| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |
Глава 31
Концепция макроэволюции и её типы
Подходы и путеводные нити
Макроэволюция подразумевает изменения гораздо большего масштаба, чем те, которые происходят при микроэволюции и видообразовании. Изменения макроэволюционного уровня состоят в развитии признаков, по которым различаются такие крупные группы, как роды, семейства, отряды, классы и типы. Подобные события происходят в геологическом масштабе времени.
Методы исследования, применяемые при изучении макроэволюции, не могут не отличаться от тех, которые используются при изучении микроэволюции и видообразования. Генетика, экология и систематика на уровне низших категорий — главные подходы к изучению микроэволюции и видообразования, тогда как всё, что нам известно о макроэволюции, получено в результате палеонтологических и сравнительно-морфологических исследований. Работы по генетике популяций и близким к ней областям позволяют получить прямые данные о факторах эволюции. Однако сведения о крупных событиях в процессе органической эволюции дают палеонтология и сравнительная анатомия.
Изучением эволюционной биологиии на макроуровне и на других уровнях обычно занимаются разные группы исследователей, и их представления о роли эксперимента, масштабах времени и других предметах нередко расходятся. Многие палеонтологи и морфологи старшего поколения недооценивали значение генетических исследований для эволюционной теории; в свою очередь биологи, изучающие популяции ныне живущих организмов, часто имеют довольно смутное представление о временном элементе макроэволюции. Источником многих противоречий в эволюционной биологии служат характерные различия между макро- и микроэволюционистами в отношении методов исследования, образа мышления и общей подготовки.
Одна из проблем эволюционной биологии касается того, представляет ли собой макроэволюция просто продолжение микроэволюции или же между этими двумя процессами имеется какоето существенное различие. Многие эволюционисты, в том числе все сторонники синтетической теории, полагают, что интерпретацию явлений макроэволюции следует основывать на наших представлениях о процессах, происходящих на микроэволюционном уровне. Эту точку зрения хорошо сформулировал Добржанский (Dobzhansky, 1937*):
«Опыт показывает…, что единственный путь к пониманию механизмов макроэволюционных изменений, для которых необходимы геологические масштабы времени, — это доскональное изучение микроэволюционных процессов, наблюдаемых на протяжении одной человеческой жизни и нередко контролируемых волей человека».
Изложенная выше точка зрения вызывает, однако, возражения со стороны фракции меньшинства в каждом почти поколении эволюционистов. Гольдшмидт (Goldschmidt, 1940, 1955*) настаивал на том, что макроэволюционное изменение детерминируется особыми (процессами — системными мутациями и скачкообразными изменениями, которые не действуют на микроэволюционном уровне. В последние годы Стэнли (Stanley, 1975, 1979*) и Гулд (Gould, 1980, 1982*) придерживаются мнения о существовании коренного различия между микро- и макроэволюцией. Гулд утверждает, что попытка объяснить макроэволюцию на основе микроэволюции представляет собой редукционизм и неоправданную экстраполяцию и что такой подход ошибочен.
Вопреки мнению Гольдшмидта (Goldschmidt, 1940*) в природе не существует разрыва между микро- и макроэволюцией. Этот предполагаемый разрыв заполняют явления видообразования. В обширной выборке филетичеоких групп эволюционные дивергенции возникают на любом уровне — от рас через виды и группы видов до родов и категорий более высокого ранга. Органическая эволюция составляет единое целое.
Огромное различие в масштабах времени между микро- и макроэволюцией в их крайних формах заставляет остерегаться сверхупрощенных экстраполяций от одного уровня к другому. Многие микроэволюционные изменения повторимы и предсказуемы. В отличие от этого макроэволюция — процесс исторический.
Говоря о длительности времени, в течение которого могли происходить макроэволюционные изменения, следует учитывать также возможность возникновения отдельных уникальных событий, имеющих далеко идущие последствия. Какой-либо примитивный организм может путем инфекции приобрести постоянного симбионта, благодаря которому он становится способным к фотосинтезу. Какой-либо наземный организм может колонизировать отдаленный необитаемый остров всего однажды, после чего начнет эволюционировать на этой занятой им территории в новом направлении.
Возможно, что известные нам эволюционные силы и факторы в разные периоды истории данной группы действовали различным образом. Быть может, частота мутаций в прошлом отличалась от их частоты в настоящее время. Среда, которая сейчас стабильна, в прошлом могла быстро изменяться, и наоборот. Некая группа на одной стадии своей истории могла состоять из большего числа симпатрических видов, способных к ограниченной гибридизации, хотя в настоящее время она монотипична и изменчивость её сильно понизилась.
И наконец, какой аспект микроэволюции следует выбирать в качестве отправной точки для экстраполяции на микроэволюцию? В ранний период синтетической теории эволюционисты экстраполировали от собственно микроэволюции, т, е. от эволюции на популяционном уровне, считая видообразование побочной темой. Однако по ряду причин следует считать, что видообразование занимает в макроэволюции более важное место, чем полагали прежде. Мы вернемся к этому вопросу в дальнейшем изложении.
Адаптивный ландшафт
Адаптивный ландшафт — полезная метафора, помогающая представить взаимодействия между организмом и окружающей средой. К этой метафоре прибегали Райт, Добржанский, Симпсон, Стеббинс и другие эволюционисты.
Представим себе ландшафт, состоящий из холмов и долин. Рельеф местности символизирует распределение адаптивных полей. Вершины холмов — «адаптивные вершины», а долины, разделяющие холмы, представляют собой «ничейную землю» и ряд «адаптивных долин». Популяции и виды занимают разные адаптивные вершины благодаря имеющимся у них сочетаниям адаптивных признаков и лежащим в основе этих признаков сочетаниям генов. Некоторые адаптивные вершины узкие, а другие широкие в соответствии с относительной широтой специализации. Вершины холмов различаются также по высоте; это означает, что на некоторые из адаптивных вершин легче взобраться и легче занять их по сравнению с другими (см. гл. 1).
Эту метафору можно расширить, с тем чтобы распространить её на группы видов, роды и более крупные филетические категории. Вершины распределяются по адаптивному ландшафту не случайным образом, а собраны, во всяком случае до некоторой степени, в отдельные цепи. При этом цепь холмов соответствует адаптивной зоне того или иного рода.
До сих пор мы рассматривали адаптивный ландшафт в статическом аспекте. А как обстоит дело с его динамикой? И организм, и среда подвержены изменениям.
Каким образам виды занимают свои адаптивные вершины, а роды — свои горные цепи? Путями, ведущими к адаптивным вершинам и горным цепям, служат направления эволюции. А скорость восхождения соответствует скоростям эволюции. Адаптив ные вершины разной высоты достигаются в результате разных сочетаний направлений эволюции и её скоростей.
Адаптивный ландшафт не всегда остается статичным и неизменным. Подобно настоящему ландшафту, он подвержен действию сил, вызывающих эрозию и горообразование. Эти силы могут действовать медленно или быстро. Изменения ландшафта отражают роль изменений среды в эволюции. Адаптивная вершина может перемещаться с той или иной скоростью, и популяция, которая её занимает, должна стараться не отставать от нее.
Необратимость
Нередко утверждают, что эволюция необратима. Это обобщение известно как закон Долло; необходимо, однако, указать, на некоторые важные ограничения.
Прежде всего необратимость нельзя считать характерной чертой микроэволюции, поскольку возникающие на этом уровне генетические изменения относительно просты и направление может легко изменяться. Частота отдельного аллеля или участка хромосомы может повышаться за счет конкурирующего с ним аллеля или участка при одних условиях среды, но понижаться относительно этого другого аллеля или участка в случае изменения среды в противоположную сторону. Необратимость становится чертой эволюции на уровнях видообразования и макроэволюции.
Здесь следует провести ещё одно различие. Макроэволюционные изменения необратимы в том, что касается морфологических признаков и их генетических детерминантов, но не в том, что касается экологии. Многие группы наземных рептилий, наземных млекопитающих и обитающих в воздушной среде птиц вернулись в водную среду — местообитание своих примитивных позвоночных предков, но морфологически они остались при этом рептилиями, млекопитающими или птицами. Общие размеры в разных группах млекопитающих, например у лошадей, могут увеличиваться в течение одного периода времени и уменьшаться в течение другого. Однако производные мелкие формы морфологически и генетически не идентичны мелким предковым типам.
Суть проблемы — в исторической природе макроэволюционного изменения (Simpson, 1953*). Генетические изменения постепенно наслаиваются друг на друга на протяжении длительных периодов времени, так что возврат к первоначальному состоянию невозможен. В длительной эволюции сложного генотипа история не повторяется в точности. Но это не исключает возможности реверсий в адаптивной морфологии.
Анагенез и кладогенез
На основании направления эволюционного изменения уже давно различают два типа эволюции. Первый — изменение во времени, происходящее в видовой популяции; второй — разветвление или дивергенций на видовом и более высоких уровнях. Для обозначения этих двух типов широко используют два термина — анагенез и кладогенез (Rensch, 1947; 1960b*), Дарвин называл эти типы эволюции «descent with modifications» («происхождение с изменениями») и «origin of species» («происхождение видов»). Другие авторы применяли термины филетическая эволюция и эволюционная дивергенция.
Майр (Mayr, 1982a*) сделал интересное замечание, указав, что ранние эволюционисты обычно уделяли внимание или по крайней мере выдвигали на первый план либо один, либо другой из этих типов. Так, для Ламарка главным был анагенез, а для Дарвина — кладогенез.
Эволюция, несомненно, происходит как тем, так и другим способом. Однако макроэволюция, возможно, не так проста, чтобы ограничиться этой терминологической дихотомией. Картина осложняется такими дополнительными типами, как сетчатая и квантовая эволюция, рассматриваемыми в дальнейших разделах этой главы. Кроме того, разграничение анагенеза и кладогенеза нарушается в случае эволюционных направлений некоторых типов, а именно видообразовательных направлений, которые также будут рассмотрены позднее.
Адаптивная радиация
При этом способе эволюции преобладает кладогенез, создающий возможность для возникновения разнообразия в данной группе, благодаря чему её члены становятся способными осваивать различные местообитания и ниши на занимаемой ею территории (см. гл. 30).
Осборн (Osborn, 1910*) описал этот способ эволюции на примере эволюции млекопитающих и дал ему название. Он различал у млекопитающих пять главных линий специализации в строении конечностей, делающих их обладателей приспособленными к различным местообитаниям и способам передвижения: 1) быстрый бег для наземных видов; 2) роющий образ жизни для видов, обитающих под землей; 3) плавание для видов, обитающих и в воде, и на суше, и для полностью водных видов; 4) лазание для древесных видов; 5) планирование и полет для видов, осваивающих воздух. Осборн различал также линии специализации в строении зубов, зависящие от характера питания (всеядность, плотоядность, растительноядность и т. п.).
Отряды млекопитающих, согласно Осборну (Osborn, 1910*), обладают этими группами признаков в различных сочетаниях, а следовательно, относятся к разным адаптивным типам. Более новые сведения об адаптивном характере отрядов млекопитающих приводит Ван Вален (Van Valen, 1971*). Современный обзор адаптивной радиации млекопитающих на всех таксономических уровнях см. Эйзенберг (Eisenberg, 1981*).
Симпсон (Simpson, 1977*) справедливо указывает, что Дарвин определил этот тип эволюции задолго до Осборна (см. Darwin, 1859*, гл. 4). Дарвин подчёркивал «дивергенцию признаков» и связи с межвидовой конкуренцией и экономическим разнообразием. Вероятно, можно оказать, что Осборн разработал далее один из аспектов рассмотренной Дарвином проблемы в более явной форме и дал ему название, которым мы теперь пользуемся.
Конвергенция и параллельное развитие
Конвергенция — это процесс, противоположный адаптивной радиации. Конвергенция состоит в независимом развитии одинаковых признаков у двух или более отдаленно родственных линий в результате адаптации к сходным экологическим условиям или обстоятельствам. Наличие общих предков у таких линий не играет роли в сходстве их признаков (перефразируя Simpson, 1961*).
Примеров конвергенции великое множество. В качестве таких примеров часто приводят различные адаптивные типы сумчатых в Австралии и их плацентарных аналогов на других континентах, а также стеблевые суккуленты молочайных и кактусов из сухих жарких областей Африки и Америки соответственно.
Параллельное развитие сходно с конвергенцией по конечным результатам, но оно идет иным филогенетическим путем. Параллельное развитие определяется как независимое развитие сходных признаков в двух или нескольких родственных линиях, сходство которых канализировано признаками общего предка этих линий (Simpson, 1961*).
Параллельное развитие также широко распространено. Здесь можно напомнить о развитии высшей нервной деятельности в различных отрядах млекопитающих. Некоторые другие примеры приводились в гл. 28.
Новая адаптивная зона часто колонизируется двумя или более неродственными или отдаленно родственными группами, конкурирующими между собой. Если прежде остававшаяся незанятой адаптивная зона оказывается доступной, т. е. если существуют преадаптированные к ней организмы, то нередко две или несколько таких преадаптированных групп хватаются за эту возможность примерно в одно и то же время. Это приводит к кон вергенции или параллельному развитию в широких масштабах.
Так, в мезозое адаптивную зону летающих млекопитающих занимали птицы и птерозавры. В эту же эру адаптивная зона растений, опыляемых насекомыми, была занята голосеменными, саговниками и, возможно, другими группами. Жизненная форма млекопитающих в этой эре развивалась сумчатыми и плацентарными, а также отдаленно родственными им однопроходными. Подобным же образом жизненную форму покрытосеменных развивали параллельно двудольные и однодольные. В более раннюю эру адаптивная зона для морских растений средних или крупных размеров была колонизирована независимо зелеными, бурыми и красными водорослями.
Одновременное проникновение в новую адаптивную зону нескольких групп отражается в таксономии некоторых категорий высокого ранга. Какой-нибудь крупный таксон часто содержит аномальную подгруппу, которая не вполне вписывается в данный таксон, но вместе с тем не заслуживает выделения в самостоятельную сестринскую группу. Однопроходные представляют собой нечто вроде побочной ветви млекопитающих, так же как бурые и красные водоросли в растительном, а губки — в животном царстве. Параллельное развитие на высших уровнях вносит известную асимметрию и неряшливость в таксономическую систему.
Ортогенез или ортоселекция?
Многие эволюционисты старшего поколения для объяснения длительных эволюционных направлений прибегали к ортогенезу в том или ином его варианте (см. гл. 2). В частности, эту теорию применяли и для объяснения направлений эволюции лошадей. Обширные данные по Equidae можно использовать для проверки теории ортогенеза. Этой проверки она не выдерживает.
Если данное эволюционное направление — результат ортогенеза, оно должно быть общим для всей филетической группы и не отклоняться. Однако в эволюции Equidae неизвестны направления, которые продолжались бы на всем протяжении истории какой-либо филетической линии или происходили бы одновременно во всех линиях. Общим направлением для эволюции Equidae было увеличение размеров тела, но в некоторых линиях иногда возникали реверсии. Филогения лошадей на самом деле сильно разветвлена, образуя много боковых ветвей, и некоторые направления, в том числе линия, приведшая к Equus, относятся к боковым ветвям (Simpson, 1951; 1953*) (см. гл. 28).
Согласно альтернативной интерпретации, берущей начало в синтетической теории, направления, наблюдаемые в эволюции лошадей, определяются взаимодействием между организмом и средой. Если отбор, связанный с факторами среды, продолжается в данном направлении достаточно долго, его можно назвать ортоселекцией и он приведет к возникновению определённого эволюционного направления (Simpson, 1944*).
Данные по эволюции Equidae соответствуют объяснению на основе ортоселекцни. Наблюдаемые направления явно адаптивны. Кроме того, они не только отклоняются, но и проявляют признаки оппортунизма, оставаясь неизменными при одном комплексе условий среды, но смещаясь при их изменении. Наконец, изучение некоторых направлений эволюции признаков зубов (см. гл. 28) показывает, что в их основе лежит полиморфизм предковых популяций (Simpson, 1951; 1953*).
Сетчатая эволюция
Во многих группах растений наиболее инклюзивной (inclusive) единицей обмена генами на каждом временном уровне является сингамеон — комплекс гибридизирующихся между собой время от времени видов, — и развивающаяся во времени эволюционная линия представляет собой изменяющееся сочетание гибридизирующихся видов. Единицей филетической эволюции служит при этом не отдельный вид, а анастомозирующие сети видов. Анагенез и кладогенез переплетены. Макроэволюция же носит сетчатый характер. Сетчатая эволюция широко распространена у сосудистых растений. Объем книги не позволяет рассмотреть этот тип эволюции во всей его полноте; подробное изложение предмета см. Grant, 1981a.*
Квантовая эволюция
Как уже было сказано в гл. 30, квантовая эволюция представляет собой относительно быстрый переход какой-либо линии из прежней адаптивной зоны в новую и от предкового состояния признаков — к явно иному, адаптированному к этой новой зоне. В этом разделе нам предстоит рассмотреть концепцию квантовой эволюции*.
Симпсон (Simpson, 1944, 1953*) выдвинул концепцию квантовой эволюции как особого способа эволюции, равного по значению филетической эволюции и дивергенции. В некоторых случаях она постепенно переходит в филетическую эволюцию. Она может также участвовать в видообразовании. Симпсон, однако, особо подчеркивает её тесную связь с филетичеокой эволюцией; квантовая эволюция — это «особый более или менее экстрёмальный и граничный случай филетической эволюции» (Simpson, 1953*). Хотя квантовая эволюция переходит в эти два других способа эволюции, она в своей хорошо развитой форме отличается от каждого из них и, таким образом, согласно Симпсону, заслуживает признания как третий способ.
Квантовая эволюция происходит в небольших популяциях. В первоначальном варианте Симпсон (1944*) говорил о маленьких изолированных популяциях, а в пересмотренном (1953*) — о подразделенных популяциях. В некоторых, но не во всех, случаях, возможно, имеет место дрейф генов (Simpson, 1953*).
Симпсон (1944, 1953*) ощущал, что популяции, переходящие из одной адаптивной зоны в другую, будут находиться в слабо адаптированном или нестабильном состоянии. Во всяком случае, переходная стадия должна быть непродолжительной.
Быстрые эволюционные скорости характерны для квантовой эволюции. Но сколь велики эти щедрости? Они рассматриваются в палеонтологическом масштабе времени. В одном месте Симпсон (1953, стр. 356*) высказал предположение, что переход из одной адаптивной зоны в другую может занять период времени порядка 1—10 млн. лет.
Квантовая эволюция носит эпизодический характер, встречаясь в истории той или иной линии в качестве исключительного события. Обширная предковая популяция эволюционирует с горотелическими или брадителическими скоростями вплоть до момента квантовой эволюции. Затем возникают быстрые скорости при сопутствующих небольших размерах популяций. После завершения квантового сдвига данная популяция, по всей вероятности, возвращается к состоянию большой популяции с горотелическими или брадителическими скоростями эволюции (Simpson, 1944, 1953*; рис. 31.1).
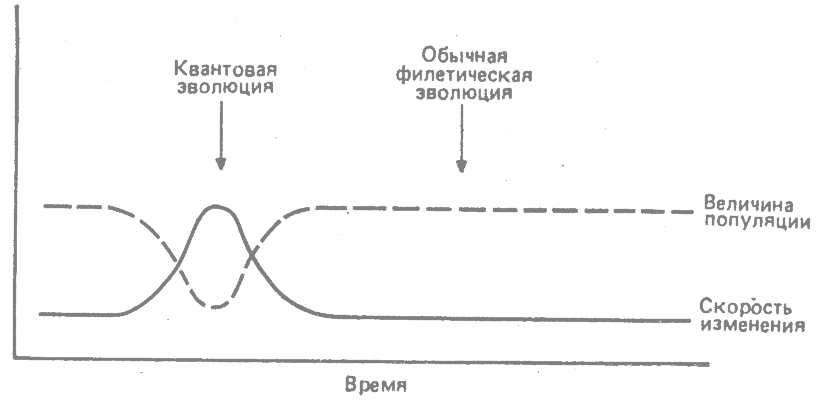 |
рис. 31.1. Сравнение скорости эволюционных изменений и величины популяции при квантовой и обычной филетической эволюции. (Simpson, 1953.*) |
Такая последовательность событий соответствует палеонтологическим данным. История многих групп представляет собой длинную, запечатлённую в палеонтологической летописи последовательность, в эволюции которой преобладали горотелические или брадителические скорости; этому предшествовал гораздо более короткий, не отраженный в летописи период, в течение которого должна была возникнуть данная группа. Палеонтологическая летопись свидетельствует о больших размерах популяций, сопровождавшихся горотеличеакими или брадителичеакими скоростями в периоды нормальной филетичеокой эволюции. И наоборот, отсутствие или редкость ископаемых остатков, относящихся к периоду возникновения и главного развития группы, согласуется с постулатом о небольших размерах популяции во время квантовой эволюции. Эти постулированные черты квантовой эволюции позволяют объяснить крайнюю редкость так называемых недостающих звеньев в палеонтологической летописи (Simpson, 1944, 1953*).
Продукт квантовой эволюции — новая группа организмов и притом нередко новая крупная группа. Полагают, что квантовая эволюция участвует в возникновении большинства новых крупных групп (Simpson, 1944, 1953*).
За несколько десятилетий, прошедших после того как Симпсон изложил свою концепцию квантовой эволюции, в изучении эволюции были достигнуты большие успехи. Настало время для некоторого пересмотра тех черт, которые были первоначально приписаны этому способу эволюции. Мы сосредоточим свои замечания на пяти аспектах: палеонтологическая картина, величина популяции, дрейф генов, скорость эволюции и соотношение с другими способами эволюции.
Первый аспект — палеонтологическая картина — очень хорошо выдержал проверку временем. Многие группы, после того как они сформировались, склонны оставаться весьма консервативными. Примеры длительной стабильности видов обнаружены, в частности, среди млекопитающих (Stanley, 1982*), двустворчатых моллюсков (Stanley, Yang, 1987*) и жуков (Coope, 1979*).
Вопрос об участии или неучастии дрейфа генов в квантовой эволюции сегодня лучше формулировать в терминах сочетания отбор — дрейф. Отбор — дрейф представляет собой силу, наиболее пригодную для того, чтобы вызывать быстрый сдвиг в небольших популяциях.
Первоначальная идея о небольших размерах популяций несомненно была шагом в верном направлении, но сегодня её можно расшифровать более детально. Эволюционный потенциал небольших изолированных популяций сильно ограничен, как было отмечено в гл. 29. Возникновение квантовой эволюции наиболее вероятно в колониальных популяционных системах и группах периферических популяций-основателей (см. гл. 16).
Это соображение ведет к дальнейшей дедукции о типе местности в которой можно ожидать возникновения новых крупных групп. Для квантовой эволюции необходимы популяционные системы островного типа. Такие популяционные системы не характерны для равнин. Они обычно встречаются на пересеченной местности с разнообразными местообитаниями; для наземных растений и животных, например, это могут быть горы. Можно ожидать, что такого рода местность служит центром возникновения крупных групп.
Что касается скоростей квантовой эволюции, то следует отметить, что тахителия охватывает довольно широкий диапазон скоростей. Палеонтологи считают тахителическими такие скорости, при которых морфологические изменения происходят за 10 млн. лет. Эволюционисты, изучающие популяции ныне живущих растений, обнаруживают доказательства возникновения существенных изменений за гораздо более короткое время, измеряемое десятками или сотнями лет. Симпсон и другие ранние исследователи квантовой эволюции (Schaeffer, 1948; Patterson, 1949; Westoll, 1949*) были палеонтологами и рассматривали этот способ эволюции в палеонтологическом масштабе времени.
Возникает вопрос, не является ли такой подход слишком консервативным, не упускаем ли мы при этом чего-то из виду? Быть может, палеонтологические примеры квантовой эволюции занимают промежуточное положение, будучи несколько сдвинуты в сторону филетичеакой эволюции, как это указывал Шаффер (Schaeffer, 1948*) в случае изучавшейся им механики конечностей у парнокопытных. Возможно, что в неонтологическом масштабе времени некоторые существенные квантовые сдвиги действительно происходят, но не оставляют следов в палеонтологической летописи.
По первоначальным представлениям Симпсона, участие видообразования в квантовой эволюции необязательно, но возможно в некоторых случаях. Я бы пошел дальше, считая, что видообразование обычно и даже непременно участвует в квантовой эволюции, если она происходит в неонтологическом масштабе времени. Вряд ли действительно быстрый квантовый переход из одной адаптивной зоны в другую может иметь место без одновременного сдвига видообразования. Способ видообразования, который я назвал квантовым видообразованием, особенно подходит для того, чтобы вызывать быстрые адаптивные сдвиги в неонтологическом масштабе времени.
Концепция квантовой эволюции в её первоначальной формулировке (Simpson, 1944, 1953*) охватывала довольно широкий диапазон условий в смысле скорости и направлений изменения. Филетическая эволюция выдвигалась на первый план, но допу скалась и дивергенция. В случае геологических масштабов времени предусматривалась тахителия, но следовало бы указать также на возможность быстрых скоростей и при неонтологических масштабах. В свете современных представлений было бы желательно подразделить квантовую эволюцию на более однородные способы. Мы предлагаем следующее деление: 1) тахителическая филетичеокая эволюция в геологическом масштабе времени; 2) образование боковых ветвей с геологически тахителичеокими скоростями; 3) квантовое видообразование.
Квантовое видообразование
Как уже было сказано в гл. 24, квантовое видообразование состоит в том, что от большой полиморфной свободно скрещивающейся предковой популяции отпочковывается дочерний вид, проходящий через промежуточную стадию небольшой локальной расы. Локальная раса основывается одной или несколькими особями из популяции предкового вида. Она пространственно изолирована или полуизолирована от предкового вида, обычно находится на периферии области, занимаемой старым видом, и может встречаться в какой-нибудь новой экологической зоне.
В начале существования локальной расы на её генофонд оказывают влияние дрейф генов и инбридинг, а позднее — сочетание отбора с дрейфом и нередко генетические революции. Эти факторы могут вызвать быстрые, а иногда и коренные изменения в экологических предпочтениях, морфологических признаках и взаимоотношениях, определяющих фертильность. Поэтому локальная раса может быстро дивергировать до видового уровня; более того, возникший таким образом дочерний вид может занять новую адаптивную зону, лежащую за пределами экологического ареала предкового вида.
Быстрый адаптивный сдвиг во время квантового видообразования облегчается, кроме того, структурой популяции, которая может избежать ограничения, налагаемого на множественную замену генов платой за отбор (см. гл. 17).
Концепция (квантового видообразования это в сущности синтетическая концепция. Она представляет собой синтез избранных частей более ранних концепций Райта (Wright, 1931*), Симпсона (Simpson, 1944*) и Майра (Mayr, 1954*), а также концепций, вытекающих из работ Льюиса по эволюции растений (Lewis, Raven, 1958; Lewis, 1962*). Однако ни одна из прежних концепций не соответствует в точности описанию квантового видообразования. Концепция отбора — дрейфа взята у Райта (но не других авторов), квантовые сдвиги (но не видообразование) — у Симпсона, а быстрое видообразование (но не отбор—дрейф) — у Майра и Льюиса. Синтетическая концепция квантового видообразования объединила в себе эти компоненты.
Генетические революции
Майр (Mayr, 1954, 1963*) высказал мнение, что популяции должны претерпевать глубокую генетическую реорганизацию при прохождении через «узкое горлышко» низкой численности. Его тезис основан на известных фактах, относящихся к гомеостазу у экспериментальных организмов, и следует начать с того, чтобы кратко суммировать эти факты (см. гл. 3).
Свободно скрещивающиеся особи у дрозофил, мышей, кур, первоцвета, кукурузы и других организмов высокогетерозиготны и гомеостатичны. Они проявляют фенотипическое постоянство на фоне изменяющихся условий среды. Гетерозиготность и гомеостаз связаны между собой: второй зависит от первой. Кроме того, хорошо забуференный нормальный фенотип возникает не при какой-то одной гетерозиготной комбинации, а может быть достигнут многими путями при участии различных гетерозиготных комбинаций и может даже зависеть от гетерозитотности по многим локусам. Одним из результатов утраты или сильного снижения мультилокусной гетерозиготности вследствие инбридинга может быть появление потомков с аберрантными признаками, так называемых феноотклонений (Дубинин, 1948; Lerner, 1954*). Майр (Mayr, 1954*) приложил эти соображения к анализу природных популяций. Он противопоставляет внутреннюю генетическую среду в больших широко распространенных популяциях свободно скрещивающегося вида этой же среде в маленьких изолированных производных популяциях того же вида.
В любую локальную субпопуляцию, входящую в состав большой непрерывной популяции всё время поступает поток генов из соседних, а иногда и из отдаленных субпопуляций. Более или менее частая иммиграция чужих генов в данную локальную популяцию создаёт давления отбора, благоприятствующие генам с хорошей комбинационной способностью. В результате в состав её генофонда входят группы генов, дающие начало нормальным жизнеспособным продуктам в весьма разнообразных гетерозиготных генных сочетаниях. Фонд «хорошо смешивающихся» генов, создающийся в такой генетической среде, обладает также в качестве естественного следствия гомеостатическими буферными свойствами (Mayr, 1954*).
Это придает известный консерватизм характеру изменчивости в большой непрерывной популяции. Такая популяция проявляет клинальную изменчивость и географическую расовую изменчивость (см. гл. 18). Но отклоняющиеся или новые формы данного вида не могут сохраняться и не сохраняются в качестве самостоятельных расовых единиц.
Посмотрим теперь, что происходит, когда нескольким особям, эмигрировавшим из большой свободно скрещивающейся попу ляции, удается основать небольшую дочернюю колонию на периферии ареала вида. Прежде всего такая популяция основателей будет содержать лишь небольшую неслучайную выборку из генофонда родительской популяции. Но, кроме того, она будет изолированна от потока генов извне. Это приводит к изменению генетической среды. Селективные ценности генов в этих изолированных дочерних генофондах изменены: отбор уже не благоприятствует «хорошо смешивающимся» генам; и напротив, гены, дающие в гомозиготном состоянии жизнеспособные типы, приобретают теперь положительное селективное значение (Mayr, 1954*).
Таким образом, переход от большой свободно скрещивающейся лопуляции к небольшой изолированной дочерней колонии уже сам по себе изменяет генетическую среду и изменяет или даже полностью ревертирует селективные ценности генов во многих локусах. Изменение внутренней генетической среды воздействует одновременно на многие генные локусы, подобно изменению, вызываемому гибридизацией или полиплоидией, и оно может послужить началом «генетической революции» (Mayr, 1954*).
Популяция может оказаться неспособной пройти через «узкое горлышко» — от высокой к низкой численности. Она может оказаться не в состоянии выдержать генетическую революцию; в таком случае она будет обречена на быстрое вымирание. Но при большом числе независимых попыток основать новые колонии есть шансы, что одной или нескольким популяциям основателей удастся выжить. В таких случаях выживание зависит от того, унаследовала ли популяция основателей такой генофонд, который может выдержать гомозиготность по многим локусам и который преадаптирован к какой-либо из имеющихся новых экологических зон.
Генетическая революция в удачной дочерней колонии приводит к более или менее коренному изменению экологических предпочтений и морфологических признаков. Возникают новые типы, аналогичные феноотклонениям у экспериментальных организмов. Интерес представляют новые типы, приспособленные к новым экологическим зонам, которые находятся за пределами зоны, занимаемой консервативной предковой популяцией. Таксономические данные до некоторой степени подтверждают это ожидание. В изолированных периферических популяциях некоторых видов и групп видов птиц и растений действительно наблюдаются новые варианты (см. гл. 23 и 24).
Короче говоря, изолированные периферические популяции, возникающие от нескольких основателей, отделившихся от большой свободно скрещивающейся предковой популяции, могут быть теми точками, где происходит быстрая эволюция и появляются новые формы (Mayr, 1954*).
Дивергирующая популяция не обязательно навсегда остается маленькой, изолированной и периферической. Она может увеличиваться в размерах, расширять занимаемую ею территорию, а в конечном счете даже вновь вернуться на территорию предкового вида. Со временем она также может стать консервативной по характеру своей изменчивости. Но после этого она может дать начало новому поколению изолированных дочерних колоний, которые снова отрываются от предковой формы (Mayr, 1954*).
Майр создал свою концепцию генетических революций в значительной мере независимо от взглядов Райта на подразделенную популяцию и от представлений Симпсона о квантовой эволюции. Тем не менее эти три концепции хорошо согласуются между собой в отношении ожидаемых конечных результатов для небольших популяций.
Вопрос об эволюционной силе, участвующей в дивергенции маленьких дочерних популяций, вызывает разногласия. Майр приписывает это эволюционное изменение эффекту основателя, отбрасывая роль дрейфа генов (Mayr, 1954, 1963*). Однако эффект основателя обусловлен случайностями выборки, возникающими тогда, когда одна или несколько особей, мигрирующих из большой предковой популяции, основывают новую дочернюю колонию, и поэтому его можно рассматривать как особый случай дрейфа генов. Кроме того, дело здесь не в дрейфе как таковом. Скорее всего фактором, детерминирующим дивергенцию маленьких дочерних популяций, служит сочетание отбора и дрейфа генов (см. гл. 16).
Филетические направления и видообразовательные направления
Рассмотрим теперь более подробно природу эволюционных направлений. В работах, относящихся к периоду с тридцатых до пятидесятых годов нашего века, обычно допускалось или утверждалось, что эволюционные направления создаются в результате анагенеза или филетичеаиой эволюции. Возможно, что во многих случаях это так. Однако позднее стали высказывать мнение, что некоторые или даже многие направления представляют собой результат видообразования. Некоторое число видообразовательных событий может быть связано друг с другом в один временной ряд, образуя эволюционное направление (Grant, 1963, 1971; Stanley, 1975, 1979*).
Согласно этой точке зрения, эволюционные направления делятся на два больших класса: филетические направления и видообразовательные направления (Grant, 1963, 1977a, b, 1989*)*. Их можно определить следующим образом.
Филетическое направление — это постепенное эволюционное изменение в пределах данной видовой линии, идущее в одну сторону и обусловленное исключительно филетической эволюцией. Отдельные стадии при этом представлены последовательными видами, а не существующими одновременно биологическими видами.
Примером филетического направления служит эволюция строения коренных зубов в линии ныне вымерших плейстоценовых европейских слонов. В эту линию входят два последовательных вида — более древний Elephas planifrons и более молодой Е. meridionalis (рис. 31.2). Другим примером служит филетическое направление Homo erectus — Н. sapiens.
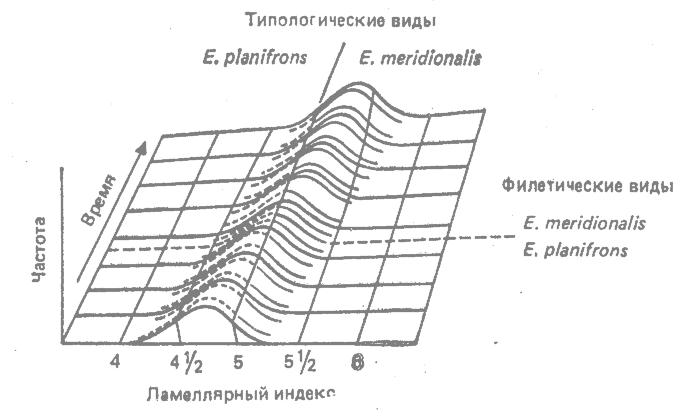 |
Рис. 31.2. Филетическое направление в линии плейстоценовых европейских слонов Elephas planifrons — Е. meridionalis. Ламеллярный индекс служит мерой количества эмали на коренных зубах. Филетические виды соответствуют последовательным видам, а типологические виды — таксономическим видам. (Из Simpson, 1953.*) |
Видообразовательное направление представляет собой ступенчатое продвижение вперед, возникающее в результате последовательного ряда видообразовательных изменений. Каждый новый вид ряда продвигается дальше в данном направлении. Это направление — линия Р—Q на рис. 31.3. Отдельные стадии этого направления представлены биологическими, а не последовательными видами. В этом случае преобладающим типом эволюции является видообразование, а не филетическая эволюция. Видообразовательное направление — суммарный результат последовательных видообразований.
При филетическом направлении эволюции на каждом данном временном уровне имеется только один биологический вид. При видообразовательном направлении консервативная предковая форма и происходящая от нее более прогрессивная форма временно существуют как два современных друг другу биологических вида. Линия Р—Q на рис. 31.3, хотя она и состоит из последовательности видов в буквальном смысле слова, не содержит последовательных видов; поэтому в соответствии с принятыми здесь критериями линия Р—Q отражает не филетическое, а видообразовательное направление.
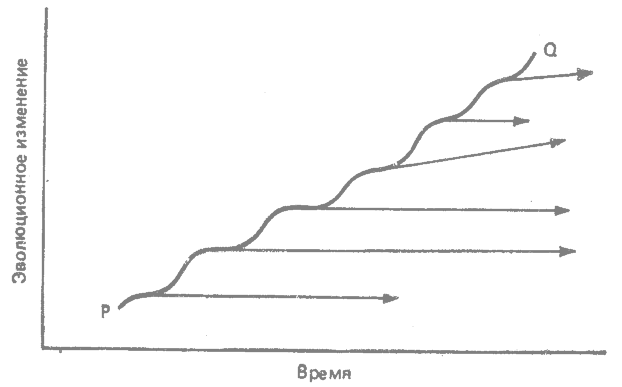 |
Рис. 31.3. Направление видообразования. Направление (линия Р—Q) представляет собой результирующий вектор последовательных видообразовательных изменений в данном направлении. (Из Grant, 1963.*) |
Скорости эволюции в данном филетическом направлении будут самыми низкими в тех случаях, когда большая непрерывная популяция подвергается массовому направленному отбору. Скорости могут быть выше, если популяция подразделена и подвергается совместному действию отбора — дрейфа генов (см. гл. 29). Однако в общем эволюционные скорости, которые могут поддерживаться в последнем случае, вероятно, никак не достигают рассматриваемых в следующем абзаце.
Как указывалось в гл. 25, самое быстрое и оперативное закрепление новых адаптивных комбинаций генов достигается путем видообразования. Если видообразование служит наилучшим способом на временном уровне 1, то оно может быть наилучшим способом и на уровнях 2, 3 и 4. Таким образом, видообразовательные направления способны вызывать достаточно крупные эволюционные изменения за относительно короткий период времени. Теоретически скорости эволюции в этих направлениях могут быть выше, чем в филетических направлениях.
Ориентация видообразовательного направления складывается в зависимости от какого-либо градиента или изменения среды, идущего в одну сторону. Среда контролирует это направление посредством межвидовой конкуренции и межвидового отбора. Допустим, что какой-то вполне сложившийся предковый вид растений адаптирован к теплому влажному климату и что этот климат становится всё холоднее и холоднее. Среди происходящих от родительского вида дочерних видов успеха сможет добиться только форма, адаптированная к холодному климату. Производные формы с другими экологическими предпочтениями будут подавлены в зародыше. Этот адаптированный к холоду вид первого поколения может затем в свою очередь произвести дочерний вид, который будет даже ещё более холодоустойчивым, и т. д. Видообразовательные сдвиги ко всё более холодным условиям — единственный путь, возможный в условиях данного градиента среды, а поэтому развитие видообразовательного направления ориентировано в эту сторону (Grant, 1963, 1977a, b, 1989; Stanley, 1979*).
Возможно, что видообразовательные направления довольно обычны. Трудно сказать, однако, насколько часто они встречаются, поскольку нелегко идентифицировать видообразовательное направление per se, если ископаемая летопись, как правило, неполна. Ископаемые остатки по какой-либо группе, поскольку они бывают собраны из разных отложений, обычно разбросаны по отношению к подлинному филогенетическому древу. В публикуемом филогенетическом древе, основанном на таких данных, т. е. в филогенетической гипотезе, имеются пробелы и неясности. В сущности по одним и тем же неполным палеонтологическим данным часто можно построить различные филогении в зависимости от того, как соединять боковые линии в тех местах, где в летописи имеются пробелы.
Недавно был описан пример видообразовательного направления у ордовикского рода Pydogus (Conodontophoridae). Более молодой вид P. anserinus замещает P. serrus в некоторых вертикальных стратиграфических колонках, что сначала наводит на мысль об анагенетических рядах; однако в дальнейшем после получения данных из других мест становится очевидным, что P. anserinus произошел от P. serrus в результате аллопатрического видообразования и в течение некоторого времени существовал одновременно со своим предковым видом (Fahraeus, 1982*).
Два вероятных примера видообразовательных направлений у растений выведены на основании изучения сравнительной биологии современных форм. У таких выводов есть слабые стороны, но также и свои достоинства, поскольку можно изучать виды в живом состоянии и определять их взаимоотношения.
Вероятное видообразовательное направление в эволюции Clarkia (Qnagraceae) на тихоокеанском побережье Северной Америки идет от форм, приспособленных к климату с умеренной влажностью, до ксерофитов. Предковый вид, обитавший в умеренно влажных местах, был близок к современным видам Clarkia amoena или С. rubicunda. Промежуточная стадия в этом ряду представлена ныне живущим видом С. biloba, а конечная ксерофитная стадия — видом С. lingutaia. Переход от С. biloba к С. Ungulata произошел в результате квантового видообразования (Lewis, Lewis, 1955; Lewis, Roberts, 1956*).
Вероятное направление эволюции у Polemonium (Polemoniaсеае) в западной части Северной Америки идет от форм, приспособленных к мягкому климату прибрежных областей умеренного пояса, до альпийских форм. Отдельные стадии этого направления представлены следующими видами:
| Potemonium carneum | Влажные прибрежные низменности |
| P. occidentale | Зона хвойных лесов |
| P. pulcherrimum | Субальпийская лесная зона и нижняя альпийская зона |
| Группа P. eximium | Высокие вершины в альпийской зоне и альпийская тундра |
Этот ряд видов встречается в указанных зонах по трансектам от побережья до высоких гор в западной части Северной Америки. При этом названные виды не образуют по таким трансектам непрерывный ряд, как это могло бы представляться теоретически, а обособлены друг от друга, составляя ступенчатую последовательность, что позволяет предполагать видообразовательное направление (Grant, 1959, 1963*).
Представляется вероятным, что видообразовательные направления вследствие их способности приводить к существенным изменениям и их тахителических скоростей могут участвовать в эволюции крупных групп в период их формирования.
Квантовые видообразовательные направления
Скорость эволюции в видообразовательных направлениях, возможно, коррелирует с тем способом видообразования, с которым они связаны. Можно ожидать, что последовательности географических видообразований в популяциях, постоянно сохраняющих большие размеры, будут происходить с умеренной или нормальной, но не с тахителической скоростью. Последовательные же квантовые видообразования теоретически могли бы возбуждать высокие скорости у соответствующего эволюционного направления.
Если особые преимущества квантового видообразования могут быть использованы однажды, на одном временном уровне, то они могут быть использованы снова и снова. Сдвиги, происходящие при квантовом видообразовании, могут циклически повторяться. Эти сдвиги иногда имеют (некое общее направление подобно линии Р—Q на рис. 31.3. Квантовое видообразовательное направление подобного рода представляет собой способ обеспечения длительной быстрой эволюции в данном направлении. Быть может, это главный способ, с помощью которого эволюционное направление происходит с высокой тахителичеокой скоростью.
Вернемся к метафоре адаптивного ландшафта, предложенной в начале этой главы. Представим себе незанятую высокую адаптивную вершину и популяцию, находящуюся у её подножия. От подножия к вершине ведет несколько различных троп. Некоторые из них пологие и, следовательно, восхождение по ним будет долгим и медленным. Эти тропы символизируют филетические направления и направления, связанные с географическим видообразованием. Они пригодны при наличии достаточного времени. Другие тропы крутые, но короткие и потенциально создают возможность для быстрого восхождения. Если популяция, находящаяся у подножия, должна быстро взобраться на вершину, потому что время для восхождения ограничено, то ей придется выбрать короткую крутую тропу, т. е. выбрать квантовое видообразование.
Прерывистые равновесия
Гипотеза прерывистого равновесия была изложена в четырёх ключевых публикациях (Eldredge, Gould, 1972; Gould, Eldredge, 1977; Stanley, 1975, 1979*). Эта гипотеза содержит следующие основные положения: 1) со времен Дарвина большинство эволюционистов, в том числе сторонники синтетической теории, рассматривают эволюцию как состояние медленного и равномерного изменения, как случай «филетического градуализма»; 2) подлинные макроэволюционные изменения носят характер вспышек, так что длительные периоды постоянства (или «застоя») чередуются с короткими эпизодами быстрого изменения; 3) эпизоды существенного изменения совпадают с видообразовательными событиями. Этот тезис сформулирован впервые.
Концепция прерывистого равновесия вызвала большие разногласия сразу же после первых публикаций, появившихся в семидесятых годах. При критическом рассмотрении этой проблемы возможны два подхода. Можно просто принять модель прерывистого равновесия и обсуждать доводы за и против, как это делают многие авторы, или же можно исследовать её в истори ческом и научном плане, чтобы выяснять, насколько она оригинальна. Мы выберем здесь этот второй подход (более детальную критику см. Grant, 1982, 1983; Simpson, 1983; Gingerich, 1984; Brown, 1987*).
Несмотря на то что взгляды Элдреджа (Eldredge) и Гулда (Gould), с одной стороны, и Станли (Stanley) — с другой, по основным теоретическим моментам сходятся, их работы заметно различаются по научному стилю и будут рассмотрены по отдельности. Главная работа Стэнли (1979*) по прерывистому равновесию хорошо документирована в отношении как фактических данных, так и эволюционной литературы и заслуживает серьезного внимания.
Элдредж и Гулд (Eldredge, Gould, 1972; Gould, Eldredge, 1977*), напротив, удивительно последовательно игнорировали основополагающую литературу по рассматриваемому (вопросу. Их работа 1972 г. была написана под влиянием Майра (Mayr) и Дарвина. Литература, на которой основана их работа 1977 г., несколько полнее, но всё ещё удручающе недостаточна.
Первый результат этого неадекватного литературного обзора — соломенное чучело, филетический градуализм. Ни один из крупных эволюционистов последних лет не пропагандировал филетический градуализм в чистом виде. Напротив, в синтетической теории эволюции непрерывно обсуждается идея о происходящих время от времени эпизодах быстрого и резкого эволюционного изменения. Эту идею рассматривали ещё до 1972 г. Райт (Wright, 1931, 1949, 1960*), Симпсон (Simpson, 1944, 1953*), Ренш (Rensch, 1947, 1960*), Майр (Mayr, 1954, 1963, 1970*), Карсон (Carson, 1959, 1971*), Льюис (Lewis, 1962*), Грант (Grant, 1963, 1971*) и другие авторы. Ни одной из этих ранних работ, за исключением вклада Майра, Элдредж и Гулд не уделили должного внимания в своих статьях 1972 и 1977 гг., а многими просто пренебрегли.
Концепция Симпсона о квантовой эволюции в статье 1972 г. не упоминается вовсе, а в статье 1977 г. о ней говорится между прочим. Позднее Гулд (Gould, 1982a*) оправдывал это упущение на том основании, что квантовая эволюция — это один из способов филетической эволюции, тогда как прерывистое равновесие —результат видообразования. В поддержку своего высказывания Гулд приводит цитату из Симпсона (Simpson, 1953*). Он не упоминает, однако, об имеющемся на той же странице утверждении Симпсона, что квантовая эволюция может происходить также и путем видообразования.
Хотя Симпсон и допускал наличие связи между видообразованием и квантовой эволюцией, он не исследовал эту связь. Однако спустя несколько лет это было сделано мною (Grant, 1963*) в работе, которую Элдредж и Гулд также игнорировали.
Здесь я сосредоточил внимание на многочисленных и серьезных литературных упущениях в статьях Элдреджа и Гулда, опубликованных в 1972 и 1977 гг. Следует отметить, что указанные упущения не были должным образом восполнены в более поздних статьях этих авторов (Gould, Eldredge, 1986*).
Так что же нового содержит гипотеза прерывистого равновесия? Ничего. В этой гипотезе нет новых идей, которые не высказывались бы в ясной и чёткой форме в эволюционной литературе, выходившей до 1972 г.
В самом деле, модель прерывистого равновесия не так хороша, как более раннее толкование этой проблемы. Модель эта носит схоластический характер. Она пытается свести все или почти все палеонтологические ряды к двум противоположным состояниям: застою и прерывистым сдвигам. Но фактически в эволюционной истории реальных групп наблюдается широкий диапазон эволюционных скоростей или смеси анагенетических и кладогенетических способов. Модель квантовой эволюции — открытая структура, способная вместить всё разнообразие действительно существующих макроэволюционных процессов.
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | ↑ |