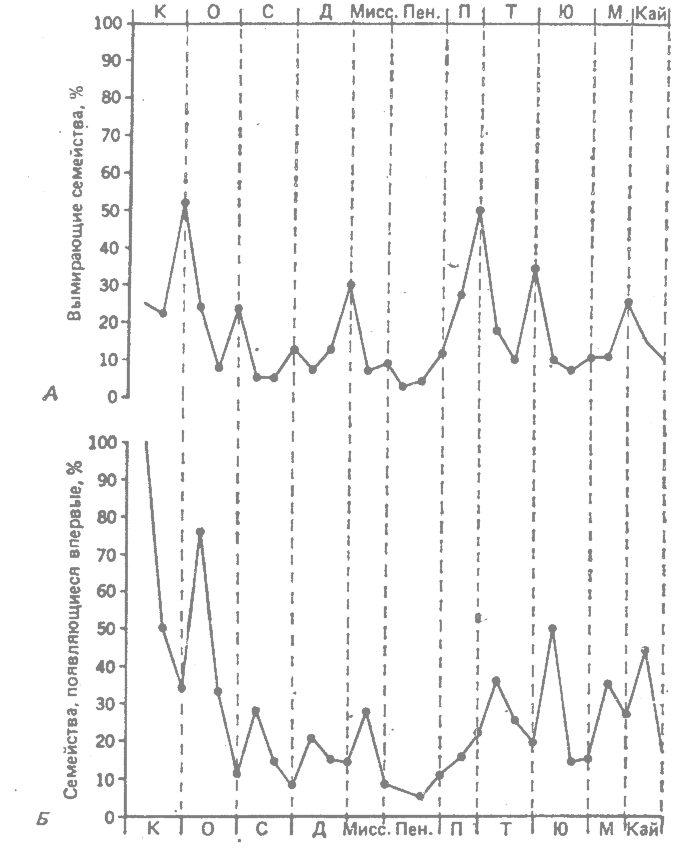 |
Рис. 36.3. Вымирание и замещение у животных. А. Частота вымирания семейств. Б. Частота первых появлений новых семейств по данным палеонтологической летописи. Объяснения см. в тексте. (Из Newell, 1967.*) |
Ньюэлл (Newell, 1967*) сопоставил и проанализировал сведения о первом и последнем появлениях в палеонтологической летописи 2250 семейств животных из всех крупных групп. Полученные им результаты представлены на рис. 36.3. На рис. 36.3, А показана частота вымирания семейств животных на всем протяжении геологической истории по данным об их последнем появлении в палеонтологической летописи. Как можно видеть, эта кривая очень изломана. Высокие пики, значительно возвышающиеся над фоновым уровнем вымирания, наблюдаются в позднем кембрии, позднем девоне, поздней перми, позднем триасе и позднем мелу; более слабо выраженные пики приходятся на концы некоторых из промежуточных периодов.
Недавно Рауп и Сепкоски (Raup, Sepkoski, 1982*) провели анализ палеонтологических данных примерно по 3300 семействам морских животных как беспозвоночных, так и позвоночных. Этот анализ выявил пять периодов массового вымирания семейств начиная с кембрия. Четыре больших пика приходятся на поздний ордовик, позднюю пермь, поздний триас и поздний мел. Пятый небольшой пик относится к девону. Эти результаты очень сходны с данными Ньюэлла.
Большая часть перечисленных выше периодов массового вымирания совпадает по времени с поднятиями материков и установлением континентального климата. Такая корреляция позволила ряду ученых (см. Newell, 1967; Axelrod, 1974*) прийти к заключению, что в конечном счете причиной вымирания (особенно наземных животных) были изменения физической среды.
Позднее Рауп и Сепкоски (Raup, Sepkoski, 1984*) собрали данные о времени существования примерно 2900 ныне вымерших семейств морских позвоночных, беспозвоночных и простейших за промежуток от поздней перми и до конца плиоцена. Они построили график зависимости доли семейств, вымерших в каждом веке, от геологического времени. На протяжении этого времени кривая вымирания семейств образует 12 максимумов. Еще одно очень интересное открытие состоит в том, что эти максимумы вымирания расположены с определённой периодичностью, со средним интервалом 26 млн. лет. Семь из двенадцати максимумов располагаются именно с такими, а пять — примерно с такими интервалами. Несколько проверок с помощью статистических методов подтверждают реальность этой периодичности. Недавно Рауп и Сепкоски (Raup, Sepkoski, 1986, 1988*) вновь провели такое исследование на более обширной выборке из 9773 родов ископаемых морских животных, относящихся примерно к тому же периоду времени. Мерой скорости вымирания служила при этом доля родов, вымиравших за 1 млн. лет. Кривая скорости вымирания родов от перми до современной эпохи показана на рис. 36.4. Эти новые данные подтверждают прежнее заключение. Максимумы вымирания ясно выражены, и интервалы между ними составляют примерно 26 млн. лет.
Такая регулярная периодичность свидетельствует о том, что массовые вымирания были вызваны каким-то астрономическим, а не земным фактором (Raup, Sepkoski, 1984; Raup, 1986*). Этот фактор мог быть связан с Солнцем, Солнечной системой или Галактикой. В настоящее время ведутся поиски астрономического фактора с периодичностью 26 млн. лет (см. Kerr, 1984*).
Ранее в этой главе я отверг гипотезу о внеземной причине массовых вымираний в позднем мелу, а здесь излагаю аналогичную гипотезу и притом с одобрением. Такая непоследовательность объясняется тем, что астероидная гипотеза, во всяком случае в её нынешней формулировке, согласуется не со всеми биологическими данными, тогда как убедительные доказательства существования циклов вымирания с периодичностью 26 млн. лет действительно указывают на какой-то внеземной фактор.
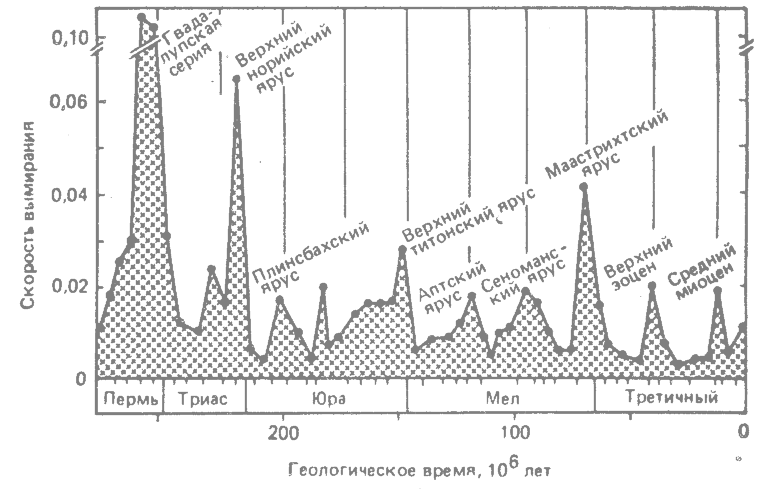 |
Рис. 36.4. Вымирание родов морских животных в течение геологического времени. По оси ординат отложены скорости вымирания родов за период 1 млн. лет. Вертикальными линиями в верхней части графика обозначены промежутки времени продолжительностью 26 млн. лет. (Raup, Sepkoski, 1984.*) |
Этот фактор может только отодвинуть испричину массового вымирания на одну ступень назад. Допустим, например, что некое астрономическое событие периодически уменьшало количество тепла, получаемого земной поверхностью, и за этим последует хорошо знакомая цепь реакций: изменение в физической среде нарушает равновесие как в пределах биотических сообществ и между ними, что в конечном счете приводит к многочисленным замещениям видов. Вернемся к картине отливов и приливов в палеонтологической летописи, которую представил Ньюэлл (Newell, 1967*). На рис. 36.3, Б можно видеть, что первые появления новых семейств животных в этой летописи распределены во времени неравномерно. На кривой появления новых групп имеются максимумы, так же как на кривой вымирания. Кроме того, максимумы на кривой появления групп отстают от максимумов на кривой вымирания. За периодами вымирания следует образование и развитие новых групп. Вымирание старых групп создаёт экологические возможности для формирования новых групп (см. гл. 30).
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |